Biological Functions of HMGN Chromosomal Proteins


Laboratory of Metabolism, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Bethesda, MD 20892, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(2), 449; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020449
Submission received: 18 December 2019
/
Revised: 2 January 2020
/
Accepted: 8 January 2020
/
Published: 10 January 2020
(This article belongs to the Special Issue HMG Proteins in Development and Disease)
Abstract
:Chromatin plays a key role in regulating gene expression programs necessary for the orderly progress of development and for preventing changes in cell identity that can lead to disease. The high mobility group N (HMGN) is a family of nucleosome binding proteins that preferentially binds to chromatin regulatory sites including enhancers and promoters. HMGN proteins are ubiquitously expressed in all vertebrate cells potentially affecting chromatin function and epigenetic regulation in multiple cell types. Here, we review studies aimed at elucidating the biological function of HMGN proteins, focusing on their possible role in vertebrate development and the etiology of disease. The data indicate that changes in HMGN levels lead to cell type-specific phenotypes, suggesting that HMGN optimize epigenetic processes necessary for maintaining cell identity and for proper execution of specific cellular functions. This manuscript contains tables that can be used as a comprehensive resource for all the English written manuscripts describing research aimed at elucidating the biological function of the HMGN protein family.
1. Introduction
High mobility group N (HMGN) is a family of non-histone chromatin architectural proteins, ubiquitously expressed in the nuclei of vertebrate cells [1,2] that binds specifically to nucleosomes, the building block of the chromatin fiber. HMGNs bind to nucleosomes dynamically without any specificity for DNA sequence [3,4,5]. The interaction of HMGNs with nucleosomes weakens the binding of linker histone H1 to nucleosomes, thereby affecting the local and global chromatin structure [6,7,8,9]. Genome-wide HMGNs preferentially localize to chromatin regulatory sites such as enhancers and promoters, thereby affecting the integrity of the chromatin epigenetic landscape and the cellular transcription profile [10,11].
The HMGN family consists of five proteins named HMGN1, 2, 3, 4 and 5, all containing a bipartite nuclear localization signal, a negatively charged chromatin regulatory domain located at C terminal region of the protein, and a positively charged nucleosome binding domain, which is highly conserved and is the consensus sequence of this protein family [5]. The major members of this family, HMGN1 and HMGN2, have been detected in every vertebrate tissue examined. The expression of HMGN3, 4 and 5 proteins seems to be more restricted and with a few notable exceptions [12,13], the level of these HMGN variants is significantly lower than that of HMGN1 and HMGN2. The genes coding for HMGN1, 2, 3 and 5 have a very similar intron-exon organization, raising the possibility that they originated from a common ancestor. The gene coding for HMGN4 is the product of a retroviral transposon and appeared to be restricted to primates [2,14]. Both the human and mouse genome contains multiple HMGN1 and HMGN2 retropseudogenes dispersed throughout their genome [15,16]; however, to date, only HMGN4 has been identified as a retropseudogene coding for a protein. In humans, HMGN1 is located on chromosome 21 (21q22.3), HMGN2 is located on Chromosome 1 (1p36.1), HMGN3 and HMGN4 are located on chromosome 6 (6q14.1 and 6p21.3) and HMGN5 is located in Chromosome X (Xp13.3) [17]. HMGN1-4 have similar molecular weights, around 10 kDa (∼90 amino acids length), while HMGN5 contains a long C-terminus; the length of this region varies among species [14,17].
Given the ubiquitous expression of HMGNs in vertebrate cells, their specific binding to nucleosomes, and their preferential association with chromatin regulatory sites it could be expected that these proteins would affect cell type-specific gene expression programs. Indeed, it has been reported that HMGNs play a role in embryogenesis and affect neuronal, ocular, reproductive, and pancreatic cell differentiation. In addition, several studies have suggested that HMGNs play a role in DNA repair processes and that the loss of HMGNs can lead to cancer, neurological disorders, and altered immune functions.
Likely, HMGNs affect the cellular phenotype by modulating epigenetic processes that affect cell type-specific gene expression [11,18], a subject of considerable interest that is not discussed here. In this review, we focus on the biological function of HMGN and summarize the available data on the role of HMGN proteins in developmental processes and in disease etiology. The two tables included in the manuscript provide a comprehensive resource for references to all the English written manuscripts on these topics that have been published in PubMed (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed) up to December 2019.
2. Genetically Altered HMGN Mice
A major approach used to study the biological function of HMGN involves correlative studies in which changes in cellular phenotypes are linked to changes in the expression level of specific HMGN variants. A second important approach involves analysis of cells and tissues in which the levels of HMGNs are experimentally altered by using specific vectors to either increase, decrease, or fully delete the expression of a specific HMGN variant. In this approach, the use of genetically altered mice can provide information on the function of HMGN variants at the biological level of an entire organism.
Table 1 lists the genetically altered HMGN mice that have been used to study HMGN function. The phenotype of most of these lines has also been analyzed in detail by the German Mouse Clinic and is posted online at https://www.mouseclinic.de. With one exception [19] the results indicate that mice lacking HMGN variants are born, appear normal and survive; however, each of the genetically altered mouse line shows phenotypic differences from its wild-type littermate mouse line, especially when exposed to stress.
3. HMGN Proteins in Development
3.1. Embryogenesis
HMGN expression is developmentally regulated; significant HMGN1 and 2 expression is seen during mouse oogenesis and early preimplantation [20,21]; however, during subsequent development and differentiation, the expression levels of these variants decreases [22,23]. Quantitative RNA blot analysis of whole mouse embryos indicates that during differentiation from E7 to E17, the relative levels of HMGN transcripts decrease by 50%, and that the relative levels of HMGN transcripts in the adult mouse heart or spleen are only 5% of those detected in 7-day embryos [22]. Nevertheless, every tissue examined in the adult mouse contains Hmgn1,2 transcripts and HMGN proteins. The high relative levels of HMGN expression during early embryogenesis suggests that these proteins may play a role in lineage commitment and cellular differentiation.
Indeed, several findings suggested that proper expression level of HMGN1 and HMGN2 may play a role during early stage embryogenesis. Thus, prominent expression of HMGN1 and HMGN2 is seen in mouse embryonic stem cells (ESCs) [20], in induced pluripotent stem cells (iPSCs) [11], and in adult mouse hair follicle bulge where hair stem cells are located [24]. Hmgn1 and Hmgn2 transcripts are detected during mouse oogenesis, and after fertilization, maternal Hmgn transcripts accumulate in the two-cell stage embryo. Transient depletion of HMGN1 and HMGN2 proteins by injecting antisense oligonucleotides into mouse oocytes, delayed cell cleavage and the onset of the blastocyst stage [21]. Both HMGN1 and HMNG2 are highly expressed throughout mouse preimplantation stages, including blastocyst stage, and both HMGN1 and HMGN2 are strongly expressed in ESCs [20,25]. In tissue culture, loss of HMGNs does not affect the rate of differentiation of ESCs into embryoid bodies; however, the gene expression profile of the Hmgn−/− cells was clearly different from that of cells obtained from genetically matched WT littermates [20]. A recent study shows that CRISPR-mediated knock out of HMGN2 in P19, a pluripotent stem cell model, alters the epigenetic landscape of these cells, leading to changes in gene expression. The authors conclude that HMGN2, and to a lesser degree, HMGN1, stabilize the epigenetic landscape necessary for maintaining the pluripotent identity of these cells [26].
Additional support that HMGNs may play a role in early development is seen in the initial reports on the Hmgn1−/− mouse line which indicate that the frequency ratio of Hmgn1−/− pups resulting from mating of Hmgn1+/− heterozygotes was 0.08 rather than 0.25, as would be expected from a Mendelian distribution of the Hmgn1−/− allele. The low number of Hmgn1−/− offspring seems to be due to events occurring in early stages of embryonic development, since the genotype distribution in 11.5-day embryos was the same as that seen in born pups. In addition, the average litter size obtained from mating of Hmgn1−/− was approximately 30% lower than that resulting from Hmgn1+/+ mating [27].
In mice, downregulation of both Hmgn1 and Hmgn2 transcripts is seen during chondrocyte, kidney, hair follicle, and corneal development and differentiation [24,28,29]. In addition, cell culture studies show downregulation of HMGN1 and HMGN2 expression during myogenesis, erythropoiesis and chondrogenesis [28,30,31]. In these tissue culture experiments, aberrant expression of HMGN proteins inhibits myoblast and chondrocyte differentiation. It may be relevant that studies by the German Mouse Clinic (https://www.mouseclinic.de) indicate that the average size of the double knock-out mice lacking both HMGN1 and HMGN2 is smaller than that of wild-type (WT) littermate mice, a finding also observed by our laboratory.
During Xenopus development, Hmgn1 and Hmgn2 transcripts are detected in the three Xenopus germinal layers; however, in the embryos, HMGN1 and HMGN2 proteins are detected and accumulate only after mid blastula transition (MBT). Morpholino mediated HMGN1 and HMGN2 depletion, or vector mediated HMGN overexpression prior to MBT does not affect Xenopus development; however, after MBT, aberrant HMGN expressions cause imperfectly closed blastopores, distorted body axis, and embryos with abnormal head structures [29]. These findings suggest that in Xenopus, correct HMGN expression is crucial for proper development. The differences between Xenopus and mice in the effect of HMGNs on differentiation raises the possibility that the requirement for HMGN functions are more stringent in “lower” vertebrates; perhaps “higher” vertebrates developed redundant mechanisms to bypass the absolute requirement for proper expression of HMGN variants.
Chromatin is not only a major regulator of gene expression but seems to play a structural role in supporting the mechanical properties of the nucleus [32]. In this aspect, overexpression of HMGN5 variant has been shown to decrease nuclear sturdiness, most likely by reducing the binding of histone H1 to nucleosomes [7], thereby leading to chromatin decompaction. In agreement with this possibility, transgenic mice overexpressing HMGN5 develop hypertrophic heart with large cardiomyocytes, deformed nuclei and disrupted lamina and die of cardiac malfunction [33]. Mice lacking HMGN5 show several phenotypes (https://www.mouseclinic.de) including changes in transcription of genes that alter glutathione metabolism [34].
3.2. Neuronal Development
The database in the Allen Institute of Brain Sciences (https://alleninstitute.org/) show high HMGN expression in both human and mouse brain, especially in the eye, hippocampus, and SVZ regions. In mouse ESCs, loss of HMGN1 and HMGN2 reduces the expression of OLIG1 and OLIG2, two transcription factors known to play an important role in oligodendrocyte differentiation. Indeed, these Hmgn1−/−Hmgn2−/− (DKO) ESCs showed a reduced ability to differentiate towards the oligodendrocyte lineage and mice lacking these HMGNs showed reduced oligodendrocyte count, decreased spinal cord myelination, and display several neurological phenotypes that could be related to the decreased oligodendrocyte count and faulty nerve myelination [20]. A recent study using a new line of Hmgn2−/− mice reports that mice lacking HMGN2 show reduced cortical surface and microcephaly, suggesting a role for HMGN2 in corticogenesis. These Hmgn2−/− mice were sub viable and most die within a short time after birth [19] an unexpected observation since previous studies showed that mice lacking either HMGN1 or HMGN2, or even both these variants survive [10]. The differences may be due to the targeting strategy and the exact timing of HMGN deletion during development.
Cell culture studies [35] revealed that forced expression of HMGNs increased the generation of astrocytes, while knockdown of HMGNs in neural progenitor cells suppressed astrocyte differentiation. Most likely, HMGNs affect astrocyte differentiation by modulating the JAK-STAT pathway. HMGN3 is expressed in several regions of adult mouse brain that are enriched in astrocytes [36]. In addition, HMGN3 regulates GLYT1 expression, a protein known to plays an essential role at glycinergic and glutamatergic synapses in the brain and central nervous system [37,38]. In the mouse neuroblastoma cell line N1E-115, the mRNA encoding HMGN5 localizes to growth cones of both neuron-like cells and of hippocampal neurons, where it has the potential to be translated, and that HMGN5 can be retrogradely transported into the nucleus along neurites. Loss of HMGN5 function induces transcriptional changes and impairs neurite outgrowth, while HMGN5 overexpression induces neurite outgrowth and chromatin decompaction suggesting that this HMGN variant plays a role in facilitating growth cone-to-nucleus signaling during neuronal development [39].
