RNAi-Mediated Knockdown of Transcription Factor E93 in Nymphs of the Desert Locust (Schistocerca gregaria) Inhibits Adult Morphogenesis and Results in Supernumerary Juvenile Stages

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Identification and Phylogenetic Analysis of SgE93
2.2. Knockdown of SgE93 Resulted in Supernumerary Nymphal Stages
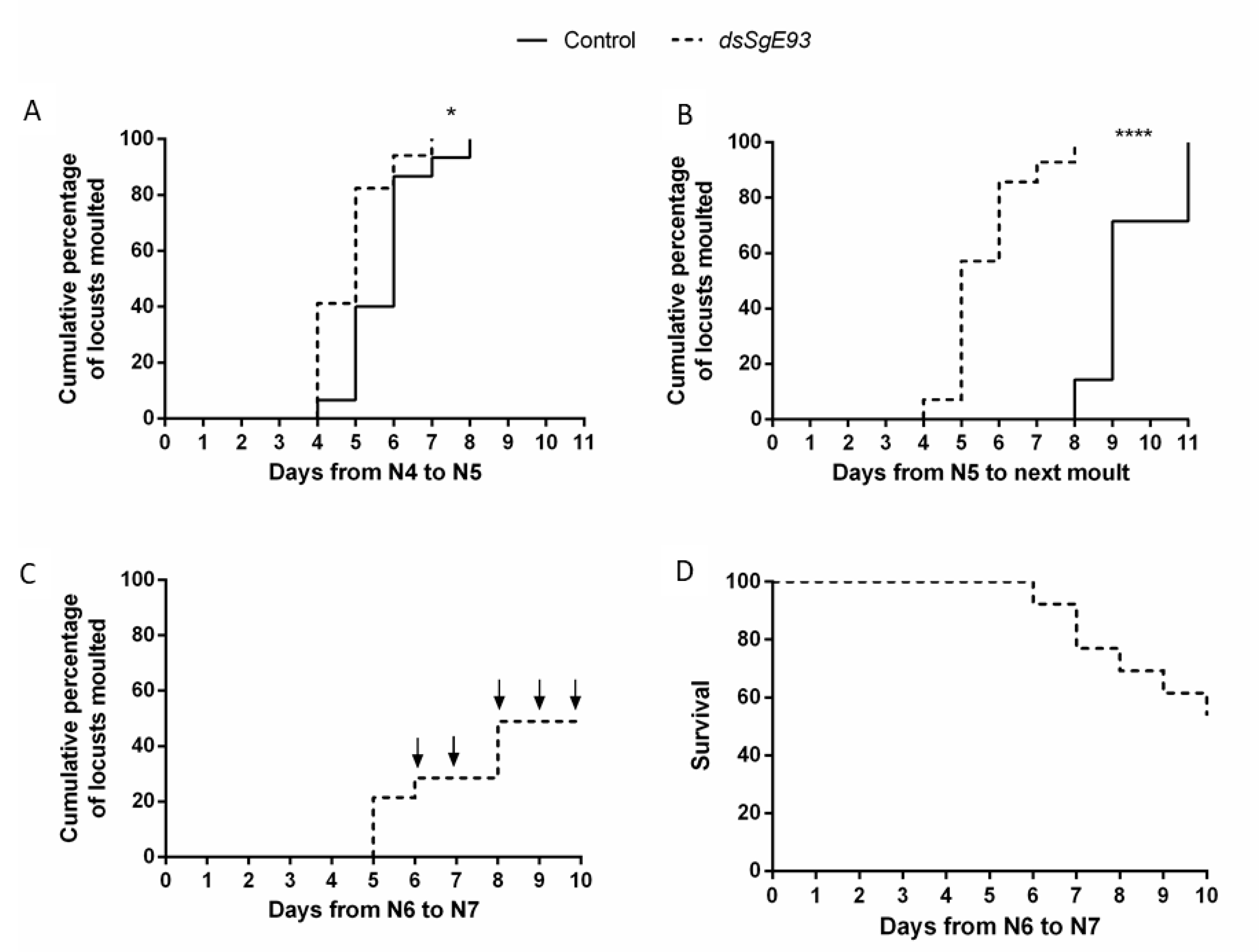
2.3. SgE93 Silencing Altered the Duration of the Molting Cycle and Affected the Success of Molting to Later Nymphal Stages
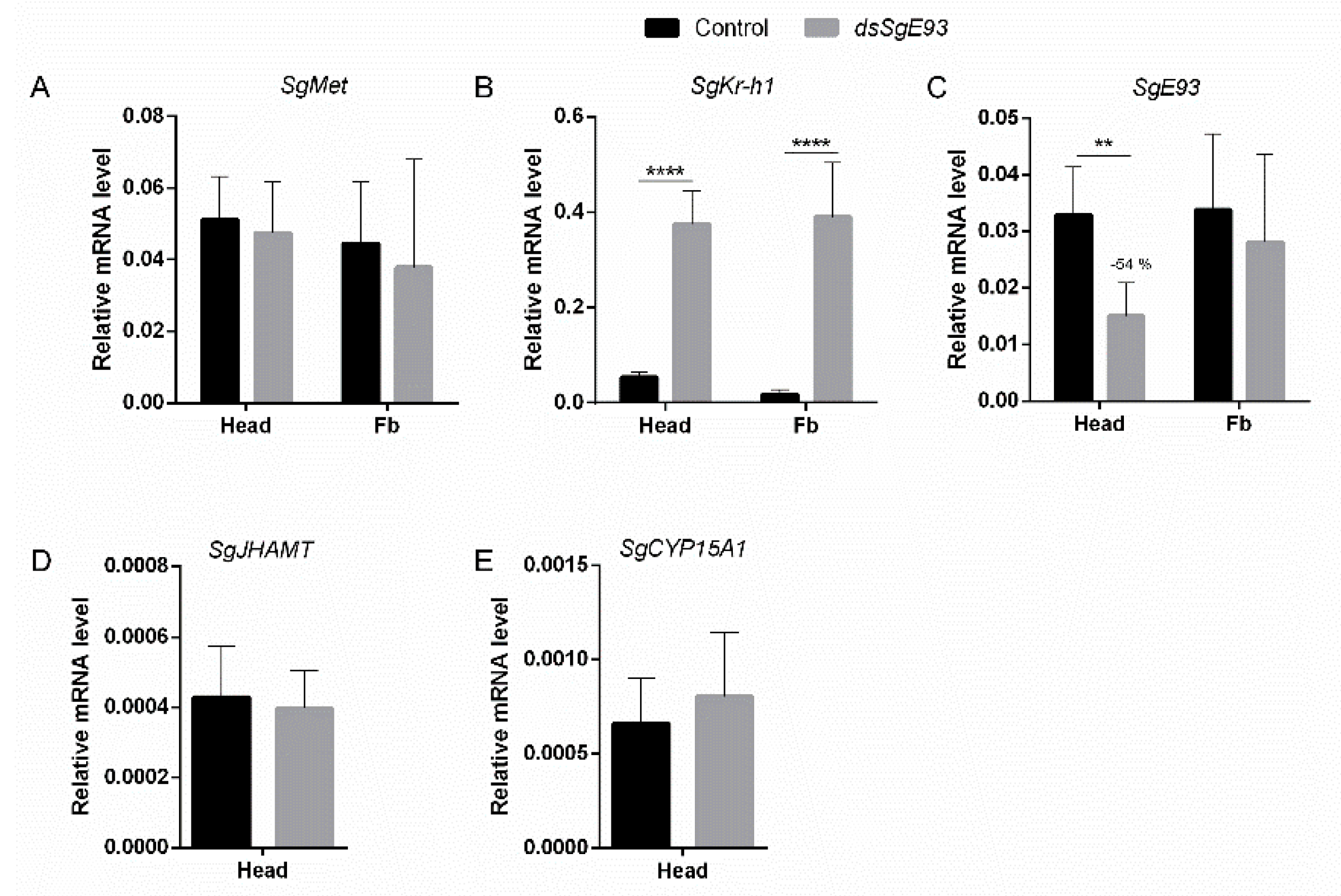
2.4. Transcript Levels of MEKRE93 and JH Biosynthesis Pathway Components in dsSgE93- and dsGFP-Injected Locusts
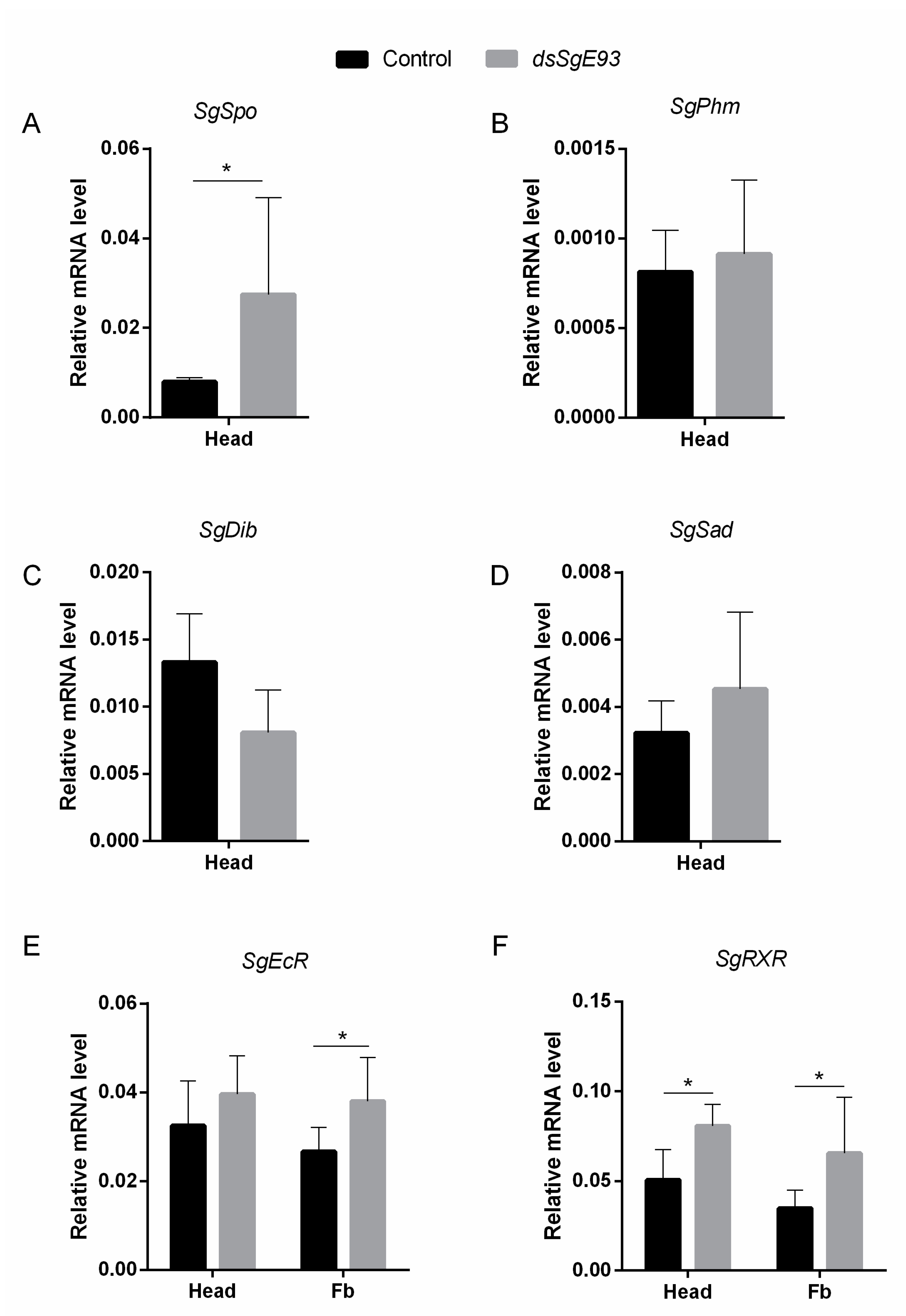
2.5. Transcript Levels of Different Halloween Genes and Components of the Ecdysone Receptor Complex in dsSgE93- and dsGFP-Injected Locusts
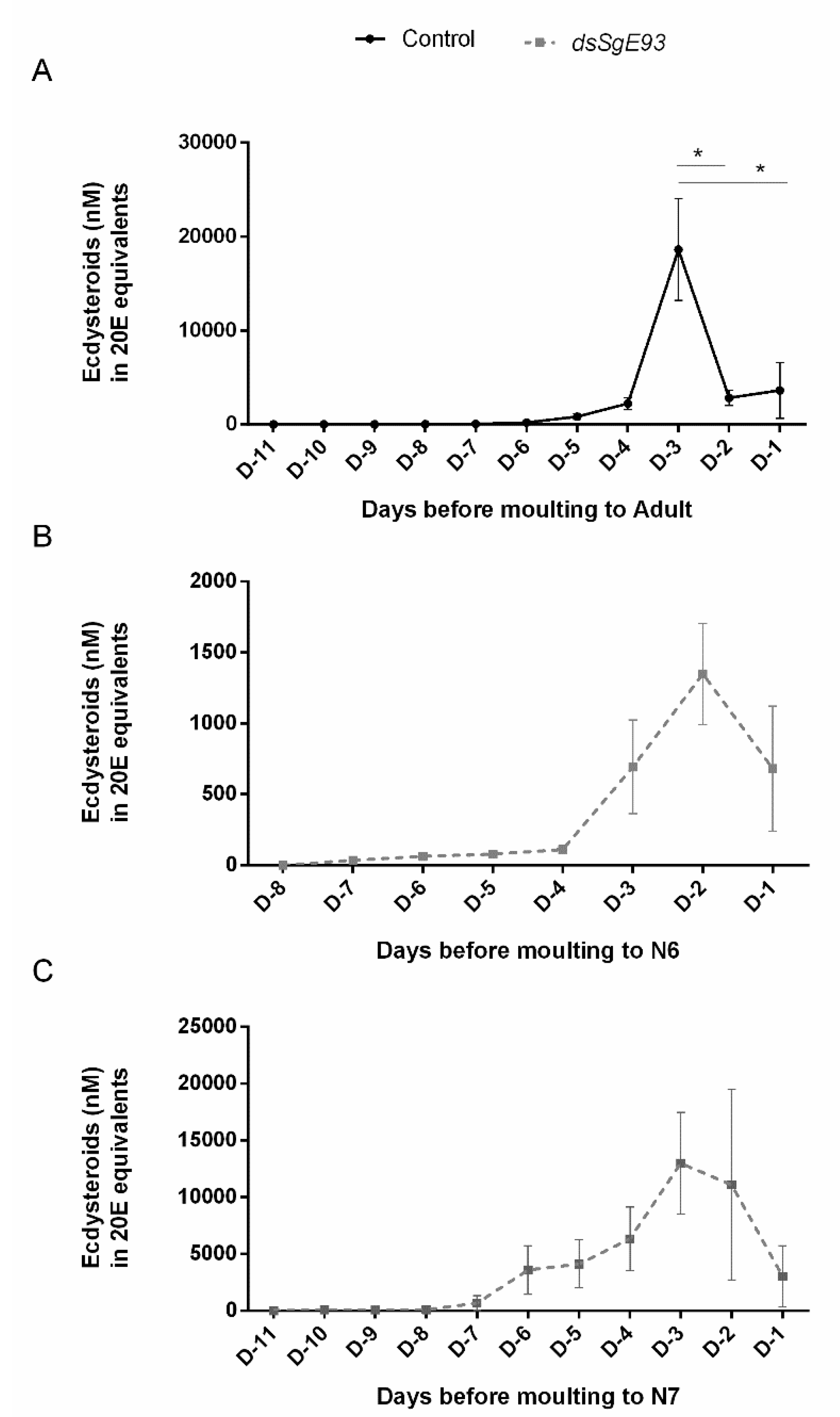
2.6. Temporal Profiles of Ecdysteroids in Hemolymph of dsSgE93- and dsGFP-Injected Locusts
3. Discussion
3.1. SgE93 is the Desert Locust E93 Ortholog
3.2. SgE93 Knockdown has Inhibited Metamorphosis and Adult Morphogenesis
3.3. SgE93 Knockdown has Influenced the Ecdysteroid Pathway
4. Materials and Methods
4.1. Rearing of Animals
4.2. Tissue Collection
4.3. RNA Extraction and cDNA Synthesis
4.4. Multispecies E93 Gene Identification and Phylogenetic Analysis
4.5. Quantitative Real-Time PCR
4.6. RNA Interference Experiments
4.7. Ecdysteroid Measurement
4.8. Observing Ecdysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 20E | 20-hydroxyecdysone |
| Ad | Adult |
| bHLH | Basic-helix-loop-helix |
| CA | Corpora allata |
| cDNA | Complementary DNA |
| CtBP-im | C-terminal-binding protein interaction motif |
| CYP15A1 | MF epoxidase |
| D | Day |
| Dib | Disembodied |
| DNA | Deoxyribonucleic acid |
| ds | Double-stranded |
| dsRNA | Double-stranded RNA |
| E93 | Ecdysone-induced protein 93 |
| EcR | Ecdysone receptor |
| EF1α | Elongation factor 1α |
| EIA | Enzyme ImmuoAssay |
| Fb | Fat body |
