Temperature-Dependent Alternative Splicing of Precursor mRNAs and Its Biological Significance: A Review Focused on Post-Transcriptional Regulation of a Cold Shock Protein Gene in Hibernating Mammals


Abstract
:1. Introduction
2. Regulation of Gene Expression by Temperature-Dependent Alternative Splicing
2.1. Flowering
2.2. Circadian Clock
2.3. Sex Determination
2.4. Others
3. Molecular Mechanism of Temperature-Dependent Alternative Splicing
4. Temperature-Dependent Regulation of Alternative Splicing in Gene Expression of CIRP
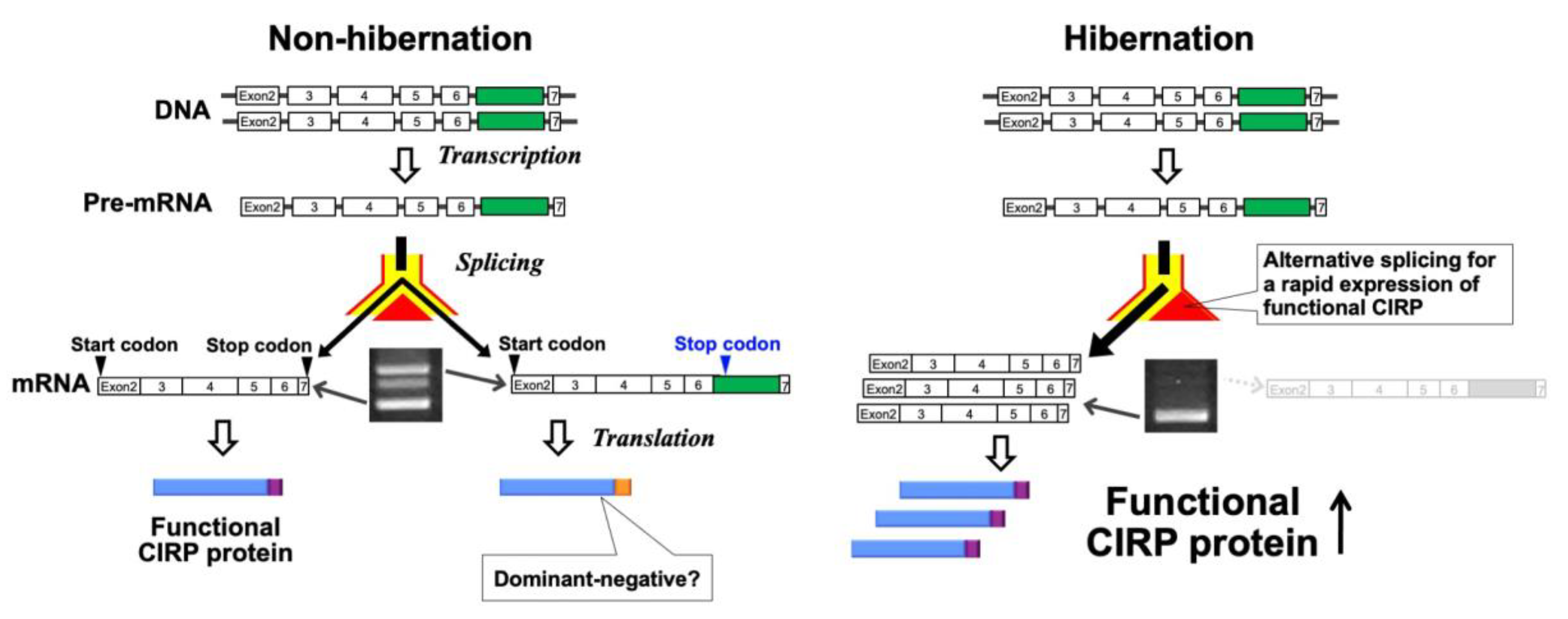
4.1. Discovery of Temperature-Dependent Alternative Splicing of a CIRP Transcript during Hibernation in Hamsters
4.2. Factors Triggering “Hibernation-Specific” Alternative Splicing
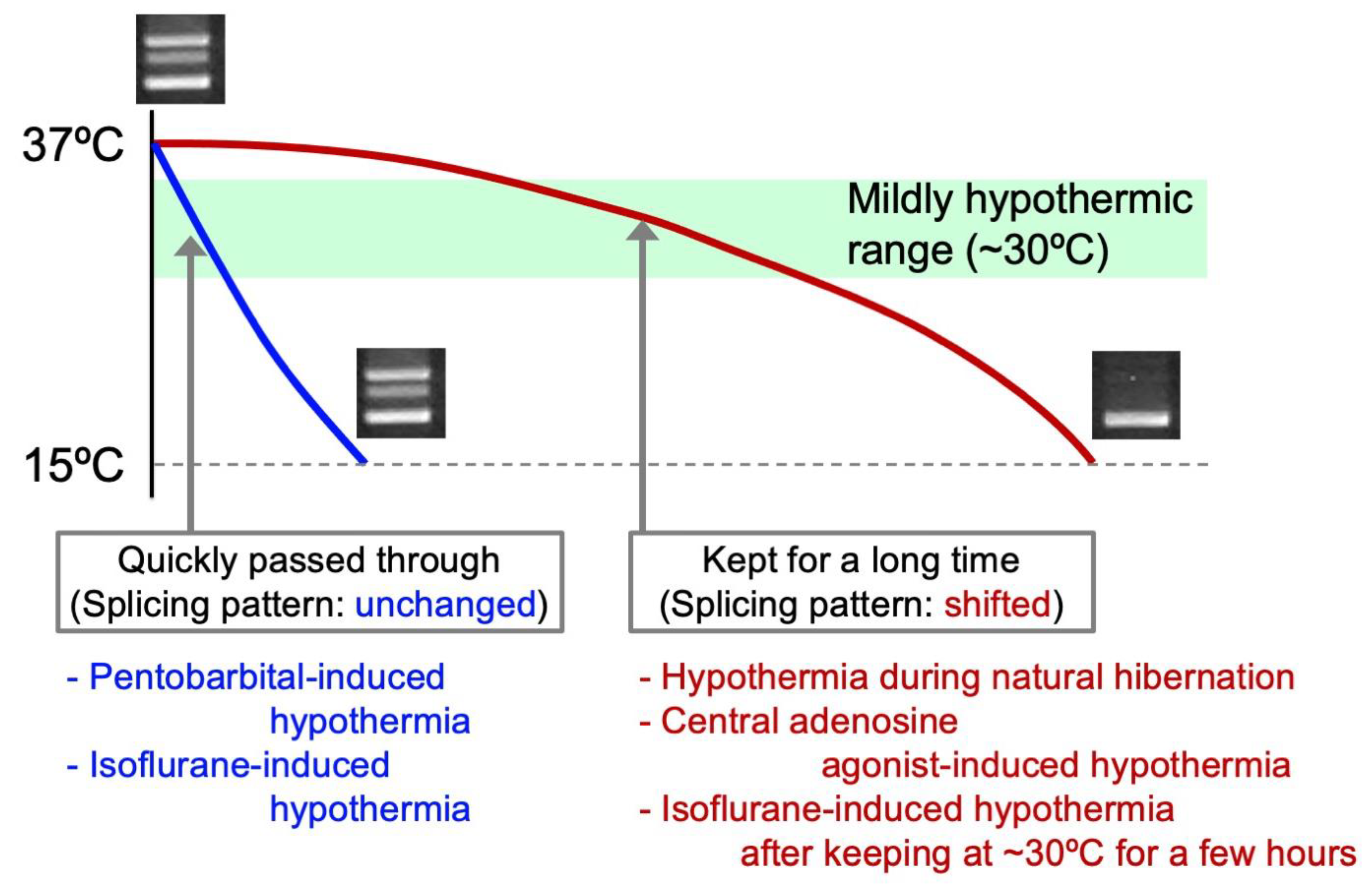
4.3. Mild Hypothermia as a Major Cause of the Shift in Alternative Splicing of CIRP Transcripts
4.4. Alternative Splicing of CIRP mRNA in a Non-Hibernator
4.5. Hypothesis for the Physiological Significance of Temperature-Dependent Alternative Splicing of CIRP mRNA

{kind=link}
{kind=link}
| Species | Genes | Related Functions | References |
|---|---|---|---|
| Arabidopsis | FLM | Flowering | [23,27,28,29,30,31,32,33,34,35,36] |
| Arabidopsis | MAF2 | Flowering | [11,25,38,39,40,41,42] |
| Arabidopsis | CCA1 | Circadian clock | [57,58,59,60,61] |
| Arabidopsis | LHY | Circadian clock | [57,59,60,64,90,141] |
| Arabidopsis | TOC1 | Circadian clock | [59,64] |
| Arabidopsis | PRR3 | Circadian clock | [59,64] |
| Arabidopsis | PRR5 | Circadian clock | [59,64] |
| Arabidopsis | PRR7 | Circadian clock | [59,64] |
| Arabidopsis | PRR9 | Circadian clock | [59,64] |
| Barley | LHY | Circadian clock | [65] |
| Barley | PPD-H1 | Circadian clock | [65] |
| Neurospora | FRQ | Circadian clock/thermosensing | [67,68,69] |
| Drosophila | PER | Circadian clock | [71,73] |
| Drosophila | TIM | Circadian clock | [66,74,77] |
| Tilapia | PER1/2 | Circadian clock | [72] |
| Mouse | U2AF26 | Circadian clock/alternative splicing | [70,75] |
| Mouse | CIRP | Circadian clock | [130,138] |
| Crocodile | Sox9 | Sex determination | [82] |
| Turtle | WT1 | Sex determination | [84,86] |
| Turtle | Dmrt1 | Sex determination | [78] |
| Mugger (Crocodile) | Dmrt1 | Sex determination | [83] |
| Arabidopsis | U2AF65A | Alternative splicing | [88] |
| Arabidopsis | CDKG1 | Alternative splicing | [33,88] |
| Arabidopsis | SR1 | Alternative splicing/temperature adaptation | [89] |
| Arabidopsis | PTB1/2 | Alternative splicing | [90] |
| Arabidopsis | SUA | Alternative splicing | [90] |
| Tilapia | SR splicing factors | Alternative splicing/temperature adaptation | [72] |
| Arabidopsis | RID1 | Plant development | [91] |
| Arabidopsis | AP3-1 | Floral phenotype | [92] |
| Rice | OsbZIP58 | Grain filling | [93] |
| Sugar beet | Bvnpcg2/3 | Sense seasonal temperature changes | [94] |
| Medicago | MtJMJC5 | Circadian clock | [95] |
| Arabidopsis | Cold stress response | [100,103] | |
| Tea plant | Cold stress response | [102] | |
| Jujuncao | Cold stress response | [101] | |
| Chlamydomonas | CGE1 | Chaperone | [107,108] |
| Yeast | APE2 | Thermosensing | [109,111] |
| Human (HeLa cell) | β-globin | [112] | |
| Human (fibroblast) | Collagen | [113] | |
| Hamster (CHO cells) | Dnml1/ Mff | [114] | |
| Syrian Hamster | CIRP | Hibernation | [13,14] |
| Mouse | CIRP | Hypothermia | [15] |
5. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| pre-mRNA | Precursor mRNA |
| CIRP | Cold-inducible RNA-binding protein |
| FLM | Flowering locus M |
| SVP | Short vegetative phase |
| FT | Flowering locus T |
| SOC1 | Suppressor of overexpression of constans 1 |
| MAF2 | MADS affecting flowering 2 |
| CCA1 | Circadian clock associated 1 |
| LHY | Late elongated hypocotyl |
| ELF | Early flowering |
| LUX | Lux arrhythmo |
| PRR | Pseudo response regulator |
| TOC1 | Timing of CAB expression 1 |
| NMD | Nonsense-mediated decay |
| PPD-H1 | Photoperiod-H1 |
| FRQ | Frequency |
| PER | Period |
| TIM | Timeless |
| U2AF | U2 snRNP auxiliary factor |
| GSD | Genotypic sex determination |
| ESD | Environmental sex determination |
| TSD | Temperature-dependent sex determination |
| Sox9 | SRY-box9 |
| WT1 | Wilms tumor 1 |
| Dmrt1 | Doublesex and mab3-related transcription factor 1 |
| CDKG1 | Cyclin-dependent kinase G1 |
| PTB | Polypyrimdine tract-binding protein |
| SUA | Suppressor of abi3-5 |