The human HMGN1 gene is located in chromosome 21 [40] in a segment known as the Down Syndrome critical region (DSCR), which is thought to play an important role in the etiology of this syndrome. Elevated HMGN1 expression is observed in human Down Syndrome cells [41] and in cells from Ts1Cje mouse, which is used as a model for Down Syndrome [42]. HMGN1 was shown to affect the expression of the methyl CpG-binding protein 2 (MeCP2) a DNA-binding protein known to affect neurological functions including autism spectrum disorders. Quantitative PCR and Western analyses of cell lines and brain tissues from mice that either overexpress or lack HMGN1 indicate that HMGN1 is a negative regulator of MeCP2 expression [43]. Alterations in HMGN1 levels lead to changes in chromatin structure and histone modifications in the MeCP2 promoter. Behavior analyses by open field test, elevated plus maze, reciprocal social interaction, and automated sociability test, link changes in HMGN1 levels to abnormalities in activity and anxiety and to social deficits in mice.
Taken together, the available findings support the possibility that aberrant expression of HMGN variants can lead to neurodevelopmental disorders.
4. Ocular Development
A systematic investigation of the expression patterns of HMGN1, HMGN2 and HMGN3 in the mouse eye development revealed that each HMGN variant is expressed in a specific manner [44]. At embryonic day 10.5 (E10.5) HMGN1 and HMGN2 proteins are evenly distributed in all ocular structures while HMGN3 expression is not detected at this stage. At E13.5, a decrease in HMGN1, but not in HMGN2 protein level is detected in the newly formed lens fiber cells. At this stage, HMGN3 is detected in the developing corneal epithelium and lens fiber cells. In adult mice, HMGN1 and HMGN2 are detected throughout the retina and lens, but in the cornea, these HMGNs are predominately located in the epithelium. In the cornea, HMGN3 is transiently upregulated in the stroma and endothelium at birth, while in adults, HMGN3 expression is restricted to the corneal epithelium. In the retina, HMGN3 expression is upregulated around 2 weeks of age and remains at relatively high levels in the inner nuclear and ganglion of the adult retina. Thus, even though all members of the HMGN protein family have similar protein structures, in the eye, each protein has a distinct regional and developmental stage expression pattern. In Hmgn1−/− mice the corneal epithelium is thin, has a reduced number of cells, is poorly stratified and shows additional abnormalities, all suggesting that loss of HMGN1 leads to transcriptional changes that ultimately affect corneal development and maturation [28].
5. Reproductive System
HMGN5 is highly expressed in placental rather than embryonic tissues, and is highly enriched in trophoblast giant cells, which play a key role in placental differentiation. In Rcho-1 cells, a cell line that serves as a model system for studies on the development of trophoblast giant cells, the expression levels of HMGN5 is highly correlated with prolactin genes and placental lactogens [12]. Hmgn3 is highly expressed in the decidua and decidualizing stromal cells or artificially stimulated decidualization leads to Hmgn3 expression. In vitro, overexpression of HMGN3 in uterine stromal cells enhanced the expression of decidualization markers, whereas inhibition of HMGN3 reduced their expression [45]. In sum, the data show a correlation between expression of specific HMGN variants in the reproductive system cells; however, it is still not clear whether proper HMGN expression in these cells is necessary for their adequate function.
6. Pancreas
In both the human and mouse pancreas, HMGN3 is specifically expressed in adult pancreatic islet cells [13]. All the endocrine cell types including the insulin-producing beta-cells and glucagon-producing alpha cells express high levels of HMGN3 [46]. Hmgn3−/− mice are viable and appear normal; however, compared to their wild-type littermates, their blood glucose levels are elevated, their serum insulin levels are reduced, and their glucose tolerance is impaired. In vitro studies using the mouse insulinoma MIN6 cell line revealed that HMGN3 modulates the expression of several genes involved in insulin secretion, including that for the glucose transporter GLUT2. In both MIN6 cells and Hmgn3−/− mice, the loss of HMGN3 protein reduces the levels of GLUT2. HMGN3, but not the HMGN1 or HMGN2 variants, binds specifically to chromatin in the promoter region of the Glut2 gene and enhances its expression by facilitating the recruitment of PDX1 and additional transcription factors, to the Glut2 promoter. Chromatin immunoprecipitation (ChIP) experiments reveal that HMGN3 enhances the acetylation of H3 at the Glut2 promoter and suggest that HMGN3 and PDX1 mutually reinforce their interaction with the chromatin at the Glut2 promoter but not throughout the chromatin or in the nucleoplasm.
In Hmgn3−/− mice, loss of HMGN3 reduces the levels of glucagon in the plasma. However, analysis of TC1-9 cells, a cell line that is widely used for studies on pancreatic alpha-cell function, did not reveal a major role for HMGN3 in glucagon synthesis and secretion. These results indicate that HMGN3 reduces the levels of glucagon in the plasma of mice without affecting glucagon gene expression [46].
Table 2 lists the references accessible in PubMed (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed) discussing the role of HMGN variants in various developmental processes. The references are limited to articles written in English. Taken together, the available data suggest that mice lacking HMGN genes are born and survive; however, loss of HMGN occasionally leads to reduced litter size, to smaller sized mice, and to epigenetic changes that play a role in the etiology of several developmental disorders.
7. HMGN Proteins in Disease
The widespread occurrence of HMGN variants in all vertebrate cells, the high degree of sequence conservation of the proteins among all vertebrates, and the localization of the proteins to chromatin regulatory sites, suggest that they are required for proper cellular function. Table 3 lists the references describing investigations of the biological function of HMGNs, especially their possible role in the etiology of disease. In addition, as described above, several mice lines that have been genetically modified to alter HMGN expression have been extensively phenotyped by the German Mouse Clinic (https://www.mouseclinic.de/); the results of these analyses are available online. Several general conclusions can be drawn from these studies: (1) Cells and mice with altered HMGN expression survive. (2) Invariably, altered levels of HMGN lead to changes in gene expression. (3) Loss or altered expression of HMGNs leads to both up and downregulation of tissue-specific gene expression. (4) HMGNs modulate the existing cell type-specific gene expression program. (5) Mice lacking HMGN show numerous phenotypes; however, the phenotypes are not strong. Part of the phenotypes seen suggest a possible role for HMGNs in cancer, neurological disorders and in immunological processes.
8. Cancer
Cancer initiation and progression often show changes in gene expression that reflect alteration in cell identity. Significantly, recent results show that HMGN1 and HMGN2 preferentially bind to cell type-specific chromatin regulatory regions and that loss of HMGN enhances the rate of transcription factor-induced reprogramming of mouse embryonic fibroblasts (MEF) into pluripotent cells, and the lineage conversion of MEFs into neuronal cells. These and additional experiments suggest that HMGN stabilizes cell identity, most likely by facilitating the binding of cell type-specific regulatory factors to cell type-specific regulatory sites [11]. A PubMed search on the role of HMGN proteins in cancer retrieved a wide range of studies involving analysis of human cell lines, mice models, and patient clinical data (Table 3). Comparative gene expression analyses of normal and cancer cells frequently show differences in HMGN expression. However, it is not yet clear whether the changes in HMGN expression levels are a cause or a consequence of the transcriptional changes seen in cancer cells.
High cytoplasmic expression and secretion of HMGN1 were associated with increased peritumor infiltration of lymphocytes in breast cancer [47,48]. In non-small cell lung cancer (NSCLC) patients, the levels of HMGN1 in the serum were correlated with overall survival after curative pneumonectomy. Likewise, the levels of HMGN1 in the serum of NSCLC individuals were higher in non-metastatic stages (I-III) as compared to the metastatic stage (IV), suggesting that HMGN1 can serve as a biomarker for early stages of NSCLC [49]. Leukemic B cells from patients with chronic lymphoblastic leukemia express HMGN2 on their surface; the HMGN2 variant acts as an autoantigen and contributes to the development of autoimmune hemolytic anemia [50]. Individuals with Down syndrome have a significantly increased risk of developing B cell acute lymphoblastic leukemia (B-ALL). HMGN1, coded by a gene located on human chromosome 21, seems to play a crucial role in the B-ALL. Studies with Ts1Rhr mice which contain a triplication of a segment of mouse chromosome 16 that is orthologous to human chromosome 21 and serve as models for Down syndrome, and analysis of cells derived from a transgenic mice overexpressing human HMGN1 [51] provide evidence that HMGN1 overexpression promotes B-ALL. In B-ALL, elevated levels of HMGN1 reduce the levels of H3K27me3, a histone modification associated with gene silencing, and increase the levels of H3K27ac, a modification associated with increased gene expression, thereby leading to transcriptional amplification and promoting B cell proliferation [52,53].
HMGN2 was also shown to promote breast cancer growth in response to prolactin. In this system, HMGN2 reduces the binding of linker histone H1 to chromatin regulatory regions, thereby enhancing STAT5 accessibility to promoter DNA, ultimately leading to increased proliferation of breast cancer cells [54,55]. Further analysis showed that the K2 residue of HMGN2 was deacetylated in primary breast tumors but highly acetylated in normal human breast tissue, implying that targeting HMGN2 deacetylation may be a viable treatment for this type of breast cancer [55].
The HMGN4 variant which is transcribed from a processed retropseudogene [56], is expressed only in humans. Upregulation of HMGN4 expression in mouse and human cells, and in the thyroid of transgenic mice, alters the cellular transcription profile, downregulates the expression of the tumor suppressors Atm, Atrx, and Brca2, and elevates the levels of the DNA damage marker γH2AX. Mouse and human cells overexpressing HMGN4 show increased tumorigenicity as measured by colony formation, by tumor generation in nude mice, and by the formation of preneoplastic lesions in the thyroid of transgenic mice. In patients with hepatocellular carcinoma (HCC), elevated level of HMGN4 expression is associated with high grade tumors and shorter overall survival, suggesting that in patients with HCC, HMGN4 could act as a candidate biomarker for poor prognosis. Thus, HMGN4 may serve as an additional diagnostic marker or perhaps even a therapeutic target in certain thyroid cancers [56]. HMGN5 expression seems to be elevated in breast, prostate and bladder cancer cell lines (Table 3); however, the oncogenic potential of this variant has not yet been studied in detail.
In mice, HMGN1 was shown to bind to the promoters of the proto-oncogene FosB and Hmgn1−/− mice show upregulated expression of the proto-oncogenes FosB, C-fos, BclL3 and N-cadherin and downregulation of JunB, and c-Jun, suggesting a possible role of this HMGN variant in modulating the action of these factor during cancer development [57,58]. Following a single DEN injection, mice lacking HMGN1 displayed earlier signs of liver tumorigenesis as compared to wild-type mice [59] suggesting that loss of HMGN1 leads to epigenetic changes that affect the rate of N-nitrosodiethylamine-induced hepatocarcinogenesis in mice. In support of this possibility, transformed Hmgn1−/− fibroblasts grow in soft agar and produce tumors in nude mice with significantly higher efficiency than WT littermate fibroblasts. Hmgn1−/− mice also show an increased incidence of tumors and metastases as compared to their Hmgn1+/+ littermates [60]. Together, these results support the general notion that the presence of HMGN stabilizes the cell epigenetic landscape that regulates cell type-specific transcription and determines cell identity [11].
It is well established that faulty repair of damaged DNA is a major factor contributing to cancer development. Several types of experiments suggest a role for HMGN proteins in DNA repair processes [61,62]. Thus, exposure of the shaved backs of Hmgn1−/− mice and their Hmgn1+/+ littermates to UV-B produced significant damage in the skin of Hmgn1−/− mice but not in the skin of control, wild-type littermates. Furthermore, mouse embryonic fibroblasts isolated from Hmgn1−/− mice were significantly more sensitive to UV radiations than MEFs isolated from WT littermates, most likely because the loss of HMGN1 impairs the removal of Cyclobutene pyrimidine dimers from the chromatin of UV irradiated cells. Taken together, these results indicate that loss of HMGN decreased the cellular ability to repair DNA damaged by UV radiation [27]. Hmgn1−/− mice and MEFs were also more sensitive to ionizing radiation than their wild-type littermates. Upon exposure to ionizing radiation, the tumor burden and mortality in Hmgn1−/− mice are higher as compared to their wild-type littermate mice [60]. The fibroblasts hypersensitivity to ionizing radiation is linked to altered G2-M checkpoint activation. Thus, in mice, loss of HMGN impairs repair of DNA damaged by either UV or ionizing radiation. Likewise, although DT40 chicken cells lacking either HMGN1, HMGN2 or both these variants survive [63], they are hypersensitive to UV irradiation. In these cells, loss of HMGNs increased the extent of G2-M checkpoint arrest and apoptosis [64]. An additional possible link between DNA damage and HMGN proteins is shown by a recent finding that the damage repressor protein Dsup of tardigrade Ramazzottius varieornatus, which binds to DNA and protects from hydroxy free radicals, exhibits sequence similarity with HMGN proteins [65].