| GFP | Green fluorescent protein |
| HTH | Helix-turn-helix |
| JH | Juvenile hormone |
| JHAMT | Juvenile hormone acid methyl transferase |
| Kr-h1 | Krüppel-homolog 1 |
| MEKRE93 | Methoprene tolerant-Krüppel homolog 1-E93 |
| Met | Methoprene-tolerant |
| mRNA | Messenger RNA |
| N | Nymph |
| NCBI | National Center for Biotechnology Information |
| NR | Nuclear receptor |
| PAS | Per-Arnt-Sim |
| PCR | Polymerase chain reaction |
| Phm | Phantom |
| qRT-PCR | Quantitative real-time PCR |
| RNA | Ribonucleic acid |
| RNAi | RNA interference |
| RXR | Retinoid-X-receptor |
| Sad | Shadow |
| Shd | Shade |
| Spo | Spook |
| Tai | Taiman |
| USP | Ultraspiracle |
References
- Riddiford, L.M. How does juvenile hormone control insect metamorphosis and reproduction? Gen. Comp. Endocrinol. 2012, 179, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.J.; Billas, I.M.; Bonneton, F.; Graham, L.D.; Lawrence, M.C. Ecdysone Receptors: From the Ashburner Model to Structural Biology. Annu. Rev. Èntomol. 2013, 58, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.-P.; Iwema, T.; Epa, V.C.; Takaki, K.; Rynes, J.; Jindra, M. Ligand-binding properties of a juvenile hormone receptor, Methoprene-tolerant. Proc. Natl. Acad. Sci. USA 2011, 108, 21128–21133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, T.G.; Fabian, J. A Drosophila melanogaster mutant resistant to a chemical analog of juvenile hormone. Dev. Biol. 1986, 118, 190–201. [Google Scholar] [CrossRef]
- Godlewski, J.; Wang, S.; Wilson, T.G. Interaction of bHLH-PAS proteins involved in juvenile hormone reception in Drosophila. Biochem. Biophys. Res. Commun. 2006, 342, 1305–1311. [Google Scholar] [CrossRef]
- Song, J.; Wu, Z.; Wang, Z.; Deng, S.; Zhou, S. Krüppel-homolog 1 mediates juvenile hormone action to promote vitellogenesis and oocyte maturation in the migratory locust. Insect Biochem. Mol. Biol. 2014, 52, 94–101. [Google Scholar] [CrossRef]
