Visceral Adipose Tissue Inflammatory Factors (TNF-Alpha, SOCS3) in Gestational Diabetes (GDM): Epigenetics as a Clue in GDM Pathophysiology

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Study Cohort
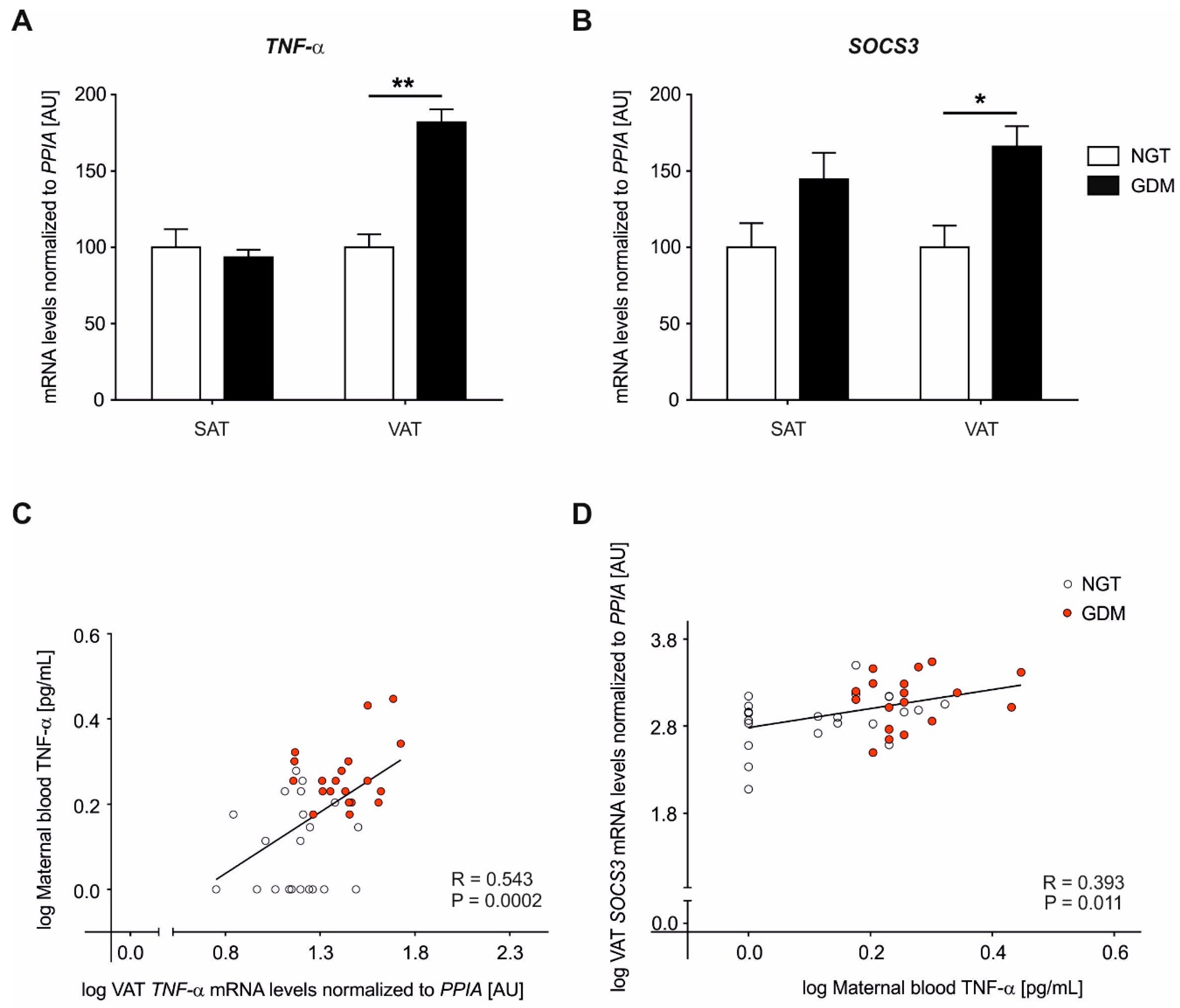
2.2. Adipose Tissue Gene Expression Analyses of TNF-α
2.3. Adipose Tissue Gene Expression Analyses of SOCS3
2.4. Maternal Circulating Plasma TNF-α Levels
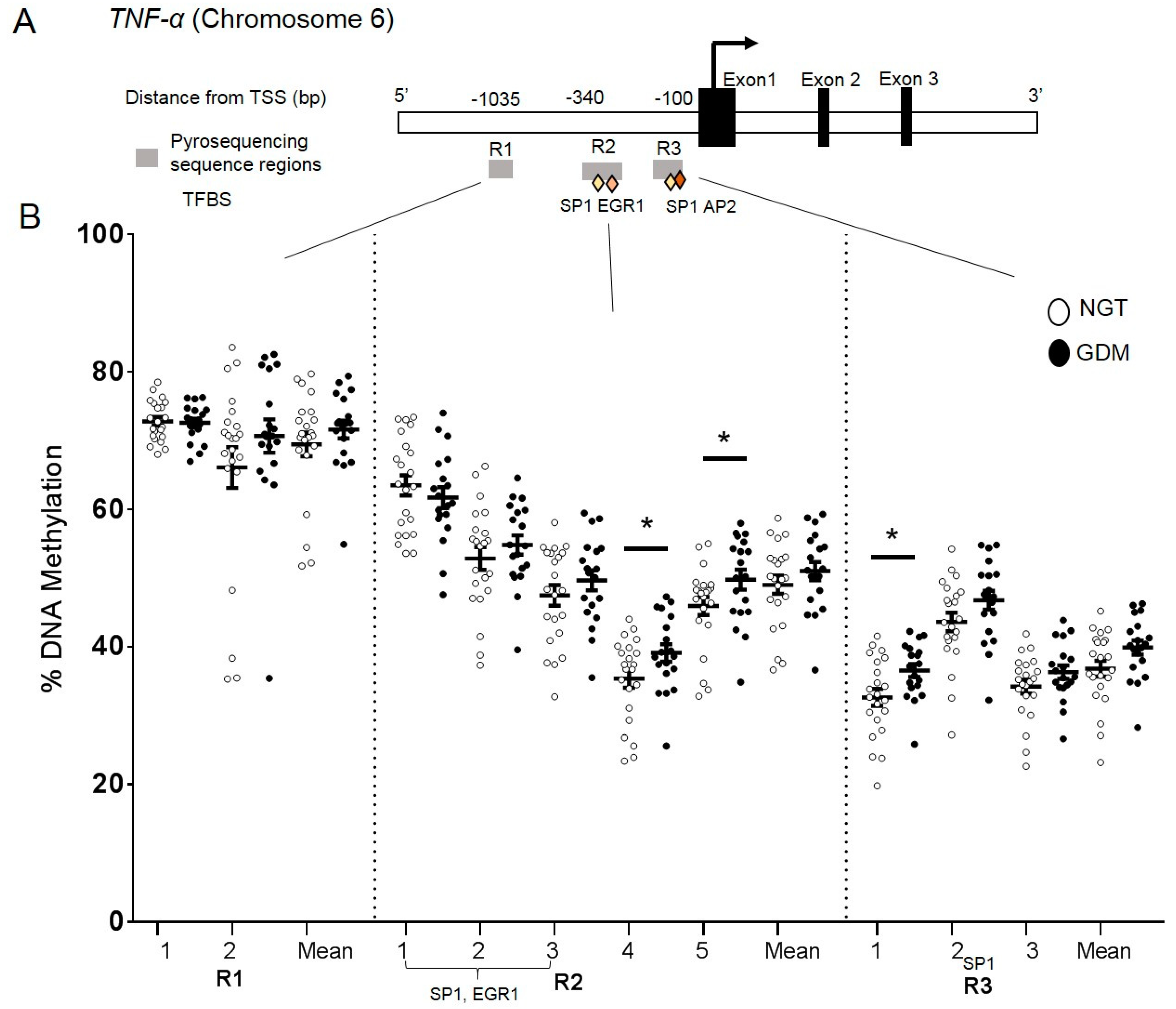
2.5. DNA Methylation at the TNF-α Promoter in Visceral Adipose Tissue
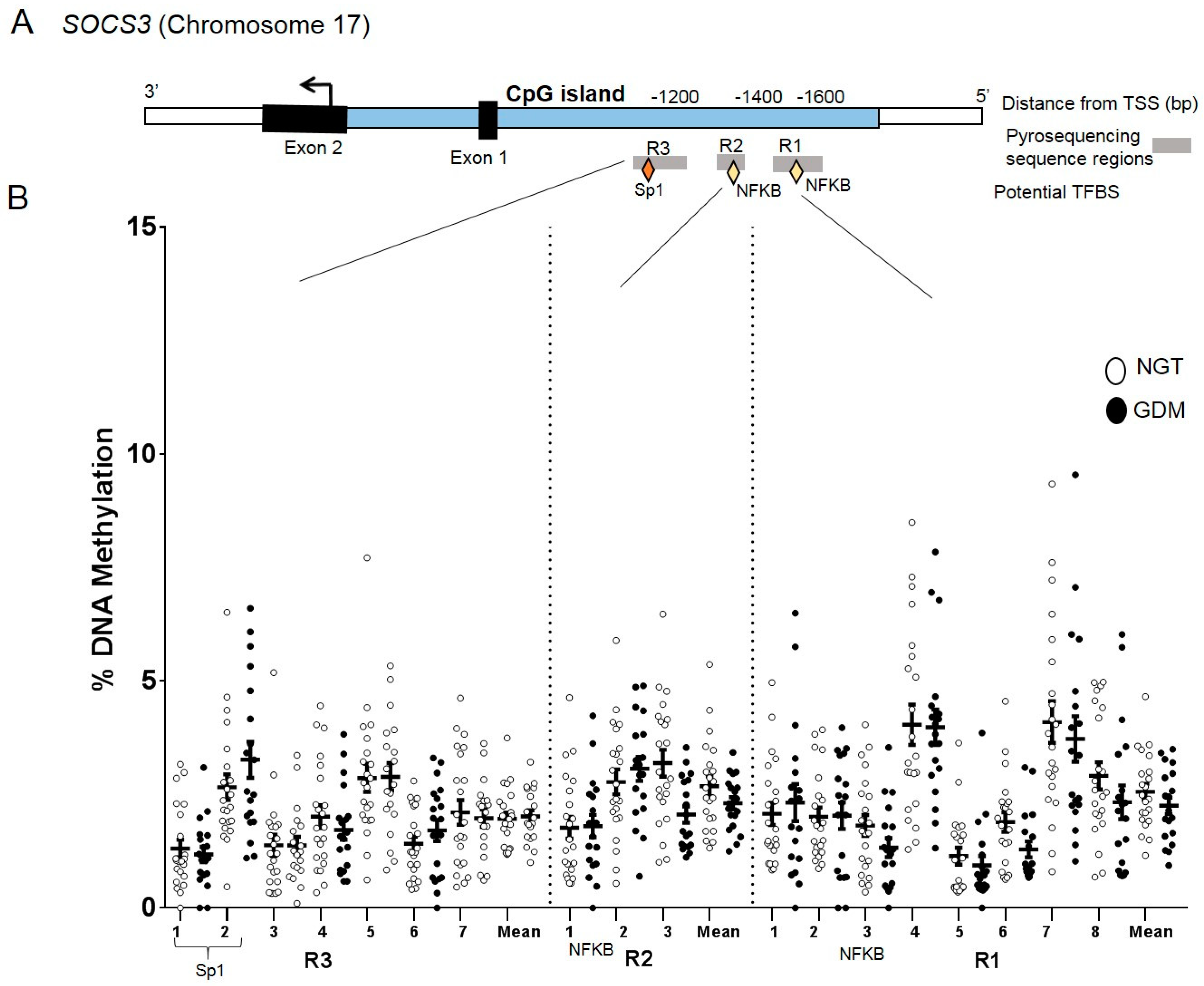
2.6. DNA Methylation at the SOCS3 Promoter in Visceral Adipose Tissue
3. Discussion
4. Materials and Methods
4.1. Subject Data
4.2. Maternal Blood and Adipose Tissue Sampling
4.3. Maternal Blood TNF-α Analysis
4.4. Gene Expression Analyses
4.5. DNA Methylation Analyses
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMI | Body-mass-index |
| CS | Cesarean section |
| GDM | Gestational diabetes mellitus |
| HOMA-IR | Homeostatic model assessment of insulin resistance |
| NGT | Normal glucose tolerance |
| oGTT | Oral glucose tolerance test |
| PPIA | Peptidylprolyl isomerase A |
| SAT | Subcutaneous adipose tissue |
| SOCS3 | Suppressor of cytokine signaling 3 |
| TNF-α | Tumor necrosis factor alpha |
| VAT | Visceral adipose tissue |
References
- Chen, C.; Xu, X.; Yan, Y. Estimated global overweight and obesity burden in pregnant women based on panel data model. PLoS ONE 2018, 13, e0202183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzago, M.; Fraticelli, F.; Stuppia, L.; Vitacolonna, E. Nutrigenetics, epigenetics and gestational diabetes: consequences in mother and child. Epigenetics 2019, 14, 215–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lain, K.Y.; Catalano, P.M. Metabolic changes in pregnancy. Clin. Obstet. Gynecol. 2007, 50, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Melchior, H.; Kurch-Bek, D.; Mund, M. The prevalence of gestational diabetes—A population based analysis of a nationwide screening program. Dtsch. Ärzteblatt Int 2017, 114, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Plagemann, A. Maternal diabetes and perinatal programming. Early Hum. Dev. 2011, 87, 743–747. [Google Scholar] [CrossRef]