In sum, the data support the possibility that the interaction of HMGN with chromatin optimizes DNA repair processes, while loss of HMGN variants decreases the efficiency of DNA repair, thereby increasing cell tumorigenicity. The mechanisms whereby HMGNs affect DNA repair processes are not fully understood; most likely, HMGNs affect DNA repair by modulating the interaction of DNA repair factors with chromatin, as was shown for ATM and for poly ADP-ribose polymerase-1 (PARP-1) [66,67,68].
9. Neurological Disorders
Several studies link altered HMGN1 to neurological disorders. Down syndrome, a common human genetic disorder affecting neural development, is caused by the presence of all, or part of an extra copy of chromosome 21 [69]. The HMGN1 gene is located in a region of human chromosome 21 that is most frequently triplicated in Down syndrome, and tissue samples obtained from Down syndrome individuals overexpress HMGN1 protein [40,41]. A region in distal mouse chromosome 16 shows conserved linkage with human chromosome 21 and mouse trisomic in this region have been used as models for Down syndrome [70]. The mouse models for Down syndrome express approximately 1.5-fold higher levels of HMGN1 mRNA and protein, as compared to normal littermates [40,71,72]; however, it is not clear whether the neurological phenotypes seen in Down syndrome individuals are directly linked to elevated HMGN1 levels.
Alterations in the DNA binding protein methyl CpG-binding protein 2 (MeCP2) levels are known to affect neurodevelopmental disorders such as autism spectrum disorders, mental retardation, learning disabilities, Rett syndrome, repetitive behavior, hypotonia, and anxiety [73,74,75]. Targeted analysis of the Autism Genetic Resource Exchange genotype collection revealed a non-random distribution of genotypes within 500 kbp of HMGN1 in a region affecting its expression in families predisposed to autism spectrum disorders [43]. In mice, HMGN1 is a negative regulator of MeCP2 and analyses of genetically altered HMGN1 mice show that changes in HMGN1 levels are linked to abnormalities in activity and anxiety and social skills [43]. As indicated in the previous section, HMGN proteins regulate the expression of Olig1 and Olig2 and mice lacking HMGN1 and HMGN2 show decreased myelination and phenotypes that can be attributed to altered nerve impulse transmission [20]. Taken together, the available results suggest a role for HMGN variants in proper differentiation and function of neural cells and support the possibility that misexpression of these proteins could play a role in the etiology of neuronal disorders.
10. Immune Functions
Several studies reported the presence of autoantibodies against HMGN1 and HMGN2 in the serum of individuals with autoimmune diseases such as systemic lupus erythematosus, rheumatoid arthritis, mixed connective tissue disease, and scleroderma [76,77,78,79]. However, there is no clear evidence that HMGN variants play a role in the etiology of autoimmune disease. The antibodies seem to have been elicited by HMGN variants or chromatin fragments containing HMGN that are immunogenic due to cellular damage and nuclear breakdown in these diseases, to defects of the immunosurveillance system against autoantigens, or to “molecular mimicry” to an invading pathogen. Nevertheless, analyses of antibodies to HMGN in autoimmune disorders could provide clues to the etiology of these disorders [77].
Support for “molecular mimicry” between bacterial pathogens and HMGN proteins is provided by several studies reporting that HMGNs and HMGN-derived peptides show antimicrobial activity. The presence of HMGN2 in the cytoplasm and extracellular environment of mononuclear leukocytes highlights the function of HMGN2 as an antimicrobial effector. Full-length HMGN2 also displayed potential antimicrobial activity against E. coli ML-35p, Pseudomonas aeruginosa ATCC 27853, and Candida albicans ATCC 10231 and in vivo, inhibited the invasion of K. pneumoniae 03182 onto mouse lungs [80]. An antimicrobial peptide with the sequence PKRKAEGDAK, which is identical to the nucleosome binding domain of HMGN2 was isolated from human peripheral blood mononuclear leukocytes [81]. In several instances, the isolated alpha-helical domain peptide of HMGN2 has been shown to possess the same antimicrobial activity as full length HMGN2 [81]. In addition, HMGN2 seems to induce autophagy, a novel antimicrobial pathway, and protects bladder epithelial cells from infection by E. coli and other Gram-negative bacteria [82,83,84]. In agreement, recent studies indicated that deficiency in HMGN2 affected macrophage polarization and non-tuberculosis mycobacterial survival in macrophages [85]. The molecular mechanisms involved in the antibacterial activity of HMGN2 has not been studied in detail. Most likely, “molecular mimicry” between HMGN and bacterial components stimulate the immune system to mount cellular responses that counteract the deleterious effects of microbial infection.
Alarmins are endogenous molecules that stimulate defense responses including innate and adaptive immune systems. In this context, HMGN1 was shown to act as an alarmin by inducing maturation in human dendric cells via Toll-like receptor 4 (TLR-4) and by recruiting antigen-presenting cells (APCs) to the site of infection [86,87]. Furthermore, when administrated along with antigen, HMGN1 acts as an adjuvant and promoted specific immune response in WT but not in Hmgn1−/− mice [87]. Absence of tumor specific immune response in Hmgn1−/− mice and increased anti-tumor immune response, when vaccinated mice with DNA vector overexpressing HMGN1, provide further support the role of HMGN proteins in regulating immune cell function and response [88]. Intratumor delivery of HMGN1 and TLT7/8 synthetic agonist R848, along with a checkpoint inhibitor Cytoxan synergistically activates dendritic cells for optimal Th1 response and eradicated large established CT26 tumors in mice [89,90]. Moreover, low doses of HMGN1 with anti-CD4 depleting antibody promoted the expansion of anti-tumor CD8 T cells by rescuing them from exhaustion [91]. In a recent study on patients with head and neck carcinoma, it was observed that cytoplasmic localization and secretion of HMGN1 was associated with the recruitment of tumor infiltering lymphocytes [92]. A proteomics study in HMGN1 knockout revealed the role of HMGN1 in complement and coagulation cascade and interferon response in cancer cells, reinforcing the role of HMGN1 in regulating immune response [93]. These and additional studies suggest that HMGN1 can be used to increase the efficiency of certain antitumor treatments [89,94,95].
11. Perspective: Biological Function of the HMGN Protein Family
The biological function of HMGNs is a major unresolved question in chromatin biology.
Every vertebrate cell examined expresses HMGN proteins, the amino acid sequence of the proteins is conserved, the levels of these proteins correlate with specific developmental stages and several studies revealed that the protein binds specifically to chromatin, preferentially targeting regulatory sites such as enhancers and promoters [10,11]. Indeed, changes in HMGN levels can lead to altered cellular transcription profiles and a wide range of studies, including analyses of genetically altered mice, do correlate changes in HMGN levels with altered cellular phenotypes (Figure 1, Table 2 and Table 3).
Yet, in humans, not a single disease or developmental abnormality is directly attributed to or is strongly correlated with, either loss or decreased expression of an HMGN variant. The only direct correlation between changes in HMGN expression and a human phenotype is Down syndrome where an increase in HMGN1 has been shown to correlate with increased incidence of B-ALL [52]. Likewise, mice lacking HMGN are born and appear normal; however, detailed examinations reveal multiple phenotypes, especially when exposed to stress. A hypothetical explanation for these observations is that in humans, loss of HMGN expression leads to embryonic lethality; embryos with disrupted HMGN expression are not born. Mice lacking HMGN survive because under the controlled conditions of an animal facility loss, HMGN does not lead to embryonic lethality; presumably, under more stressful conditions found outside an animal facility, the mice would not be born or survive. In Xenopus, changes in HMGN levels lead to embryonic lethality, provided that the changes in HMGN levels are induced after midblastula transition [29].
A possible clue to molecular mechanisms regulating the biological function of the HMGN protein family comes from experiments showing that HMGNs bind to cell type-specific enhancers [11,96], that they do regulate the cell type-specific gene expression [18] and that they do stabilize, rather than determine cell identity, most likely by modulating the dynamic establishment and maintenance of a cell type-specific chromatin epigenetic landscape [11]. These findings suggest that cells do indeed survive without these proteins because they optimize and fine-tune rather than determine cell type-specific biological processes. The widespread occurrence and sequence conservation of HMGN proteins in vertebrate cells, and their association with cell type-specific chromatin regulatory sites, suggest that they do play important roles in safeguarding cell type-specific gene expression programs that optimize cellular functions necessary for proper development and prevention of disease.
Funding
This research was funded by the Center for Cancer Research, the Intramural Research Program of the National Cancer Institute, NIH. No additional external funding was received.