| RID1 | Root initiation defective1 |
| SR | Serine-arginine-rich |
| AP3 | Apetala3 |
| TRPs | Transient receptor potentials |
| CHO | Chinese hamster ovary |
| CLK | CDC-like kinase |
| SF | Splicing factor |
| PSI | P-element somatic inhibitor |
| H3K36me3 | Trimethylated histone H3 at lysine 36 |
| RBM3 | RNA-binding motif 3 |
| ECG | Electrocardiograms |
| PTC | Premature termination codon |
References
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Bowler, E.; Oltean, S. Alternative Splicing in Angiogenesis. Int. J. Mol. Sci. 2019, 20, 2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Manley, J.L. Mechanisms of alternative splicing regulation: Insights from molecular and genomics approaches. Nat. Rev. Mol. Cell Biol. 2009, 10, 741–754. [Google Scholar] [CrossRef]
- Furlanis, E.; Scheiffele, P. Regulation of Neuronal Differentiation, Function, and Plasticity by Alternative Splicing. Annu. Rev. Cell Dev. Biol. 2018, 34, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, O.; Convertini, P.; Zhang, Z.; Wen, Y.; Shen, M.; Falaleeva, M.; Stamm, S. Function of alternative splicing. Gene 2013, 514, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipscombe, D.; Lopez Soto, E.J. Alternative splicing of neuronal genes: New mechanisms and new therapies. Curr. Opin. Neurobiol. 2019, 57, 26–31. [Google Scholar] [CrossRef] [PubMed]
- De Conti, L.; Baralle, M.; Buratti, E. Exon and intron definition in pre-mRNA splicing. Wiley Interdiscip. Rev. RNA 2013, 4, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Rio, D.C. Mechanisms and Regulation of Alternative Pre-mRNA Splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeiwa, T.; Mitobe, Y.; Ikeda, K.; Horie-Inoue, K.; Inoue, S. Roles of Splicing Factors in Hormone-Related Cancer Progression. Int. J. Mol. Sci. 2020, 21, 1551. [Google Scholar] [CrossRef] [Green Version]
- Okabe, K.; Sakaguchi, R.; Shi, B.; Kiyonaka, S. Intracellular thermometry with fluorescent sensors for thermal biology. Pflugers Arch. 2018, 470, 717–731. [Google Scholar] [CrossRef]
- Susila, H.; Nasim, Z.; Ahn, J.H. Ambient Temperature-Responsive Mechanisms Coordinate Regulation of Flowering Time. Int. J. Mol. Sci. 2018, 19, 3196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tansey, E.A.; Johnson, C.D. Recent advances in thermoregulation. Adv. Physiol. Educ. 2015, 39, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Horii, Y.; Shimaoka, H.; Horii, K.; Shiina, T.; Shimizu, Y. Mild hypothermia causes a shift in the alternative splicing of cold-inducible RNA-binding protein transcripts in Syrian hamsters. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R240–R247. [Google Scholar] [CrossRef] [PubMed]
- Sano, Y.; Shiina, T.; Naitou, K.; Nakamori, H.; Shimizu, Y. Hibernation-specific alternative splicing of the mRNA encoding cold-inducible RNA-binding protein in the hearts of hamsters. Biochem. Biophys. Res. Commun. 2015, 462, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Horii, Y.; Shiina, T.; Uehara, S.; Nomura, K.; Shimaoka, H.; Horii, K.; Shimizu, Y. Hypothermia induces changes in the alternative splicing pattern of cold-inducible RNA-binding protein transcripts in a non-hibernator, the mouse. Biomed. Res. 2019, 40, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Capovilla, G.; Pajoro, A.; Immink, R.G.; Schmid, M. Role of alternative pre-mRNA splicing in temperature signaling. Curr. Opin. Plant Biol. 2015, 27, 97–103. [Google Scholar] [CrossRef]
- Staiger, D.; Brown, J.W. Alternative splicing at the intersection of biological timing, development, and stress responses. Plant Cell 2013, 25, 3640–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streitner, C.; Simpson, C.G.; Shaw, P.; Danisman, S.; Brown, J.W.; Staiger, D. Small changes in ambient temperature affect alternative splicing in Arabidopsis thaliana. Plant Signal Behav. 2013, 8, e24638. [Google Scholar] [CrossRef] [Green Version]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol. 2009, 19, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Lee, H.J.; Gil, K.E.; Kim, J.Y.; Lee, J.H.; Lee, H.; Cho, H.T.; Vu, L.D.; De Smet, I.; Park, C.M. Developmental Programming of Thermonastic Leaf Movement. Plant Physiol. 2019, 180, 1185–1197. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Lee, J.H.; Kim, J.Y.; Park, C.M. Alternative RNA Splicing Expands the Developmental Plasticity of Flowering Transition. Front. Plant Sci. 2019, 10, 606. [Google Scholar] [CrossRef] [Green Version]