- Sacks, D.A.; Hadden, D.R.; Maresh, M.; Deerochanawong, C.; Dyer, A.R.; Metzger, B.E.; Lowe, L.P.; Coustan, D.R.; Hod, M.; Oats, J.J.N.; et al. Frequency of Gestational Diabetes Mellitus at Collaborating Centers Based on IADPSG Consensus Panel–Recommended Criteria The Hyperglycemia and Adverse Pregnancy Outcome (HAPO) Study. Diabetes Care 2012, 35, 526–528. [Google Scholar] [CrossRef] [Green Version]
- Metzger, B.E.; Buchanan, T.A.; Coustan, D.R.; De Leiva, A.; Dunger, D.B.; Hadden, D.R.; Hod, M.; Kitzmiller, J.L.; Kjos, S.L.; Oats, J.N.; et al. Summary and recommendations of the Fifth International Workshop-Conference on Gestational Diabetes Mellitus. Diabetes Care 2007, 30, S251–S260. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, E.P.; Avalos, G.; O’Reilly, M.; Dennedy, M.C.; Gaffney, G.; Dunne, F.; the Atlantic DIP Collaborators. Atlantic Diabetes in Pregnancy (DIP): the prevalence and outcomes of gestational diabetes mellitus using new diagnostic criteria. Diabetologia 2011, 54, 1670–1675. [Google Scholar]
- Catalano, P.; DeMouzon, S.H. Maternal obesity and metabolic risk to the offspring: why lifestyle interventionsmay have not achieved the desired outcomes. Int. J. Obes. (Lond.) 2015, 39, 642–649. [Google Scholar] [CrossRef] [Green Version]
- Tchernof, A.; Després, J.P. Pathophysiology of human visceral obesity: An update. Physiol. Rev. 2013, 93, 359–404. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- De Souza, L.R.; Berger, H.; Retnakaran, R.; Maguire, J.L.; Nathens, A.B.; Connelly, P.W.; Ray, J.G. First-Trimester Maternal Abdominal Adiposity Predicts Dysglycemia and Gestational Diabetes Mellitus in Midpregnancy. Diabetes Care 2016, 39, 61–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano-Castillo, D.; Moreno-Indias, I.; Sanchez-Alcoholado, L.; Ramos-Molina, B.; Alcaide-Torres, J.; Morcillo, S.; Ocaña-Wilhelmi, L.; Tinahones, F.; Queipo-Ortuño, M.I.; Cardona, F. Altered Adipose Tissue DNA Methylation Status in Metabolic Syndrome: Relationships Between Global DNA Methylation and Specific Methylation at Adipogenic, Lipid Metabolism and Inflammatory Candidate Genes