Acknowledgments
We thank Yuri V. Postnikov and Shaofei Zhang for critical comments on this review.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hock, R.; Furusawa, T.; Ueda, T.; Bustin, M. HMG chromosomal proteins in development and disease. Trends Cell Biol. 2007, 17, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Romero, R.; Eirin-Lopez, J.M.; Ausio, J. Evolution of high mobility group nucleosome-binding proteins and its implications for vertebrate chromatin specialization. Mol. Biol. Evol. 2015, 32, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catez, F.; Lim, J.H.; Hock, R.; Postnikov, Y.V.; Bustin, M. HMGN dynamics and chromatin function. Biochem. Cell Biol. 2003, 81, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, H.; Herrera, J.E.; Bustin, M.; Postnikov, Y. Targeting of high mobility group-14/-17 proteins in chromatin is independent of DNA sequence. J. Biol. Chem. 2000, 275, 37937–37944. [Google Scholar] [CrossRef] [Green Version]
- Postnikov, Y.; Bustin, M. Regulation of chromatin structure and function by HMGN proteins. Biochim. Biophys. Acta 2010, 1799, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Catez, F.; Brown, D.T.; Misteli, T.; Bustin, M. Competition between histone H1 and HMGN proteins for chromatin binding sites. EMBO Rep. 2002, 3, 760–766. [Google Scholar] [CrossRef] [Green Version]
- Rochman, M.; Postnikov, Y.; Correll, S.; Malicet, C.; Wincovitch, S.; Karpova, T.S.; McNally, J.G.; Wu, X.; Bubunenko, N.A.; Grigoryev, S.; et al. The interaction of NSBP1/HMGN5 with nucleosomes in euchromatin counteracts linker histone-mediated chromatin compaction and modulates transcription. Mol. Cell 2009, 35, 642–656. [Google Scholar] [CrossRef] [Green Version]
- Catez, F.; Ueda, T.; Bustin, M. Determinants of histone H1 mobility and chromatin binding in living cells. Nat. Struct. Mol. Biol. 2006, 13, 305–310. [Google Scholar] [CrossRef]
- Postnikov, Y.V.; Bustin, M. Functional interplay between histone H1 and HMG proteins in chromatin. Biochim. Biophys. Acta 2016, 1859, 462–467. [Google Scholar] [CrossRef] [Green Version]
- Deng, T.; Zhu, Z.I.; Zhang, S.; Postnikov, Y.; Huang, D.; Horsch, M.; Furusawa, T.; Beckers, J.; Rozman, J.; Klingenspor, M.; et al. Functional compensation among HMGN variants modulates the DNase I hypersensitive sites at enhancers. Genome Res. 2015, 25, 1295–1308. [Google Scholar] [CrossRef] [Green Version]
- He, B.; Deng, T.; Zhu, I.; Furusawa, T.; Zhang, S.; Tang, W.; Postnikov, Y.; Ambs, S.; Li, C.C.; Livak, F.; et al. Binding of HMGN proteins to cell specific enhancers stabilizes cell identity. Nat. Commun. 2018, 9, 5240. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, H.; Rochman, M.; Furusawa, T.; Kuehn, M.R.; Horigome, S.; Haketa, K.; Sugita, Y.; Inada, T.; Komai, M.; Bustin, M. The nucleosomal binding protein NSBP1 is highly expressed in the placenta and modulates the expression of differentiation markers in placental Rcho-1 cells. J. Cell. Biochem. 2009, 106, 651–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, T.; Furusawa, T.; Kurahashi, T.; Tessarollo, L.; Bustin, M. The nucleosome binding protein HMGN3 modulates the transcription profile of pancreatic beta cells and affects insulin secretion. Mol. Cell. Biol. 2009, 29, 5264–5276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugler, J.E.; Deng, T.; Bustin, M. The HMGN family of chromatin-binding proteins: Dynamic modulators of epigenetic processes. Biochim. Biophys. Acta 2012, 1819, 652–656. [Google Scholar] [CrossRef] [Green Version]
- Landsman, D.; McBride, O.W.; Soares, N.; Crippa, M.P.; Srikantha, T.; Bustin, M. Chromosomal protein HMG-14. Identification, characterization, and chromosome localization of a functional gene from the large human multigene family. J. Biol. Chem. 1989, 264, 3421–3427. [Google Scholar]
- Landsman, D.; McBride, O.W.; Bustin, M. Human non-histone chromosomal protein HMG-17: Identification, characterization, chromosome localization and RFLPs of a functional gene from the large multigene family. Nucleic Acids Res. 1989, 17, 2301–2314. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, T.; Cherukuri, S. Developmental function of HMGN proteins. Biochim. Biophys. Acta 2010, 1799, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Kugler, J.E.; Horsch, M.; Huang, D.; Furusawa, T.; Rochman, M.; Garrett, L.; Becker, L.; Bohla, A.; Holter, S.M.; Prehn, C.; et al. High mobility group N proteins modulate the fidelity of the cellular transcriptional profile in a tissue-and variant-specific manner. J. Biol. Chem. 2013, 288, 16690–16703. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.L.; Tian, W.J.; Liu, B.; Wu, J.; Xie, W.; Shen, Q. High mobility group nucleosomal binding domain 2 protects against microcephaly by maintaining global chromatin accessibility during corticogenesis. J. Biol. Chem. 2019. [Google Scholar] [CrossRef]
- Deng, T.; Postnikov, Y.; Zhang, S.; Garrett, L.; Becker, L.; Racz, I.; Holter, S.M.; Wurst, W.; Fuchs, H.; Gailus-Durner, V.; et al. Interplay between H1 and HMGN epigenetically regulates OLIG1&2 expression and oligodendrocyte differentiation. Nucleic Acids Res. 2017, 45, 3031–3045. [Google Scholar]
- Mohamed, O.A.; Bustin, M.; Clarke, H.J. High-mobility group proteins 14 and 17 maintain the timing of early embryonic development in the mouse. Dev. Biol. 2001, 229, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furusawa, T.; Lim, J.H.; Catez, F.; Birger, Y.; Mackem, S.; Bustin, M. Down-regulation of nucleosomal binding protein HMGN1 expression during embryogenesis modulates Sox9 expression in chondrocytes. Mol. Cell. Biol. 2006, 26, 592–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtonen, S.; Lehtonen, E. HMG-17 is an early marker of inductive interactions in the developing mouse kidney. Differentiation 2001, 67, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, T.; Ko, J.H.; Birger, Y.; Bustin, M. Expression of nucleosomal protein HMGN1 in the cycling mouse hair follicle. Gene Expr. Patterns 2009, 9, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Deng, T.; Zhu, Z.I.; Zhang, S.; Leng, F.; Cherukuri, S.; Hansen, L.; Marino-Ramirez, L.; Meshorer, E.; Landsman, D.; Bustin, M. HMGN1 modulates nucleosome occupancy and DNase I hypersensitivity at the CpG island promoters of embryonic stem cells. Mol. Cell. Biol. 2013, 33, 3377–3389. [Google Scholar] [CrossRef] [Green Version]
- Garza-Manero, S.; Sindi, A.A.A.; Mohan, G.; Rehbini, O.; Jeantet, V.H.M.; Bailo, M.; Latif, F.A.; West, M.P.; Gurden, R.; Finlayson, L.; et al. Maintenance of active chromatin states by HMGN2 is required for stem cell identity in a pluripotent stem cell model. Epigenetics Chromatin 2019, 12, 73. [Google Scholar] [CrossRef]
- Birger, Y.; West, K.L.; Postnikov, Y.V.; Lim, J.H.; Furusawa, T.; Wagner, J.P.; Laufer, C.S.; Kraemer, K.H.; Bustin, M. Chromosomal protein HMGN1 enhances the rate of DNA repair in chromatin. EMBO J. 2003, 22, 1665–1675. [Google Scholar] [CrossRef] [Green Version]
- Birger, Y.; Davis, J.; Furusawa, T.; Rand, E.; Piatigorsky, J.; Bustin, M. A role for chromosomal protein HMGN1 in corneal maturation. Differentiation 2006, 74, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Korner, U.; Bustin, M.; Scheer, U.; Hock, R. Developmental role of HMGN proteins in Xenopus laevis. Mech. Dev. 2003, 120, 1177–1192. [Google Scholar] [CrossRef]
- Pash, J.M.; Bhorjee, J.S.; Patterson, B.M.; Bustin, M. Persistence of chromosomal proteins HMG-14/-17 in myotubes following differentiation-dependent reduction of HMG mRNA. J. Biol. Chem. 1990, 265, 4197–4199. [Google Scholar]
- Crippa, M.P.; Nickol, J.M.; Bustin, M. Developmental changes in the expression of high mobility group chromosomal proteins. J. Biol. Chem. 1991, 266, 2712–2714. [Google Scholar]
- Bustin, M.; Misteli, T. Nongenetic functions of the genome. Science 2016, 352, aad6933. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, T.; Rochman, M.; Taher, L.; Dimitriadis, E.K.; Nagashima, K.; Anderson, S.; Bustin, M. Chromatin decompaction by the nucleosomal binding protein HMGN5 impairs nuclear sturdiness. Nat. Commun. 2015, 6, 6138. [Google Scholar] [CrossRef] [Green Version]
- Ciappio, E.D.; Krausz, K.W.; Rochman, M.; Furusawa, T.; Bonzo, J.A.; Tessarollo, L.; Gonzalez, F.J.; Bustin, M. Metabolomics reveals a role for the chromatin-binding protein HMGN5 in glutathione metabolism. PLoS ONE 2014, 9, e84583. [Google Scholar] [CrossRef]
- Nagao, M.; Lanjakornsiripan, D.; Itoh, Y.; Kishi, Y.; Ogata, T.; Gotoh, Y. High mobility group nucleosome-binding family proteins promote astrocyte differentiation of neural precursor cells. Stem Cells 2014, 32, 2983–2997. [Google Scholar] [CrossRef]
- Ito, Y.; Bustin, M. Immunohistochemical localization of the nucleosome-binding protein HMGN3 in mouse brain. J. Histochem. Cytochem. 2002, 50, 1273–1275. [Google Scholar] [CrossRef] [Green Version]
