Lymphocyte Landscape after Chronic Hepatitis C Virus (HCV) Cure: The New Normal

Institute of Human Virology, University of Maryland School of Medicine, Baltimore, MD 21201, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(20), 7473; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207473
Submission received: 9 September 2020
/
Revised: 5 October 2020
/
Accepted: 7 October 2020
/
Published: 10 October 2020
(This article belongs to the Special Issue Anti-viral Therapy for Chronic Hepatitis)
Abstract
:Chronic HCV (CHC) infection is the only chronic viral infection for which curative treatments have been discovered. These direct acting antiviral (DAA) agents target specific steps in the viral replication cycle with remarkable efficacy and result in sustained virologic response (SVR) or cure in high (>95%) proportions of patients. These treatments became available 6–7 years ago and it is estimated that their real impact on HCV related morbidity, including outcomes such as cirrhosis and hepatocellular carcinoma (HCC), will not be known for the next decade or so. The immune system of a chronically infected patient is severely dysregulated and questions remain regarding the immune system’s capacity in limiting liver pathology in a cured individual. Another important consequence of impaired immunity in patients cleared of HCV with DAA will be the inability to generate protective immunity against possible re-infection, necessitating retreatments or developing a prophylactic vaccine. Thus, the impact of viral clearance on restoring immune homeostasis is being investigated by many groups. Among the important questions that need to be answered are how much the immune system normalizes with cure, how long after viral clearance this recalibration occurs, what are the consequences of persisting immune defects for protection from re-infection in vulnerable populations, and does viral clearance reduce liver pathology and the risk of developing hepatocellular carcinoma in individuals cured with these agents. Here, we review the recent literature that describes the defects present in various lymphocyte populations in a CHC patient and their status after viral clearance using DAA treatments.
1. Introduction
The ability to cure chronic hepatitis C virus (HCV) infection with direct acting antiviral (DAA) treatments [1] has provided scientists with a unique opportunity to investigate the immune recalibration that is possible once a chronic viral antigen has been removed. This is also an excellent model system to study basic immunology questions such as plasticity of the immune system and epigenetic regulation of immune responses in chronic infections and their reversibility. Our group has conducted pioneer clinical trials of DAA for HCV cure [2,3,4,5,6,7,8] and we investigate immune dysfunction in chronic HCV (CHC) and its recovery in cured individuals [9,10,11,12,13,14]. This focused review provides current understanding of innate and adaptive lymphocytes perturbations in CHC patients, the status of these immune cells after viral clearance with DAA, and implications thereof (summarized in Table 1 and in Figure 1).
2. Innate Immune Lymphocytes during and after Clearance of Chronic HCV Infection
Innate lymphoid cells are abundantly present in the liver [81,82], which is the site of HCV replication, and these are expected to provide the first line of defense against hepatotropic pathogens. A paradox common to most innate lymphoid cells is their function as early antiviral effector cells and their contribution to liver tissue injury by way of producing pro-inflammatory cytokines in response to pathogens.
2.1. Natural Killer (NK) Cells
Natural Killer (NK) cells are the dominant lymphocyte population in the liver and contribute to control of HCV infection as well perform anticancer effector function [83,84,85]. NK cells are a diverse population with multiple subtypes defined by unique combinations of surface markers. Traditionally, these cells are divided into types based on surface expression levels of CD16 and CD56 [83]. CD56brightCD16dim NK cells produce antiviral cytokines [84], while CD56dim/-CD16brigh NK cells express higher levels of killer immunoglobulin-like receptors (KIR) and perforin and are cytotoxic [83,85]. Additionally, activating and inhibitory NK cell receptors, NKG2D, NKp30, NKp46, and NKG2A, respectively, regulate NK cell functionality [86], and the balance shifts from inhibition to activation during HCV infection [15,16].
NK cells have been well investigated during and after cure of chronic HCV infection. The NK cell landscape is altered in the blood of chronically infected patients [17,18,19], reflected in increased frequencies of CD56bright and decreased frequencies of CD56dim subsets; DAA treatment-mediated HCV clearance results in an increase in CD56dim and a decrease in CD56bright NK cell frequencies, resulting in normalization [18]. Most of the other alterations in NK cell phenotypic markers observed in a chronically infected patient, including NKG2A, CD94, NKp30, NKp46, IFNγ, and perforin expression, also normalize rapidly upon virus clearance [19]. The majority of these changes are observed around week 12 or 24 after treatment with sofosbuvir and ledipasvir. In patients with undetectable viral loads treated with daclatasvir/asunaprevir, NKp46 and NKG2A expression levels normalized by week 8. The expression of inhibitory receptor NKG2A [19,20,21] and activating receptors NKp30 [18,19,22] and NKp46 [22,23] that increase on NK cells during HCV infection is also reduced with DAA treatment [18,19,22,24]. Expression of activating receptor NKG2D in chronic HCV infection remains unchanged during infection and with a cure in most studies [19,22,25,26] or was shown decreased during infection [27].
Functional impairment in NK cells during chronic infection is consistently reflected in reduced IFNγ production and higher CD107a degranulation response; these alterations return to normal responses upon virus clearance [17,22]. Whether improved NK cell functional responses upon virus clearance are indicative of generation of NK cell types which possess antigen-specific “memory” was investigated by Wijaya et al. [28]. This study showed that NK cells from patients have a stronger cytotoxic response against HCV infected cell lines compared with NK cells from healthy individuals, suggesting an antigen-specific response. Further, this cytotoxic ability was increased at 12 weeks after virus clearance. KLRG1 was identified as the marker of these antigen-specific NK cells; KLRG1+CD56dim HCV-specific memory NK cells thus develop following HCV infection and may have relevance for HCV vaccine design.
A novel subset of “adaptive” NK cells representing a minor proportion of bulk NK cells, has been identified recently [29]. Compared with conventional NK cells, these CD57+ cells express lower levels of intracellular γ-signaling chain of Fc receptor (FcεRIγ), Siglec-7, NKG2A, NKp30, and Tim-3, higher levels of NKG2C and KIR, and possess higher ADCC activity. During HCV infection, an association of these adaptive NK cells with low levels of liver enzymes and fibrosis suggests a possible role of these cells in limiting fibrosis progression [87]. Phenotypically, the proportion of these cells is higher and they have increased PD-1 and reduced Siglec-7 expression in CHC patients with advanced fibrosis. A healthy individual’s adaptive NK subset is a greater IFNγ producer compared with its FcεRIγ+ conventional NK counterpart, and during chronic HCV infection, IFNγ production from adaptive NK is reduced. Upregulation of PD-1 expression during chronic infection is likely involved in negative regulation of this cell’s function, as evident from lower IFNγ and CD107a expression in PD-1+ compared with the PD-1- subset. Importantly, both the altered phenotype and function of these cells are restored with DAA-mediated virus clearance [29].
Strunz et al. [27] have comprehensively examined the NK cell repertoire using high-dimensional single cell analysis combined with novel metrics that capture the diversity and heterogeneity of these cells. This elegant work shows HCV infection results in increasing NK cell repertoire diversity in an individual, while inter-individual diversity in the repertoire decreases. In contrast with most studies however, no change in expression levels of multiple NK cell markers including CD38, NKp46, NKp30, NKG2D, and Programmed cell death protein 1 (PD-1) was observed during treatment and up to 36 weeks after DAA treatment. Some of the discrepancies described in different studies are potentially related to variables such as demographics of patient populations, regimen of DAA, viral genotype, and different methodologies, among others. For example, HCV genotype [20], the race of CHC patients [88,89], different DAA regimens [20], and non-response to pegylated interferon/ribavirin treatment [19] are all assumed to have an impact on the dynamic changes of NK cells during IFN free therapy.
Based on consensus, it can be concluded that most altered NK cell phenotypes and function observed in a chronically infected patient normalize with rapid clearance of HCV using DAA treatments.
2.2. Mucosal-Associated Invariant T (MAIT) Cells
Mucosal-associated invariant T (MAIT) cells are an innate-like T cell subset that comprise between 5 and 10% of peripheral T cells and about 12–50% of T cells in the liver and the gastrointestinal tract [30,31]. Human MAIT cells have a semi-invariant T cell receptor determining bacterial specificity, high expression of CD161, CD26, and IL-18Rα, and are able to produce cytokines including TNF-α, IFNγ, and IL-17 [30,31]. MAIT cells are readily activated by viral pathogens, including HCV and have a potential role in both host defense and immunopathology. During Chronic HCV infection, the frequency of MAIT cells decreases both in peripheral blood and liver [32,33,34,35]. In fact, MAIT cells were shown to be the most profoundly impacted immune cell type during CHC [32]. Intrahepatic MAIT cells are more activated and cytotoxic compared with their blood counterparts and frequencies of intrahepatic MAIT cells in CHC are inversely correlated to levels of liver inflammation and fibrosis [33]. This reflects the intrahepatic recruitment of MAIT cells in infected patients, where they become activated in the inflammatory cytokine environment and undergo apoptosis. MAIT cells are activated during CHC in a TCR-independent manner dependent on cytokines, predominantly IL-18 that is triggered by viral replication; these activated cells are believed to contribute to liver pathology [35]. The activation status of cells in periphery is reflected in increased expression of CD69, HLA-DR, and PD-1 and lower expression levels of CD127 [32]. Conversely, in vitro-activated MAIT cells have antiviral activity and limit HCV replication by IFNγ induction [35]. Thus, reduced frequencies of MAIT in HCV infection can limit their antiviral effect; at the same time, excess activation and cytokine production can contribute to liver pathology.
Hengst et al. demonstrated that interferon-free DAA therapy does not restore the peripheral MAIT cell compartment either phenotypically or functionally [32]. The frequency of peripheral MAIT cells remained at a low level in treated HCV-infected patients and these residual cells had high activation levels and low functional response. Significantly lower levels of IFNγ and TNF, less pronounced upregulation of CD69 and granzyme B, as well as lower degranulation in response to E. coli persist despite viral clearance. Similar results were reported by others for HCV [33,36] and for HCV/HIV patients treated with DAA [36,37]. The fate of intrahepatic MAIT cells, however, is different from peripheral MAIT cells after virus clearance. DAA treatment elevated the frequency of intrahepatic MAIT cells and resulted in their reduced activation and cytotoxicity [33]. On the other hand, it has been reported that IFNα-based therapies increase activation and frequency of MAIT cells in treated HCV patients [35].
Thus, clearance of chronic HCV infection with DAA therapy fails to recover the frequencies and function of this innate lymphocyte subset even long after cure. This is in contrast with the NK cell recovery that is both rapid and substantial. A consequence of persisting MAIT cell dysfunction is continued impaired bacterial pathogen immune surveillance in cured individuals.
2.3. Gamma Delta (γδ) T Cells
Gamma delta (γδ) T cells are a subgroup of T cells, expressing the TCR with γ and δ chains. Two main subsets of these cells are Vδ1 cells, present in gut and other epithelial tissues and Vδ2/Vγ9, the major γδ subset in the peripheral blood. γδ T cells are abundant in the liver, forming up to 5–15% of hepatic lymphocyte population in humans. In humans, a recent study described the CD27loCD45RAlo subset of Vδ1+ T cells expressing enhanced levels of liver residence-associated markers CD69, CXCR3, and CXCR6 in the liver [90].
Similar to other innate lymphocytes, both protective and pathogenic roles for these lymphoid cells have been suggested in the context of CHC. During HCV infection, a redistribution of γδ T cells to the liver is observed, and these cells express high levels of cytotoxicity markers and produce inflammatory cytokines IL-8, TNFα, and IFNγ [38]. An inflammatory subset of γδ T cells expressing CD161 was also found enriched in livers of patients with chronic HCV infection [39]. Agrati C et al. showed increased concentration of Vδ1 T cells in livers of HCV infected patients and these cells produced IFNγ, levels of which correlated with degree of necroinflammation, further suggesting a role of these cells in hepatic pathology [40]. At the same time, IFNγ produced by Vδ2 T cells resulted in a reduction in HCV replication in vitro in HuH7 infected cells [41]. These contrasting outcomes of role of IFNγ indicate different effects of cytokines produced by different gamma delta T cell subsets in HCV pathogenesis or controls and require further investigations. Even the same subset of γδ T cell can have dual function in protection and pathogenesis. Vδ2 T cells in the circulation of CHC patients are activated and express NK-like cytotoxic molecules including CD56 and CD16, and have high expression of cytolytic molecules, which correlate positively with serum ALT levels. At the same time, these cells had reduced IFNγ-producing capacity, suggesting impaired antiviral function of the Vδ2 T cell in CHC [42]. Intrahepatic Vδ2 T cells from CHC patients had exhausted phenotype and reduced IFNγ response; however, they still maintained antiviral capacity [43].
Not many studies have examined the fate of these cells post HCV cure. We showed that the circulating Vδ2 T cells in patients who were cured of HCV with DAA remain dysfunctional. Patient Vδ2 T cells were activated and had poor proliferative response to phosphoantigens [12] and these defects did not normalize with virus clearance. Another study reported a similar fate of these cells in HIV/HCV co-infected patients [44]. Clonal diversity of γδ T cells during and after HCV infection was examined by Ravens S et al. using mRNA-based next generation sequencing. Both Vγ9+ and Vγ9- cells from patients had comparably complex repertoires than healthy controls and DAA treatment did not alter this at least in 1 year follow up [45].
Impact of HCV clearance on Vδ2/Vγ9 cells can be summarized as persisting dysfunctional phenotype and functional state in a cured patient. An important consequence of remaining defects can manifest in long-term tumor surveillance by this innate effector subset.
3. Adaptive Immune Lymphocytes during and after Clearance of Chronic HCV Infection
The adaptive immune responses are crucial in determining the fate, either acute or chronic, of HCV infection. While about 30% of HCV-infected adult individuals are able to eliminate virus-infected cells via strong and sustained CD8+ and CD4+ T cell responses, the rest develop chronicity. Even though the adaptive immune system does not clear a persistent HCV infection and spontaneous resolutions are rare, immunity exerts some control over infection as evident from exacerbated liver disease in settings of immunosuppressive therapy or co-infection with HIV [91,92,93]. The inability of the immune system to clear a persistent infection is a result of exhaustion and dysfunction of the immune system. Global and antigen-specific T cells are severely dysfunctional in a chronically infected patient, characterized by expression of multiple exhaustion markers and poor HCV-specific cytokine production. With HCV cure now achievable in most treated patients, a significant question that remains is whether the dysregulated immunity is restored upon viral clearance to levels where it protects from re-infection. This is important for determining a need for developing a prophylactic vaccine, especially in high-risk populations. Development of chronicity after reinfection was observed in a chimpanzee model after successful clearance of HCV with DAA [94]. Several cases of reinfection with HCV have been reported in human patients who had previously cleared HCV with DAA therapy [95,96], most with high-risk behaviors. Understanding the adaptive immune restoration in a cured patient will allow drawing of conclusions regarding persisting immune dysfunction as a potential risk factor for re-infection. Additionally, alterations in global T cell populations have potential relevance for overall immune competence and protection against unrelated pathogens.
3.1. Helper T Cells
CD4+ T cells or T helper cells execute diverse tasks after differentiating into lineages such as Th1, Th2, Th17, Tregs, follicular T helper, etc. Although broadly directed HCV-specific CD4+ T cell response is observed at early stages of infection in both acute and chronic patients, persistent viremia is associated with early proliferative defects of these cells and subsequent rapid depletion of virus-specific response [46]. Difficulty in detecting antigen-specific T cells in peripheral blood from chronically infected patients is due both to the small frequency of these cells and possible migration of cells to the site of virus replication in the liver. Recent studies have investigated class II tetramer binding CD4+ T cells in chronic HCV infection and their fate after virus clearance. In an elegant study with samples from 44 patients tested at baseline, during and after DAA-mediated cure, Smits et al. [47] showed a significant increase in HCV tetramer binding CD4+ T cells within 2 weeks of therapy initiation. This likely reflects the egress of virus-specific cells from the liver in an IP-10 gradient-dependent mechanism. Analyses of global CD4+ T cells revealed that frequency of T cells expressing CXCR3, which is expressed on the vast majority of liver-infiltrating CD4+ T cells in chronic HCV infection [48], is increased in the peripheral blood 1 to 2 weeks after DAA therapy initiation, suggesting an early emigration of liver-infiltrating lymphocytes from liver to blood [49]. This is accompanied by a rapid decline in levels of serum IP-10 (CXCL10, the ligand for CXCR3) [50].
Concomitantly, there are dynamic changes in exhaustion and activation levels of these cells that tightly control their differentiation, proliferation, and effector function. T cell exhaustion is characterized by sustained expression of multiple inhibitory receptors on T cells, including PD-1, 2B4, Cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), CD160, T cell immunoglobulin and mucin domain-containing protein 3 (TIM-3), and Killer Cell Lectin Like Receptor G1 (KLRG1) [51,52,53,69,70]. Expression of inhibitory receptors, PD-1 and CTLA-4, on global CD4+ T cells is almost universal in the early phase of the infection, regardless of the outcome. With spontaneous resolution of the infection, expression of these inhibitory receptors fades away. In contrast, persisting viremia continues to drive T cell activation, PD-1 and CTLA-4 expression, and blocks T cell differentiation until the virus-specific cells disappear from the circulation [52].
The impact of virus clearance on these exhaustion and activation markers on CD4+ T cells is well documented. The frequency of virus-specific CD4+ T cells expressing PD-1, BTLA, and T cell Ig and ITIM domain (TGIT) stays high during and after treatment, although intensity of PD-1 expression is reduced significantly [47]. Higher levels of TIGIT expression on HCV-specific CD4+ T cells observed during CHC remains persistently elevated at least until 24 weeks from the end of DAA treatment [53]. There is a however normalization of activation status of antigen-specific CD4+ T cells, as indicated by the significant reduction in CD38, ICOS, and OX40 expression [47].
Interestingly, an increase in expression of memory-associated markers CD127 and TCF-1 on antigen-specific cells occurs with viral clearance, this however is not accompanied by changes in cytokine expression patterns of these cells [47]. During CHC, a redistribution of memory CD4+ T cells at the expenses of naïve cells occurs, with memory T lymphocytes displaying an activated and exhaustive phenotype; these changes are only partially restored with virus clearance [54].
With respect to the global CD4+ T cell population, frequency of peripheral CD4+ T cells increases significantly with virus clearance [49,55] and a re-differentiation of the T lymphocyte memory compartment results in a higher effector memory population with increased Tbet expression [55]. The expression of various exhaustion markers on global CD4+ T cells is not uniformly impacted by viral clearance. While no change in PD-1 expression on CD4+ T cells with virus clearance occurs, expression of TIM-3 reduces on several innate and adaptive cells including CD4+ T, NK, and NK-like T cells [56]. There is no consensus regarding the expressions of activation markers HLA-DR and CD38 on the CD4+ T cells with HCV clearance. Both a decreased expression of HLA-DR and CD38 post DAA therapy [13,49] and no significant change in their expressions at 12 weeks after end of therapy [57], is reported by different investigators. Functionally, an increase in IFNγ, IL-17, and IL-22 producing global CD4+ and CD8+ T cells is observed after DAA therapy [57].
Although DAA treatments successfully clear the virus, the observations of re-infection in cured patients warrant investigations into prophylactic vaccination strategies. Adenoviral-vectored vaccines encoding non-structural proteins of HCV have shown multi-specific, high-magnitude, and durable CD4+ and CD8+ T cell responses in healthy volunteers; these responses resemble “protective memory” detected in patients who undergo spontaneous HCV resolution [97,98,99]. However, similar robust CD4+ T cell response is not present in immunized CHC patients, including in those treated with interferon/ribavirin [98,99] or DAA therapy [97]. Thus, viral clearance likely does not promote resurrection of exhausted memory CD4+ T cells, resulting in poor CD4+ T cell response with these vaccines. Unlike CD4+ T cells, vaccine-induced CD8+ T cell response was readily detectable in the majority of CHC patients treated with interferon (IFN)-based therapy [98] or in chimpanzees treated with DAA [94], to suppress viral load during vaccination. In spite of detectable CD8+ T cell response, these vaccination strategies failed to control HCV replication due to sequence variation of circulating virus, lack of help from CD4+ T cells, and expression of inhibitory receptors, particularly on virus-specific intrahepatic CD8+ T cells (discussed in details in [55]) [100].
Interestingly, several studies have indicated that many persistently HCV-infected pregnant women achieve substantial decline in HCV viral load after childbirth, which is associated with re-appearance of HCV-specific CD4+ T cells with an effector-memory phenotype at 3 months postpartum [101,102,103]. In addition, significantly increased frequencies of IL-2+, IFNγ+, and IL-2+IFNγ+ HCV-specific CD4+ T cells are detected at 3 months postpartum compared to the third trimester of pregnancy. However, PD-1 expression on HCV-specific CD4+ T cells remained high and correlated with viral control, which is indicative of its role in CD4+ T cell activation rather than exhaustion in these cases. In contrast, reduced CTLA-4 expression was associated with higher viral control, indicating its role as a negative regulator of HCV-specific CD4+ T cell activity in the postpartum period [103]. Such studies indicate that revival of HCV-specific CD4+ T cell immunity is possible after persistent infection and it can play a crucial role in controlling HCV replication.
PD-1 is one of the most well studied negative regulators of T cell response and its high expression level is an indicator of an exhausted immune response. We showed that in a cohort of patients that were treated for an ultra-short duration of 4 weeks with DAA, those that achieved SVR had higher levels of PD-1 and other “inhibitory receptor” co-expression on their CD4+ and CD8+ T cells before and at the end of treatment, although the levels of these markers significantly decreased 12 weeks after the end of treatment in the only SVR group [11]. Since these markers are expressed in response to continuous antigenic stimulation, it makes sense that their expression is high on virus-specific cells. We believe that the T cells co-expressing multiple inhibitory markers reflect antigen-specific cells and once the viral antigen level was reduced with 4 weeks of treatment, patients who had higher levels of such antigen-specific cells continued to control virus replication even when treatment was stopped at 4 weeks. This suggests that concentration of these markers reflects an activation of T cells in response to antigenic stimulation and not necessarily a terminally exhausted phenotype, which cannot be restored.
In summary, DAA therapy partially improves the CD4+ T cell compartment after HCV clearance. The frequencies of global CD4+ T cells are increased after DAA therapy and virus-specific CD4+ T cells transiently increase in circulation, possibly due to emergence from intrahepatic compartment. Expression of activation markers on antigen-specific CD4+ T cells is significantly reduced after virus clearance but expression of inhibitory receptors is reduced only moderately with modest improvement in cytokine production.
3.2. Follicular Helper T (Tfh) Cells
Humoral immune response requires interaction between antigen-specific B cells and CD4+ helper T cells. CD4+CXCR5+ follicular helper T cells (Tfh) cells are a specialized subset of CD4+ T cell that are essential for germinal center formation and production of high-affinity antibody by B cells in lymphoid follicles [104,105]. These cells express BCL-6 transcription factor after being primed and activated by dendritic cells and in turn, exhibit a unique CXCR5hiCCR7loPD-1+ICOS+ phenotype [106,107,108,109]. Tfh cells promote proliferation, differentiation, and maturation of antigen-specific B cells inside lymphoid follicles through various cytokines, including the signature cytokine IL-21 [104,110,111].
Unlike acute HCV infection, where circulating virus-specific Tfh cells can be detected in periphery, these cells disappear from peripheral blood but remain detectable in the liver in CHC patients [48]. Comparing CHC and healthy donors, Spaan M et al. found no difference in the frequency of circulating CD4+CXCR5+ CXCR5+, CXCR5+PD-1+, CXCR5+ICOS+, and CXCR5+PD-1+ICOS+ subsets of CD4+ T cells between the groups [58]. In CHC patients with Cryoglobulinemia Vasculitis however, frequency of Tfh cells is higher compared to healthy donors and a reduction in Tfh frequency occurs after DAA therapy [59]. Although circulating Tfh cells in CHC patients express and secrete less IL-21 compared to healthy subjects, they are capable of stimulating memory B cells into IgG and IgM-producing plasmablasts [58,60]. Smits et al. demonstrated that the frequency of HCV-specific CD4+ T cells increased in circulation within two weeks of the initiation of DAA therapy [47] and the majority of these HCV-specific CD4+ T cells exhibited phenotypic and transcriptional characteristics of follicular T helper cells; these are maintained after therapy-induced elimination of persistent HCV infection. Following viral clearance with DAA, a downregulation in expression of exhaustion (PD1, BTLA, CD39, and TIGIT) and activation (CD38, ICOS, and OX40) markers and an upregulation in memory-associated markers (CD127) are detected on Tfh cells [47]. Thus, the antigen-specific Tfh reappear in circulation in a cured patient, and are maintained for months thereafter. Moreover, this reshaping of circulating HCV-specific Tfh is accompanied by reduced germinal center activity and preceded reduction on HCV-specific neutralizing antibody titers [47]. Whether these antigen-specific Tfh can induce long lasting protective antibody response will be an important consideration of HCV vaccine efforts.
A special subset of follicular CD4+ T cells, the T follicular regulatory (Tfr) cells, expresses regulatory markers CD25 and FoxP3 and functions to suppress Tfh cells, thus negatively regulating the germinal center [112,113,114,115]. The frequency of Tfr cells is increased both in the circulation and liver of a chronically infected patient [116,117]. This increase in intrahepatic Tfr cells could be a consequence of the immune modulatory effect exerted by HCV. Cobb, J.A. et al. demonstrated that HCV-infected hepatocyte-derived exosomes lead to an expansion of Tfr cells in a TGFβ-dependent manner [117]. However, it is not clear whether these Tfr cells are HCV-specific or how much role they play in suppression of HCV-specific Tfh. Virus clearance results in a reduction and normalization of their numbers with the concomitant increase in frequencies of Tfh cells [118].
3.3. T Regulatory Cells (Treg)
T regulatory cells (Treg) are CD4+ or CD8+ T cells engaged in promoting self-tolerance and suppressing non-self immune responses. Thus, they can play beneficial (prevent excess inflammation of the liver) or harmful (suppress anti-HCV immune responses) roles in the setting of chronic HCV infection. CD4+ Tregs, defined as CD4+CD25+CD127lowFoxp3+ cells, are a specialized T cell population that inhibit the activation, proliferation, differentiation, and effector functions of multiple immune cell types, including T cells, B cells, NK cells, and dendritic cells [119,120,121,122]. Once activated, they exert their suppressive function in a non-specific manner through cell-to-cell contact inhibition and production of regulatory cytokines TGFβ, IL10, and IL35 [123]. In CHC, CD4+ Treg frequency gradually expands, resulting in reduced overall antiviral immune responses. A mechanism for increased Treg numbers during CHC is altered differentiation of CD4+ T cells in a persistently infected patient. While chronically infected patients often present with higher HCV-specific CD4+ T cell responses, both in terms of breadth and magnitude, this is largely comprised of regulatory T cells (CD4+CD25highCD134+CD39+). In comparison, HCV-specific CD4+ T cell responses in subjects who cleared the virus are comprised of effector T cells (CD4+CD25highCD134+CD39−) phenotype producing higher levels of IFNγ [61]. Galectin-9 (Gal-9), the ligand for TIM-3, is upregulated by Kupffer cells and monocytes and induces the expansion of CD4+ regulatory T cells (Treg) in a TGF-β-dependent manner in HCV-infected patients [62,63].
Abundant intrahepatic infiltration of Treg cells is present in CHC patients compared to healthy individuals [64,65], due to migration of these cells to liver. Monocyte-derived dendritic cells (mDCs) possess the capacity to generate Treg from naive CD4+ T cells in CHC patients and their interaction with HCV-infected hepatocytes results in increased production of the Treg-attracting chemokines CCL17 and CCL22, which facilitate intrahepatic recruitment of Tregs [124,125]. Whether Tregs limit or promote liver fibrosis is less clear [64,126,127]. Claassen et al. showed that activated Tregs migrate to the livers of CHC patients and limit the extent of HCV-induced immune activation, thereby limiting the extent of fibrosis [128]. Increased numbers of TGFβ producing HCV-specific Treg cells in chronic HCV-infected livers with limited fibrosis and normal serum ALT levels compared with CHC patients with elevated ALT levels support the role of this T cell in controlling liver pathology [64,129]. Langhans et al. however described a minor subset of intrahepatic IL-8 producing Tregs, present in close proximity to the areas of liver fibrosis, whose numbers correlated with stage of fibrosis; this subset also upregulated profibrogenic markers TIMP1, MMP2, TGF-beta1, alpha-SMA, collagen, and on hepatic stellate cells [126].
HCV clearance with DAA treatment does not affect percentages of Tregs in peripheral blood, even long-term after virus elimination [66,67,68]. Although the frequency of Tregs during DAA therapy gradually decreases until the end of therapy, in parallel with reduction in IL10 and TGFβ production, these changes are not sustained during the follow-up period, in spite of patient achieving SVR [67]. In spite of this, the Treg cells that reappear and expand after the end of DAA therapy might be functionally different from the Tregs present before therapy. Wei K et al. recently demonstrated that stimulation with Pam3Csk4, a TLR2 agonist, enhances the suppressive function and IL-35 production by T-regs that were isolated from a CHC patient before DAA therapy but not from Tregs isolated at SVR time-point. This indicates that therapy results in altered responsiveness of Tregs to TLR2 stimulation [66]. The fibrosis-limiting Tregs are retained in the liver up to at least 1 year after DAA-mediated cure [128], while the status of IL-8 producing fibrosis-promoting Tregs in the absence of HCV infection is unknown.
CD8 Tregs, defined as CD8+CD25+FoxP3+ cells, are a more recent discovery and are present at very low frequency in blood, making their characterization more difficult. One study has shown elevated frequency of CD8 Tregs both in the liver and peripheral blood of chronic patients, and this is associated with liver inflammation but not with fibrosis [130]. The status of this subset after HCV cure is not known.
The presence of Tregs in circulation long after the end of DAA therapy can impact the success of prophylactic vaccine candidates that are intended to prevent re-infection in cured individuals. In CHC patients, Han JW et al. evaluated the immunogenicity of an IFNL3-adjuvanted vaccine. Recombinant human IFNL3 reduced Treg cell frequency by inducing apoptotic cell death particularly of the activated Treg subpopulation (CD45RA−CD4+FoxP3hi); this was accompanied by an increase in virus-specific T cell responses [131]. Thus, innovative vaccination strategies that take into account the altered lymphocyte landscape in a cured patient might be critical for their success.
3.4. Th17 Cells
The Th17 subset of CD4+ T cells is distinguished by production of signature cytokines IL-17 and IL-22 and these cells play a key role in host resistance to microbial infections and to development of autoimmune diseases [132,133,134,135]. The transcription factors, orphan nuclear receptors ROR gamma t and ROR alpha (RORγt and RORα), determine the differentiation of CD4+ T cell to Th17 lineage with the influence of IL-6 and TGF-β [136,137,138]. Th17 are not well characterized in HCV infection. It is possible that these cells may have role in HCV-associated liver pathology as increased numbers of these cells correlate with levels of liver inflammation [139]. The Th17/IL-17 axis is believed to play a pro-fibrogenic role by way of its involvement in the production of cytokine IL-6 and profibrotic factors like TGFβ [140]. A skewing of intrahepatic T cells towards a higher Th17/FoxP3 ratio associates with advanced liver fibrosis in CHC and this is determined by TGFβ and TNFα concentrations in liver microenvironment [141]. On the other hand, IL-21 produced by Th17 cells can limit exhaustion of CD4 and CD8 T cells [142]. Different roles of this cell type in the context of both impact on antiviral effector function and in liver pathology have been explored. One report has examined the fate of these cells in DAA-cured patients so far. Comarmond C et al. showed increased frequencies of these cells in HCV patients with cryoglobulinemia vasculitis (CV) but not in those without CV. This Th17 polarization was normalized by end of treatment with DAA [59]. Thus, much remains to be known regarding their fate and significance in a cured patient.
3.5. Cytotoxic T Lymphocytes
Failure to eliminate HCV from infected hepatocytes is largely attributed to functional impairment of virus-specific CD8+ T cells, which is characterized by limited proliferative capacity, defective cytokine production and reduced cytolytic activity of these effector cells [71,72,73]. Two widely accepted mechanisms for the impairment of virus-specific CD8 T cells are the emergence of escape mutant variants of the wild type virus and CD8 T cell exhaustion [46,69,72,74,75]. Expression of Interleukin-7 (IL-7) receptor alpha chain (CD127), a key molecule associated with the maintenance of memory T cell populations, on CD8+ T cells is typically only observed when the respective antigen is controlled [143,144]. Cytokines, such as IL-7, and signaling through CD127 are critical for long-lived memory cells to survive and proliferate in the absence of antigen [145]. Whilst most resolved HCV patients express a high level of CD127 on more than 90% of the virus-specific CD8+ T cells [74], a substantial proportion (ranging from 10 to 100%) of the virus-specific CD8+ T cells express CD127 in long-established chronic HCV infection [74,76]. The high expression of CD127 in the virus-specific CD8 T cells in chronic infection is attributed to the emergence of escape mutant variant with mutation in the CD8+ T cell targeting epitopes due to immune selection pressure. In the absence of the targeting epitope, the effector CD8+ T cells re-differentiate into CD127 expressing memory-like CD8 T cells, while the HCV infection persists with mutant variants of viral antigens [74]. On the other hand, CD8+ T cells in CHC patients express a low level of CD127, generally accompanied by a higher level of activation marker CD38 and a lower level of CCR7 expression, insinuating re-activation-induced downregulation of CD127 on them [76]. The common feature observed in several chronic viral infections is exhaustion of T cells; persistent antigenic exposure of the virus-specific CD8 T cells to viral antigens leads to severe CD8 T cell exhaustion in chronic HCV-infected patients. Expression of multiple co-regulatory molecules such as PD-1, 2B4, TIM-3, Lag-3, CD5, CD160, and TIGIT is upregulated on HCV-specific CD8 T cells during chronic HCV infection [11,70], and is associated with poor proliferative and functional response. Varying levels of restoration of proliferation capacity of these virus-specific CD8+ T cells are observed following cessation of antigenic stimulation after DAA therapy-mediated viral clearance [77,78]. Factors that impact the proliferation capacity of the HCV-specific CD8 T cells upon peptide stimulation include the presence of cells recognizing the conserved epitopes [77], gender, presence of advance fibrosis [78], and combination of drugs used in the DAA regimen. Aregay A et al. showed a significant decrease in activation markers, CD38 and HLA-DR, on HCV-specific CD8+ T cells after DAA-mediated clearance of HCV and no significant reduction in the expression exhaustion markers, PD-1, TIM3, LAG3, and CD5 [78]. Likewise, the cytokine production from virus-specific CD8+ T cells was not restored after DAA therapy. However, a significant restoration of the proliferation capacity of these cells occurred in a subset of HCV patients that were treated with a combination of DAA drugs that included protease inhibitors but not sofosbuvir [78].
Whether expression of inhibitory molecules is indicative of persisting dysfunctional immunity in a cured individual is something not well understood. Chimpanzee studies showed that PD-1 is expressed on functional memory CD8+ T cells after resolution of infection and thus, is not always a marker of exhaustion [146]. We recently found that higher-level co-expression of PD-1 with other inhibitory receptors including CD160, 2B4, CTLA-4, Blimp-1, or Tim-3 on CD8+ T cells is present on HCV tetramer-positive cells and high expression of these molecules both at treatment start and at end of treatment time points correlated with the patients’ ability to clear chronic infection when treated with an ultra-short regimen involving 4 weeks of DAA treatment [11]. Importantly, expression of these molecules went down significantly in those who achieved SVR at week 16 but not in those that eventually relapsed. It is known from other viral infections that antigen-specific CD8+ T cells express PD-1, which preserves them from overstimulation, excessive proliferation, and terminal differentiation [147]. Thus, maintenance of PD-1 on T cells after HCV clearance potentially has a beneficial role in the preservation of these cells.
Recently, the role of high mobility group box (TOX) transcription factor as an important transcriptional regulator of exhausted CD8+ T cells in chronic viral infection was described [79,80]. Alfei et al. showed that TOX is highly expressed in HCV antigen-specific CD8+ T cells in CHC patients but not in those who spontaneously resolved HCV or in influenza-specific CD8+ T cells. Moreover, high expression of TOX in antigen-specific CD8+ T cells correlates with high PD1 expression in CHC patients [80]. In fact, TOX transcriptionally increases PD1 expression on antigen-specific CD8+ T cells in chronic infections, resulting in their reduced polyfunctionality [80]. After HCV clearance with DAA therapy, TOX expression is reduced in HCV-specific CD8+ T cells with concomitant reduction in PD1 expression [80]. Considering these data, targeting TOX could be a promising immunotherapeutic endeavor to reverse CD8+ T cell exhaustion and their disrupted polyfunctionality.
In an elegant work, Wolski et al. demonstrated differences in the transcriptional landscape of HCV-specific CD8+ T cells from patients during early acute HCV infection that either developed persistent infection or spontaneously resolved their infection [148]. This study identified molecular signatures of genes that govern the fate of virus-specific T cells towards memory or exhaustive phenotype; these signatures involved metabolic, nucleosomal, and immune processes. Among the differences, those who spontaneously resolved the infection showed higher levels expression of genes related to T cell differentiation and memory, while those who developed chronic infection had higher expression levels of genes related to T cell function, proliferation, and survival. Metabolic dysregulations at the transcriptional level involved genes related to nucleosomal regulation and those linked to T cell differentiation and the inflammatory environment. How clearance of HCV in these chronically infected subjects will reprogram these transcriptional signatures remains to be seen.
An emerging area in the field of T cell function is bioenergetic requirements of cells during a chronic viral infection [149]. Dynamic reprogramming of the metabolism occurs to meet excess requirements of T cell response under viral stress. Importantly, PD-1 along with mTOR signaling regulates these mitochondrial dynamics by repressing transcriptional coactivator PGC1a, leading to reduced cellular respiration, glycolysis, and dysregulated mitochondrial energetics [149]. Mitochondrial plasticity and bioenergetics are modulated significantly in chronic HBV and HCV infections [78,150]. Polarization status, superoxide level, and mass of the mitochondria dynamically influence survival, activation, and function of T lymphocytes [149,151,152,153,154]. Aregay A et al. elucidated the presence of mitochondrial dysfunction, characterized by depolarized mitochondria with increased mass and higher mitochondrial ROS in HCV-specific CD8 T cells, in CHC patients; these changes did not normalize following DAA-mediated clearance of HCV [78]. It remains to be investigated whether mitochondrial dysfunction is an effect of chronic infection or if it is a causal factor in impaired cellular function, exhaustion, and eventually, failure of antiviral T cells to control virus replication.
To summarize, DAA-mediated clearance of chronic HCV infection has partial impact on the recovery of CD8+ T cell dysfunction. While reduction in activation status and a partial reduction in the expression levels of exhaustion markers, specifically on antigen-specific cells, are evident, cytokine production, proliferation capacity and mitochondrial function of the CD8+ T cells remains dysfunctional.
3.6. B Cells
The role of B cells in HCV infection is not well understood. Neutralizing antibodies against the virus are present in all infected patients irrespective of outcome and it is not known if they play a significant role in virus control. Despite this, perturbations in B cell phenotypes are observed in chronically infected patients. The most common observation with respect to abnormality in B cell compartment is the presence of circulating mixed cryoglobulins in about 50% of patients, with overt cryoglobulinemia vasculitis (CV) in up to 10% [59]; these patients also have an accumulation of atypical memory (AtM) B cells (CD21lo/- IgM+). In cohorts of patients with mixed CV that were treated with DAA, a decrease in percentages of these autoreactive B cells is reported, suggesting some recovery in B cell with virus clearance [59]. An extensive analysis of the mechanism behind Tbet+CD11c+CD27+CD21− B cell-mediated autoimmunity showed the AtM derived antibodies do not react against ubiquitous autoantigens or HCV antigens, including NS3 and E2 proteins. These AtM-derived antibodies possess rheumatoid factor activity and target unique epitopes on the human IgG–Fc region. Virus clearance with DAA results in elimination of these deleterious B cells [155].
4. Liver Resident Lymphocytes in Chronic HCV Infection
While peripheral blood lymphocytes are recruited to tissues under conditions of viral infection, there are specialized populations of lymphocytes that reside in tissues that do not recirculate in the blood. These are called tissue resident cells and their role in chronic infections is being revealed now. Most peripheral lymphocytes have liver resident counterparts—memory CD8+ T (TRM) cells, invariant natural killer T (iNKT) cells, mucosal-associated invariant T (MAIT) cells, γδT cells, and innate lymphoid cells (ILCs) such as natural killer (NK) cells and other ILCs. These cells express phenotypic markers CD44, CD103, and CD49a, which mediate the adhesion and retention of these cells and possess a “memory-like” phenotype and function to provide long-lasting and robust protection against subsequent infection. For example, virus-specific CD8 TRM can reside in the liver for a long time after primary infection and can control secondary infection with their cytolytic function [156]. On the other hand, these cells participate in liver pathology as well. Not much has been revealed about these cells during CHC yet. IL-2highT-betloEomesloBlimp-1hiHobitlo T cells were described enriched in livers from chronic HBV-infected patients who had partial immune control of the virus; these cells are believed to provide local surveillance [157]. The proportions of CD8+ T cells expressing CD103 was higher in CHC patients compared with healthy controls and these cells have an altered differentiation program indicated by lower levels of Hobit expression [158]. Further studies are required to understand the role of resident lymphocytes in HCV pathogenesis or control as well as their fate with and after chronic infection.
5. Summary
The human immune system efficiently controls most of the viral infections through strong sterilizing immunity mediated by various innate (non-specific; e.g., NK cells, gamma-delta T cells, MAIT cells) and adaptive (virus-specific; e.g., CD4+ Th1, Tfh, Th17 cells, CD8+ cytotoxic T cells, B cells) immune cell types. However, certain viruses including HCV can evade the immune system and develop chronic infection in some individuals. For development of chronicity, two fundamental events need to occur; first, the virus must evade the host sterilizing immunity and second, the host immune system must adjust to the continuous presence of viral antigen-driven inflammatory responses in order to limit viral replication to an acceptable level without permanent damage to infected tissues [159]. This adaptation comes in the form of transcriptional, metabolic, and epigenetic changes in immune cells. Whether these changes are permanent or reversible once the antigen is removed is a significant question in immunology, which can now be addressed in CHC patients that are cured with highly effective DAA treatments.
In this review, we discussed phenotypic and functional changes that occur in various lymphocyte populations during chronic HCV infection and their recovery upon clearance of HCV with DAA therapy. In our observation, NK cells, MAIT cells, CD4+ Th1 cells, and CD8+ T cells have been studied relatively extensively in comparison with other lymphocyte populations like γδ T cells, Th17 cells, Tfh cells, T regs, and B cells in the context of DAA-mediated clearance of HCV. In particular, studies on B cells and Tfh cells are scarce. Except NK cells, no other lymphocyte population undergoes significant recovery in up to one year follow-up studies after DAA therapy. A partial recovery manifested in reduced levels of activation and exhaustion markers and improved antigen-specific functional response is observed for CD4+ and CD8+ T lymphocytes. No significant recovery occurs in MAIT and γδ T cells. However, it is barely a decade since the implementation of these treatments and longer-term follow-up studies will provide more clear understanding of the reversibility of these immune defects.
6. Final Remarks
HCV clearance with DAA treatments has thus provided scientists with a first of its kind opportunity to investigate the extent of immune restoration that is possible once the antigen responsible for persistent immune dysfunction has been removed. Viral clearance has differential impact on different lymphocyte subsets, but overall viral clearance does not result in a “normalization” of lymphocyte landscape. Further studies can examine whether certain immune alterations take more time to normalize after cure or whether they are epigenetically imprinted by viral infection. The foremost significance of this is for a cured individual’s future immune competence, both in the context of re-exposure to HCV and for risk of associated pathology-like fibrosis and HCC.
Funding
This research was funded by National Institutes of Health: 5R01DA043396-05.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Holmes, J.A.; Rutledge, S.M.; Chung, R.T. Direct-acting antiviral treatment for hepatitis C. Lancet 2019, 393, 1392–1394. [Google Scholar] [CrossRef]
- Osinusi, A.; Meissner, E.G.; Lee, Y.J.; Bon, D.; Heytens, L.; Nelson, A.; Sneller, M.; Kohli, A.; Barrett, L.; Proschan, M.; et al. Sofosbuvir and ribavirin for hepatitis C genotype 1 in patients with unfavorable treatment characteristics: A randomized clinical trial. JAMA 2013, 310, 804–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osinusi, A.; Kohli, A.; Marti, M.M.; Nelson, A.; Zhang, X.; Meissner, E.G.; Silk, R.; Townsend, K.; Pang, P.S.; Subramanian, G.M.; et al. Re-treatment of chronic hepatitis C virus genotype 1 infection after relapse: An open-label pilot study. Ann. Intern. Med. 2014, 161, 634–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, A.; Osinusi, A.; Sims, Z.; Nelson, A.; Meissner, E.G.; Barrett, L.L.; Bon, D.; Marti, M.M.; Silk, R.; Kotb, C.; et al. Virological response after 6 week triple-drug regimens for hepatitis C: A proof-of-concept phase 2A cohort study. Lancet 2015, 385, 1107–1113. [Google Scholar] [CrossRef] [Green Version]
- Osinusi, A.; Townsend, K.; Kohli, A.; Nelson, A.; Seamon, C.; Meissner, E.G.; Bon, D.; Silk, R.; Gross, C.; Price, A.; et al. Virologic response following combined ledipasvir and sofosbuvir administration in patients with HCV genotype 1 and HIV co-infection. JAMA 2015, 313, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Kohli, A.; Kapoor, R.; Sims, Z.; Nelson, A.; Sidharthan, S.; Lam, B.; Silk, R.; Kotb, C.; Gross, C.; Teferi, G.; et al. Ledipasvir and sofosbuvir for hepatitis C genotype 4: A proof-of-concept, single-centre, open-label phase 2a cohort study. Lancet Infect. Dis. 2015, 15, 1049–1054. [Google Scholar] [CrossRef] [Green Version]
- Kattakuzhy, S.; Wilson, E.; Sidharthan, S.; Sims, Z.; McLaughlin, M.; Price, A.; Silk, R.; Gross, C.; Akoth, E.; McManus, M.; et al. Moderate Sustained Virologic Response Rates With 6-Week Combination Directly Acting Anti-Hepatitis C Virus Therapy in Patients With Advanced Liver Disease. Clin. Infect. Dis. 2016, 62, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, E.S.; Silk, R.; Mathur, P.; Gross, C.; Eyasu, R.; Nussdorf, L.; Hill, K.; Brokus, C.; D’Amore, A.; Sidique, N.; et al. Concurrent Initiation of Hepatitis C and Opioid Use Disorder Treatment in People Who Inject Drugs. Clin. Infect. Dis. 2020, 10.1093/cid/ciaa105. [Google Scholar] [CrossRef]
- Shrivastava, S.; Wilson, E.; Poonia, B.; Tang, L.; Osinusi, A.; Kohli, A.; Kottilil, S. Augmentation of hepatitis C virus-specific immunity and sustained virologic response. J. Viral. Hepat. 2017, 24, 742–749. [Google Scholar] [CrossRef]
- Shrivastava, S.; Bhatta, M.; Ward, H.; Romani, S.; Lee, R.; Rosenthal, E.; Osinusi, A.; Kohli, A.; Masur, H.; Kottilil, S.; et al. Multitarget Direct-Acting Antiviral Therapy Is Associated With Superior Immunologic Recovery in Patients Coinfected With Human Immunodeficiency Virus and Hepatitis C Virus. Hepatol. Commun. 2018, 2, 1451–1466. [Google Scholar] [CrossRef]
- Romani, S.; Stafford, K.; Nelson, A.; Bagchi, S.; Kottilil, S.; Poonia, B. Peripheral PD-1(+) T Cells Co-expressing Inhibitory Receptors Predict SVR With Ultra Short Duration DAA Therapy in HCV Infection. Front. Immunol. 2019, 10, 1470. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Mondal, R.K.; Romani, S.; Bagchi, S.; Cairo, C.; Pauza, C.D.; Kottilil, S.; Poonia, B. Persistent gamma delta T-cell dysfunction in chronic HCV infection despite direct-acting antiviral therapy induced cure. J. Viral. Hepat. 2019, 26, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, B.; El-Kamary, S.S.; Magder, L.S.; Stafford, K.A.; Charurat, M.E.; Poonia, B.; Chairez, C.; McLaughlin, M.; Hadigan, C.; Masur, H.; et al. Immunological recovery in T-cell activation after sustained virologic response among HIV positive and HIV negative chronic Hepatitis C patients. Hepatol. Int. 2019, 13, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Mathur, P.; Emmanuel, B.; Sneller, M.; Zhang, X.; Poonia, B.; Kottilil, S. Recovery of hepatitis C specific T-cell responses after rituximab therapy in hepatitis C mixed cryoglobulinemic vasculitis. J. Med. Virol. 2018, 90, 936–941. [Google Scholar] [CrossRef] [Green Version]
- Mondelli, M.U.; Varchetta, S.; Oliviero, B. Natural killer cells in viral hepatitis: Facts and controversies. Eur. J. Clin. Investig. 2010, 40, 851–863. [Google Scholar] [CrossRef]
- Kokordelis, P.; Kramer, B.; Korner, C.; Boesecke, C.; Voigt, E.; Ingiliz, P.; Glassner, A.; Eisenhardt, M.; Wolter, F.; Kaczmarek, D.; et al. An effective interferon-gamma-mediated inhibition of hepatitis C virus replication by natural killer cells is associated with spontaneous clearance of acute hepatitis C in human immunodeficiency virus-positive patients. Hepatology 2014, 59, 814–827. [Google Scholar] [CrossRef]
- Oliviero, B.; Varchetta, S.; Paudice, E.; Michelone, G.; Zaramella, M.; Mavilio, D.; De Filippi, F.; Bruno, S.; Mondelli, M.U. Natural killer cell functional dichotomy in chronic hepatitis B and chronic hepatitis C virus infections. Gastroenterology 2009, 137, 1151–1160. [Google Scholar] [CrossRef]
- Spaan, M.; van Oord, G.; Kreefft, K.; Hou, J.; Hansen, B.E.; Janssen, H.L.; de Knegt, R.J.; Boonstra, A. Immunological Analysis During Interferon-Free Therapy for Chronic Hepatitis C Virus Infection Reveals Modulation of the Natural Killer Cell Compartment. J. Infect. Dis. 2016, 213, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.X.; Luo, B.F.; Jiang, H.J.; Cong, X.; Jin, Q.; Ma, D.L.; Wei, L.; Feng, B. Recovery of natural killer cells is mainly in post-treatment period in chronic hepatitis C patients treated with sofosbuvir plus ledipasvir. World J. Gastroenterol. 2018, 24, 4554–4564. [Google Scholar] [CrossRef]
- Ning, G.; Li, Y.T.; Chen, Y.M.; Zhang, Y.; Zeng, Y.F.; Lin, C.S. Dynamic Changes of the Frequency of Classic and Inflammatory Monocytes Subsets and Natural Killer Cells in Chronic Hepatitis C Patients Treated by Direct-Acting Antiviral Agents. Can. J. Gastroenterol. Hepatol. 2017, 2017, 3612403. [Google Scholar] [CrossRef]
- Ahlenstiel, G.; Titerence, R.H.; Koh, C.; Edlich, B.; Feld, J.J.; Rotman, Y.; Ghany, M.G.; Hoofnagle, J.H.; Liang, T.J.; Heller, T.; et al. Natural killer cells are polarized toward cytotoxicity in chronic hepatitis C in an interferon-alfa-dependent manner. Gastroenterology 2010, 138, 325–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serti, E.; Chepa-Lotrea, X.; Kim, Y.J.; Keane, M.; Fryzek, N.; Liang, T.J.; Ghany, M.; Rehermann, B. Successful Interferon-Free Therapy of Chronic Hepatitis C Virus Infection Normalizes Natural Killer Cell Function. Gastroenterology 2015, 149, 190–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozzano, F.; Picciotto, A.; Costa, P.; Marras, F.; Fazio, V.; Hirsch, I.; Olive, D.; Moretta, L.; De Maria, A. Activating NK cell receptor expression/function (NKp30, NKp46, DNAM-1) during chronic viraemic HCV infection is associated with the outcome of combined treatment. Eur. J. Immunol. 2011, 41, 2905–2914. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-j.; Wang, X.-x.; Luo, B.-f.; Cong, X.; Jin, Q.; Qin, H.; Zhang, H.-y.; Kong, X.-s.; Wei, L.; Feng, B. Direct antiviral agents upregulate natural killer cell potential activity in chronic hepatitis C patients. Clin. Exp. Med. 2019, 19, 299–308. [Google Scholar] [CrossRef]
- De Maria, A.; Fogli, M.; Mazza, S.; Basso, M.; Picciotto, A.; Costa, P.; Congia, S.; Mingari, M.C.; Moretta, L. Increased natural cytotoxicity receptor expression and relevant IL-10 production in NK cells from chronically infected viremic HCV patients. Eur. J. Immunol. 2007, 37, 445–455. [Google Scholar] [CrossRef]
- Nattermann, J.; Feldmann, G.; Ahlenstiel, G.; Langhans, B.; Sauerbruch, T.; Spengler, U. Surface expression and cytolytic function of natural killer cell receptors is altered in chronic hepatitis C. Gut 2006, 55, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Strunz, B.; Hengst, J.; Deterding, K.; Manns, M.P.; Cornberg, M.; Ljunggren, H.-G.; Wedemeyer, H.; Björkström, N.K. Chronic hepatitis C virus infection irreversibly impacts human natural killer cell repertoire diversity. Nat. Commun. 2018, 9, 2275. [Google Scholar] [CrossRef]
- Wijaya, R.S.; Read, S.A.; Selvamani, S.P.; Schibeci, S.; Azardaryany, M.K.; Ong, A.; van der Poorten, D.; Lin, R.; Douglas, M.W.; George, J.; et al. Hepatitis C virus eradication with interferon free, DAA-based therapy results in KLRG1+, hepatitis C virus-specific memory natural killer cells. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Lombardi, A.; Genco, F.; Gulminetti, R.; Novati, S.; Mondelli, M.U.; Varchetta, S. Adaptive Natural Killer Cell Functional Recovery in Hepatitis C Virus Cured Patients. Hepatology 2020. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Uldrich, A.P.; McCluskey, J.; Rossjohn, J.; Moody, D.B. The burgeoning family of unconventional T cells. Nat. Immunol. 2015, 16, 1114–1123. [Google Scholar] [CrossRef]
- Franciszkiewicz, K.; Salou, M.; Legoux, F.; Zhou, Q.; Cui, Y.; Bessoles, S.; Lantz, O. MHC class I-related molecule, MR1, and mucosal-associated invariant T cells. Immunol. Rev. 2016, 272, 120–138. [Google Scholar] [CrossRef] [PubMed]
- Hengst, J.; Strunz, B.; Deterding, K.; Ljunggren, H.G.; Leeansyah, E.; Manns, M.P.; Cornberg, M.; Sandberg, J.K.; Wedemeyer, H.; Bjorkstrom, N.K. Nonreversible MAIT cell-dysfunction in chronic hepatitis C virus infection despite successful interferon-free therapy. Eur. J. Immunol. 2016, 46, 2204–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolte, F.J.; O’Keefe, A.C.; Webb, L.M.; Serti, E.; Rivera, E.; Liang, T.J.; Ghany, M.; Rehermann, B. Intra-Hepatic Depletion of Mucosal-Associated Invariant T Cells in Hepatitis C Virus-Induced Liver Inflammation. Gastroenterology 2017, 153, 1392–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beudeker, B.J.B.; van Oord, G.W.; Arends, J.E.; Schulze Zur, W.J.; van der Heide, M.S.; de Knegt, R.J.; Verbon, A.; Boonstra, A.; Claassen, M.A.A. Mucosal-associated invariant T-cell frequency and function in blood and liver of HCV mono- and HCV/HIV co-infected patients with advanced fibrosis. Liver International. 2018, 38, 458–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wilgenburg, B.; Scherwitzl, I.; Hutchinson, E.C.; Leng, T.; Kurioka, A.; Kulicke, C.; de Lara, C.; Cole, S.; Vasanawathana, S.; Limpitikul, W.; et al. MAIT cells are activated during human viral infections. Nat. Commun. 2016, 7, 11653. [Google Scholar] [CrossRef] [Green Version]
- Spaan, M.; Hullegie, S.J.; Beudeker, B.J.; Kreefft, K.; van Oord, G.W.; Groothuismink, Z.M.; van Tilborg, M.; Rijnders, B.; de Knegt, R.J.; Claassen, M.A.; et al. Frequencies of Circulating MAIT Cells Are Diminished in Chronic HCV, HIV and HCV/HIV Co-Infection and Do Not Recover during Therapy. PLoS ONE 2016, 11, e0159243. [Google Scholar] [CrossRef] [PubMed]
- Cannizzo, E.S.; Cerrone, M.; Merlini, E.; van Wilgenburg, B.; Swadling, L.; Ancona, G.; De Bona, A.; d’Arminio Monforte, A.; Klenerman, P.; Marchetti, G. Successful direct-acting antiviral therapy in HIV/HCV co-infected patients fails to restore circulating mucosal-associated invariant T cells. Eur. J. Immunol. 2019, 49, 1127–1129. [Google Scholar] [CrossRef]
- Tseng, C.T.; Miskovsky, E.; Houghton, M.; Klimpel, G.R. Characterization of liver T-cell receptor gammadelta T cells obtained from individuals chronically infected with hepatitis C virus (HCV): Evidence for these T cells playing a role in the liver pathology associated with HCV infections. Hepatology 2001, 33, 1312–1320. [Google Scholar] [CrossRef]
- Rajoriya, N.; Fergusson, J.R.; Leithead, J.A.; Klenerman, P. Gamma Delta T-lymphocytes in Hepatitis C and Chronic Liver Disease. Front. Immunol. 2014, 5, 400. [Google Scholar] [CrossRef]
- Agrati, C.; D’Offizi, G.; Narciso, P.; Abrignani, S.; Ippolito, G.; Colizzi, V.; Poccia, F. Vdelta1 T lymphocytes expressing a Th1 phenotype are the major gammadelta T cell subset infiltrating the liver of HCV-infected persons. Mol. Med. 2001, 7, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Agrati, C.; Alonzi, T.; De Santis, R.; Castilletti, C.; Abbate, I.; Capobianchi, M.R.; D’Offizi, G.; Siepi, F.; Fimia, G.M.; Tripodi, M.; et al. Activation of Vgamma9Vdelta2 T cells by non-peptidic antigens induces the inhibition of subgenomic HCV replication. Int. Immunol. 2006, 18, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Tong, S.; Zhang, Q.; Shao, J.; Liu, Q.; Peng, H.; Hu, H.; Peng, M.; Hu, P.; Ren, H.; et al. Functional dichotomy of Vdelta2 gammadelta T cells in chronic hepatitis C virus infections: Role in cytotoxicity but not for IFN-gamma production. Sci. Rep. 2016, 6, 26296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimini, E.; Bordoni, V.; Sacchi, A.; Visco-Comandini, U.; Montalbano, M.; Taibi, C.; Casetti, R.; Lalle, E.; D’Offizi, G.; Capobianchi, M.R.; et al. Intrahepatic Vgamma9Vdelta2 T-cells from HCV-infected patients show an exhausted phenotype but can inhibit HCV replication. Virus Res. 2018, 243, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Cimini, E.; Sacchi, A.; Grassi, G.; Casetti, R.; Notari, S.; Bordoni, V.; Forini, O.; Grilli, E.; Vergori, A.; Capobianchi, M.R.; et al. Persistent gamma delta T-cell dysfunction in HCV/HIV co-infection despite direct-acting antiviral therapy-induced cure. J. Viral. Hepat. 2020, 27, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Ravens, S.; Hengst, J.; Schlapphoff, V.; Deterding, K.; Dhingra, A.; Schultze-Florey, C.; Koenecke, C.; Cornberg, M.; Wedemeyer, H.; Prinz, I. Human gammadelta T Cell Receptor Repertoires in Peripheral Blood Remain Stable Despite Clearance of Persistent Hepatitis C Virus Infection by Direct-Acting Antiviral Drug Therapy. Front. Immunol. 2018, 9, 510. [Google Scholar] [CrossRef] [Green Version]
- Schulze Zur Wiesch, J.; Ciuffreda, D.; Lewis-Ximenez, L.; Kasprowicz, V.; Nolan, B.E.; Streeck, H.; Aneja, J.; Reyor, L.L.; Allen, T.M.; Lohse, A.W.; et al. Broadly directed virus-specific CD4+ T cell responses are primed during acute hepatitis C infection, but rapidly disappear from human blood with viral persistence. J. Exp. Med. 2012, 209, 61–75. [Google Scholar] [CrossRef]
- Smits, M.; Zoldan, K.; Ishaque, N.; Gu, Z.; Jechow, K.; Wieland, D.; Conrad, C.; Eils, R.; Fauvelle, C.; Baumert, T.F.; et al. Follicular T helper cells shape the HCV-specific CD4+ T cell repertoire after virus elimination. J. Clin. Investig. 2020, 130, 998–1009. [Google Scholar] [CrossRef] [Green Version]
- Raziorrouh, B.; Sacher, K.; Tawar, R.G.; Emmerich, F.; Neumann-Haefelin, C.; Baumert, T.F.; Thimme, R.; Boettler, T. Virus-Specific CD4+ T Cells Have Functional and Phenotypic Characteristics of Follicular T-Helper Cells in Patients With Acute and Chronic HCV Infections. Gastroenterology 2016, 150, 696–706. [Google Scholar] [CrossRef]
- Meissner, E.G.; Kohli, A.; Higgins, J.; Lee, Y.J.; Prokunina, O.; Wu, D.; Orr, C.; Masur, H.; Kottilil, S. Rapid changes in peripheral lymphocyte concentrations during interferon-free treatment of chronic hepatitis C virus infection. Hepatol. Commun. 2017, 1, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Meissner, E.G.; Decalf, J.; Casrouge, A.; Masur, H.; Kottilil, S.; Albert, M.L.; Duffy, D. Dynamic Changes of Post-Translationally Modified Forms of CXCL10 and Soluble DPP4 in HCV Subjects Receiving Interferon-Free Therapy. PLoS ONE 2015, 10, e0133236. [Google Scholar] [CrossRef]
- Schlaphoff, V.; Lunemann, S.; Suneetha, P.V.; Jaroszewicz, J.; Grabowski, J.; Dietz, J.; Helfritz, F.; Bektas, H.; Sarrazin, C.; Manns, M.P.; et al. Dual function of the NK cell receptor 2B4 (CD244) in the regulation of HCV-specific CD8+ T cells. PLoS Pathog. 2011, 7, e1002045. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Y.; Wolski, D.; Aneja, J.; Matsubara, L.; Robilotti, B.; Hauck, G.; de Sousa, P.S.F.; Subudhi, S.; Fernandes, C.A.; Hoogeveen, R.C.; et al. Hepatitis C virus-specific CD4+ T cell phenotype and function in different infection outcomes. J. Clin. Investig. 2020, 130, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, C.; Smits, M.; Woost, R.; Eberhard, J.M.; Peine, S.; Kummer, S.; Marget, M.; Kuntzen, T.; Kwok, W.W.; Lohse, A.W.; et al. HCV-specific CD4+ T cells of patients with acute and chronic HCV infection display high expression of TIGIT and other co-inhibitory molecules. Sci. Rep. 2019, 9, 10624. [Google Scholar] [CrossRef] [Green Version]
- Telatin, V.; Nicoli, F.; Frasson, C.; Menegotto, N.; Barbaro, F.; Castelli, E.; Erne, E.; Palu, G.; Caputo, A. In Chronic Hepatitis C Infection, Myeloid-Derived Suppressor Cell Accumulation and T Cell Dysfunctions Revert Partially and Late After Successful Direct-Acting Antiviral Treatment. Front. Cell Infect. Microbiol. 2019, 9, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burchill, M.A.; Golden-Mason, L.; Wind-Rotolo, M.; Rosen, H.R. Memory re-differentiation and reduced lymphocyte activation in chronic HCV-infected patients receiving direct-acting antivirals. J. Viral. Hepat. 2015, 22, 983–991. [Google Scholar] [CrossRef]
- Szereday, L.; Meggyes, M.; Berki, T.; Miseta, A.; Farkas, N.; Gervain, J.; Par, A.; Par, G. Direct-acting antiviral treatment downregulates immune checkpoint inhibitor expression in patients with chronic hepatitis C. Clin. Exp. Med. 2020, 20, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Najafi Fard, S.; Schietroma, I.; Corano Scheri, G.; Giustini, N.; Serafino, S.; Cavallari, E.N.; Pinacchio, C.; De Girolamo, G.; Ceccarelli, G.; Scagnolari, C.; et al. Direct-acting antiviral therapy enhances total CD4+ and CD8+ T-cells responses, but does not alter T-cells activation among HCV mono-infected, and HCV/HIV-1 co-infected patients. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 319–329. [Google Scholar] [CrossRef]
- Spaan, M.; Kreefft, K.; de Graav, G.N.; Brouwer, W.P.; de Knegt, R.J.; ten Kate, F.J.; Baan, C.C.; Vanwolleghem, T.; Janssen, H.L.; Boonstra, A. CD4+ CXCR5+ T cells in chronic HCV infection produce less IL-21, yet are efficient at supporting B cell responses. J. Hepatol. 2015, 62, 303–310. [Google Scholar] [CrossRef]
- Comarmond, C.; Garrido, M.; Pol, S.; Desbois, A.C.; Costopoulos, M.; Le Garff-Tavernier, M.; Si Ahmed, S.N.; Alric, L.; Fontaine, H.; Bellier, B.; et al. Direct-Acting Antiviral Therapy Restores Immune Tolerance to Patients With Hepatitis C Virus-Induced Cryoglobulinemia Vasculitis. Gastroenterology 2017, 152, 2052–2062 e2052. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.H.; Jiang, B.C.; Liu, X.H.; Zhang, M.X.; Li, Z.S.; Zhu, G.Z. Interleukin-7 Regulates T Follicular Helper Cell Function in Patients with Chronic Hepatitis C. Viral. Immunol. 2018, 31, 417–425. [Google Scholar] [CrossRef]
- Keoshkerian, E.; Hunter, M.; Cameron, B.; Nguyen, N.; Sugden, P.; Bull, R.; Zekry, A.; Maher, L.; Seddiki, N.; Zaunders, J.; et al. Hepatitis C-specific effector and regulatory CD4 T-cell responses are associated with the outcomes of primary infection. J. Viral. Hepat. 2016, 23, 985–993. [Google Scholar] [CrossRef]
- Mengshol, J.A.; Golden-Mason, L.; Arikawa, T.; Smith, M.; Niki, T.; McWilliams, R.; Randall, J.A.; McMahan, R.; Zimmerman, M.A.; Rangachari, M.; et al. A crucial role for Kupffer cell-derived galectin-9 in regulation of T cell immunity in hepatitis C infection. PLoS ONE 2010, 5, e9504. [Google Scholar] [CrossRef]
- Elsegood, C.L.; Tirnitz-Parker, J.E.; Olynyk, J.K.; Yeoh, G.C. Immune checkpoint inhibition: Prospects for prevention and therapy of hepatocellular carcinoma. Clin. Transl. Immunol. 2017, 6, e161. [Google Scholar] [CrossRef] [PubMed]
- Claassen, M.A.; de Knegt, R.J.; Tilanus, H.W.; Janssen, H.L.; Boonstra, A. Abundant numbers of regulatory T cells localize to the liver of chronic hepatitis C infected patients and limit the extent of fibrosis. J. Hepatol. 2010, 52, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.M.; Fox, B.C.; Brown, P.J.; Worthington, J.; Fox, S.B.; Chapman, R.W.; Fleming, K.A.; Banham, A.H.; Klenerman, P. Quantification and localisation of FOXP3+ T lymphocytes and relation to hepatic inflammation during chronic HCV infection. J. Hepatol. 2007, 47, 316–324. [Google Scholar] [CrossRef]
- Wei, K.; Jiang, B.C.; Guan, J.H.; Zhang, D.N.; Zhang, M.X.; Wu, J.L.; Zhu, G.Z. Decreased CD4(+)CD25(+)CD127(dim/-) Regulatory T Cells and T Helper 17 Cell Responsiveness to Toll-Like Receptor 2 in Chronic Hepatitis C Patients with Daclatasvir Plus Asunaprevir Therapy. Viral Immunol. 2018, 31, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.F.; Tseng, C.W.; Ho, Y.C.; Chen, Y.C.; Ko, P.H.; He, Y.T.; Tseng, K.C. Regulatory T Cell Function Modulated After Successful Direct-Acting Antiviral Treatment for Chronic Hepatitis C Patients. Dig. Dis. Sci. 2020, 65, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Langhans, B.; Nischalke, H.D.; Kramer, B.; Hausen, A.; Dold, L.; van Heteren, P.; Huneburg, R.; Nattermann, J.; Strassburg, C.P.; Spengler, U. Increased peripheral CD4(+) regulatory T cells persist after successful direct-acting antiviral treatment of chronic hepatitis C. J. Hepatol. 2017, 66, 888–896. [Google Scholar] [CrossRef]
- Owusu Sekyere, S.; Suneetha, P.V.; Kraft, A.R.; Zhang, S.; Dietz, J.; Sarrazin, C.; Manns, M.P.; Schlaphoff, V.; Cornberg, M.; Wedemeyer, H. A heterogeneous hierarchy of co-regulatory receptors regulates exhaustion of HCV-specific CD8 T cells in patients with chronic hepatitis C. J. Hepatol. 2015, 62, 31–40. [Google Scholar] [CrossRef]
- Bengsch, B.; Seigel, B.; Ruhl, M.; Timm, J.; Kuntz, M.; Blum, H.E.; Pircher, H.; Thimme, R. Coexpression of PD-1, 2B4, CD160 and KLRG1 on exhausted HCV-specific CD8+ T cells is linked to antigen recognition and T cell differentiation. PLoS Pathog. 2010, 6, e1000947. [Google Scholar] [CrossRef]
- Wedemeyer, H.; He, X.S.; Nascimbeni, M.; Davis, A.R.; Greenberg, H.B.; Hoofnagle, J.H.; Liang, T.J.; Alter, H.; Rehermann, B. Impaired effector function of hepatitis C virus-specific CD8+ T cells in chronic hepatitis C virus infection. J. Immunol. 2002, 169, 3447–3458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitschke, K.; Flecken, T.; Schmidt, J.; Gostick, E.; Marget, M.; Neumann-Haefelin, C.; Blum, H.E.; Price, D.A.; Thimme, R. Tetramer enrichment reveals the presence of phenotypically diverse hepatitis C virus-specific CD8+ T cells in chronic infection. J. Virol. 2015, 89, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penna, A.; Pilli, M.; Zerbini, A.; Orlandini, A.; Mezzadri, S.; Sacchelli, L.; Missale, G.; Ferrari, C. Dysfunction and functional restoration of HCV-specific CD8 responses in chronic hepatitis C virus infection. Hepatology 2007, 45, 588–601. [Google Scholar] [CrossRef] [PubMed]
- Kasprowicz, V.; Kang, Y.H.; Lucas, M.; Schulze zur Wiesch, J.; Kuntzen, T.; Fleming, V.; Nolan, B.E.; Longworth, S.; Berical, A.; Bengsch, B.; et al. Hepatitis C virus (HCV) sequence variation induces an HCV-specific T-cell phenotype analogous to spontaneous resolution. J. Virol. 2010, 84, 1656–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, S.D.; Shin, H.; Haining, W.N.; Zou, T.; Workman, C.J.; Polley, A.; Betts, M.R.; Freeman, G.J.; Vignali, D.A.; Wherry, E.J. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nat. Immunol. 2009, 10, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Bengsch, B.; Spangenberg, H.C.; Kersting, N.; Neumann-Haefelin, C.; Panther, E.; von Weizsacker, F.; Blum, H.E.; Pircher, H.; Thimme, R. Analysis of CD127 and KLRG1 expression on hepatitis C virus-specific CD8+ T cells reveals the existence of different memory T-cell subsets in the peripheral blood and liver. J. Virol. 2007, 81, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.; Hennecke, N.; Lohmann, V.; Kayser, A.; Neumann-Haefelin, C.; Kukolj, G.; Bocher, W.O.; Thimme, R. Restoration of HCV-specific CD8+ T cell function by interferon-free therapy. J. Hepatol. 2014, 61, 538–543. [Google Scholar] [CrossRef]
- Aregay, A.; Owusu Sekyere, S.; Deterding, K.; Port, K.; Dietz, J.; Berkowski, C.; Sarrazin, C.; Manns, M.P.; Cornberg, M.; Wedemeyer, H. Elimination of hepatitis C virus has limited impact on the functional and mitochondrial impairment of HCV-specific CD8+ T cell responses. J. Hepatol. 2019, 71, 889–899. [Google Scholar] [CrossRef]
- Khan, O.; Giles, J.R.; McDonald, S.; Manne, S.; Ngiow, S.F.; Patel, K.P.; Werner, M.T.; Huang, A.C.; Alexander, K.A.; Wu, J.E.; et al. TOX transcriptionally and epigenetically programs CD8(+) T cell exhaustion. Nature 2019, 571, 211–218. [Google Scholar] [CrossRef]
- Alfei, F.; Kanev, K.; Hofmann, M.; Wu, M.; Ghoneim, H.E.; Roelli, P.; Utzschneider, D.T.; von Hoesslin, M.; Cullen, J.G.; Fan, Y.; et al. TOX reinforces the phenotype and longevity of exhausted T cells in chronic viral infection. Nature 2019, 571, 265–269. [Google Scholar] [CrossRef]
- Crispe, I.N. The liver as a lymphoid organ. Annu. Rev. Immunol. 2009, 27, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, C. The Role of Innate Lymphoid Cells in Immune-Mediated Liver Diseases. Front. Immunol. 2017, 8, 695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunemann, S.; Schlaphoff, V.; Cornberg, M.; Wedemeyer, H. NK cells in hepatitis C: Role in disease susceptibility and therapy. Dig. Dis. 2012, 30 (Suppl. 1), 48–54. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Szabo, G. Innate immune cell networking in hepatitis C virus infection. J. Leukoc. Biol. 2014, 96, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsumi, T.; Takehara, T. Impact of natural killer cells on chronic hepatitis C and hepatocellular carcinoma. Hepatol. Res. 2016, 46, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Oh, J.S.; Ali, A.K.; Kim, S.; Corsi, D.J.; Cooper, C.L.; Lee, S.H. NK cells lacking FcepsilonRIgamma are associated with reduced liver damage in chronic hepatitis C virus infection. Eur. J. Immunol. 2016, 46, 1020–1029. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.K.; Guirgis, M.; Dinh, T.; George, J.; Dev, A.; Lee, A.; Zekry, A. Treatment responses in Asians and Caucasians with chronic hepatitis C infection. World J. Gastroenterol. 2008, 14, 3416–3420. [Google Scholar] [CrossRef]
- Su, F.; Green, P.K.; Berry, K.; Ioannou, G.N. The association between race/ethnicity and the effectiveness of direct antiviral agents for hepatitis C virus infection. Hepatology 2017, 65, 426–438. [Google Scholar] [CrossRef]
- Hunter, S.; Willcox, C.R.; Davey, M.S.; Kasatskaya, S.A.; Jeffery, H.C.; Chudakov, D.M.; Oo, Y.H.; Willcox, B.E. Human liver infiltrating gammadelta T cells are composed of clonally expanded circulating and tissue-resident populations. J. Hepatol. 2018, 69, 654–665. [Google Scholar] [CrossRef]
- Thein, H.H.; Yi, Q.; Dore, G.J.; Krahn, M.D. Natural history of hepatitis C virus infection in HIV-infected individuals and the impact of HIV in the era of highly active antiretroviral therapy: A meta-analysis. AIDS 2008, 22, 1979–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuyama, A.C.; Hong, F.; Saiman, Y.; Wang, C.; Ozkok, D.; Mosoian, A.; Chen, P.; Chen, B.K.; Klotman, M.E.; Bansal, M.B. Human immunodeficiency virus (HIV)-1 infects human hepatic stellate cells and promotes collagen I and monocyte chemoattractant protein-1 expression: Implications for the pathogenesis of HIV/hepatitis C virus-induced liver fibrosis. Hepatology 2010, 52, 612–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockstroh, J.K.; Spengler, U.; Sudhop, T.; Ewig, S.; Theisen, A.; Hammerstein, U.; Bierhoff, E.; Fischer, H.P.; Oldenburg, J.; Brackmann, H.H.; et al. Immunosuppression may lead to progression of hepatitis C virus-associated liver disease in hemophiliacs coinfected with HIV. Am. J. Gastroenterol. 1996, 91, 2563–2568. [Google Scholar] [PubMed]
- Callendret, B.; Eccleston, H.B.; Satterfield, W.; Capone, S.; Folgori, A.; Cortese, R.; Nicosia, A.; Walker, C.M. Persistent hepatitis C viral replication despite priming of functional CD8+ T cells by combined therapy with a vaccine and a direct-acting antiviral. Hepatology 2016, 63, 1442–1454. [Google Scholar] [CrossRef] [Green Version]
- Ingiliz, P.; Martin, T.C.; Rodger, A.; Stellbrink, H.J.; Mauss, S.; Boesecke, C.; Mandorfer, M.; Bottero, J.; Baumgarten, A.; Bhagani, S.; et al. HCV reinfection incidence and spontaneous clearance rates in HIV-positive men who have sex with men in Western Europe. J. Hepatol. 2017, 66, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Grebely, J.; Dore, G.J.; Greenwald, R.; Swan, T.; Barua, S.; Taylor, L.E. Hepatitis C Virus Treatment and Persons Who Inject Drugs. Ann. Intern. Med. 2016, 164, 203. [Google Scholar] [CrossRef]
- Hartnell, F.; Esposito, I.; Swadling, L.; Brown, A.; Phetsouphanh, C.; de Lara, C.; Gentile, C.; Turner, B.; Kopycinski, J.; Dorrell, L.; et al. Characterising HCV specific CD4+ T-cells following viral-vectored vaccination, directly acting anti-virals and spontaneous viral cure. Hepatology 2020. [Google Scholar] [CrossRef]
- Kelly, C.; Swadling, L.; Capone, S.; Brown, A.; Richardson, R.; Halliday, J.; von Delft, A.; Oo, Y.; Mutimer, D.; Kurioka, A.; et al. Chronic hepatitis C viral infection subverts vaccine-induced T-cell immunity in humans. Hepatology 2016, 63, 1455–1470. [Google Scholar] [CrossRef]
- Swadling, L.; Halliday, J.; Kelly, C.; Brown, A.; Capone, S.; Ansari, M.A.; Bonsall, D.; Richardson, R.; Hartnell, F.; Collier, J.; et al. Highly-Immunogenic Virally-Vectored T-cell Vaccines Cannot Overcome Subversion of the T-cell Response by HCV during Chronic Infection. Vaccines 2016, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Wieland, D.; Thimme, R. Vaccine-induced hepatitis C virus-specific CD8+ T cells do not always help. Hepatology 2016, 63, 1411–1414. [Google Scholar] [CrossRef]
- Honegger, J.R.; Tedesco, D.; Kohout, J.A.; Prasad, M.R.; Price, A.A.; Lindquist, T.; Ohmer, S.; Moore-Clingenpeel, M.; Grakoui, A.; Walker, C.M. Influence of IFNL3 and HLA-DPB1 genotype on postpartum control of hepatitis C virus replication and T-cell recovery. Proc. Natl. Acad. Sci. USA 2016, 113, 10684–10689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, M.; Jhaveri, R.; Saleh, D.A.; Sharaf, S.A.; El-Mougy, F.; Abdelsalam, L.; Shardell, M.D.; El-Ghazaly, H.; El-Kamary, S.S. Spontaneous Viral Load Decline and Subsequent Clearance of Chronic Hepatitis C Virus in Postpartum Women Correlates With Favorable Interleukin-28B Gene Allele. Clin. Infect. Dis. 2017, 65, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Coss, S.L.; Torres-Cornejo, A.; Prasad, M.R.; Moore-Clingenpeel, M.; Grakoui, A.; Lauer, G.M.; Walker, C.M.; Honegger, J.R. CD4+ T cell restoration and control of hepatitis C virus replication after childbirth. J. Clin. Investig. 2020, 130, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Breitfeld, D.; Ohl, L.; Kremmer, E.; Ellwart, J.; Sallusto, F.; Lipp, M.; Forster, R. Follicular B helper T cells express CXC chemokine receptor 5, localize to B cell follicles, and support immunoglobulin production. J. Exp. Med. 2000, 192, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Schaerli, P.; Willimann, K.; Lang, A.B.; Lipp, M.; Loetscher, P.; Moser, B. CXC chemokine receptor 5 expression defines follicular homing T cells with B cell helper function. J. Exp. Med. 2000, 192, 1553–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goenka, R.; Barnett, L.G.; Silver, J.S.; O’Neill, P.J.; Hunter, C.A.; Cancro, M.P.; Laufer, T.M. Cutting edge: Dendritic cell-restricted antigen presentation initiates the follicular helper T cell program but cannot complete ultimate effector differentiation. J. Immunol. 2011, 187, 1091–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, R.J.; Poholek, A.C.; DiToro, D.; Yusuf, I.; Eto, D.; Barnett, B.; Dent, A.L.; Craft, J.; Crotty, S. Bcl6 and Blimp-1 are reciprocal and antagonistic regulators of T follicular helper cell differentiation. Science 2009, 325, 1006–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurieva, R.I.; Chung, Y.; Martinez, G.J.; Yang, X.O.; Tanaka, S.; Matskevitch, T.D.; Wang, Y.H.; Dong, C. Bcl6 mediates the development of T follicular helper cells. Science 2009, 325, 1001–1005. [Google Scholar] [CrossRef] [Green Version]
- Vinuesa, C.G.; Cyster, J.G. How T cells earn the follicular rite of passage. Immunity 2011, 35, 671–680. [Google Scholar] [CrossRef] [Green Version]
- King, C. A fine romance: T follicular helper cells and B cells. Immunity 2011, 34, 827–829. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Li, X.; Liu, D.; Li, J.; Zhang, X.; Chen, X.; Hou, S.; Peng, L.; Xu, C.; Liu, W.; et al. Follicular T-helper cell recruitment governed by bystander B cells and ICOS-driven motility. Nature 2013, 496, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Linterman, M.A.; Pierson, W.; Lee, S.K.; Kallies, A.; Kawamoto, S.; Rayner, T.F.; Srivastava, M.; Divekar, D.P.; Beaton, L.; Hogan, J.J.; et al. Foxp3+ follicular regulatory T cells control the germinal center response. Nat. Med. 2011, 17, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.; Tanaka, S.; Chu, F.; Nurieva, R.I.; Martinez, G.J.; Rawal, S.; Wang, Y.H.; Lim, H.; Reynolds, J.M.; Zhou, X.H.; et al. Follicular regulatory T cells expressing Foxp3 and Bcl-6 suppress germinal center reactions. Nat. Med. 2011, 17, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, I.; Agua-Doce, A.; Hernandez, A.; Almeida, C.; Oliveira, V.G.; Faro, J.; Graca, L. Regulation of the germinal center reaction by Foxp3+ follicular regulatory T cells. J. Immunol. 2011, 187, 4553–4560. [Google Scholar] [CrossRef] [Green Version]
- Sage, P.T.; Sharpe, A.H. T follicular regulatory cells in the regulation of B cell responses. Trends Immunol. 2015, 36, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Qiu, J.; Yu, L.; Hu, X.; Zhao, P.; Jiang, Y. Increased numbers of CD5+CD19+CD1dhighIL-10+ Bregs, CD4+Foxp3+ Tregs, CD4+CXCR5+Foxp3+ follicular regulatory T (TFR) cells in CHB or CHC patients. J. Transl. Med. 2014, 12, 251. [Google Scholar] [CrossRef] [Green Version]
- Cobb, D.A.; Kim, O.K.; Golden-Mason, L.; Rosen, H.R.; Hahn, Y.S. Hepatocyte-derived exosomes promote T follicular regulatory cell expansion during hepatitis C virus infection. Hepatology 2018, 67, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cao, D.; Wang, L.; Zhao, J.; Nguyen, L.N.; Dang, X.; Ji, Y.; Wu, X.Y.; Morrison, Z.D.; Xie, Q.; et al. HCV-associated exosomes promote myeloid-derived suppressor cell expansion via inhibiting miR-124 to regulate T follicular cell differentiation and function. Cell Discov. 2018, 4, 51. [Google Scholar] [CrossRef]
- Shevach, E.M.; DiPaolo, R.A.; Andersson, J.; Zhao, D.M.; Stephens, G.L.; Thornton, A.M. The lifestyle of naturally occurring CD4+ CD25+ Foxp3+ regulatory T cells. Immunol. Rev. 2006, 212, 60–73. [Google Scholar] [CrossRef]
- Zhao, D.M.; Thornton, A.M.; DiPaolo, R.J.; Shevach, E.M. Activated CD4+CD25+ T cells selectively kill B lymphocytes. Blood 2006, 107, 3925–3932. [Google Scholar] [CrossRef] [Green Version]
- Ghiringhelli, F.; Menard, C.; Martin, F.; Zitvogel, L. The role of regulatory T cells in the control of natural killer cells: Relevance during tumor progression. Immunol. Rev. 2006, 214, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, K.; Enk, A.H. Dendritic cells: Key cells for the induction of regulatory T cells? Curr. Top. Microbiol. Immunol. 2005, 293, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Barjon, C.; Dahlqvist, G.; Calmus, Y.; Conti, F. Role of regulatory T-cells during hepatitis C infection: From the acute phase to post-transplantation recurrence. Dig. Liver Dis. 2015, 47, 913–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riezu-Boj, J.I.; Larrea, E.; Aldabe, R.; Guembe, L.; Casares, N.; Galeano, E.; Echeverria, I.; Sarobe, P.; Herrero, I.; Sangro, B.; et al. Hepatitis C virus induces the expression of CCL17 and CCL22 chemokines that attract regulatory T cells to the site of infection. J. Hepatol. 2011, 54, 422–431. [Google Scholar] [CrossRef]
- Higashitani, K.; Kanto, T.; Kuroda, S.; Yoshio, S.; Matsubara, T.; Kakita, N.; Oze, T.; Miyazaki, M.; Sakakibara, M.; Hiramatsu, N.; et al. Association of enhanced activity of indoleamine 2,3-dioxygenase in dendritic cells with the induction of regulatory T cells in chronic hepatitis C infection. J. Gastroenterol. 2013, 48, 660–670. [Google Scholar] [CrossRef]
- Langhans, B.; Kramer, B.; Louis, M.; Nischalke, H.D.; Huneburg, R.; Staratschek-Jox, A.; Odenthal, M.; Manekeller, S.; Schepke, M.; Kalff, J.; et al. Intrahepatic IL-8 producing Foxp3(+)CD4(+) regulatory T cells and fibrogenesis in chronic hepatitis C. J. Hepatol. 2013, 59, 229–235. [Google Scholar] [CrossRef]
- Zhao, H.Q.; Li, W.M.; Lu, Z.Q.; Yao, Y.M. Roles of Tregs in development of hepatocellular carcinoma: A meta-analysis. World J. Gastroenterol. 2014, 20, 7971–7978. [Google Scholar] [CrossRef]
- Claassen, M.A.; de Knegt, R.J.; Janssen, H.L.; Boonstra, A. Retention of CD4+ CD25+ FoxP3+ regulatory T cells in the liver after therapy-induced hepatitis C virus eradication in humans. J. Virol. 2011, 85, 5323–5330. [Google Scholar] [CrossRef] [Green Version]
- Bolacchi, F.; Sinistro, A.; Ciaprini, C.; Demin, F.; Capozzi, M.; Carducci, F.C.; Drapeau, C.M.; Rocchi, G.; Bergamini, A. Increased hepatitis C virus (HCV)-specific CD4+CD25+ regulatory T lymphocytes and reduced HCV-specific CD4+ T cell response in HCV-infected patients with normal versus abnormal alanine aminotransferase levels. Clin. Exp. Immunol. 2006, 144, 188–196. [Google Scholar] [CrossRef]
- Hartling, H.J.; Gaardbo, J.C.; Ronit, A.; Knudsen, L.S.; Ullum, H.; Vainer, B.; Clausen, M.R.; Skogstrand, K.; Gerstoft, J.; Nielsen, S.D. CD4(+) and CD8(+) regulatory T cells (Tregs) are elevated and display an active phenotype in patients with chronic HCV mono-infection and HIV/HCV co-infection. Scand. J. Immunol. 2012, 76, 294–305. [Google Scholar] [CrossRef]
- Han, J.W.; Sung, P.S.; Hong, S.H.; Lee, H.; Koh, J.Y.; Lee, H.; White, S.; Maslow, J.N.; Weiner, D.B.; Park, S.H.; et al. IFNL3-adjuvanted HCV DNA vaccine reduces regulatory T cell frequency and increases virus-specific T cell responses. J. Hepatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006, 441, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Boniface, K.; Bernard, F.X.; Garcia, M.; Gurney, A.L.; Lecron, J.C.; Morel, F. IL-22 inhibits epidermal differentiation and induces proinflammatory gene expression and migration of human keratinocytes. J. Immunol. 2005, 174, 3695–3702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilloteau, K.; Paris, I.; Pedretti, N.; Boniface, K.; Juchaux, F.; Huguier, V.; Guillet, G.; Bernard, F.X.; Lecron, J.C.; Morel, F. Skin Inflammation Induced by the Synergistic Action of IL-17A, IL-22, Oncostatin M, IL-1{alpha}, and TNF-{alpha} Recapitulates Some Features of Psoriasis. J. Immunol. 2010, 184, 5263–5270. [Google Scholar] [CrossRef] [Green Version]
- Hirota, K.; Duarte, J.H.; Veldhoen, M.; Hornsby, E.; Li, Y.; Cua, D.J.; Ahlfors, H.; Wilhelm, C.; Tolaini, M.; Menzel, U.; et al. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat. Immunol. 2011, 12, 255–263. [Google Scholar] [CrossRef]
- Yang, X.O.; Pappu, B.P.; Nurieva, R.; Akimzhanov, A.; Kang, H.S.; Chung, Y.; Ma, L.; Shah, B.; Panopoulos, A.D.; Schluns, K.S.; et al. T helper 17 lineage differentiation is programmed by orphan nuclear receptors ROR alpha and ROR gamma. Immunity 2008, 28, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [Green Version]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.; Wang, Y.K.; Zhao, Q.; Wang, C.Z.; Hu, Y.Z.; Wu, B.Y. Th17 cells are increased with severity of liver inflammation in patients with chronic hepatitis C. J. Gastroenterol. Hepatol. 2012, 27, 273–278. [Google Scholar] [CrossRef]
- Paquissi, F.C. Immunity and Fibrogenesis: The Role of Th17/IL-17 Axis in HBV and HCV-induced Chronic Hepatitis and Progression to Cirrhosis. Front. Immunol. 2017, 8, 1195. [Google Scholar] [CrossRef] [Green Version]
- Rios, D.A.; Valva, P.; Casciato, P.C.; Frias, S.; Soledad Caldirola, M.; Gaillard, M.I.; Bezrodnik, L.; Bandi, J.; Galdame, O.; Ameigeiras, B.; et al. Chronic hepatitis C liver microenvironment: Role of the Th17/Treg interplay related to fibrogenesis. Sci. Rep. 2017, 7, 13283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kared, H.; Fabre, T.; Bedard, N.; Bruneau, J.; Shoukry, N.H. Galectin-9 and IL-21 mediate cross-regulation between Th17 and Treg cells during acute hepatitis C. PLoS Pathog. 2013, 9, e1003422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Leeuwen, E.M.; de Bree, G.J.; Remmerswaal, E.B.; Yong, S.L.; Tesselaar, K.; ten Berge, I.J.; van Lier, R.A. IL-7 receptor alpha chain expression distinguishes functional subsets of virus-specific human CD8+ T cells. Blood 2005, 106, 2091–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boettler, T.; Panther, E.; Bengsch, B.; Nazarova, N.; Spangenberg, H.C.; Blum, H.E.; Thimme, R. Expression of the interleukin-7 receptor alpha chain (CD127) on virus-specific CD8+ T cells identifies functionally and phenotypically defined memory T cells during acute resolving hepatitis B virus infection. J. Virol. 2006, 80, 3532–3540. [Google Scholar] [CrossRef] [Green Version]
- Fry, T.J.; Mackall, C.L. The many faces of IL-7: From lymphopoiesis to peripheral T cell maintenance. J. Immunol. 2005, 174, 6571–6576. [Google Scholar] [CrossRef]
- Bowen, D.G.; Shoukry, N.H.; Grakoui, A.; Fuller, M.J.; Cawthon, A.G.; Dong, C.; Hasselschwert, D.L.; Brasky, K.M.; Freeman, G.J.; Seth, N.P.; et al. Variable patterns of programmed death-1 expression on fully functional memory T cells after spontaneous resolution of hepatitis C virus infection. J. Virol. 2008, 82, 5109–5114. [Google Scholar] [CrossRef] [Green Version]
- Odorizzi, P.M.; Pauken, K.E.; Paley, M.A.; Sharpe, A.; Wherry, E.J. Genetic absence of PD-1 promotes accumulation of terminally differentiated exhausted CD8+ T cells. J. Exp. Med. 2015, 212, 1125–1137. [Google Scholar] [CrossRef]
- Wolski, D.; Foote, P.K.; Chen, D.Y.; Lewis-Ximenez, L.L.; Fauvelle, C.; Aneja, J.; Walker, A.; Tonnerre, P.; Torres-Cornejo, A.; Kvistad, D.; et al. Early Transcriptional Divergence Marks Virus-Specific Primary Human CD8(+) T Cells in Chronic versus Acute Infection. Immunity 2017, 47, 648–663.e648. [Google Scholar] [CrossRef] [Green Version]
- Bengsch, B.; Johnson, A.L.; Kurachi, M.; Odorizzi, P.M.; Pauken, K.E.; Attanasio, J.; Stelekati, E.; McLane, L.M.; Paley, M.A.; Delgoffe, G.M.; et al. Bioenergetic Insufficiencies Due to Metabolic Alterations Regulated by the Inhibitory Receptor PD-1 Are an Early Driver of CD8(+) T Cell Exhaustion. Immunity 2016, 45, 358–373. [Google Scholar] [CrossRef] [Green Version]
- Fisicaro, P.; Barili, V.; Montanini, B.; Acerbi, G.; Ferracin, M.; Guerrieri, F.; Salerno, D.; Boni, C.; Massari, M.; Cavallo, M.C.; et al. Targeting mitochondrial dysfunction can restore antiviral activity of exhausted HBV-specific CD8 T cells in chronic hepatitis B. Nat. Med. 2017, 23, 327–336. [Google Scholar] [CrossRef]
- Schurich, A.; Pallett, L.J.; Jajbhay, D.; Wijngaarden, J.; Otano, I.; Gill, U.S.; Hansi, N.; Kennedy, P.T.; Nastouli, E.; Gilson, R.; et al. Distinct Metabolic Requirements of Exhausted and Functional Virus-Specific CD8 T Cells in the Same Host. Cell Rep. 2016, 16, 1243–1252. [Google Scholar] [CrossRef] [Green Version]
- Sena, L.A.; Li, S.; Jairaman, A.; Prakriya, M.; Ezponda, T.; Hildeman, D.A.; Wang, C.R.; Schumacker, P.T.; Licht, J.D.; Perlman, H.; et al. Mitochondria are required for antigen-specific T cell activation through reactive oxygen species signaling. Immunity 2013, 38, 225–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, M.D.; O’Sullivan, D.; Pearce, E.L. T cell metabolism drives immunity. J. Exp. Med. 2015, 212, 1345–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devadas, S.; Zaritskaya, L.; Rhee, S.G.; Oberley, L.; Williams, M.S. Discrete generation of superoxide and hydrogen peroxide by T cell receptor stimulation: Selective regulation of mitogen-activated protein kinase activation and fas ligand expression. J. Exp. Med. 2002, 195, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comarmond, C.; Lorin, V.; Marques, C.; Maciejewski-Duval, A.; Joher, N.; Planchais, C.; Touzot, M.; Biard, L.; Hieu, T.; Quiniou, V.; et al. TLR9 signalling in HCV-associated atypical memory B cells triggers Th1 and rheumatoid factor autoantibody responses. J. Hepatol. 2019, 71, 908–919. [Google Scholar] [CrossRef]
- Shoukry, N.H.; Grakoui, A.; Houghton, M.; Chien, D.Y.; Ghrayeb, J.; Reimann, K.A.; Walker, C.M. Memory CD8+ T cells are required for protection from persistent hepatitis C virus infection. J. Exp. Med. 2003, 197, 1645–1655. [Google Scholar] [CrossRef]
- Pallett, L.J.; Davies, J.; Colbeck, E.J.; Robertson, F.; Hansi, N.; Easom, N.J.W.; Burton, A.R.; Stegmann, K.A.; Schurich, A.; Swadling, L.; et al. IL-2(high) tissue-resident T cells in the human liver: Sentinels for hepatotropic infection. J. Exp. Med. 2017, 214, 1567–1580. [Google Scholar] [CrossRef]
- Stelma, F.; de Niet, A.; Sinnige, M.J.; van Dort, K.A.; van Gisbergen, K.; Verheij, J.; van Leeuwen, E.M.M.; Kootstra, N.A.; Reesink, H.W. Human intrahepatic CD69 + CD8+ T cells have a tissue resident memory T cell phenotype with reduced cytolytic capacity. Sci. Rep. 2017, 7, 6172. [Google Scholar] [CrossRef] [Green Version]
- Virgin, H.W.; Wherry, E.J.; Ahmed, R. Redefining chronic viral infection. Cell 2009, 138, 30–50. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Visual summary of perturbations in frequency, phenotypes, and function of innate (A) and adaptive (B) lymphocytes in chronic hepatitis C virus (HCV) (CHC) patients and restoration associated with direct acting antiviral (DAA)-mediated viral clearance.
Figure 1.
Visual summary of perturbations in frequency, phenotypes, and function of innate (A) and adaptive (B) lymphocytes in chronic hepatitis C virus (HCV) (CHC) patients and restoration associated with direct acting antiviral (DAA)-mediated viral clearance.



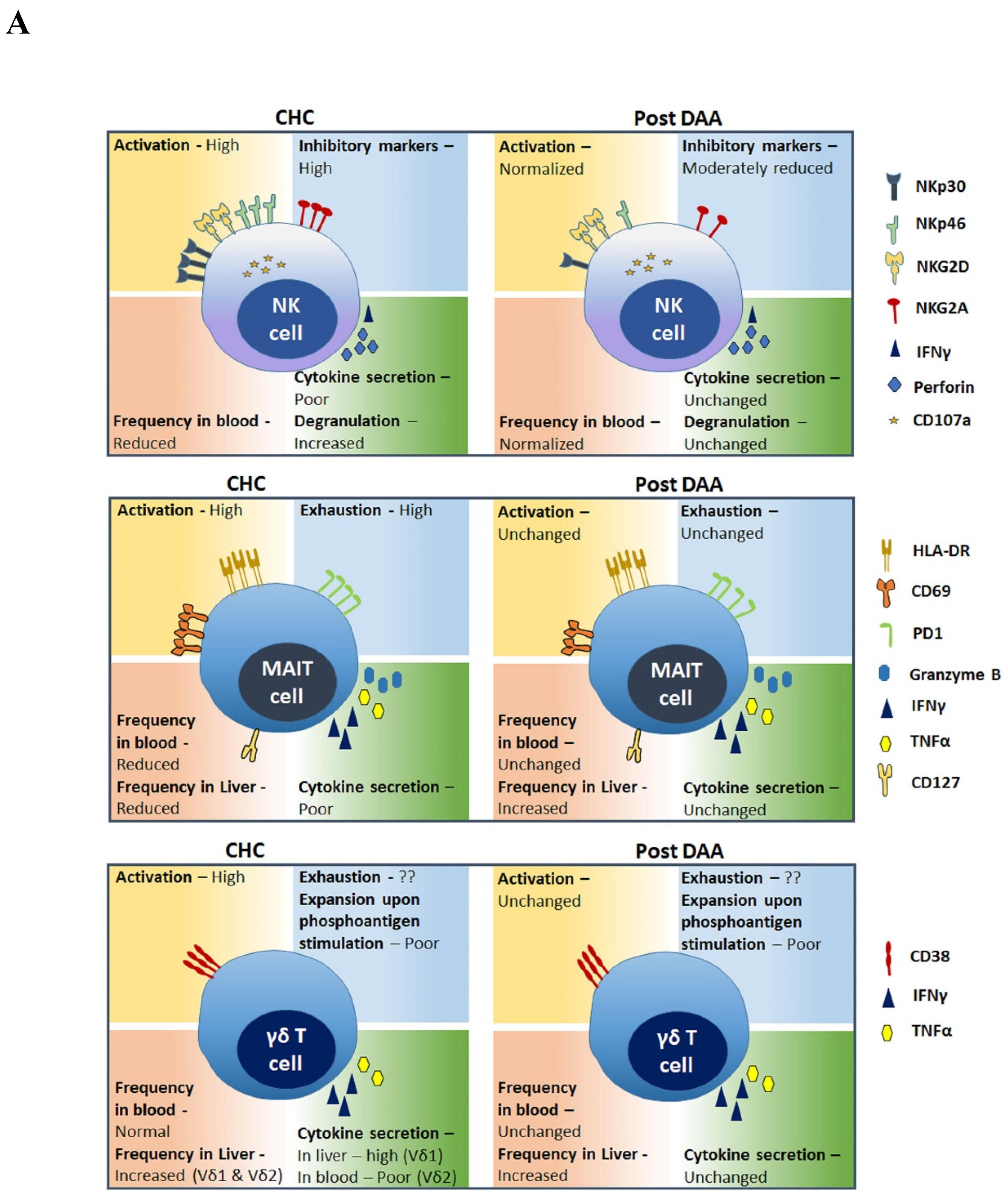{kind=link}
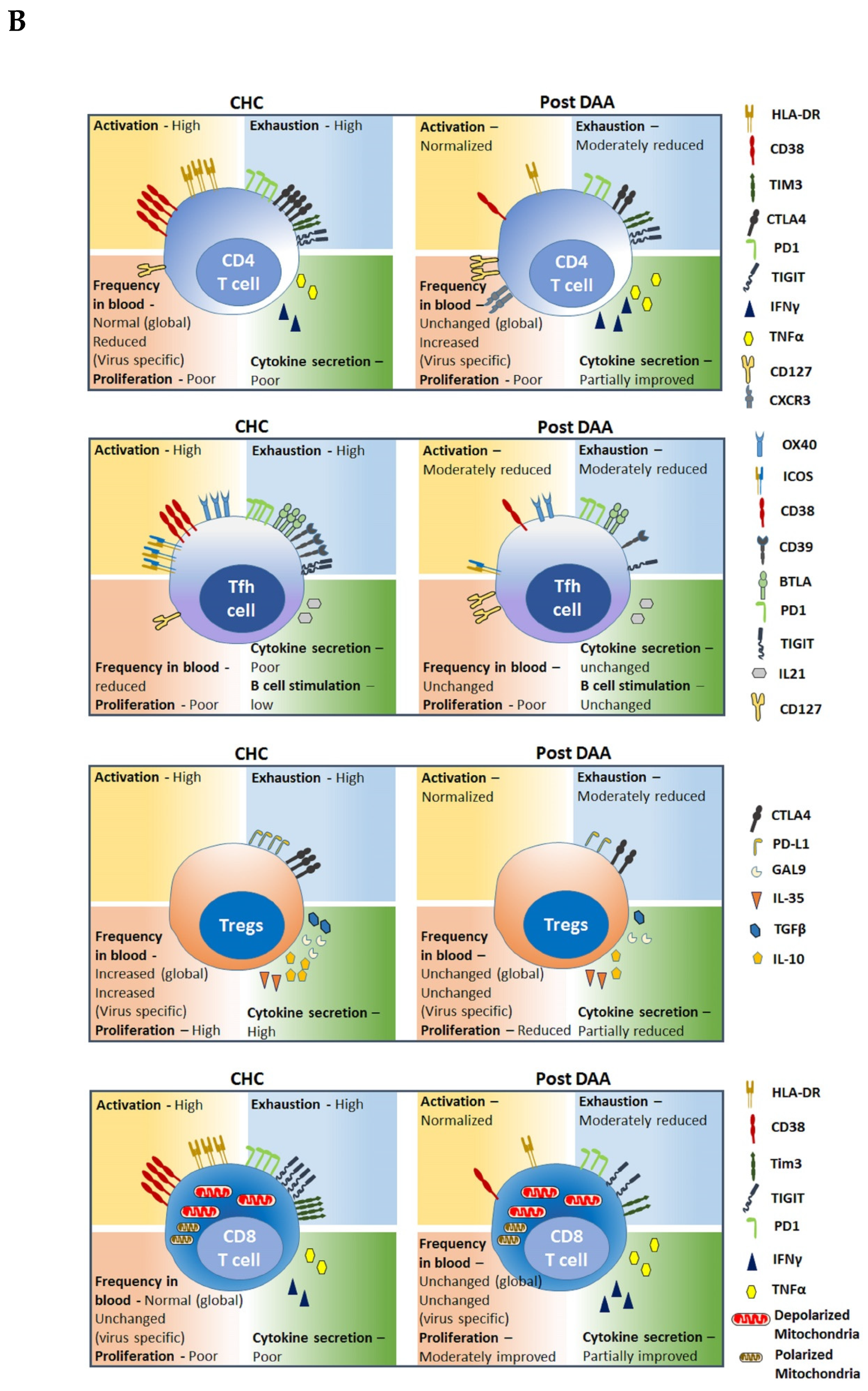{kind=link}
Table 1.
Lymphocyte landscape present in a chronic hepatitis C virus (HCV)-infected patient and its status after virus clearance using direct acting antiviral (DAA) therapy.
Table 1.
Lymphocyte landscape present in a chronic hepatitis C virus (HCV)-infected patient and its status after virus clearance using direct acting antiviral (DAA) therapy.
| Lymphocyte Populations | Dysfunction in CHC | Post DAA Therapy | Reference | ||||
|---|---|---|---|---|---|---|---|
| Frequency | Phenotype | Function | Frequency | Phenotype | Function | ||
| Natural Killer (NK) cells | Reduced in blood | NKG2A↑, NKp30↑, NKp46↑ | Reduced cytotoxicity, INFγ↓, CD107a ↑ | Increase in CD56dim and decrease in CD56bright | NKG2A↓, NKp30↓, NKp46↓, CD16↑, KLRG1↑, CD69↑ | Improved cytotoxicity, INFγ↑ | [15,16,17,18,19,20,21,22,23,24,25,26,27,28,29] |
| Mucosal-associated invariant T (MAIT) cells | Low in blood and liver | CD69↑, PD-1↑, HLA-DR↑, CD127↓ | Increased cytotoxicity, INFγ↓, INFα↓, GrB↑ | Low in circulation, Increase in liver | CD69↑, PD1↑, HLA-DR↑, CD127↓ | Increased cytotoxicity, INFγ↓, INFα↓, GrB↑ | [30,31,32,33,34,35,36,37] |
| Gamma detes (γδ) T cells | |||||||
| Vδ1 | Increased in liver | CD161↑ | Increased cytotoxicity, INFγ↑, INFα↑, | ND | ND | ND | [38,39,40] |
| Vδ2 | Unchanged in blood, Increased in liver | CD56↑, CD16↑, PD-1↑ | Poor proliferation in response to phospho-antigen stimulation, INFγ ↓ | Unchanged ih blood | ND | Poor proliferation in response to phospho-antigen stimulation, INFγ↓ | [12,41,42,43,44,45] |
| Helper T Cells | Virus-specific CD4+ T cell reduced in blood | CD38↑, HLA-DR↑, PD-1↑, CTLA-4↑, TIM-3↑, KLRG1↑, TIGIT↑, CD127↓ | INFγ↓, INFα↓ | Virus-specific CD4+ T cell increased in blood | CD38↓, HLA-DR↓, PD-1↑, CTLA-4↓, TIM-3↓, TCF1↑, TIGIT↑, CD127↑, CXCR3↑ | INFγ↓, INFα↓, IL-22↑, IL-17↑ | [46,47,48,49,50,51,52,53,54,55,56,57] |
| Follicular Helper T (Tfh) Cells | Reduced in blood | CD38↑, CD39↑, ICOS ↑, OX40↑, BTLA↑, TIGIT↑, CD127↓ | IL-21↓ | Reduced in blood | CD38↓, CD39↓, ICOS↓, OX40↓, BTLA↓, TIGIT↑, CD127↑ | IL-21↓ | [48,58,59,60] |
| T Regulatory Cells (Treg) | Increased in blood and liver | PD-1↑, CTLA-4↑, | IL-10↑, GAL9↑, IL-35↑, TGFβ↑ | Increased in blood | PD-1↑, CTLA-4↑, | IL-10↓, GAL9↓, IL-35↓, TGFβ↓ During therapy | [61,62,63,64,65,66,67,68] |
| Cytotoxic T Lymphocytes | Unchanged in blood | CD38↑, HLA-DR↑, PD-1↑, LAG-3↑, TIM-3↑, CD5↑, CD160↑, TIGIT↑, CD127↓, TOX↑ | Reduced cytotoxicity, INFγ↓, INFα↓ | Unchanged in blood | CD38↓, HLA-DR↓, PD-1↑, LAG-3 ↑, TIM-3↑, CD5↑, CD160↑, TIGIT↑, CD127↓, TOX↓ | INFγ ↑, INFα ↑ | [11,69,70,71,72,73,74,75,76,77,78,79,80] |
ND: Not defined.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ghosh, A.; Romani, S.; Kottilil, S.; Poonia, B. Lymphocyte Landscape after Chronic Hepatitis C Virus (HCV) Cure: The New Normal. Int. J. Mol. Sci. 2020, 21, 7473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207473
AMA Style
Ghosh A, Romani S, Kottilil S, Poonia B. Lymphocyte Landscape after Chronic Hepatitis C Virus (HCV) Cure: The New Normal. International Journal of Molecular Sciences. 2020; 21(20):7473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207473
Chicago/Turabian StyleGhosh, Alip, Sara Romani, Shyam Kottilil, and Bhawna Poonia. 2020. "Lymphocyte Landscape after Chronic Hepatitis C Virus (HCV) Cure: The New Normal" International Journal of Molecular Sciences 21, no. 20: 7473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207473
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.