and Metabolic Variables. J. Clin. Med. 2019, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo-Flores, E.; Mancilla-Herrera, I.; Espino y Sosa, S.; Ortiz-Ramirez, M.; Flores-Rueda, V.; Ibargüengoitia-Ochoa, F.; Ibañez, C.A.; Zambrano, E.; Solis-Paredes, M.; Perichart-Perera, O.; et al. Macrophage Populations in Visceral Adipose Tissue from Pregnant Women: Potential Role of Obesity in Maternal Inflammation. Int. J. Mol. Sci. 2018, 19, 1074. [Google Scholar] [CrossRef] [Green Version]
- Reilly, S.M.; Saltiel, A.R. Adapting to obesity with adipose tissue inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef]
- Ott, R.; Stupin, J.H.; Melchior, K.; Schellong, K.; Ziska, T.; Dudenhausen, J.W.; Henrich, W.; Rancourt, R.C.; Plagemann, A. Alterations of adiponectin gene expression and DNA methylation in adipose tissues and blood cells are associated with gestational diabetes and neonatal outcome. Clin. Epigenetics 2018, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Ott, R.; Melchior, K.; Stupin, J.H.; Ziska, T.; Schellong, K.; Henrich, W.; Rancourt, R.C.; Plagemann, A. Reduced insulin receptor expression and altered DNA methylation in fat tissues and blood of women with GDM and offspring. JCEM 2019, 104, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Rancourt, R.C.; Ott, R.; Schellong, K.; Melchior, K.; Ziska, T.; Henrich, W.; Plagemann, A. Visceral adipose tissue alteration of PI3KR1 expression is associated with gestational diabetes but not promoter DNA methylation. Adipocyte 2019, 8, 339–346. [Google Scholar] [CrossRef]
- Ahima, R.S.; Stanley, T.L.; Khor, V.K.; Zanni, M.V.; Grinspoon, S.K. Estrogen sulfotransferase is expressed in subcutaneous adipose tissue of obese humans in association with TNF-alpha and SOCS3. J. Clin. Endocrinol. Metab. 2011, 96, E1153–E1158. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Gan, L.; Zhou, Z.; Jin, W.; Sun, C. SOCS3 promotes inflammation and apoptosis via inhibiting JAK2/STAT3 signaling pathway in 3T3-L1 adipocyte. Immunobiology 2015, 220, 947–953. [Google Scholar] [CrossRef]