- West, K.L.; Castellini, M.A.; Duncan, M.K.; Bustin, M. Chromosomal proteins HMGN3a and HMGN3b regulate the expression of glycine transporter 1. Mol. Cell. Biol. 2004, 24, 3747–3756. [Google Scholar] [CrossRef] [Green Version]
- Barkess, G.; Postnikov, Y.; Campos, C.D.; Mishra, S.; Mohan, G.; Verma, S.; Bustin, M.; West, K.L. The chromatin-binding protein HMGN3 stimulates histone acetylation and transcription across the Glyt1 gene. Biochem. J. 2012, 442, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Moretti, F.; Rolando, C.; Winker, M.; Ivanek, R.; Rodriguez, J.; Von Kriegsheim, A.; Taylor, V.; Bustin, M.; Pertz, O. Growth Cone Localization of the mRNA Encoding the Chromatin Regulator HMGN5 Modulates Neurite Outgrowth. Mol. Cell. Biol. 2015, 35, 2035–2050. [Google Scholar] [CrossRef] [Green Version]
- Pash, J.; Popescu, N.; Matocha, M.; Rapoport, S.; Bustin, M. Chromosomal protein HMG-14 gene maps to the Down syndrome region of human chromosome 21 and is overexpressed in mouse trisomy 16. Proc. Natl. Acad. Sci. USA 1990, 87, 3836–3840. [Google Scholar] [CrossRef] [Green Version]
- Pash, J.; Smithgall, T.; Bustin, M. Chromosomal protein HMG-14 is overexpressed in Down syndrome. Exp. Cell Res. 1991, 193, 232–235. [Google Scholar] [CrossRef]
- Potier, M.C.; Rivals, I.; Mercier, G.; Ettwiller, L.; Moldrich, R.X.; Laffaire, J.; Personnaz, L.; Rossier, J.; Dauphinot, L. Transcriptional disruptions in Down syndrome: A case study in the Ts1Cje mouse cerebellum during post-natal development. J. Neurochem. 2006, 97 (Suppl. 1), 104–109. [Google Scholar] [CrossRef] [PubMed]
- Abuhatzira, L.; Shamir, A.; Schones, D.E.; Schaffer, A.A.; Bustin, M. The chromatin-binding protein HMGN1 regulates the expression of methyl CpG-binding protein 2 (MECP2) and affects the behavior of mice. J. Biol. Chem. 2011, 286, 42051–42062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucey, M.M.; Wang, Y.; Bustin, M.; Duncan, M.K. Differential expression of the HMGN family of chromatin proteins during ocular development. Gene Expr. Patterns 2008, 8, 433–437. [Google Scholar] [CrossRef] [Green Version]
- Li, D.D.; Guo, C.H.; Yue, L.; Duan, C.C.; Yang, Z.Q.; Cao, H.; Guo, B.; Yue, Z.P. Expression, regulation and function of Hmgn3 during decidualization in mice. Mol. Cell. Endocrinol. 2015, 413, 13–25. [Google Scholar] [CrossRef]
- Kurahashi, T.; Furusawa, T.; Ueda, T.; Bustin, M. The nucleosome binding protein HMGN3 is expressed in pancreatic alpha-cells and affects plasma glucagon levels in mice. J. Cell. Biochem. 2010, 109, 49–57. [Google Scholar]
- Lee, H.J.; Kim, J.Y.; Song, I.H.; Park, I.A.; Yu, J.H.; Ahn, J.H.; Gong, G. High mobility group B1 and N1 (HMGB1 and HMGN1) are associated with tumor-infiltrating lymphocytes in HER2-positive breast cancers. Virchows Arch. 2015, 467, 701–709. [Google Scholar] [CrossRef]
- Park, I.A.; Heo, S.H.; Song, I.H.; Kim, Y.A.; Park, H.S.; Bang, W.S.; Park, S.Y.; Jo, J.H.; Lee, H.J.; Gong, G. Endoplasmic reticulum stress induces secretion of high-mobility group proteins and is associated with tumor-infiltrating lymphocytes in triple-negative breast cancer. Oncotarget 2016, 7, 59957–59964. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Yang, F.; Jiang, X.; Yu, W.; Ren, X. High-mobility group nucleosome-binding protein 1 is a novel clinical biomarker in non-small cell lung cancer. Tumour Biol. 2015, 36, 9405–9410. [Google Scholar] [CrossRef]
- Morande, P.E.; Borge, M.; Abreu, C.; Galletti, J.; Zanetti, S.R.; Nannini, P.; Bezares, R.F.; Pantano, S.; Dighiero, G.; Oppezzo, P.; et al. Surface localization of high-mobility group nucleosome-binding protein 2 on leukemic B cells from patients with chronic lymphocytic leukemia is related to secondary autoimmune hemolytic anemia. Leuk. Lymphoma 2015, 56, 1115–1122. [Google Scholar] [CrossRef]
- Bustin, M.; Alfonso, P.J.; Pash, J.M.; Ward, J.M.; Gearhart, J.D.; Reeves, R.H. Characterization of transgenic mice with an increased content of chromosomal protein HMG-14 in their chromatin. DNA Cell Biol. 1995, 14, 997–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, A.A.; Chapuy, B.; Lin, C.Y.; Tivey, T.; Li, H.; Townsend, E.C.; van Bodegom, D.; Day, T.A.; Wu, S.C.; Liu, H.; et al. Triplication of a 21q22 region contributes to B cell transformation through HMGN1 overexpression and loss of histone H3 Lys27 trimethylation. Nat. Genet. 2014, 46, 618–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowery, C.T.; Reyes, J.M.; Cabal-Hierro, L.; Higby, K.J.; Karlin, K.L.; Wang, J.H.; Kimmerling, R.J.; Cejas, P.; Lim, K.; Li, H.; et al. Trisomy of a Down Syndrome Critical Region Globally Amplifies Transcription via HMGN1 Overexpression. Cell Rep. 2018, 25, 1898–1911.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauwecker, S.M.; Kim, J.J.; Licht, J.D.; Clevenger, C.V. Histone H1 and Chromosomal Protein HMGN2 Regulate Prolactin-induced STAT5 Transcription Factor Recruitment and Function in Breast Cancer Cells. J. Biol. Chem. 2017, 292, 2237–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medler, T.R.; Craig, J.M.; Fiorillo, A.A.; Feeney, Y.B.; Harrell, J.C.; Clevenger, C.V. HDAC6 Deacetylates HMGN2 to Regulate Stat5a Activity and Breast Cancer Growth. Mol. Cancer Res. 2016, 14, 994–1008. [Google Scholar] [CrossRef] [Green Version]
- Kugler, J.; Postnikov, Y.V.; Furusawa, T.; Kimura, S.; Bustin, M. Elevated HMGN4 expression potentiates thyroid tumorigenesis. Carcinogenesis 2017, 38, 391–401. [Google Scholar] [CrossRef]
- Lim, J.H.; Catez, F.; Birger, Y.; West, K.L.; Prymakowska-Bosak, M.; Postnikov, Y.V.; Bustin, M. Chromosomal protein HMGN1 modulates histone H3 phosphorylation. Mol. Cell 2004, 15, 573–584. [Google Scholar] [CrossRef]
- Rubinstein, Y.R.; Furusawa, T.; Lim, J.H.; Postnikov, Y.V.; West, K.L.; Birger, Y.; Lee, S.; Nguyen, P.; Trepel, J.B.; Bustin, M. Chromosomal protein HMGN1 modulates the expression of N-cadherin. FEBS J. 2005, 272, 5853–5863. [Google Scholar] [CrossRef] [Green Version]
- Postnikov, Y.V.; Furusawa, T.; Haines, D.C.; Factor, V.M.; Bustin, M. Loss of the nucleosome-binding protein HMGN1 affects the rate of N-nitrosodiethylamine-induced hepatocarcinogenesis in mice. Mol. Cancer Res. 2014, 12, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Birger, Y.; Catez, F.; Furusawa, T.; Lim, J.H.; Prymakowska-Bosak, M.; West, K.L.; Postnikov, Y.V.; Haines, D.C.; Bustin, M. Increased tumorigenicity and sensitivity to ionizing radiation upon loss of chromosomal protein HMGN1. Cancer Res. 2005, 65, 6711–6718. [Google Scholar] [CrossRef] [Green Version]
- West, K.L. HMGN proteins play roles in DNA repair and gene expression in mammalian cells. Biochem. Soc. Trans. 2004, 32, 918–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlitz, G. HMGNs, DNA repair and cancer. Biochim. Biophys. Acta 2010, 1799, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Strahler, J.R.; Dodgson, J.B. Neither HMG-14a nor HMG-17 gene function is required for growth of chicken DT40 cells or maintenance of DNaseI-hypersensitive sites. Nucleic Acids Res. 1997, 25, 283–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, M.; Gonzalez, R.W.; Patil, H.; Ueda, T.; Lim, J.H.; Kraemer, K.H.; Bustin, M.; Bergel, M. The nucleosome-binding protein HMGN2 modulates global genome repair. FEBS J. 2009, 276, 6646–6657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez, C.; Cruz-Becerra, G.; Fei, J.; Kassavetis, G.A.; Kadonaga, J.T. The tardigrade damage suppressor protein binds to nucleosomes and protects DNA from hydroxyl radicals. Elife 2019, 8, e47682. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Gerlitz, G.; Furusawa, T.; Catez, F.; Nussenzweig, A.; Oh, K.S.; Kraemer, K.H.; Shiloh, Y.; Bustin, M. Activation of ATM depends on chromatin interactions occurring before induction of DNA damage. Nat. Cell Biol. 2009, 11, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Gerlitz, G.; Bustin, M. Nucleosome binding proteins potentiate ATM activation and DNA damage response by modifying chromatin. Cell Cycle 2009, 8, 1641. [Google Scholar] [CrossRef] [Green Version]
- Masaoka, A.; Gassman, N.R.; Kedar, P.S.; Prasad, R.; Hou, E.W.; Horton, J.K.; Bustin, M.; Wilson, S.H. HMGN1 protein regulates poly(ADP-ribose) polymerase-1 (PARP-1) self-PARylation in mouse fibroblasts. J. Biol. Chem. 2012, 287, 27648–27658. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, K.; Herault, Y.; Lott, I.T.; Antonarakis, S.E.; Reeves, R.H.; Dierssen, M. Down syndrome: From understanding the neurobiology to therapy. J. Neurosci. 2010, 30, 14943–14945. [Google Scholar] [CrossRef] [Green Version]
- Cabin, D.E.; McKee-Johnson, J.W.; Matesic, L.E.; Wiltshire, T.; Rue, E.E.; Mjaatvedt, A.E.; Huo, Y.K.; Korenberg, J.R.; Reeves, R.H. Physical and comparative mapping of distal mouse chromosome 16. 5 p5. Genome Res. 1998, 8, 940–950. [Google Scholar] [CrossRef] [Green Version]