- Minakuchi, C.; Namiki, T.; Shinoda, T. Krüppel homolog 1, an early juvenile hormone-response gene downstream of Methoprene-tolerant, mediates its anti-metamorphic action in the red flour beetle Tribolium castaneum. Dev. Biol. 2009, 325, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Gujar, H.; Palli, S.R. Krüppel homolog 1 and E93 mediate Juvenile hormone regulation of metamorphosis in the common bed bug, Cimex lectularius. Sci. Rep. 2016, 6, 26092. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Tomonari, S.; Watanabe, T.; Noji, S.; Mito, T. Regulatory mechanisms underlying the specification of the pupal-homologous stage in a hemimetabolous insect. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190225. [Google Scholar] [CrossRef] [Green Version]
- Konopova, B.; Smykal, V.; Jindra, M. Common and Distinct Roles of Juvenile Hormone Signaling Genes in Metamorphosis of Holometabolous and Hemimetabolous Insects. PLoS ONE 2011, 6, e28728. [Google Scholar] [CrossRef] [Green Version]
- Li, K.L.; Yuan, S.Y.; Nanda, S.; Wang, W.X.; Lai, F.X.; Fu, Q.; Wan, P.J. The Roles of E93 and Kr-h1 in Metamorphosis of Nilaparvata lugens. Front. Physiol. 2018, 9, 1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, J.; Belles, X. Conserved repressive function of Krüppel homolog 1 on insect metamorphosis in hemimetabolous and holometabolous species. Sci. Rep. 2011, 1, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smykal, V.; Bajgar, A.; Provaznik, J.; Fexova, S.; Buricova, M.; Takaki, K.; Hodkova, M.; Jindra, M.; Dolezel, D. Juvenile hormone signaling during reproduction and development of the linden bug, Pyrrhocoris apterus. Insect Biochem. Mol. Biol. 2014, 45, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Chafino, S.; Ureña, E.; Casanova, J.; Casacuberta, E.; Franch-Marro, X.; Martín, D. Upregulation of E93 Gene Expression Acts as the Trigger for Metamorphosis Independently of the Threshold Size in the Beetle Tribolium castaneum. Cell Rep. 2019, 27, 1039–1049.e2. [Google Scholar] [CrossRef] [PubMed]