- Emanuelli, B.; Peraldi, P.; Filloux, C.; Chavey, C.; Freidinger, K.; Hilton, D.J.; Hotamisligil, G.S.; Van Obberghen, E. SOCS-3 inhibits insulin signaling and is up-regulated in response to tumor necrosis factor-alpha in the adipose tissue of obese mice. J. Biol. Chem. 2001, 276, 47944–47949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, A.; Akhter, N.; Al-Roub, A.; Thomas, R.; Kochumon, S.; Wilson, A.; Koshy, M.; Al-Ozairi, E.; Al-Mulla, F.; Ahmad, R. TNF-α in Combination with Palmitate Enhances IL-8 Production via The MyD88- Independent TLR4 Signaling Pathway: Potential Relevance to Metabolic Inflammation. Int. J. Mol. Sci. 2019, 20, 4112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanti, J.F.; Ceppo, F.; Jager, J.; Berthou, F. Implication of inflammatory signaling pathways in obesity-induced insulin resistance. Front. Endocrinol. (Lausanne) 2013, 3, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlting, C.; Lai, W.S.; Schaper, F.; Brenndörfer, E.D.; Matthes, R.J.; Heinrich, P.C.; Ludwig, S.; Blackshear, P.J.; Gaestel, M.; Häussinger, D.; et al. Regulation of suppressor of cytokine signaling 3 (SOCS3) mRNA stability by TNF-alpha involves activation of the MKK6/p38MAPK/MK2 cascade. J. Immunol. 2007, 178, 2813–2826. [Google Scholar] [CrossRef] [Green Version]
- Abell, S.; De Courten, B.; Boyle, J.; Teede, H. Inflammatory and other biomarkers: Role in pathophysiology and prediction of gestational diabetes mellitus. Int. J. Mol. Sci. 2015, 16, 13442–13473. [Google Scholar] [CrossRef]
- Lekva, T.; Norwitz, E.R.; Aukrust, P.; Ueland, T. Impact of Systemic Inflammation on the Progression of Gestational Diabetes Mellitus. Curr. Diabetes Rep. 2016, 16, 26. [Google Scholar] [CrossRef]
- Dong, Y.; Chauhan, M.; Betancourt, A.; Belfort, M.; Yallampalli, C. Adipose Tissue Inflammation and Adrenomedullin Overexpression Contribute to Lipid Dysregulation in Diabetic Pregnancies. J. Clin. Endocrinol. Metab. 2018, 103, 3810–3818. [Google Scholar] [CrossRef] [Green Version]
- Cawthorn, W.P.; Sethi, J.K. TNF-α and adipocyte biology. FEBS Lett. 2008, 582, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose tissue in obesity-related inflammation and insulin resistance: Cells, cytokines, and chemokines. ISRN Inflamm. 2013, 2013, 139239. [Google Scholar] [CrossRef] [Green Version]