- Ling, K.H.; Hewitt, C.A.; Tan, K.L.; Cheah, P.S.; Vidyadaran, S.; Lai, M.I.; Lee, H.C.; Simpson, K.; Hyde, L.; Pritchard, M.A.; et al. Functional transcriptome analysis of the postnatal brain of the Ts1Cje mouse model for Down syndrome reveals global disruption of interferon-related molecular networks. BMC Genom. 2014, 15, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.D.; Liu, N.; Xu, X.J. Bioinformatics analysis of biomarkers and transcriptional factor motifs in Down syndrome. Braz. J. Med. Biol. Res. 2014, 47, 834–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carney, R.M.; Wolpert, C.M.; Ravan, S.A.; Shahbazian, M.; Ashley-Koch, A.; Cuccaro, M.L.; Vance, J.M.; Pericak-Vance, M.A. Identification of MeCP2 mutations in a series of females with autistic disorder. Pediatr. Neurol. 2003, 28, 205–211. [Google Scholar] [CrossRef]
- Yamazaki, M.; Itoh, S.; Sasaki, N.; Tanabe, K.; Uchiyama, M. Modification of the dialysis membrane method for drug release from suppositories. Pharm. Res. 1993, 10, 927–929. [Google Scholar] [CrossRef]
- Ramocki, M.B.; Tavyev, Y.J.; Peters, S.U. The MECP2 duplication syndrome. Am. J. Med. Genet. A 2010, 152A, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Bustin, M.; Reisch, J.; Einck, L.; Klippel, J.H. Autoantibodies to nucleosomal proteins: Antibodies to HMG-17 in autoimmune diseases. Science 1982, 215, 1245–1247. [Google Scholar] [CrossRef]
- Neuer, G.; Bautz, F.A.; Bustin, M.; Michels, H.; Truckenbrodt, H. Sera from JRA patients contain antibodies against a defined epitope in chromosomal protein HMG-17. Autoimmunity 1994, 17, 23–30. [Google Scholar] [CrossRef]
- Neuer, G.; Bustin, M.; Michels, H.; Truckenbrodt, H.; Bautz, F.A. Autoantibodies to the chromosomal protein HMG-17 in juvenile rheumatoid arthritis. Arthritis Rheumatol. 1992, 35, 472–475. [Google Scholar] [CrossRef]
- Vlachoyiannopoulos, P.G.; Boumba, V.A.; Tzioufas, A.G.; Seferiadis, C.; Tsolas, O.; Moutsopoulos, H.M. Autoantibodies to HMG-17 nucleosomal protein in patients with scleroderma. J. Autoimmun. 1994, 7, 193–201. [Google Scholar] [CrossRef]
- Zheng, S.; Ren, L.; Li, H.; Shen, X.; Yang, X.; Li, N.; Wang, X.; Guo, X.; Wang, X.; Huang, N. High-mobility group nucleosome-binding domain 2 protein inhibits the invasion of Klebsiella pneumoniae into mouse lungs in vivo. Mol. Med. Rep. 2015, 12, 1279–1285. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, N.; Wu, Q.; Wang, B. HMGN2: A novel antimicrobial effector molecule of human mononuclear leukocytes? J. Leukoc. Biol. 2005, 78, 1136–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.; Miao, J.; Zhang, F.; Xiong, F.; Zhu, F.; Li, J.; Wang, X.; Chen, S.; Chen, J.; Huang, N.; et al. Non-histone nuclear protein HMGN2 differently regulates the urothelium barrier function by altering expression of antimicrobial peptides and tight junction protein genes in UPEC J96-infected bladder epithelial cell monolayer. Acta Biochim. Pol. 2018, 65, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhu, F.; Yang, J.; Zhang, W.; Liu, K.; Ren, L.; Xiong, F.; Lu, K.; Li, D.; Zeng, R.; et al. High-mobility group protein N2 induces autophagy by activating AMPK/ULK1 pathway and thereby boosts UPEC proliferation within bladder epithelial cells. Biochem. Biophys. Res. Commun. 2019, 513, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, X.F.; Zhou, X.E.; Shi, Y.E.; Deng, L.X.; Ma, Y.; Wang, X.Y.; Li, J.Y.; Huang, N. Antibacterial mechanism of high-mobility group nucleosomal-binding domain 2 on the Gram-negative bacteria Escherichia coli. J. Zhejiang Univ. Sci. B 2017, 18, 410–420. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, S.; Ren, H.; Chen, J.; Li, J.; Wang, Y.; Hua, Y.; Wang, X.; Huang, N. HMGN2 regulates non-tuberculous mycobacteria survival via modulation of M1 macrophage polarization. J. Cell. Mol. Med. 2019, 23, 7985–7998. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Yang, H.; Tracey, K.J.; Bustin, M.; Oppenheim, J.J. High mobility group box-1 protein induces the migration and activation of human dendritic cells and acts as an alarmin. J. Leukoc. Biol. 2007, 81, 59–66. [Google Scholar] [CrossRef]
- Yang, D.; Postnikov, Y.V.; Li, Y.; Tewary, P.; de la Rosa, G.; Wei, F.; Klinman, D.; Gioannini, T.; Weiss, J.P.; Furusawa, T.; et al. High-mobility group nucleosome-binding protein 1 acts as an alarmin and is critical for lipopolysaccharide-induced immune responses. J. Exp. Med. 2012, 209, 157–171. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Yang, D.; Tewary, P.; Li, Y.; Li, S.; Chen, X.; Howard, O.M.; Bustin, M.; Oppenheim, J.J. The Alarmin HMGN1 contributes to antitumor immunity and is a potent immunoadjuvant. Cancer Res. 2014, 74, 5989–5998. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Yang, D.; Trivett, A.; Han, Z.; Xin, H.; Chen, X.; Oppenheim, J.J. Development of a Curative Therapeutic Vaccine (TheraVac) for the Treatment of Large Established Tumors. Sci. Rep. 2017, 7, 14186. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.M.; Yang, D.; Trivett, A.; Meyer, T.J.; Oppenheim, J.J. HMGN1 and R848 Synergistically Activate Dendritic Cells Using Multiple Signaling Pathways. Front. Immunol. 2018, 9, 2982. [Google Scholar] [CrossRef]
- Chen, C.Y.; Ueha, S.; Ishiwata, Y.; Yokochi, S.; Yang, D.; Oppenheim, J.J.; Ogiwara, H.; Shichino, S.; Deshimaru, S.; Shand, F.H.W.; et al. Combined treatment with HMGN1 and anti-CD4 depleting antibody reverses T cell exhaustion and exerts robust anti-tumor effects in mice. J. Immunother Cancer 2019, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zeng, Z.; Li, J.; Zheng, Y.; Wei, F.; Ren, X. PD-1/PD-L1 Axis, Rather Than High-Mobility Group Alarmins or CD8+ Tumor-Infiltrating Lymphocytes, Is Associated with Survival in Head and Neck Squamous Cell Carcinoma Patients Who Received Surgical Resection. Front. Oncol. 2018, 8, 604. [Google Scholar] [CrossRef] [PubMed]
- Mehnert, M.; Li, W.; Wu, C.; Salovska, B.; Liu, Y. Combining Rapid Data Independent Acquisition and CRISPR Gene Deletion for Studying Potential Protein Functions: A Case of HMGN1. Proteomics 2019, 19, e1800438. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Bustin, M.; Oppenheim, J.J. Harnessing the alarmin HMGN1 for anticancer therapy. Immunotherapy 2015, 7, 1129–1131. [Google Scholar] [CrossRef]
- Yang, D.; Han, Z.; Alam, M.M.; Oppenheim, J.J. High-mobility group nucleosome binding domain 1 (HMGN1) functions as a Th1-polarizing alarmin. Semin. Immunol. 2018, 38, 49–53. [Google Scholar] [CrossRef]
- Zhang, S.; Deng, T.; Tang, W.; He, B.; Furusawa, T.; Ambs, S.; Bustin, M. Epigenetic regulation of REX1 expression and chromatin binding specificity by HMGNs. Nucleic Acids Res. 2019, 47, 4449–4461. [Google Scholar] [CrossRef] [Green Version]
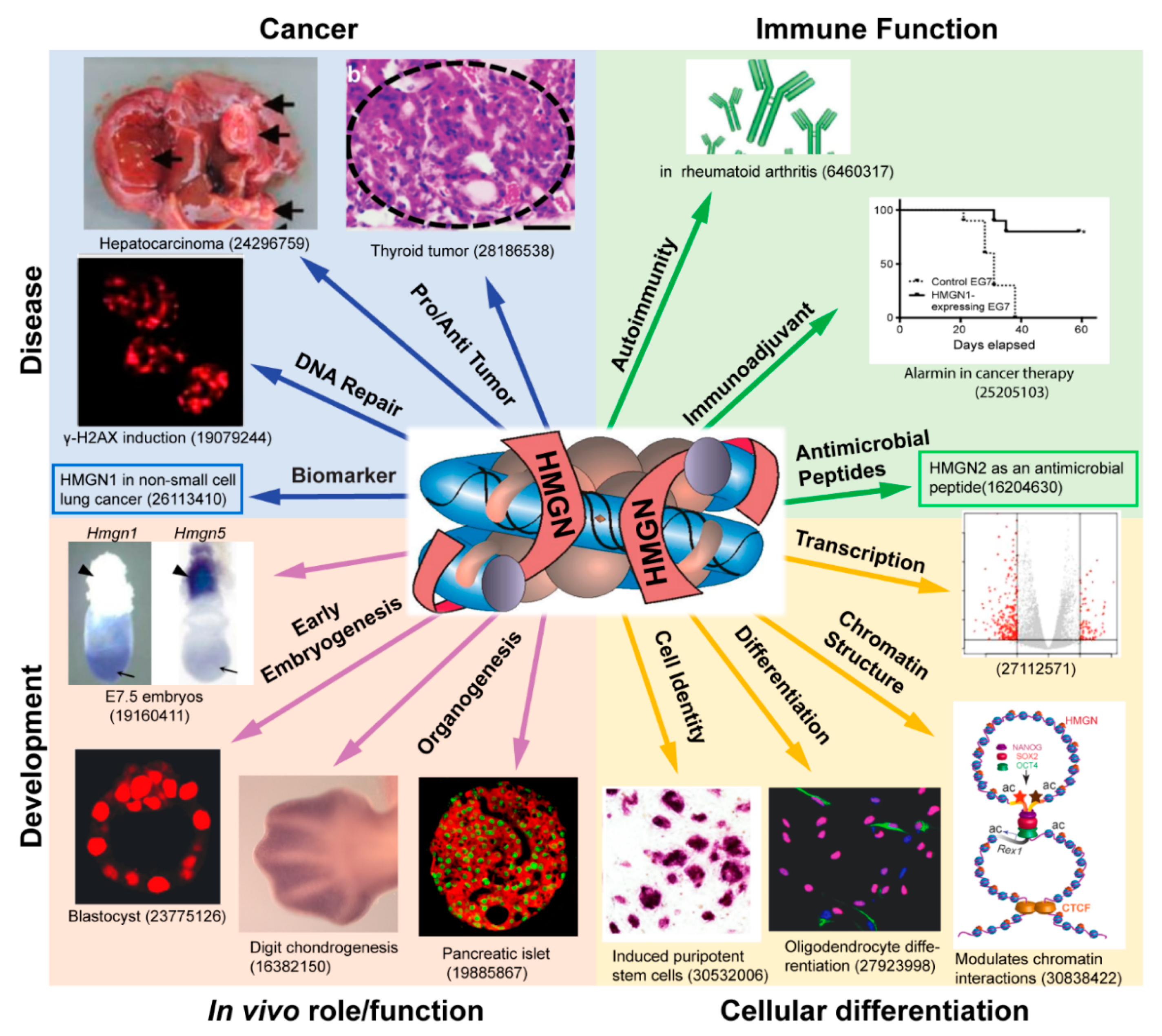
Figure 1.
HMGN proteins in development and disease. Shown is a schematic illustration of the biological processes reported to be affected by members of the HMGN protein family. The center panel depicts a nucleosome complexed with two molecules of HMGN protein. The number shown below each panel is the PMID of an article that discusses a role of HMGN in a specific biological process. Not all the biological processes affected by HMGN are depicted. A comprehensive list of reference to articles on the biological functions of HMGN is presented in Table 2 and Table 3.
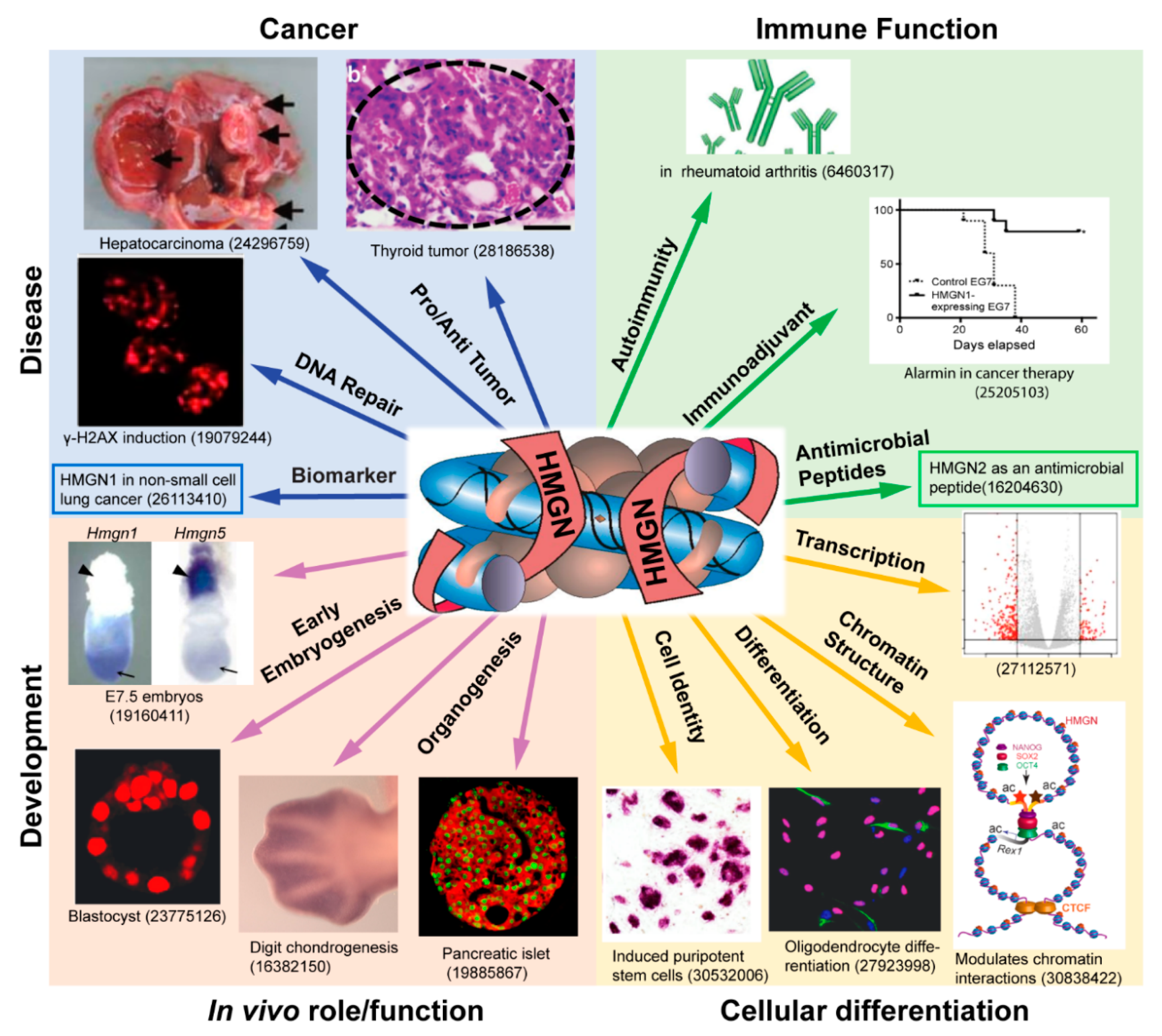
Figure 1.
HMGN proteins in development and disease. Shown is a schematic illustration of the biological processes reported to be affected by members of the HMGN protein family. The center panel depicts a nucleosome complexed with two molecules of HMGN protein. The number shown below each panel is the PMID of an article that discusses a role of HMGN in a specific biological process. Not all the biological processes affected by HMGN are depicted. A comprehensive list of reference to articles on the biological functions of HMGN is presented in Table 2 and Table 3.

{kind=link}
Table 1.
Genetically modified high mobility group N (HMGN) mice *.
| Gene | Modification | References |
|---|---|---|
| Hmgn1 | Knockout | Birger, Y. et al., 2003 |
| Hmgn2 | Knockout | Deng, T. et al., 2015 |
| Hmgn2# | Knockout | Gao, X.L. et al., 2019 |
| Hmgn3 | Knockout | Ueda, T. et al., 2009 |
| Hmgn5 | Knockout | Ciappio, E.D. et al., 2014 |
| Hmgn1/Hmgn2 | Knockout | Deng, T. et al., 2015 |
| Human HMGN1 | Transgenic | Bustin, M. et al., 1995 |
| Human HMGN4 | Transgenic | Kugler, J. et al., 2017 |
| Hmgn5 | Transgenic | Furusawa, T. et al., 2016 |
* All mouse lines listed except the line denoted by # were generated in the author’s laboratory. Additional commercial resources for genetically altered mice may be available.
Table 2.
HMGN in Development. Studies on the role of HMGN proteins in development and differentiation published in the English language. Reference given as PubMed Identifier (PMID).
Table 2.