- Konopova, B.; Jindra, M. Juvenile hormone resistance gene Methoprene-tolerant controls entry into metamorphosis in the beetle Tribolium castaneum. Proc. Natl. Acad. Sci. USA 2007, 104, 10488–10493. [Google Scholar] [CrossRef] [Green Version]
- Lozano, J.; Belles, X. Role of Methoprene-Tolerant (Met) in Adult Morphogenesis and in Adult Ecdysis of Blattella germanica. PLoS ONE 2014, 9, e103614. [Google Scholar] [CrossRef] [Green Version]
- Gijbels, M.; Schellens, S.; Schellekens, T.; Bruyninckx, E.; Marchal, E.; Vanden Broeck, J. Precocious Downregulation of Krüppel-Homolog 1 in the Migratory Locust, Locusta migratoria, Gives Rise to An Adultoid Phenotype with Accelerated Ovarian Development but Disturbed Mating and Oviposition. Int. J. Mol. Sci. 2020, 21, 6058. [Google Scholar] [CrossRef]
- Baehrecke, E.H.; Thummel, C.S. The Drosophila E93 Gene from the 93F Early Puff Displays Stage- and Tissue-Specific Regulation by 20-Hydroxyecdysone. Dev. Biol. 1995, 171, 85–97. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Wendel, D.P.; Reid, P.; Lam, G.; Thummel, C.S.; Baehrecke, E.H. E93 Directs Steroid-Triggered Programmed Cell Death in Drosophila. Mol. Cell 2000, 6, 433–443. [Google Scholar] [CrossRef]
- Mou, X.; Duncan, D.M.; Baehrecke, E.H.; Duncan, I. Control of target gene specificity during metamorphosis by the steroid response gene E. Proc. Natl. Acad. Sci. USA 2012, 109, 2949–2954. [Google Scholar] [CrossRef] [Green Version]
- Ureña, E.; Manjón, C.; Franch-Marro, X.; Martín, D. Transcription factor E93 specifies adult metamorphosis in hemimetabolous and holometabolous insects. Proc. Natl. Acad. Sci. USA 2014, 111, 7024–7029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belles, X.; Santos, C.G. The MEKRE93 (Methoprene tolerant-Krüppel homolog 1-E93) pathway in the regulation of insect metamorphosis, and the homology of the pupal stage. Insect Biochem. Mol. Biol. 2014, 52, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Belles, X. Krüppel homolog 1 and E93: The doorkeeper and the key to insect metamorphosis. Arch. Insect Biochem. Physiol. 