- Pantham, P.; Aye, I.L.; Powell, T.L. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta 2015, 36, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Moller, D.E. Potential role of TNF-alpha in the pathogenesis of insulin resistance and type 2 diabetes. Trends Endocrinol. Metab. 2000, 11, 212–217. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Peraldi, P.; Budavari, A.; Ellis, R.; White, M.F.; Spiegelman, B.M. IRS-1-Mediated Inhibition of Insulin Receptor Tyrosine Kinase Activity in TNF-α- and Obesity-Induced Insulin Resistance. Science 1996, 271, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Kanety, H.; Feinstein, R.; Papa, M.Z.; Hemi, R.; Karasik, A. Tumor necrosis factor alpha-induced phosphorylation of insulin receptor substrate-1 (IRS-1). Possible mechanism for suppression of insulin-stimulated tyrosine phosphorylation of IRS-1. J. Biol. Chem. 1995, 270, 23780–23784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, K.; Rakesh, K.; Pankajakshan, D.; Agrawal, D.K. SOCS3 promotor hypermethylation and STAT3-NF-κB interaction downregulate SOCS3 expression in human coronary artery smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H776–H785. [Google Scholar] [CrossRef] [Green Version]
- Ronn, S.G.; Billestrup, N.; Mandrup-Poulsen, T. Diabetes and Suppressors of Cytokine Signaling Proteins. Diabetes 2007, 56, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Kleiblova, P.; Dostalova, I.; Bartlova, M.; Lacinova, Z.; Ticha, I.; Krejci, V.; Springer, D.; Kleibl, Z.; Haluzik, M. Expression of adipokines and estrogen receptors in adipose tissue and placenta of patients with gestational diabetes mellitus. Mol. Cell. Endocrinol. 2010, 314, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Tsiotra, P.C.; Halvatsiotis, P.; Patsouras, K.; Maratou, E.; Salamalekis, G.; Raptis, S.A.; Dimitriadis, G.; Boutati, E. Circulating adipokines and mRNA expression in adipose tissue and the placenta in women with gestational diabetes mellitus. Peptides 2018, 101, 157–166. [Google Scholar] [CrossRef]
- Santangelo, C.; Filardi, T.; Perrone, G.; Mariani, M.; Mari, E.; Scazzocchio, B.; Masella, R.; Brunelli, R.; Lenzi, A.; Zicari, A.; et al. Cross-talk between fetal membranes and visceral adipose tissue involves HMGB1-RAGE and VIP-VPAC2 pathways in human gestational diabetes mellitus. Acta Diabetol. 2019, 56, 681–689. [Google Scholar] [CrossRef]
- Deng, X.; Yang, Y.; Sun, H.; Qi, W.; Duan, Y.; Qian, Y. Analysis of whole genome-wide methylation and gene expression profiles in visceral omental adipose tissue of pregnancies with gestational diabetes mellitus. J. Chin. Med. Assoc. 2018, 81, 623–630. [Google Scholar] [CrossRef]
- Qian, Y.; Sun, H.; Xiao, H.; Ma, M.; Xiao, X.; Qu, Q. Microarray analysis of differentially expressed genes and their functions in omental visceral adipose tissues of pregnant women with vs. without gestational diabetes mellitus. Biomed. Rep. 2017, 6, 503–512. [Google Scholar] [CrossRef] [Green Version]
- D’Ambrosi, F.; Crovetto, F.; Colosi, E.; Fabietti, I.; Carbone, F.; Tassis, B.; Motta, S.; Bulfoni, A.; Fedele, L.; Rossi, G.; et al. Maternal Subcutaneous and Visceral Adipose Ultrasound Thickness in Women with Gestational Diabetes Mellitus at 24–28 Weeks’ Gestation. Fetal Diagn. Ther. 2018, 43, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Gur, E.B.; Ince, O.; Turan, G.A.; Karadeniz, M.; Tatar, S.; Celik, E.; Yalcin, M.; Guclu, S. Ultrasonographic visceral fat thickness in the first trimester can predict metabolic syndrome and gestational diabetes mellitus. Endocrine 2014, 47, 478–484. [Google Scholar] [CrossRef]
- Kelstrup, L.; Hjort, L.; Houshmand-Oeregaard, A.; Clausen, T.D.; Hansen, N.S.; Broholm, C.; Borch-Johnsen, L.; Mathiesen, E.R.; Vaag, A.A.; Damm, P. Gene Expression and DNA Methylation of PPARGC1A in Muscle and Adipose Tissue From Adult Offspring of Women With Diabetes in Pregnancy. Diabetes 2016, 65, 2900–2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, R.; Stupin, J.H.; Loui, A.; Eilers, E.; Melchior, K.; Rancourt, R.C.; Schellong, K.; Ziska, T.; Dudenhausen, J.W.; Henrich, W.; et al. Maternal overweight is not an independent risk factor for increased birth weight, leptin and insulin in newborns of gestational diabetic women: Observations from the prospective “EaCH” cohort study. BMC