HMGN in Development. Studies on the role of HMGN proteins in development and differentiation published in the English language. Reference given as PubMed Identifier (PMID).
| HMGN | Species | Study Model | Summary | Reference (PMID) |
|---|---|---|---|---|
| HMGN1 | Mouse | Myoblast differentiation | Overexpression of human HMGN1 inhibited the differentiation of mouse C2C12 myoblast cells into myotubes | 8514795 |
| HMGN1 | Mouse | Transcription in MEFs | HMGN1 is a negative regulator of N-Cadherin gene. N-Cadherin was also higher in the early embryos of HMGN1 KO compared to WT | 16279949 |
| HMGN1 | Mouse | Cornea differentiation and maturation | In corneal epithelium of Hmgn1 KO mice, corneal differentiation, maturation and gene expression were affected. In the wildtype mice, HMGN1 is colocalized with p63 and p63 expression was absent in the central region of the Hmgn1 KO cornea | 16466397 |
| HMGN1 | Mouse | Chondrocyte differentiation | In developing limb bud, Hmgn1 expression is inversely correlated to Sox9 expression and it is down regulated during chondrogenesis | 16382150 |
| HMGN1 | Mouse | Myogenesis and adipogenesis | Myogenesis or adipogenesis were induced in Hmgn1 knock down (KD) NIH3T3 cell line by overexpressing MyoD or C/EBPalpha. Hmgn1 Knockdown cells did not show any differences when compared with control cells | 16451822 |
| HMGN1 | Mouse | Digit formation | At E13.5 mouse autopod, interdigital expression of hmgn1 mRNA is highest at the digit-interdigit junction and is greatly down-regulated in the Raldh2 KO mice | 20034106 |
| HMGN1 | Mouse | embryonic stem cells (ESCs) | HMGN1 is strongly expressed in oocytes and throughout all preimplantation stages. Loss of HMGN1 reduces the DNase I hypersensitivity (DHS) at CpG island promoters in ES cells | 23775126 |
| HMGN1 | Human | Transgenic mice | HMGN1 is negative regulator for methyl CpG-binding protein 2. Hence, mice overexpressing HMGN1 displayed abnormalities in activity and anxiety and to social deficits | 22009741 |
| HMGN1 | Mouse | Down syndrome model mouse. | HMGN1 is overexpressed in Down syndrome and in all down syndrome model mice | 2140193 |
| 16635258 | ||||
| HMGN2 | Mouse | Kidney differentiation | During metanephric kidney development, HMGN2 expression is highly linked to induction and differentiation in kidney organogenesis | 11683498 |
| HMGN1 HMGN2 | Mouse | Myoblast differentiation | During myogenesis the level of Hmgn1 and Hmgn2 mRNA were decreased and this downregulation is a differentiation related event | 1689723 |
| HMGN1 HMGN2 | Chicken | Erythroid cells | Erythroid cells from 5-day chicken embryos contain higher level of Hmgn mRNAs than cells from 14-day embryos. Change from primitive to definitive erythroid lineage during embryogenesis is accompanied by a change in the expression of HMGN proteins. | 1993650 |
| HMGN1 HMGN2 | Rat Human | Osteoblast differentiation and monocyte differentiation | In both primary cultures of calvarial-derived rat osteoblasts and human leukemia HL-60 cells, HMGN1 and HMGN2 are preferentially expressed in the proliferating state, and the levels are down-regulated during differentiation. | 8496248 |
| HMGN1 HMGN2 | Mouse | Early embryonic development | HMGN1 and HMGN2 were detected throughout oogenesis and embryonic transcripts accumulate beyond the two-cell stage. Depletion of HMGNs in one-cell or two-cell mouse embryos delays subsequent progression. | 11133167 |
| HMGN1 HMGN2 | Xenopus | Early embryonic development | Depletion of Hmgn1 or Hmgn2 resulted in developmental defects after mid blastula transition (MBT) in Xenopus embryos, and re-expression of HMGN2 rescued these phenotypes | 14568106 |
| HMGN1 HMGN2 | Bovine | Maternal-zygotic transition | During bovine embryo early stage development, HMGN1 and HMGN2 are expressed in oocytes but are downregulated after fertilization and almost disappear by the 8-cell stage | 14960490 |
| HMGN1 HMGN2 | Mouse | Hair follicle cycle | Hmgn mRNAs was expressed at the onset of hair follicle development and during the adult hair follicle cycle | 19303948 |
| HMGN1 HMGN2 | Mouse | MEF, mouse behavior | Genome wide mapping of DHSs sites in MEFs from HMGN1 KO, HMGN2 KO and HMGN DKO revealed that loss of both, but not single HMGN variant leads to significant remodeling of DHS especially at enhancer regions | 26156321 |
| HMGN1 HMGN2 | Mouse | B lymphocyte | Loss of HMGN1 and HMGN2 proteins affect gene expression by modulating chromatin regulatory sites during activation of naïve B cells | 27112571 |
| HMGN1 HMGN2 | Mouse | Oligodendrocyte differentiation | Loss of HMGNs affect the expression of OLIG2 and OLIG1, thereby affecting oligodendrocyte development and nerve myelination, and mouse behavior | 27923998 |
| HMGN1 HMGN2 | Mouse | MEFs, induced pluripotent stem cells (iPSCs) | MEFs from Hmgn1/2 DKO mice showed enhanced rate of iPSCs induction by OSKM factors, and into neurons induced by ASCL1 | 30532006 |
| HMGN1 HMGN2 HMGN3 | Mouse | Eye development | HMGN1, HMGN2 and HMGN3 displayed dynamic and specific expression patterns during ocular development (E10.5, E13.5 and newborn) and adult eye | 18502697 |
| HMGN1 HMGN2 HMGN3 | Mouse | Astrocyte differentiation | Ectopic expression of either HMGN1, HMGN2 or HMGN3 in neural precursor cells (NPCs) or in late embryonic neocortex promoted the generation of astrocytes while knockdown suppressed astrocyte differentiation. | 25069414 |
| HMGN2 | Bovine | Early embryonic development | An in vitro fertilization study of bovine embryos revealed that embryos with enhanced levels of HMGN2 failed to develop into blastocyst due to the altered chromatin remodeling which might be caused by hyperacetylation of Histone H3 at lysine 14 residue | 17712799 |
| HMGN2 | Mouse | Tooth development | HMGN2 binds to Pitx2 and inhibits Pitx2 mediated Amelogenin expression during tooth development | 23975681 |
| HMGN2 | Mouse | Corticogenesis (Cerebral cortex development) | Hmgn2 KO mice exhibited microcephalus phenotype with reduced cortical surface area and almost normal radial corticogenesis | 31699896 |
| HMGN3 | Xenopus | Anuran metamorphosis | Hmgn3 expression is upregulated during thyroid hormone dependent anuran metamorphosis. | 11921340 |
| HMGN3 | Mouse | Retina | HMGN3 is highly expressed in glia cells and eye. HMGN3 binds to glycine transporter 1 (Glyt1) gene and upregulates its expression. | 15082770 |
| HMGN3 | Mouse | Pancreas (beta cells) | Loss of HMGN3 abrogates glucose stimulated insulin secretion and HMGN3 KO mice possess diabetic phenotype. HMGN3 along with PDX1regulates Glu2 gene | 19651901 |
| HMGN3 | Bovine | Early embryonic development | Hmgn3 mRNA was expressed at similar levels in bovine matured oocytes and 2–4-cell embryos, but expression was higher in 8-16-cell embryos, morulae and blastocysts | 19393058 |
| HMGN3 | Mouse | Pancreas (alpha cells) | HMGN3 is highly expressed in pancreatic alpha cells, and that Hmgn3 KO mice displayed reduced glucagon levels in their blood | 19885867 |
| HMGN3 | Mouse | Decidualization | HMGN3 is highly expressed in decidua and decidualizing stromal cells, and also in the artificially stimulated decidualization | 26112184 |
| HMGN5 | Mouse | Placenta development | Hmgn5 mRNA is strongly expressed in ectoplacental cone in E7.5 embryo and its expression retained in developing placenta and in trophoblast giant cells | 19160411 |
| HMGN5 | Mouse | Metabolomics | Metabolomic analysis of liver extracts and urine from Hmgn5 KO revealed that loss of HMGN5 leads to downregulation of glutathione peroxidase 6 (Gpx6) and hexokinase 1 (Hk1), thereby affects glutathione metabolism | 24392144 |
| HMGN5 | Mouse | Heart development | Transgenic mice overexpressing HMGN5 (both ubiquitous or heart specific) are normal at birth but develop hypertrophic heart with large cardiomyocytes, deformed nuclei and disrupted lamina and die of cardiac malfunction | 25609380 |
Table 3.
HMGN in Disease. Studies on the role of HMGN proteins in different diseases published in English language. Reference given as PMID.
Table 3.
HMGN in Disease. Studies on the role of HMGN proteins in different diseases published in English language. Reference given as PMID.
| HMGN | Study Model | Disease | Role/Function | Summary | Reference (PMID) |
|---|---|---|---|---|---|
| HMGN1 | Mice | Cancer | Immunoadjuvant | Augments antitumor immunity of alpha-fetoprotein expressing lentiviral vaccines in hepatocellular carcinoma mice | 31281528 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | Treatment with HMGN1 and anti-CD4 depleting antibody reverses T cell exhaustion and exerts robust anti-tumor effects in mice | 30696484 |
| HMGN1 | Cell culture | Cancer | Immunoadjuvant | HMGN1 promotes dendritic cells (DC) recruitment through interacting with a Gαi protein-coupled receptor and activates DCs predominantly through triggering toll like receptor 4 (TLR4) | 29503123 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | Intratumoral delivery of HMGN1 and R848 along with cytoxan (TheraVac) eradicated large established CT26, Renca and EG7 tumors in mice | 29079801 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | Intratumoral injections cured mice harboring large established subcutaneous Hepa1-6 hepatomas | 28881713 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | HMGN1 contributes to antitumor immunity | 25205103 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | HMGN1 induced DC maturation via TLR4, recruitment of antigen presenting cells at sites of injection and promoted antigen-specific immune response upon co-administration with antigens | 22184635 |
| HMGN1 | Mice | Cancer | Immunoadjuvant | DNA(TA) encapsulated within peptide hydrogels encodes for a melanoma-specific gp100 antigen fused to the HMGN1 enhanced immunostimulatory effects in tumor mice model | 25890750 |
| HMGN1 | Mice | Cancer | Protumor activity | Down syndrome associated triplication of a 21q22 region contributes to B cell proliferation with increased risk of B cell acute lymphoblastic leukemia through HMGN1 overexpression | 24747640 |
| HMGN1 | Cell culture | Cancer | Protumor activity | High expression of HMGN1 in highly metastatic cells of MDA-MB-435 suggests the association of HMGN1 with metastasis of breast cancer cells | 16979889 |
| HMGN1 | Patient’s studies | Cancer | Protumor activity | Endoplasmic reticulum stress induces secretion of HMGN1 and is associated with tumor-infiltrating lymphocytes in triple-negative breast cancer | 27494867 |
| HMGN1 | Patient’s studies | Cancer | Protumor activity | High cytoplasmic expression of HMGN1 was associated with a high histological grade, high levels of tumor infiltering lymphocytes (TILs) in HER2-positive breast cancer tissues | 26445971 |
| HMGN1 | Patient’s studies | Cancer | Biomarker | Patients with high serum HMGN1 had a poorer overall survival after curative pneumonectomy in non-small cell lung cancer | 26113410 |
| HMGN1 | Cell culture | Cancer | Transcriptional regulator | HMGN1 and HMGN2 are synthesized throughout the S-phase and persists through the cell cycle in Burkitt’s lymphoma | 8670211 |
| HMGN1 | Mice | Cancer | Antitumor activity | Loss of HMGN1 accelerates the progression of N-nitrosodiethylamine-induced hepatocarcinogenesis | 24296759 |
| HMGN1 | Mice | Cancer | Antitumor activity | Loss of HMGN1 increases the tumorigenicity and sensitivity to Ionizing radiations | 16061652 |
| HMGN1 | Patient’s studies | Down Syndrome | Transcriptional regulator | Gene dosage studies in cultured cells and brain tissue samples obtained from Down syndrome patients suggest that HMGN1 as contributing factor in the etiology of Down syndrome | 1825298 |
| HMGN1 | Mice | Down Syndrome | Gene locus | HMGN1 maps to the 21q22.3 in humans, the region associated with the pathogenesis of Down syndrome, and to chromosome 16 in mice | 2140193 |
| HMGN1 | Mice | Down Syndrome | Transcriptional regulator | Overexpression of the HMGN1 (encoded on chr21q22) reiterates transcriptional changes seen with triplication of Down syndrome critical region on distal chromosome 21 | 30428356 |
| HMGN1 | Mice | Diabetic nephropathy | Transcriptional regulator | HMGN1 expression correlates with epithelial mesenchymal transition, renal fibrosis in diabetic nephropathy mice | 31203283 |
| HMGN1 | Mice | Diabetic nephropathy | Transcriptional regulator | Insulin growth factor 1 receptor inhibitor attenuates diabetic nephropathy by suppressing HMGN1-TLR4 pathway | 29384065 |
| HMGN1 | Rats | Amnesia | Biomarker | Increased HMGN1 phosphorylation by protein kinase CK2 decreases amnesia in aged rats | 14635996 |
| HMGN1 | Patient’s studies | Systemic sclerosis | Immunogen in autoimmunity | Autoantibodies against HMGN1 and HMGN2 were found in systemic sclerosis patients’ sera | 7869312 |
| HMGN1 | Patient’s studies | Drug induced lupus | Immunogen in autoimmunity | Autoantibodies against HMGN1 and HMGN2 were found in drug induced lupus patients | 7907477 |
| HMGN1 | Patient’s studies | Primary pulmonary hypertension | Immunogen in autoimmunity | Autoantibodies against HMGN1 were found in sera from primary pulmonary hypertension patients | 8918589 |
| HMGN1 | Mice | Immune Tolerance | TLR-4 agonist | HMGN1 induces innate immune tolerance in a TLR4 dependent Sirtuin-1 mediated deacetylation in human blood peripheral mononuclear cells | 29593748 |
| HMGN2 | Mice | Cancer | Antitumor activity | The tumor volumes in SaO2 and U2-OS osteocarcinoma subcutaneous nude mouse models treated with overexpressing HMGN2 lentivirus were significantly decreased | 25530340 |
| HMGN2 | Mice | Cancer | Antitumor activity | Overexpression of HMGN2 significantly decreased migration capacity, increased apoptosis of MCF7 breast cancer cells both in vitro and in vivo | 30655878 |
| HMGN2 | Cell culture | Cancer | Protumor activity | HMGN2 was increased in tumor cell line MDA-MB-468 and also in metastatic oral squamous cell carcinoma tissues | 29541221 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | Isopentenyl pyrophosphate activated γδ T cells expresses high level of HMGN2 which contributes to antitumor activity | 29401165 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | Overexpression of HMGN2 in by lentivirus, inhibits the growth and migration of osteosarcoma cells | 25530340 |
| HMGN2 | Cell culture | Cancer | Signal transducer | Surface expressed of HMGN2 on leukemic B cells binds to autoantigen erythrocyte protein band 3 and contributes to initiation of autoimmune hemolytic anemia | 25156469 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | Phytohemagglutinin activated CD8 T cells releases high level of HMGN2 and kill tumor cells | 25060707 |
| HMGN2 | Cell culture | Cancer | Protumor activity | Prolactin induced H1 disassociation mediated by HMGN2 induce STAT5 recruitment and breast cancer pathogenesis | 28035005 |
| HMGN2 | Patient studies | Cancer | Protumor activity | Expression of HMGN2 was higher in human leukemia cells with chronic myelogenic leukemia | 8528142 |
| HMGN2 | Cell culture | Cancer | Protumor activity | HDAC6 mediated deacetylation of HMGN2 enhances Stat5a transcriptional activity, thereby regulating prolactin-induced gene transcription and breast cancer growth | 27358110 |
| HMGN2 | Cell culture | Cancer | Biomarker | HMGN1 and HMGN2 are synthesized throughout the S-phase and persists through the cell cycle in Burkitt’s lymphoma | 8670211 |
| HMGN2 | Cell culture | Cancer | Antitumor activity | HMGN2 is a direct target of MicroRNA-23a which plays crucial role in epithelial mesenchymal transition (EMT) in lung cancer cells suggests its role | 23437179 |
| HMGN2 | Mice | Cancer | Antitumor activity | HMGN2 exerts inhibitory effect on tumors in human tongue carcinoma transplanted nude mice | 24665631 |
| HMGN2 | Mice | Cancer | Antitumor activity | HMGN2 treatment inhibited the growth, increased apoptosis of Tca8113 cells in vitro and in vivo | 24348831 |
| HMGN2 | Cell culture | Cancer | Antimicrobial activity | 31-aa peptide (F3) in the HMGN2 sequence homed to HL-60 and MDA-MB-435 tumors upon I.v. injection | 12032302 |
| HMGN2 | Patient studies | Cancer | Immunogen in autoimmunity | High throughput screening of autoantibodies in ovarian cancer ascitic fluids revealed HMGN2 as an antigen | 31428516 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | HMGN2 regulates the expression of antimicrobial peptides in Uropathogenic Escherichia coli J96-infected bladder epithelial cell monolayer | 29549670 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 displayed anti-bacterial and anti-biofilm activity against Escherichia coli K12 | 28471113 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 inhibits Pseudomonas aeruginosa internalization in A549 cells | 28408162 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 protects bladder epithelial cells from Klebsiella invasion | 21720014 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | Knockdown of HMGN2 affected the survival of non-tuberculosis mycobacteria in macrophage | 31596045 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | HMGN2 is a target of miR-155, which is downregulated in Klebsiella pneumoniae infection suggested the role in controlling infection | 27534887 |
| HMGN2 | Cell culture | Infection | Transcriptional regulator | Knockdown of HMGN2 increases the internalization of Klebsiella pneumoniae by respiratory epithelial cells | 27460641 |
| HMGN2 | Mice | Infection | Antimicrobial activity | HMGN2 is crucial for LPS induced expression of murine β-defensin-3 and -4 in mice | 21594618 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 is crucial for LPS induced expression of murine β-defensin-3 and -4 in A549 cells | 21518253 |
| HMGN2 | Mice | Infection | Antimicrobial activity | HMGN2 inhibits the invasion of Klebsiella pneumoniae into mouse lungs in vivo | 25760831 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 inhibited hepatitis B virus infection and replication in HepG2.2.15 cell line | 19150374 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 inhibited internalization of Klebsiella pneumoniae into cultured bladder epithelial cells | 21778192 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | N-terminal amino sequence was PKRKAEGDAK of HMGN2 is a novel antimicrobial peptide | 16204630 |
| HMGN2 | Cell culture | Infection | Antimicrobial activity | HMGN2 showed potent antimicrobial activity against E coli ML-35p, P aeruginosa ATCC 27853 | 16115376 |
| HMGN2 | Patient’s studies | Systemic lupus erythromatosus | Immunogen in autoimmunity | Autoantibodies against HMGN2 were found in systemic lupus erythromatosis (SLE) patients | 8037838 |
| HMGN2 | Patient’s studies | Systemic lupus erythromatosus | Immunogen in autoimmunity | Autoantibodies to HMGN2 were found in different autoimmune diseases and are more frequent in Systemic lupus erythromatosus | 8318042 |
| HMGN2 | Patient’s studies | Systemic lupus erythromatosus | Immunogen in autoimmunity | Autoantibodies to HMGN2 were present in serums from patients with systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), and mixed connective tissue disease (MCTD) | 6460317 |
| HMGN2 | Patient’s studies | Drug induced lupus | Immunogen in autoimmunity | Autoantibodies against HMGN1 and HMGN2 were found in drug induced lupus patients | 7907477 |
| HMGN2 | Patient’s studies | Juvenile Rheumatoid Arthritis | Immunogen in autoimmunity | Autoantibodies to HMGN2 were detected in ANA-positive patients with periarticular-onset Juvenile Rheumatoid Arthritis | 7517709 |
| HMGN2 | Patient’s studies | Juvenile Rheumatoid Arthritis | Immunogen in autoimmunity | Autoantibodies to HMGN2 were detected in patients with Juvenile Rheumatoid Arthritis | 1567496 |
| HMGN2 | Patient’s studies | Periodontitis | Biomarker | HMGN2 in saliva of periodontitis patients was higher especially with severe periodontitis | 28591948 |
| HMGN2 | Mice | Microcephaly | Transcriptional regulator | HMGN2 protects against Microcephaly by maintaining chromatin accessibility during corticogenesis | 31699896 |
| HMGN3 | Mice | Diabetes | Anti-diabetic | Hmgn3-/- mice which have a mild diabetic phenotype, with reduced glucagon levels in the blood | 19885867 |
| HMGN3 | Mice | Diabetes | Anti-diabetic | In mice, loss of HMGN3 blunt glucose-stimulated insulin secretion and have a diabetic phenotype. HMGN3 regulates Glut4 gene in pancreatic beta cells | 19651901 |
| HMGN4 | Patient’s studies | Cancer | Biomarker | Patients with high expression of HMGN4 have high grade tumors, shorter overall survival in hepatocellular carcinoma | 30272824 |
| HMGN4 | Mice | Cancer | Protumor activity | Mouse and human cells overexpressing HMGN4 displayed increased tumorigenicity in thyroid tumor mouse model | 28186538 |
| HMGN5 | Cell culture | Cancer | Protumor activity | microRNA-183-3p directly inhibits the expression of HMGN5 and prostate cancer progression | 31314587 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 expression was upregulated in pancreatic ductal adenocarcinoma tissues and cell lines. | 30128022 |
| HMGN5 | Cell culture | Cancer | Protumor activity | miR-488 inhibits cell growth and metastasis in renal cell carcinoma by abrogating HMGN5 expression | 29713189 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 expression was significantly upregulated in bladder transitional cell carcinoma tissues | 29509244 |
| HMGN5 | Cell culture | Cancer | Protumor activity | Silencing HMGN5 promotes chemosensitivity of human bladder cancer cells to cisplatin | 29163683 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 was significantly upregulated in esophageal squamous cell carcinoma cells. Silencing of HMGN5 significantly inhibited cell growth and induced cell apoptosis of ESCC cells | 28914995 |
| HMGN5 | Cell culture | Cancer | Protumor activity | microRNA-495 inhibits the proliferation and invasion and induces the apoptosis of osteosarcoma cells by inhibiting HMGN5 | 28627703 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-140-5p expression enhances osteosarcoma chemoresistance by targeting HMGN5 and autophagy | 28341864 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-409-3p decreases glioma cell invasion and proliferation by targeting HMGN5 | 28109076 |
| HMGN5 | Cell culture | Cancer | Protumor activity | microRNA-340 inhibits tumorigenic potential of prostate cancer cells by abrogating HMGN5 expression | 26394192 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing enhances apoptosis, suppresses invasion and increases chemosensitivity to temozolomide in meningiomas | 26315299 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing suppresses the viability and invasion of human urothelial bladder cancer 5637 cells | 25796505 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 is over expressed in prostate cancer cells and activates MAPK and its expression is correlated with chemosensitivity | 25572120 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 is highly expressed in breast cancer cells and prompts proliferation and invasion | 25315189 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing in cells exhibit increased apoptosis rate and decreased colonogenic survival in response to 2–8 Gy ionizing radiation | 25307178 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 was positively correlated with pathologic staging and TNM staging of osteosarcoma | 24687550 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 increases drug resistance in osteosarcoma cell lines U-2OS and MG63 through upregulating autophagy | 24664583 |
| HMGN5 | Cell culture | Cancer | Protumor activity | Silencing of HMGN5 induces apoptosis through the modulation of a mitochondrial pathway and Bcl-2 family proteins in prostate cancer cells | 22504871 |
| HMGN5 | Cell culture | Cancer | Protumor activity | Knockdown of HMGN5 slowed down the cell cycle at the G0/G1 phase in human lung cancer cell lines A549 and H1299 | 22994738 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 silencing inhibits the growth and invasion of clear cell renal cell carcinoma cells and its expression is associated with tumor grade | 22420896 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 promotes the viability of bladder cancer cells through increased cell proliferation, invasion ability of metastatic bladder cancer cells through the upregulation of MMP-9 | 21695596 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown causes cell cycle arrest in human glioma U251 and U87 cells and it is highly expressed in high grade and low-grade tumor tissues | 21373965 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown causes cell cycle arrest in prostate cancer cell line DU145 | 20531280 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown inhibits proliferation and tumorigenicity of BGC823 and SGC7901 gastric cancer cell lines | 30854121 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-186 suppresses the growth and metastasis of bladder cancer by targeting HMGN5 | 26290438 |
| HMGN5 | Cell culture | Cancer | Protumor activity | HMGN5 knockdown inhibits proliferation and tumorogenecity of BGC823 and SGC7901 gastric cancer cell lines | 30854121 |
| HMGN5 | Cell culture | Cancer | Protumor activity | MicroRNA-186 suppresses the growth and metastasis of bladder cancer by targeting HMGN5 | 26290438 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nanduri, R.; Furusawa, T.; Bustin, M. Biological Functions of HMGN Chromosomal Proteins. Int. J. Mol. Sci. 2020, 21, 449. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020449
AMA Style
Nanduri R, Furusawa T, Bustin M. Biological Functions of HMGN Chromosomal Proteins. International Journal of Molecular Sciences. 2020; 21(2):449. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020449
Chicago/Turabian StyleNanduri, Ravikanth, Takashi Furusawa, and Michael Bustin. 2020. "Biological Functions of HMGN Chromosomal Proteins" International Journal of Molecular Sciences 21, no. 2: 449. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020449
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.