2019, 103, e21609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ureña, E.; Chafino, S.; Manjón, C.; Franch-Marro, X.; Martín, D. The Occurrence of the Holometabolous Pupal Stage Requires the Interaction between E93, Krüppel-Homolog 1 and Broad-Complex. PLoS Genet. 2016, 12, e1006020. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, R.; Tanaka, S.; Shiotsuki, T. RNAi-mediated knockdown of SPOOK reduces ecdysteroid titers and causes precocious metamorphosis in the desert locust Schistocerca gregaria. Dev. Biol. 2017, 429, 71–80. [Google Scholar] [CrossRef]
- Verlinden, H.; Sterck, L.; Li, J.; Li, Z.; Yssel, A.; Gansemans, Y.; Verdonck, R.; Holtof, M.; Song, H.; Behmer, S.T.; et al. First draft genome assembly of the desert locust, Schistocerca gregaria. F1000Research 2020, 9, 775. [Google Scholar] [CrossRef]
- Marchal, E.; Badisco, L.; Verlinden, H.; Vandersmissen, T.; Van Soest, S.; Van Wielendaele, P.; Vanden Broeck, J. Role of the Halloween genes, Spook and Phantom in ecdysteroidogenesis in the desert locust, Schistocerca gregaria. J. Insect Physiol. 2011, 57, 1240–1248. [Google Scholar] [CrossRef]
- Marchal, E.; Verlinden, H.; Badisco, L.; Van Wielendaele, P.; Vanden Broeck, J. RNAi-mediated knockdown of Shade negatively affects ecdysone-20-hydroxylation in the desert locust, Schistocerca gregaria. J. Insect Physiol. 2012, 58, 890–896. [Google Scholar] [CrossRef]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L.; Tian, L.; Cao, Y.; Zhang, G.; Palli, S.R.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E93 Modulates 20E Signaling to Promote Bombyx Larval-Pupal Metamorphosis. J. Biol. Chem. 2015, 290, 27370–27383. [Google Scholar] [CrossRef] [Green Version]
- Chafino, S.; López-Escardó, D.; Benelli, G.; Kovac, H.; Casacuberta, D.M.; Franch-Marro, X.; Kathirithamby, J.; Martín, D. Differential expression of the adult specifier E93 in the strepsipteran Xenos vesparum Rossi suggests a role in female neoteny. Sci. Rep. 2018, 8, 14176. [Google Scholar] [CrossRef] [Green Version]
- Minakuchi, C.; Zhou, X.; Riddiford, L.M. Krüppel homolog 1 (Kr-h1) mediates juvenile hormone action during metamorphosis of Drosophila melanogaster. Mech. Dev. 2008, 125, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smykal, V.; Daimon, T.; Kayukawa, T.; Takaki, K.; Shinoda, T.; Jindra, M. Importance of juvenile hormone signaling arises with competence of insect larvae to metamorphose. Dev. Biol. 2014, 390, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Nijhout, H. Stretch-induced moulting in Oncopeltus fasciatus. J. Insect Physiol. 1979, 25, 277–281. [Google Scholar] [CrossRef]
- Santos, C.G.; Fernandez-Nicolas, A.; Belles, X. Smads and insect hemimetabolan metamorphosis. Dev. Biol. 2016, 417, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Kamsoi, O.; Belles, X. E93-depleted adult insects preserve the prothoracic gland and molt again. bioRxiv 2020, preprint. [Google Scholar]
- De Mendonça, R.L.; Bouton, D.; Bertin, B.; Escriva, H.; Noël, C.; Vanacker, J.-M.; Cornette, J.; Laudet, V.; Pierce, R.J. A functionally conserved member of the FTZ-F1 nuclear receptor family from Schistosoma mansoni. JBIC J. Biol. Inorg. Chem. 2002, 269, 5700–5711. [Google Scholar] [CrossRef] [PubMed]
- Ohno, C.K.; Ueda, H.; Petkovich, M. The Drosophila nuclear receptors FTZ-F1 alpha and FTZ-F1 beta compete as monomers for binding to a site in the fushi tarazu gene. Mol. Cell. Biol. 1994, 14, 3166–3175. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, N.; Rewitz, K.F.; O’Connor, M.B. Ecdysone Control of Developmental Transitions: Lessons fromDrosophilaResearch. Annu. Rev. Èntomol. 2013, 58, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Ono, H.; Rewitz, K.F.; Shinoda, T.; Itoyama, K.; Petryk, A.; Rybczynski, R.; Jarcho, M.; Warren, J.T.; Marqués, G.; Shimell, M.J.; et al. Spook and Spookier code for stage-specific components of the ecdysone biosynthetic pathway in Diptera. Dev. Biol. 2006, 298, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Joly, P.; Joly, L.; Halbwachs, M. Contrôle humoral du développement chez Locusta migratoria. Ann. Sci. Nat. Zool. 1956, 18, 256–261. [Google Scholar]