- Quint, M.; Delker, C.; Franklin, K.A.; Wigge, P.A.; Halliday, K.J.; van Zanten, M. Molecular and genetic control of plant thermomorphogenesis. Nat. Plants 2016, 2, 15190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Ryu, H.S.; Chung, K.S.; Pose, D.; Kim, S.; Schmid, M.; Ahn, J.H. Regulation of temperature-responsive flowering by MADS-box transcription factor repressors. Science 2013, 342, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Michaels, S.D. Flowering time regulation produces much fruit. Curr. Opin. Plant Biol. 2009, 12, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Shang, X.; Cao, Y.; Ma, L. Alternative Splicing in Plant Genes: A Means of Regulating the Environmental Fitness of Plants. Int. J. Mol. Sci. 2017, 18, 432. [Google Scholar] [CrossRef] [PubMed]
- Capovilla, G.; Schmid, M.; Pose, D. Control of flowering by ambient temperature. J. Exp. Bot. 2015, 66, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pose, D.; Verhage, L.; Ott, F.; Yant, L.; Mathieu, J.; Angenent, G.C.; Immink, R.G.; Schmid, M. Temperature-dependent regulation of flowering by antagonistic FLM variants. Nature 2013, 503, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Scortecci, K.; Michaels, S.D.; Amasino, R.M. Genetic interactions between FLM and other flowering-time genes in Arabidopsis thaliana. Plant Mol. Biol. 2003, 52, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Sureshkumar, S.; Lempe, J.; Weigel, D. Potent induction of Arabidopsis thaliana flowering by elevated growth temperature. PLoS Genet. 2006, 2, e106. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Weigel, D. Temperature Induced Flowering in Arabidopsis thaliana. Plant Signal Behav. 2006, 1, 227–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capovilla, G.; Symeonidi, E.; Wu, R.; Schmid, M. Contribution of major FLM isoforms to temperature-dependent flowering in Arabidopsis thaliana. J. Exp. Bot. 2017, 68, 5117–5127. [Google Scholar] [CrossRef] [PubMed]
- Lutz, U.; Pose, D.; Pfeifer, M.; Gundlach, H.; Hagmann, J.; Wang, C.; Weigel, D.; Mayer, K.F.; Schmid, M.; Schwechheimer, C. Modulation of Ambient Temperature-Dependent Flowering in Arabidopsis thaliana by Natural Variation of FLOWERING LOCUS M. PLoS Genet. 2015, 11, e1005588. [Google Scholar] [CrossRef] [PubMed]
- Nibau, C.; Gallemi, M.; Dadarou, D.; Doonan, J.H.; Cavallari, N. Thermo-Sensitive Alternative Splicing of FLOWERING LOCUS M Is Modulated by Cyclin-Dependent Kinase G2. Front. Plant Sci. 2019, 10, 1680. [Google Scholar] [CrossRef]
- Scortecci, K.C.; Michaels, S.D.; Amasino, R.M. Identification of a MADS-box gene, FLOWERING LOCUS M, that represses flowering. Plant J. 2001, 26, 229–236. [Google Scholar] [CrossRef]
- Sureshkumar, S.; Dent, C.; Seleznev, A.; Tasset, C.; Balasubramanian, S. Nonsense-mediated mRNA decay modulates FLM-dependent thermosensory flowering response in Arabidopsis. Nat. Plants 2016, 2, 16055. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Xiong, F.; Ren, Q.P.; Wang, X.L. Regulation of flowering transition by alternative splicing: The role of the U2 auxiliary factor. J. Exp. Bot. 2020, 71, 751–758. [Google Scholar] [CrossRef]
- Lee, J.H.; Yoo, S.J.; Park, S.H.; Hwang, I.; Lee, J.S.; Ahn, J.H. Role of SVP in the control of flowering time by ambient temperature in Arabidopsis. Genes Dev. 2007, 21, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, O.J.; Kumimoto, R.W.; Wong, B.J.; Riechmann, J.L. Analysis of the Arabidopsis MADS AFFECTING FLOWERING gene family: MAF2 prevents vernalization by short periods of cold. Plant Cell 2003, 15, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Rosloski, S.M.; Jali, S.S.; Balasubramanian, S.; Weigel, D.; Grbic, V. Natural diversity in flowering responses of Arabidopsis thaliana caused by variation in a tandem gene array. Genetics 2010, 186, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Rosloski, S.M.; Singh, A.; Jali, S.S.; Balasubramanian, S.; Weigel, D.; Grbic, V. Functional analysis of splice variant expression of MADS AFFECTING FLOWERING 2 of Arabidopsis thaliana. Plant Mol. Biol. 2013, 81, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salome, P.A.; Bomblies, K.; Laitinen, R.A.; Yant, L.; Mott, R.; Weigel, D. Genetic architecture of flowering-time variation in Arabidopsis thaliana. Genetics 2011, 188, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airoldi, C.A.; McKay, M.; Davies, B. MAF2 Is Regulated by Temperature-Dependent Splicing and Represses Flowering at Low Temperatures in Parallel with FLM. PLoS ONE 2015, 10, e0126516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartok, O.; Kyriacou, C.P.; Levine, J.; Sehgal, A.; Kadener, S. Adaptation of molecular circadian clockwork to environmental changes: A role for alternative splicing and miRNAs. Proc. Biol. Sci. 2013, 280, 20130011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmer, S.L. The circadian system in higher plants. Annu. Rev. Plant Biol. 2009, 60, 357–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClung, C.R. Plant circadian rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef] [Green Version]
- McClung, C.R. The genetics of plant clocks. Adv. Genet. 2011, 74, 105–139. [Google Scholar] [PubMed]
- Nakamichi, N. Molecular mechanisms underlying the Arabidopsis circadian clock. Plant Cell Physiol. 2011, 52, 1709–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salome, P.A.; McClung, C.R. The Arabidopsis thaliana clock. J. Biol. Rhythms 2004, 19, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Helfer, A.; Nusinow, D.A.; Chow, B.Y.; Gehrke, A.R.; Bulyk, M.L.; Kay, S.A. LUX ARRHYTHMO encodes a nighttime repressor of circadian gene expression in the Arabidopsis core clock. Curr. Biol. 2011, 21, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, E.; Kolmos, E.; Bujdoso, N.; Yuan, Y.; Wang, M.; Berns, M.C.; Uhlworm, H.; Coupland, G.; Saini, R.; Jaskolski, M.; et al. EARLY FLOWERING4 recruitment of EARLY FLOWERING3 in the nucleus sustains the Arabidopsis circadian clock. Plant Cell 2012, 24, 428–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, B.Y.; Kay, S.A. Global approaches for telling time: Omics and the Arabidopsis circadian clock. Semin. Cell Dev. Biol. 2013, 24, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Farre, E.M.; Harmer, S.L.; Harmon, F.G.; Yanovsky, M.J.; Kay, S.A. Overlapping and distinct roles of PRR7 and PRR9 in the Arabidopsis circadian clock. Curr. Biol. 2005, 15, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, N.; Kiba, T.; Henriques, R.; Mizuno, T.; Chua, N.H.; Sakakibara, H. PSEUDO-RESPONSE REGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell 2010, 22, 594–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabadi, D.; Oyama, T.; Yanovsky, M.J.; Harmon, F.G.; Mas, P.; Kay, S.A. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 2001, 293, 880–883. [Google Scholar] [CrossRef]
- Green, R.M.; Tobin, E.M. The role of CCA1 and LHY in the plant circadian clock. Dev. Cell 2002, 2, 516–518. [Google Scholar] [CrossRef] [Green Version]
- Kojima, S.; Shingle, D.L.; Green, C.B. Post-transcriptional control of circadian rhythms. J. Cell Sci. 2011, 124, 311–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filichkin, S.A.; Mockler, T.C. Unproductive alternative splicing and nonsense mRNAs: A widespread phenomenon among plant circadian clock genes. Biol. Direct. 2012, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.K.; Mockler, T.C. Genome-wide mapping of alternative splicing in Arabidopsis thaliana. Genome Res. 2010, 20, 45–58. [Google Scholar] [CrossRef] [Green Version]
- James, A.B.; Syed, N.H.; Bordage, S.; Marshall, J.; Nimmo, G.A.; Jenkins, G.I.; Herzyk, P.; Brown, J.W.; Nimmo, H.G. Alternative splicing mediates responses of the Arabidopsis circadian clock to temperature changes. Plant Cell 2012, 24, 961–981. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.M.; Tartaglio, V.; Duarte, M.; Harmon, F.G. The Arabidopsis sickle Mutant Exhibits Altered Circadian Clock Responses to Cool Temperatures and Temperature-Dependent Alternative Splicing. Plant Cell 2016, 28, 2560–2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, P.J.; Park, M.J.; Lim, M.H.; Kim, S.G.; Lee, M.; Baldwin, I.T.; Park, C.M. A self-regulatory circuit of CIRCADIAN CLOCK-ASSOCIATED1 underlies the circadian clock regulation of temperature responses in Arabidopsis. Plant Cell 2012, 24, 2427–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, X.; Sugano, S.; Tobin, E.M. CK2 phosphorylation of CCA1 is necessary for its circadian oscillator function in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 3292–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.J.; Seo, P.J.; Park, C.M. CCA1 alternative splicing as a way of linking the circadian clock to temperature response in Arabidopsis. Plant Signal Behav. 2012, 7, 1194–1196. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.J.; Park, M.J.; Kim, S.G.; Baldwin, I.T.; Park, C.M. Alternative splicing and nonsense-mediated decay of circadian clock genes under environmental stress conditions in Arabidopsis. BMC Plant Biol. 2014, 14, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calixto, C.P.; Simpson, C.G.; Waugh, R.; Brown, J.W. Alternative Splicing of Barley Clock Genes in Response to Low Temperature. PLoS ONE 2016, 11, e0168028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrieux, A.; Xue, Y.; Cai, Y.; Lewald, K.M.; Nguyen, H.N.; Zhang, Y.; Chiu, J.C. EYES ABSENT and TIMELESS integrate photoperiodic and temperature cues to regulate seasonal physiology in Drosophila. Proc. Natl. Acad. Sci. USA 2020, 117, 15293–15304. [Google Scholar] [CrossRef] [PubMed]
- Colot, H.V.; Loros, J.J.; Dunlap, J.C. Temperature-modulated alternative splicing and promoter use in the Circadian clock gene frequency. Mol. Biol. Cell 2005, 16, 5563–5571. [Google Scholar] [CrossRef] [Green Version]
- Diernfellner, A.; Colot, H.V.; Dintsis, O.; Loros, J.J.; Dunlap, J.C.; Brunner, M. Long and short isoforms of Neurospora clock protein FRQ support temperature-compensated circadian rhythms. FEBS Lett. 2007, 581, 5759–5764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diernfellner, A.C.; Schafmeier, T.; Merrow, M.W.; Brunner, M. Molecular mechanism of temperature sensing by the circadian clock of Neurospora crassa. Genes Dev. 2005, 19, 1968–1973. [Google Scholar] [CrossRef] [Green Version]
- Goldammer, G.; Neumann, A.; Strauch, M.; Muller-McNicoll, M.; Heyd, F.; Preussner, M. Characterization of cis-acting elements that control oscillating alternative splicing. RNA Biol. 2018, 15, 1081–1092. [Google Scholar] [CrossRef]
- Hughes, M.E.; Grant, G.R.; Paquin, C.; Qian, J.; Nitabach, M.N. Deep sequencing the circadian and diurnal transcriptome of Drosophila brain. Genome Res. 2012, 22, 1266–1281. [Google Scholar] [CrossRef] [Green Version]
- Li, B.J.; Zhu, Z.X.; Qin, H.; Meng, Z.N.; Lin, H.R.; Xia, J.H. Genome-Wide Characterization of Alternative Splicing Events and Their Responses to Cold Stress in Tilapia. Front. Genet. 2020, 11, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majercak, J.; Sidote, D.; Hardin, P.E.; Edery, I. How a circadian clock adapts to seasonal decreases in temperature and day length. Neuron 1999, 24, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Martin Anduaga, A.; Evantal, N.; Patop, I.L.; Bartok, O.; Weiss, R.; Kadener, S. Thermosensitive alternative splicing senses and mediates temperature adaptation in Drosophila. Elife 2019, 8, e44642. [Google Scholar] [CrossRef]
- Preussner, M.; Goldammer, G.; Neumann, A.; Haltenhof, T.; Rautenstrauch, P.; Muller-McNicoll, M.; Heyd, F. Body Temperature Cycles Control Rhythmic Alternative Splicing in Mammals. Mol. Cell 2017, 67, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Preussner, M.; Wilhelmi, I.; Schultz, A.S.; Finkernagel, F.; Michel, M.; Moroy, T.; Heyd, F. Rhythmic U2af26 alternative splicing controls PERIOD1 stability and the circadian clock in mice. Mol. Cell 2014, 54, 651–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, L.E.; Ling, J.; Joshi, R.; Evantal, N.; Kadener, S.; Emery, P. Drosophila PSI controls circadian period and the phase of circadian behavior under temperature cycle via tim splicing. Elife 2019, 8, e50063. [Google Scholar] [CrossRef]
- Mizoguchi, B.; Valenzuela, N. Alternative splicing and thermosensitive expression of Dmrt1 during urogenital development in the painted turtle, Chrysemys picta. PeerJ 2020, 8, e8639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarre, S.D.; Georges, A.; Quinn, A. The ends of a continuum: Genetic and temperature-dependent sex determination in reptiles. Bioessays 2004, 26, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, N.; Adams, D.C.; Janzen, F.J. Pattern does not equal process: Exactly when is sex environmentally determined? Am. Nat. 2003, 161, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, R.; Wessely, O.; Anand, A.; Singh, L.; Aggarwal, R.K. Male-specific expression of Sox9 during gonad development of crocodile and mouse is mediated by alternative splicing of its proline-glutamine-alanine rich domain. FEBS J. 2009, 276, 4184–4196. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Patel, M.; Lalremruata, A.; Singh, A.P.; Agrawal, R.; Singh, L.; Aggarwal, R.K. Multiple alternative splicing of Dmrt1 during gonadogenesis in Indian mugger, a species exhibiting temperature-dependent sex determination. Gene 2008, 425, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Rhen, T.; Fagerlie, R.; Schroeder, A.; Crossley, D.A., 2nd; Lang, J.W. Molecular and morphological differentiation of testes and ovaries in relation to the thermosensitive period of gonad development in the snapping turtle, Chelydra serpentina. Differentiation 2015, 89, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, C.M.; Crews, D. Analyzing the coordinated gene network underlying temperature-dependent sex determination in reptiles. Semin. Cell Dev. Biol. 2009, 20, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Spotila, L.D.; Hall, S.E. Expression of a new RNA-splice isoform of WT1 in developing kidney-gonadal complexes of the turtle, Trachemys scripta. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 119, 761–767. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Iwasa, Y. Temperature-dependent sex determination, realized by hormonal dynamics with enzymatic reactions sensitive to ambient temperature. J. Theor. Biol. 2018, 453, 146–155. [Google Scholar] [CrossRef]
- Cavallari, N.; Nibau, C.; Fuchs, A.; Dadarou, D.; Barta, A.; Doonan, J.H. The cyclin-dependent kinase G group defines a thermo-sensitive alternative splicing circuit modulating the expression of Arabidopsis ATU2AF65A. Plant J. 2018, 94, 1010–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazar, G.; Goodman, H.M. The Arabidopsis splicing factor SR1 is regulated by alternative splicing. Plant Mol. Biol. 2000, 42, 571–581. [Google Scholar] [CrossRef]
- James, A.B.; Calixto, C.P.G.; Tzioutziou, N.A.; Guo, W.; Zhang, R.; Simpson, C.G.; Jiang, W.; Nimmo, G.A.; Brown, J.W.S.; Nimmo, H.G. How does temperature affect splicing events? Isoform switching of splicing factors regulates splicing of LATE ELONGATED HYPOCOTYL (LHY). Plant Cell Environ. 2018, 41, 1539–1550. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, M.; Demura, T.; Sugiyama, M. Arabidopsis root initiation defective1, a DEAH-box RNA helicase involved in pre-mRNA splicing, is essential for plant development. Plant Cell 2013, 25, 2056–2069. [Google Scholar] [CrossRef] [Green Version]
- Sablowski, R.W.; Meyerowitz, E.M. Temperature-sensitive splicing in the floral homeotic mutant apetala3-1. Plant Cell 1998, 10, 1453–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Li, X.; Zhang, H.; Wang, L.; Zhu, Z.; Gao, J.; Li, C.; Zhu, Y. High temperature inhibits the accumulation of storage materials by inducing alternative splicing of OsbZIP58 during filling stage in rice. Plant Cell Environ. 2020, 43, 1879–1896. [Google Scholar] [CrossRef]
- Rotthues, A.; Kappler, J.; Lichtfuss, A.; Kloos, D.U.; Stahl, D.J.; Hehl, R. Post-harvest regulated gene expression and splicing efficiency in storage roots of sugar beet (Beta vulgaris L.). Planta 2008, 227, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wu, X.; Liu, D.; Song, S.; Liu, D.; Wang, H. Cold-dependent alternative splicing of a Jumonji C domain-containing gene MtJMJC5 in Medicago truncatula. Biochem. Biophys. Res. Commun. 2016, 474, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, M.W.; Ko, M.C. Functional evolution of vertebrate sensory receptors. Horm. Behav. 2020, 124, 104771. [Google Scholar] [CrossRef]
- Lamas, J.A.; Rueda-Ruzafa, L.; Herrera-Perez, S. Ion Channels and Thermosensitivity: TRP, TREK, or Both? Int. J. Mol. Sci. 2019, 20, 2371. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Fukuda, N.; Matsumoto, T.; Iribe, Y.; Tsunemi, A.; Kazama, T.; Yoshida-Noro, C.; Hayashi, N. Moderate low temperature preserves the stemness of neural stem cells and suppresses apoptosis of the cells via activation of the cold-inducible RNA binding protein. Brain Res. 2010, 1358, 20–29. [Google Scholar] [CrossRef]
- Takayama, Y.; Derouiche, S.; Maruyama, K.; Tominaga, M. Emerging Perspectives on Pain Management by Modulation of TRP Channels and ANO1. Int. J. Mol. Sci. 2019, 20, 3411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calixto, C.P.G.; Guo, W.; James, A.B.; Tzioutziou, N.A.; Entizne, J.C.; Panter, P.E.; Knight, H.; Nimmo, H.G.; Zhang, R.; Brown, J.W.S. Rapid and Dynamic Alternative Splicing Impacts the Arabidopsis Cold Response Transcriptome. Plant Cell 2018, 30, 1424–1444. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Xiang, C.; Xu, L.; Cui, J.; Fu, S.; Chen, B.; Yang, S.; Wang, P.; Xie, Y.; Wei, M.; et al. SMRT sequencing of a full-length transcriptome reveals transcript variants involved in C18 unsaturated fatty acid biosynthesis and metabolism pathways at chilling temperature in Pennisetum giganteum. BMC Genom. 2020, 21, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Mi, X.; Zhao, S.; Zhu, J.; Guo, R.; Xia, X.; Liu, L.; Liu, S.; Wei, C. Comprehensive profiling of alternative splicing landscape during cold acclimation in tea plant. BMC Genom. 2020, 21, 65. [Google Scholar] [CrossRef] [Green Version]
- Vyse, K.; Faivre, L.; Romich, M.; Pagter, M.; Schubert, D.; Hincha, D.K.; Zuther, E. Transcriptional and Post-Transcriptional Regulation and Transcriptional Memory of Chromatin Regulators in Response to Low Temperature. Front. Plant Sci. 2020, 11, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, P.J.; Park, M.J.; Park, C.M. Alternative splicing of transcription factors in plant responses to low temperature stress: Mechanisms and functions. Planta 2013, 237, 1415–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 chaperone machines. Cell 1998, 92, 351–366. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willmund, F.; Muhlhaus, T.; Wojciechowska, M.; Schroda, M. The NH2-terminal domain of the chloroplast GrpE homolog CGE1 is required for dimerization and cochaperone function in vivo. J. Biol. Chem. 2007, 282, 11317–11328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroda, M.; Vallon, O.; Whitelegge, J.P.; Beck, C.F.; Wollman, F.A. The chloroplastic GrpE homolog of Chlamydomonas: Two isoforms generated by differential splicing. Plant Cell 2001, 13, 2823–2839. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Plass, M.; Perez-Valle, J.; Eyras, E.; Vilardell, J. Deciphering 3′ss selection in the yeast genome reveals an RNA thermosensor that mediates alternative splicing. Mol. Cell 2011, 43, 1033–1039. [Google Scholar] [CrossRef] [Green Version]
- Somero, G.N. RNA thermosensors: How might animals exploit their regulatory potential? J. Exp. Biol. 2018, 221, jeb162842. [Google Scholar] [CrossRef] [Green Version]
- Yassour, M.; Kaplan, T.; Fraser, H.B.; Levin, J.Z.; Pfiffner, J.; Adiconis, X.; Schroth, G.; Luo, S.; Khrebtukova, I.; Gnirke, A.; et al. Ab initio construction of a eukaryotic transcriptome by massively parallel mRNA sequencing. Proc. Natl. Acad. Sci. USA 2009, 106, 3264–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemignani, F.; Sazani, P.; Morcos, P.; Kole, R. Temperature-dependent splicing of beta-globin pre-mRNA. Nucleic Acids Res. 2002, 30, 4592–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weil, D.; D’Alessio, M.; Ramirez, F.; Steinmann, B.; Wirtz, M.K.; Glanville, R.W.; Hollister, D.W. Temperature-dependent expression of a collagen splicing defect in the fibroblasts of a patient with Ehlers-Danlos syndrome type VII. J. Biol. Chem. 1989, 264, 16804–16809. [Google Scholar]
- Tzani, I.; Monger, C.; Motheramgari, K.; Gallagher, C.; Hagan, R.; Kelly, P.; Costello, A.; Meiller, J.; Floris, P.; Zhang, L.; et al. Subphysiological temperature induces pervasive alternative splicing in Chinese hamster ovary cells. Biotechnol. Bioeng. 2020, 117, 2489–2503. [Google Scholar] [CrossRef]
- Ip, J.Y.; Schmidt, D.; Pan, Q.; Ramani, A.K.; Fraser, A.G.; Odom, D.T.; Blencowe, B.J. Global impact of RNA polymerase II elongation inhibition on alternative splicing regulation. Genome Res. 2011, 21, 390–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiller, M.; Zhang, Z.; Backofen, R.; Stamm, S. Pre-mRNA secondary structures influence exon recognition. PLoS Genet. 2007, 3, e204. [Google Scholar] [CrossRef] [Green Version]
- Haltenhof, T.; Kotte, A.; De Bortoli, F.; Schiefer, S.; Meinke, S.; Emmerichs, A.K.; Petermann, K.K.; Timmermann, B.; Imhof, P.; Franz, A.; et al. A Conserved Kinase-Based Body-Temperature Sensor Globally Controls Alternative Splicing and Gene Expression. Mol. Cell 2020, 78, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Iida, K.; Seki, M.; Sakurai, T.; Satou, M.; Akiyama, K.; Toyoda, T.; Konagaya, A.; Shinozaki, K. Genome-wide analysis of alternative pre-mRNA splicing in Arabidopsis thaliana based on full-length cDNA sequences. Nucleic Acids Res. 2004, 32, 5096–5103. [Google Scholar] [CrossRef] [Green Version]
- Palusa, S.G.; Ali, G.S.; Reddy, A.S. Alternative splicing of pre-mRNAs of Arabidopsis serine/arginine-rich proteins: Regulation by hormones and stresses. Plant J. 2007, 49, 1091–1107. [Google Scholar] [CrossRef]
- Melzer, R. Regulation of flowering time: A splicy business. J. Exp. Bot. 2017, 68, 5017–5020. [Google Scholar] [CrossRef] [Green Version]
- Steffen, A.; Staiger, D. Chromatin marks and ambient temperature-dependent flowering strike up a novel liaison. Genome Biol. 2017, 18, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.L.; Luo, G.; Wise, J.A.; Lou, H. Regulation of alternative splicing by local histone modifications: Potential roles for RNA-guided mechanisms. Nucleic Acids Res. 2014, 42, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Pajoro, A.; Severing, E.; Angenent, G.C.; Immink, R.G.H. Histone H3 lysine 36 methylation affects temperature-induced alternative splicing and flowering in plants. Genome Biol. 2017, 18, 102. [Google Scholar] [CrossRef]
- Carey, H.V.; Andrews, M.T.; Martin, S.L. Mammalian hibernation: Cellular and molecular responses to depressed metabolism and low temperature. Physiol. Rev. 2003, 83, 1153–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruf, T.; Geiser, F. Daily torpor and hibernation in birds and mammals. Biol. Rev. Camb. Philos. Soc. 2015, 90, 891–926. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, K.P. Physiological blocking of the mechanisms of cold death: Theoretical and experimental considerations. J. Therm. Biol. 2000, 25, 467–479. [Google Scholar] [CrossRef]
- Horii, Y.; Shiina, T.; Shimizu, Y. The Mechanism Enabling Hibernation in Mammals. Adv. Exp. Med. Biol. 2018, 1081, 45–60. [Google Scholar] [PubMed]
- Miyazawa, S.; Shimizu, Y.; Shiina, T.; Hirayama, H.; Morita, H.; Takewaki, T. Central A1-receptor activation associated with onset of torpor protects the heart against low temperature in the Syrian hamster. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R991–R996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Buhrer, C.; Wellmann, S. Cold-inducible proteins CIRP and RBM3, a unique couple with activities far beyond the cold. Cell Mol. Life Sci. 2016, 73, 3839–3859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotic, I.; Omidi, S.; Fleury-Olela, F.; Molina, N.; Naef, F.; Schibler, U. Temperature regulates splicing efficiency of the cold-inducible RNA-binding protein gene Cirbp. Genes Dev. 2016, 30, 2005–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, H.; Itoh, K.; Kaneko, Y.; Kishishita, M.; Yoshida, O.; Fujita, J. A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth. J. Cell. Biol. 1997, 137, 899–908. [Google Scholar] [CrossRef]
- Lleonart, M.E. A new generation of proto-oncogenes: Cold-inducible RNA binding proteins. Biochim. Biophys. Acta 2010, 1805, 43–52. [Google Scholar] [CrossRef]
- Gualerzi, C.O.; Giuliodori, A.M.; Pon, C.L. Transcriptional and post-transcriptional control of cold-shock genes. J. Mol. Biol. 2003, 331, 527–539. [Google Scholar] [CrossRef]
- Saitongdee, P.; Milner, P.; Becker, D.L.; Knight, G.E.; Burnstock, G. Increased connexin43 gap junction protein in hamster cardiomyocytes during cold acclimatization and hibernation. Cardiovasc. Res. 2000, 47, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Jinka, T.R.; Toien, O.; Drew, K.L. Season primes the brain in an arctic hibernator to facilitate entrance into torpor mediated by adenosine A(1) receptors. J. Neurosci. 2011, 31, 10752–10758. [Google Scholar] [CrossRef] [PubMed]
- Tamura, Y.; Shintani, M.; Nakamura, A.; Monden, M.; Shiomi, H. Phase-specific central regulatory systems of hibernation in Syrian hamsters. Brain Res. 2005, 1045, 88–96. [Google Scholar] [CrossRef]
- Horwitz, B.A.; Chau, S.M.; Hamilton, J.S.; Song, C.; Gorgone, J.; Saenz, M.; Horowitz, J.M.; Chen, C.Y. Temporal relationships of blood pressure, heart rate, baroreflex function, and body temperature change over a hibernation bout in Syrian hamsters. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R759–R768. [Google Scholar] [CrossRef]
- Gotic, I.; Schibler, U. Posttranscriptional mechanisms controlling diurnal gene expression cycles by body temperature rhythms. RNA Biol. 2017, 14, 1294–1298. [Google Scholar] [CrossRef] [Green Version]
- De Leeuw, F.; Zhang, T.; Wauquier, C.; Huez, G.; Kruys, V.; Gueydan, C. The cold-inducible RNA-binding protein migrates from the nucleus to cytoplasmic stress granules by a methylation-dependent mechanism and acts as a translational repressor. Exp. