1-Cinnamoyltrichilinin from Melia azedarach Causes Apoptosis through the p38 MAPK Pathway in HL-60 Human Leukemia Cells

 , , , and
, , , and 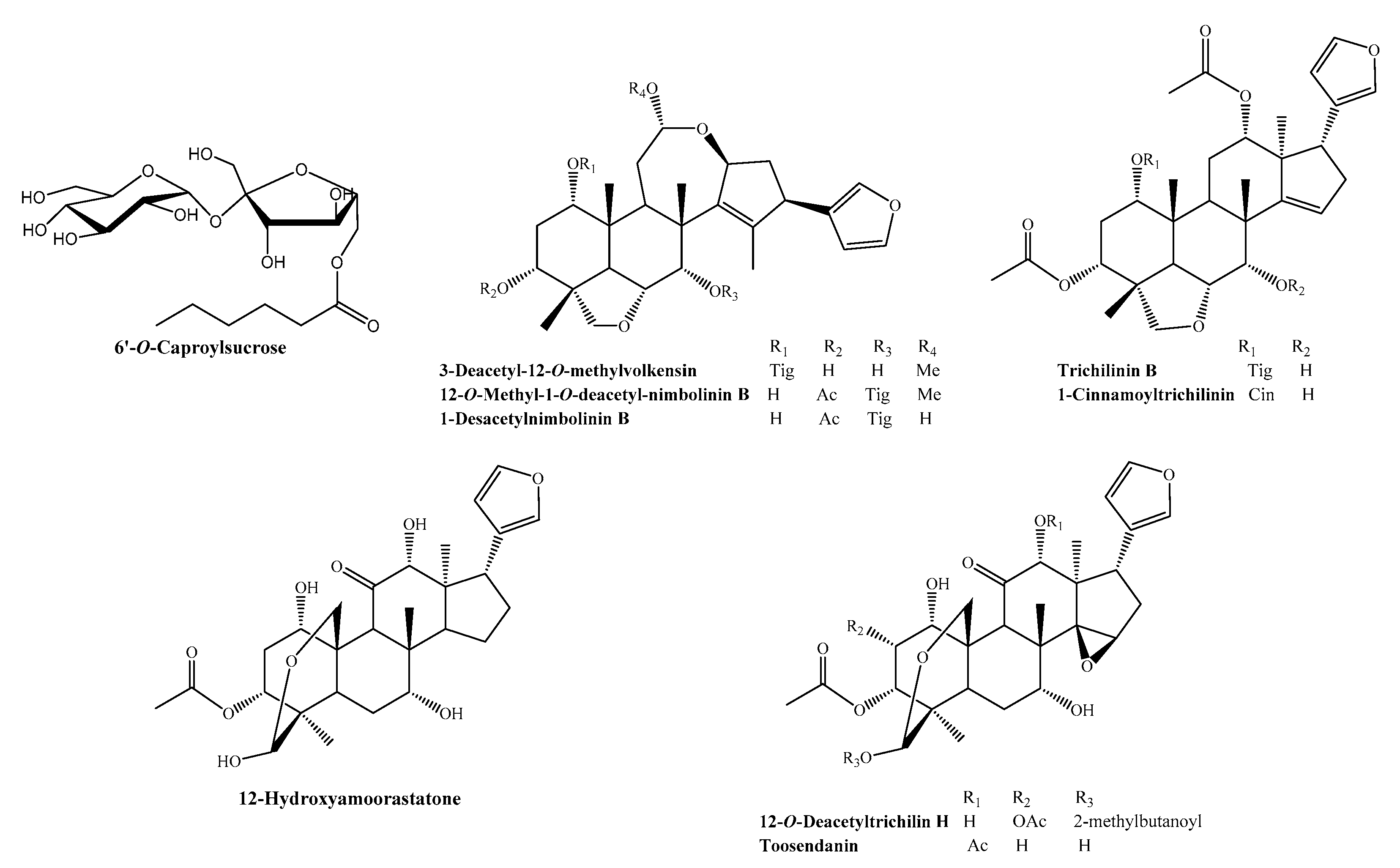{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
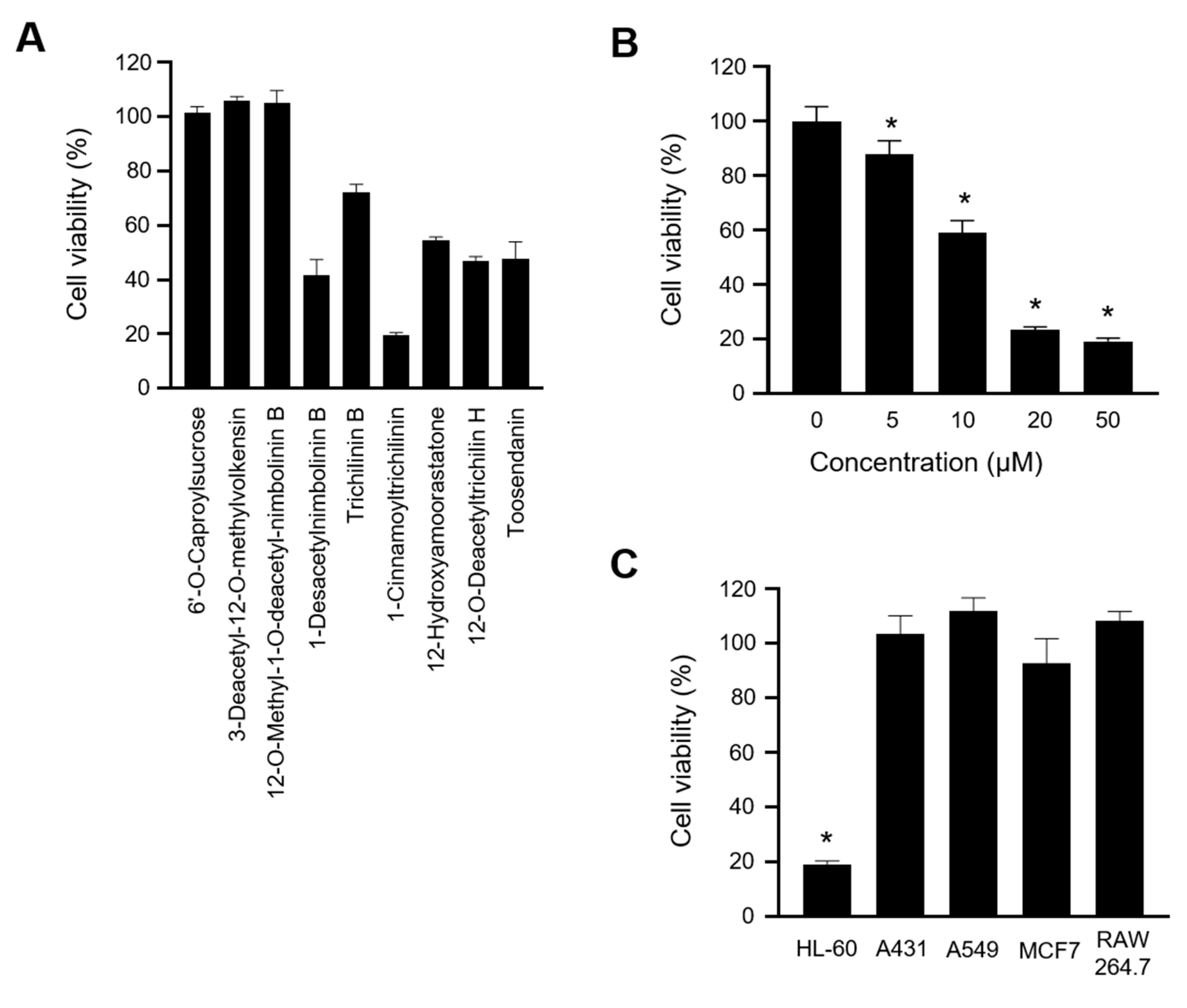
2.1. 1-Cinnamoyltrichilinin (CT) Suppressed the Viability of HL-60 Cells
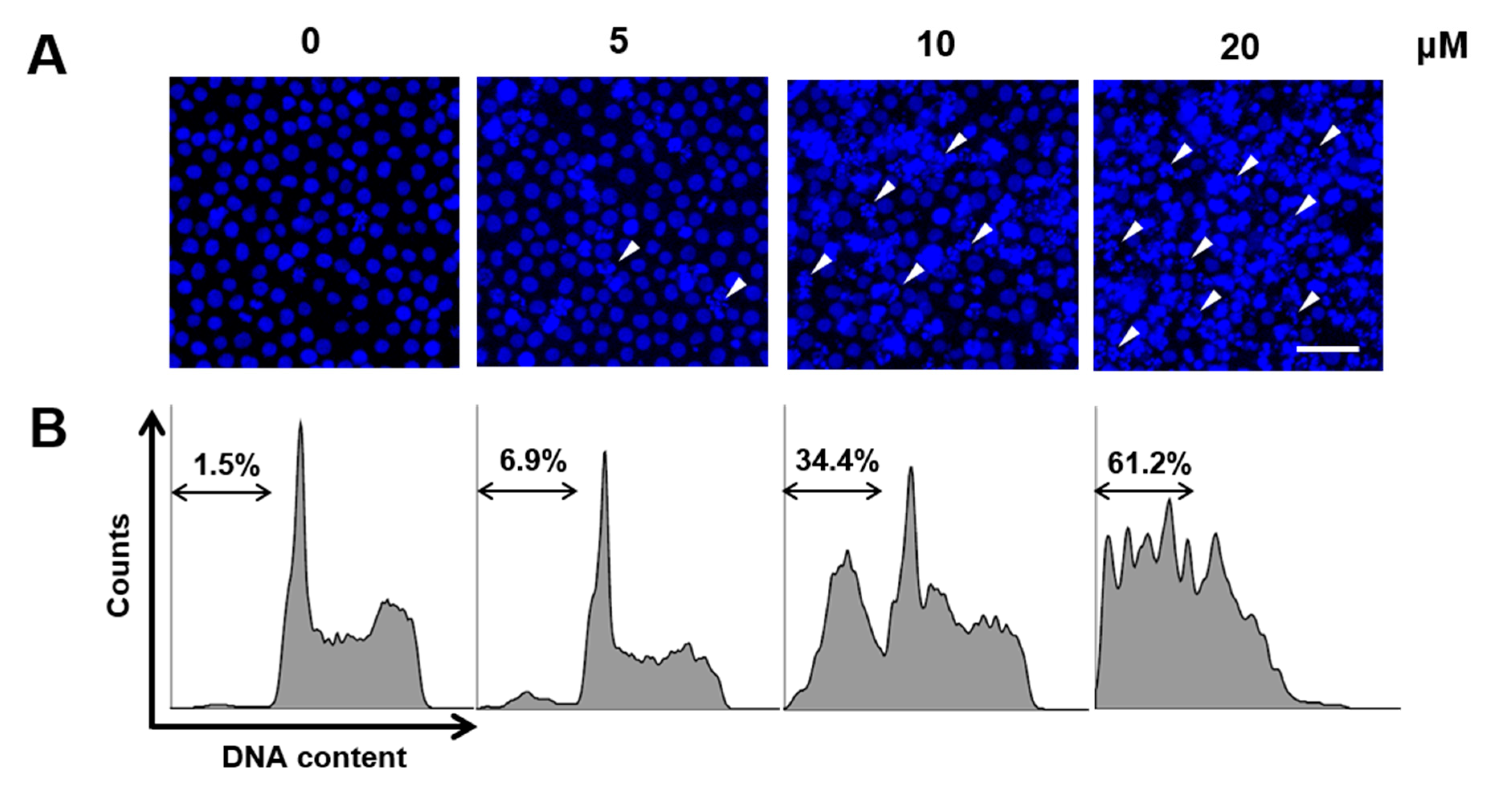
2.2. CT Induced Apoptosis in HL-60 Cells
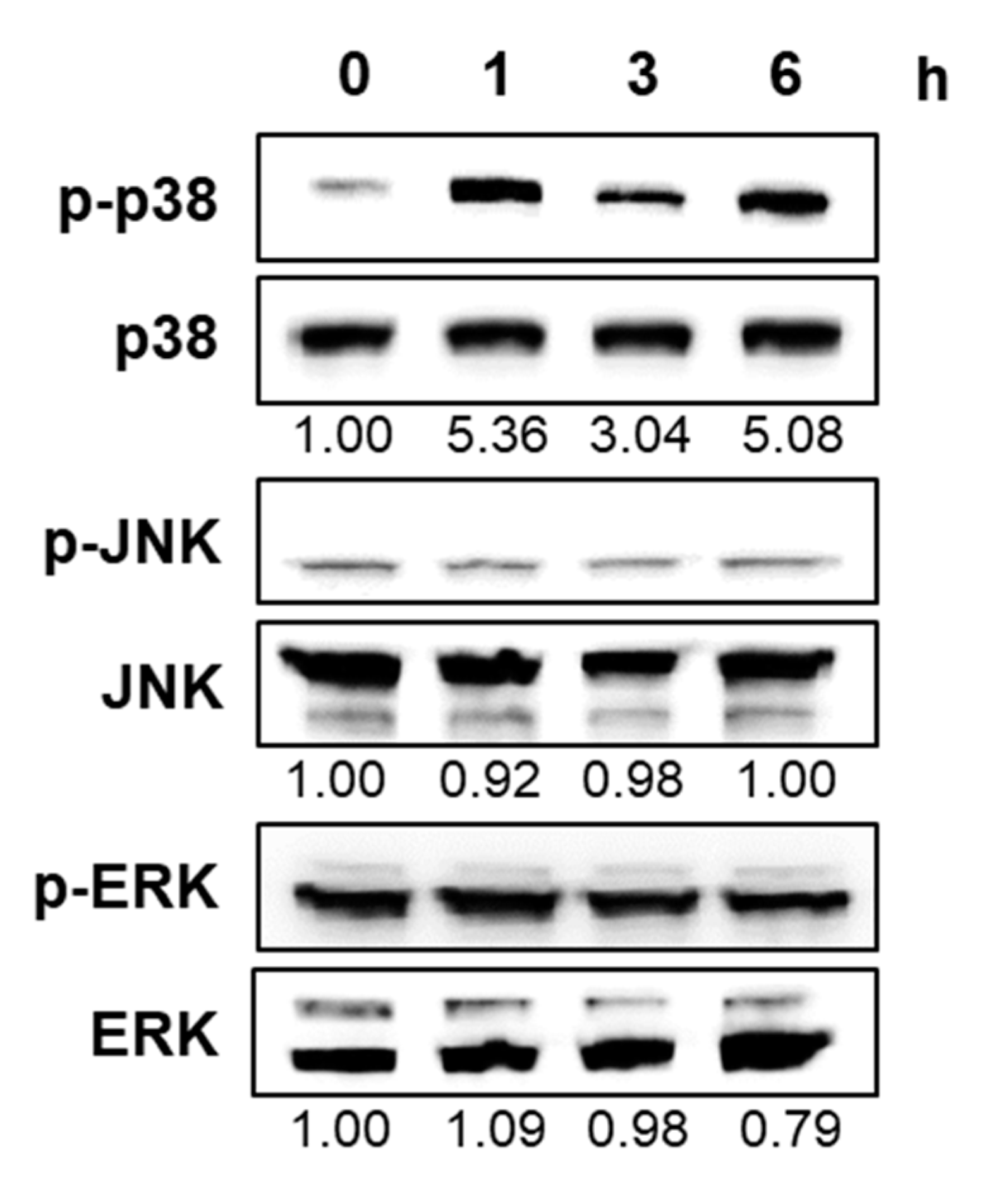
2.3. CT Activated p38 Mitogen-Activated Protein Kinase in HL-60 Cells
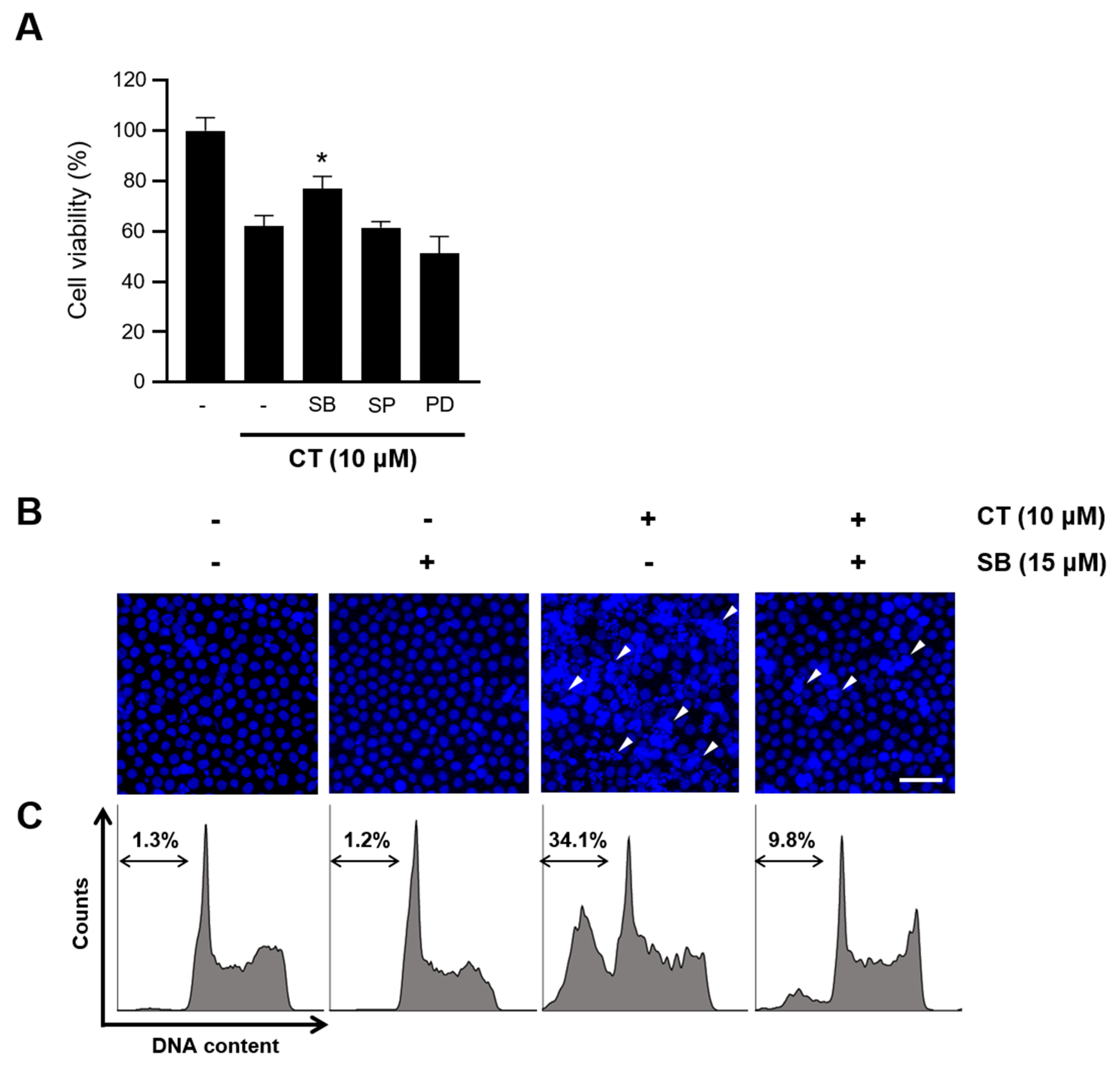
2.4. CT-Induced Apoptosis Was Prevented by p38 Inhibition
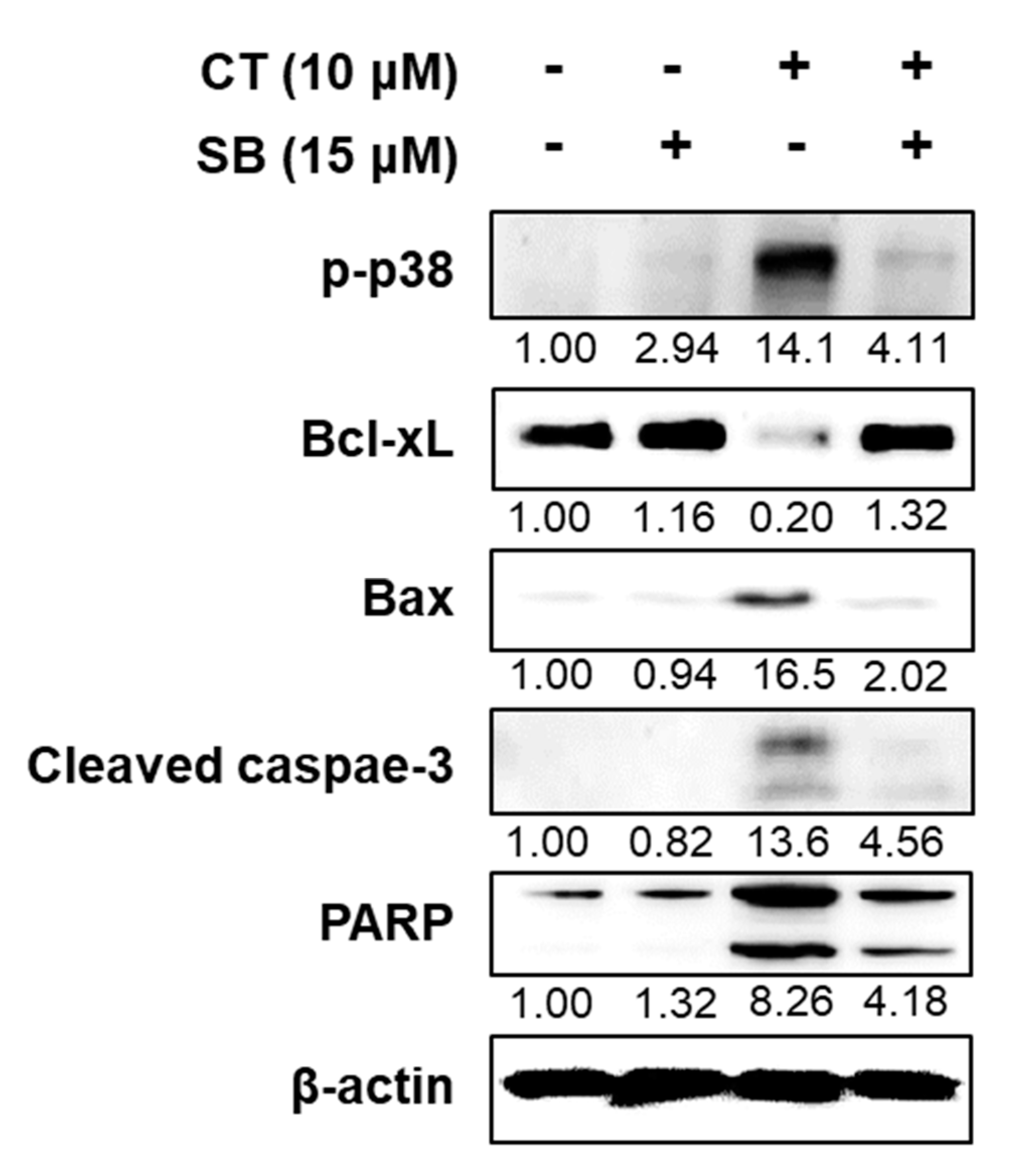
2.5. Inhibition of p38 Improved HL-60 Viability by Regulating Apoptotic Factors
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Material
4.3. Extraction and Isolation
4.4. Cell Culture and Materials
4.5. Cell Growth Inhibition Assays
4.6. Nuclear Staining with Hoechst 33342
4.7. Sub-G1 Population Measurement by Flow Cytometry
4.8. Western Blot
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AML | Acute myeloid leukemia |
| CT | 1-Cinnamoyltrichilinin |
| HR-ESI-MS | High-resolution electrospray ionization mass spectroscopy |
| NMR | Nuclear magnetic resonance |
| MAPK | Mitogen-activated protein kinase |
| JNK | c-Jun-N-terminal kinase |
| ERK | Extracellular signal-regulated kinase |
| PARP | Poly (ADP-ribose) polymerase |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| PI | Propidium iodide |
| CC | Column chromatography |
| HPLC | High-performance liquid chromatography |
References
- Döhner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute Myeloid Leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corces, M.R.; Chang, H.Y.; Majeti, R. Preleukemic Hematopoietic Stem Cells in Human Acute Myeloid Leukemia. Front. Oncol. 2017, 7, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kouchkovsky, I.; Abdul-Hay, M. ‘Acute myeloid leukemia: A comprehensive review and 2016 update’. Blood Cancer J. 2016, 6, e441. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Luo, Y.; Tian, M.; Zhang, S.-Y.; Lu, R.; Wang, J.-H.; Kasimu, R.; Li, X. Plant natural products: From traditional compounds to new emerging drugs in cancer therapy. Cell Prolif. 2014, 47, 506–515. [Google Scholar] [CrossRef]
- Demain, A.L.; Vaishnav, P. Natural products for cancer chemotherapy. Microb. Biotechnol. 2010, 4, 687–699. [Google Scholar] [CrossRef] [Green Version]
- Hanan, N.A.; Chiu, H.I.; Ramachandran, M.R.; Tung, W.H.; Zain, N.; Yahaya, N.; Lim, V. Cytotoxicity of Plant-Mediated Synthesis of Metallic Nanoparticles: A Systematic Review. Int. J. Mol. Sci. 2018, 19, 1725. [Google Scholar] [CrossRef] [Green Version]
- Della Bona, A.; Nedel, F. Evaluation of Melia azedarach Extracts Against Streptococcus mutans. J. Med. Food 2015, 18, 259–263. [Google Scholar] [CrossRef]
- Marimuthu, S.; Balakrishnan, P.; Nair, S. Phytochemical investigation and radical scavenging activities ofMelia azedarachand its DNA protective effect in cultured lymphocytes. Pharm. Biol. 2013, 51, 1331–1340. [Google Scholar] [CrossRef]
- Nerome, K.; Ito-Kureha, T.; Paganini, T.; Fukuda, T.; Igarashi, Y.; Ashitomi, H.; Ikematsu, S.; Yamamoto, T. Potent and broad anticancer activities of leaf extracts from Melia azedarach L. of the subtropical Okinawa islands. Am. J. Cancer Res. 2020, 10, 581–594. [Google Scholar]
- Malar, T.J.; Antonyswamy, J.; Vijayaraghavan, P.; Kim, Y.O.; Al-Ghamdi, A.A.; Elshikh, M.S.; Hatamleh, A.A.; Al-Dosary, M.A.; Na, S.W.; Kim, H.-J. In-vitro phytochemical and pharmacological bio-efficacy studies on Azadirachta indica A. Juss and Melia azedarach Linn for anticancer activity. Saudi J. Biol. Sci. 2020, 27, 682–688. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Hotz, M.A.; Gong, J.; Traganos, F.; Darzynkiewicz, Z. Flow cytometric detection of apoptosis: Comparison of the assays of in situ DNA degradation and chromatin changes. Cytometry 1994, 15, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.; López, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, R.; Chaudhry, G.-E.-S. Understanding Apoptosis and Apoptotic Pathways Targeted Cancer Therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualdani, R.; Cavalluzzi, M.M.; Lentini, G.; Habtemariam, S. The Chemistry and Pharmacology of Citrus Limonoids. Molecules 2016, 21, 1530. [Google Scholar] [CrossRef]
- Poulose, S.M.; Harris, E.D.; Patil, B.S. Citrus Limonoids Induce Apoptosis in Human Neuroblastoma Cells and Have Radical Scavenging Activity. J. Nutr. 2005, 135, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, M.F.; Hamdan, D.I.; Wink, M.; El-Shazly, A.M. Hepatoprotective effect of limonin, a natural limonoid from the seed of Citrus aurantium var. bigaradia, on D-galactosamine-induced liver injury in rats. Naunyn Schmiedebergs Arch. Pharmacol. 2013, 387, 251–261. [Google Scholar] [CrossRef]
- Wang, N.; Fan, Y.; Yuan, C.M.; Song, J.; Yao, Y.; Liu, W.; Gajendran, B.; Zacksenhaus, E.; Li, Y.; Liu, J.; et al. Selective ERK1/2 agonists isolated from Melia azedarach with potent anti-leukemic activity. BMC Cancer 2019, 19, 764. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.; Tait, S.W.G. Mitochondrial apoptosis: Killing cancer using the enemy within. Br. J. Cancer 2015, 112, 957–962. [Google Scholar] [CrossRef] [Green Version]
- Wada, T.; Penninger, J.M. Mitogen-activated protein kinases in apoptosis regulation. Oncogene 2004, 23, 2838–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Rauch, J.; Kolch, W. Targeting MAPK Signaling in Cancer: Mechanisms of Drug Resistance and Sensitivity. Int. J. Mol. Sci. 2020, 21, 1102. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-E.; Du, F.; Fang, M.; Wang, X. Formation of apoptosome is initiated by cytochrome c-induced dATP hydrolysis and subsequent nucleotide exchange on Apaf-1. Proc. Natl. Acad. Sci. USA 2005, 102, 17545–17550. [Google Scholar] [CrossRef] [Green Version]
- Kraus, W.; Bokel, M. Neue Tetranortriterpenoide aus Melia azedarach Linn. (Meliaceae). Ber. Dtsch. Chem. Ges. 1981, 114, 267–275. [Google Scholar] [CrossRef]
- Ahn, J.-W.; Choi, S.-U.; Lee, C.-O. Cytotoxic limonoids from Melia azedarach var. Japonica. Phytochemistry 1994, 36, 1493–1496. [Google Scholar] [CrossRef]
- Zhou, H.; Hamazaki, A.; Fontana, J.D.; Takahashi, H.; Wandscheer, C.B.; Fukuyama, Y. Cytotoxic Limonoids from Brazilian Melia azedarach. Chem. Pharm. Bull. 2005, 53, 1362–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.-F.; Fan, H.; Wang, L.-J.; Wu, S.-B.; Zhao, Y. Limonoids from the fruits of Melia toosendan. Phytochem. Lett. 2011, 4, 292–297. [Google Scholar] [CrossRef]
- Zhu, G.-Y.; Bai, L.-P.; Liu, L.; Jiang, Z.-H. Limonoids from the fruits of Melia toosendan and their NF-κB modulating activities. Phytochemistry 2014, 107, 175–181. [Google Scholar] [CrossRef]
- Park, S.; Nhiem, N.X.; Subedi, L.; Oh, I.; Kim, J.-Y.; Kim, S.Y.; Kim, S.H. Isolation of bioactive limonoids from the fruits of Melia azedarach. J. Asian Nat. Prod. Res. 2019, 22, 1–9. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.; Park, S.; Kim, S.-Y.; Cho, S.-H.; Jeong, M.S.; Kim, S.-R.; Seo, J.B.; Kim, S.H.; Kim, K.-N. 1-Cinnamoyltrichilinin from Melia azedarach Causes Apoptosis through the p38 MAPK Pathway in HL-60 Human Leukemia Cells. Int. J. Mol. Sci. 2020, 21, 7506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207506
Jeong H, Park S, Kim S-Y, Cho S-H, Jeong MS, Kim S-R, Seo JB, Kim SH, Kim K-N. 1-Cinnamoyltrichilinin from Melia azedarach Causes Apoptosis through the p38 MAPK Pathway in HL-60 Human Leukemia Cells. International Journal of Molecular Sciences. 2020; 21(20):7506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207506
Chicago/Turabian StyleJeong, Hoibin, SeonJu Park, Seo-Young Kim, Su-Hyeon Cho, Myeong Seon Jeong, Song-Rae Kim, Jong Bok Seo, Seung Hyun Kim, and Kil-Nam Kim. 2020. "1-Cinnamoyltrichilinin from Melia azedarach Causes Apoptosis through the p38 MAPK Pathway in HL-60 Human Leukemia Cells" International Journal of Molecular Sciences 21, no. 20: 7506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207506