- Morgan, E.; Poole, C. The pattern of ecdysone levels during development in the desert locust, Schistocerca gregaria. J. Insect Physiol. 1976, 22, 885–889. [Google Scholar] [CrossRef]
- Lenaerts, C.; Cools, D.; Verdonck, R.; Verbakel, L.; Vanden Broeck, J.; Marchal, E. The ecdysis triggering hormone system is essential for successful moulting of a major hemimetabolous pest insect, Schistocerca gregaria. Sci. Rep. 2017, 7, 46502. [Google Scholar] [CrossRef] [Green Version]
- Tawfik, A.I.; Mat’Hová, A.; Sehnal, F.; Ismail, S.H. Haemolymph ecdysteroids in the solitary and gregarious larvae of Schistocerca gregaria. Arch. Insect Biochem. Physiol. 1996, 31, 427–438. [Google Scholar] [CrossRef]
- Marchal, E.; Zhang, J.; Badisco, L.; Verlinden, H.; Hult, E.F.; Van Wielendaele, P.; Yagi, K.J.; Tobe, S.S.; Vanden Broeck, J. Final steps in juvenile hormone biosynthesis in the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2011, 41, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Lenaerts, C.; Marchal, E.; Peeters, P.; Vanden Broeck, J. The ecdysone receptor complex is essential for the reproductive success in the female desert locust, Schistocerca gregaria. Sci. Rep. 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symmons, P.M.; Cressman, K. Desert Locust Guidelines, Biology and Behaviour; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; pp. 1–43. [Google Scholar]
- Pener, M.P.; Simpson, S.J. Locust Phase Polyphenism: An Update. Adv. Insect Physiol. 2009, 36, 1–272. [Google Scholar]
- Roussi, A. The Battle to contain gigantic locust swarms. Nature 2020, 579, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations. GIEWS Update, South America, Outbreaks of Locusts Could Threaten Wheat and Barley Crops in Argentina, Brazil and Uruguay. 2020. Available online: http://www.fao.org/3/a-i7876e.pdf (accessed on 1 September 2020).
- Stephen, S. ‘Biblical’ Swarm of Locusts Plague Italian Farmlands in Worst Infestation since WWII; Fox News. 2019. Available online: https://www.foxnews.com/world/biblical-locusts-italy-sardinia-farmers (accessed on 14 August 2020).
- Carlone, M.; Sestito, D. ‘There’s Nothing Left’—Sardinian Farmland Stripped by Locust Swarms. CGTN. 2020. Available online: https://newseu.cgtn.com/news/2020-06-25/-There-s-nothing-left-Sardinian-farmland-stripped-by-locust-swarms-RzaYxkBVuw/index.html (accessed on 14 August 2020).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Van Hiel, M.B.; Van Wielendaele, P.; Temmerman, L.; Van Soest, S.; Vuerinckx, K.; Huybrechts, R.; Vanden Broeck, J.; Simonet, G. Identification and validation of housekeeping genes in brains of the desert locust Schistocerca gregaria under different developmental conditions. BMC Mol. Biol. 2009, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2020, 3. [Google Scholar] [CrossRef] [Green Version]
- Porcheron, P.; Morinière, M.; Grassi, J.; Pradelles, P. Development of an enzyme immunoassay for ecdysteroids using acetylcholinesterase as label. Insect Biochem. 1989, 19, 117–122. [Google Scholar] [CrossRef]
- Pascual, N.; Belles, X.; Delbecque, J.-P.; Hua, Y.-J.; Koolmanc, J. Quantification of Ecdysteroids by Immunoassay: Comparison of Enzyme Immunoassay and Radioimmunoassay. Z. Naturforsch. C J. Biosci. 1995, 50, 862–867. [Google Scholar] [CrossRef]
- Lafont, R.; Dauphin-Villemant, C.; Warren, J.T.; Rees, H. Ecdysteroid Chemistry and Biochemistry. In Insect Endocrinology; Academic Press: Cambdrige, MA, USA, 2012; pp. 106–176. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gijbels, M.; Marchal, E.; Verdonckt, T.W.; Bruyninckx, E.; Vanden Broeck, J. RNAi-Mediated Knockdown of Transcription Factor E93 in Nymphs of the Desert Locust (Schistocerca gregaria) Inhibits Adult Morphogenesis and Results in Supernumerary Juvenile Stages. Int. J. Mol. Sci. 2020, 21, 7518. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207518
Gijbels M, Marchal E, Verdonckt TW, Bruyninckx E, Vanden Broeck J. RNAi-Mediated Knockdown of Transcription Factor E93 in Nymphs of the Desert Locust (Schistocerca gregaria) Inhibits Adult Morphogenesis and Results in Supernumerary Juvenile Stages. International Journal of Molecular Sciences. 2020; 21(20):7518. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207518
Chicago/Turabian StyleGijbels, Marijke, Elisabeth Marchal, Thomas Wolf Verdonckt, Evert Bruyninckx, and Jozef Vanden Broeck. 2020. "RNAi-Mediated Knockdown of Transcription Factor E93 in Nymphs of the Desert Locust (Schistocerca gregaria) Inhibits Adult Morphogenesis and Results in Supernumerary Juvenile Stages" International Journal of Molecular Sciences 21, no. 20: 7518. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207518