Pregnancy Childbirth 2018, 18, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutsche Gesellschaft für Gynäkologie und Geburtshilfe. Diabetes und Schwangerschaft [Internet] 2008. Available online: https://www.awmf.org/uploads/tx_szleitlinien/057-008l_S3_Gestationsdiabetes-mellitus-GDM-Diagnostik-Therapie-Nachsorge_2019-06.pdf (accessed on 19 November 2019).
- Kleinwechter, H. Gestationsdiabetes mellitus (GDM). Dtsch. Med. Wochenschr. 2012, 137, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.R.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Neville, M.J.; Collins, J.M.; Gloyn, A.L.; McCarthy, M.I.; Karpe, F. Comprehensive human adipose tissue mRNA and microRNA endogenous control selection for quantitative real-time-PCR normalization. Obesity 2011, 19, 888–892. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Stahlberg, A.; Kubista, M.; Pfaffl, M. Comparison of reverse transcriptases in gene expression analysis. Clin. Chem. 2004, 50, 1678–1680. [Google Scholar] [CrossRef]
- Sato, H.; Watanabe, A.; Tanaka, T.; Koitabashi, N.; Arai, M.; Kurabayashi, M.; Yokoyama, T. Regulation of the human tumor necrosis factor-alpha promoter by angiotensin II and lipopolysaccharide in cardiac fibroblasts: different cis-acting promoter sequences and transcriptional factors. J. Mol. Cell. Cardiol. 2003, 35, 1197–1205. [Google Scholar] [CrossRef]
- Tsai, E.Y.; Falvo, J.V.; Tsytsykova, A.V.; Barczak, A.K.; Reimold, A.M.; Glimcher, L.H.; Fenton, M.J.; Gordon, D.C.; Dunn, I.F.; Goldfeld, A.E. A lipopolysaccharide-specific enhancer complex involving Ets, Elk-1, Sp1, and CREB binding protein and p300 is recruited to the tumor necrosis factor alpha promoter in vivo. Mol. Cell. Biol. 2000, 20, 6084–6094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowers, I.R.; Walters, K.; Kiss-Toth, E.; Read, R.C.; Duff, G.W.; Wilson, A.G. Age-related loss of CpG methylation in the tumour necrosis factor promoter. Cytokine 2011, 56, 792–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| NGT n = 22 [18] | GDM n = 19 | p-Value * | Spearman’s vs. MB TNF-α r (p-Value *) | |
|---|---|---|---|---|
| Maternal age (years) | 32.0 ± 5.3 | 32.5 ± 4.2 | 0.72 | 0.043 (0.78) |
| Prepregnancy BMI (kg/m2) | 26.8 ± 7.9 | 28.2 ± 6.7 | 0.32 | 0.107 (0.50) |
| BMI at delivery (kg/m2) | 33.1 ± 9.2 | 33.2 ± 6.6 | 0.50 | 0.099 (0.53) |
| Blood glucose at oGTT (mg/dL) | ||||
| Fasting | 79.5 ± 8.1 | 100 ± 30.9 | <0.0004 | 0.380 (0.01) |
| 1-h | 120.9 ± 29.3 | 213 ± 36.8 | <0.0001 | 0.505 (0.0007) |
| 2-h | 90.3 ± 19.9 | 167 ± 49.5 | <0.0001 | 0.516 (0.0006) |
| Area under the curve (mg/dL*h) | 205.8 ± 38.3 | 346 ± 71.9 | <0.0001 | 0.516 (0.0006) |
| Maternal fasting plasma levels at delivery: | ||||
| Glucose (mg/dL) | 71.1 ± 10.7 | 82.8 ± 8.4 | 0.001 | 0.319 (0.04) |
| Insulin (µU/mL) | 21.5 ± 16.1 | 40.9 ± 36.8 | 0.05 | 0.111 (0.48) |
| HOMA-IR | 3.2 ± 1.3 | 8.4 ± 7.4 | 0.006 | 0.208 (0.19) |
| C-peptide (ng/mL) | 2.0 ± 0.8 | 5.0 ± 3.1 | <0.0001 | 0.338 (0.03) |
| TNF-α (pg/mL) | 0.32 ± 0.35 | 0.86 ± 0.35 | <0.0001 | n.a. |
| Infant parameters: | ||||
| Birth weight (g) | 3365 ± 495.9 | 3585 ± 454.4 | 0.06 | 0.064 (0.68) |
| Relative Birth weight (g/cm) | 66.4 ± 6.8 | 70.16 ± 8.1 | 0.10 | 0.029 (0.85) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rancourt, R.C.; Ott, R.; Ziska, T.; Schellong, K.; Melchior, K.; Henrich, W.; Plagemann, A. Visceral Adipose Tissue Inflammatory Factors (TNF-Alpha, SOCS3) in Gestational Diabetes (GDM): Epigenetics as a Clue in GDM Pathophysiology. Int. J. Mol. Sci. 2020, 21, 479. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020479
Rancourt RC, Ott R, Ziska T, Schellong K, Melchior K, Henrich W, Plagemann A. Visceral Adipose Tissue Inflammatory Factors (TNF-Alpha, SOCS3) in Gestational Diabetes (GDM): Epigenetics as a Clue in GDM Pathophysiology. International Journal of Molecular Sciences. 2020; 21(2):479. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020479
Chicago/Turabian StyleRancourt, Rebecca C., Raffael Ott, Thomas Ziska, Karen Schellong, Kerstin Melchior, Wolfgang Henrich, and Andreas Plagemann. 2020. "Visceral Adipose Tissue Inflammatory Factors (TNF-Alpha, SOCS3) in Gestational Diabetes (GDM): Epigenetics as a Clue in GDM Pathophysiology" International Journal of Molecular Sciences 21, no. 2: 479. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020479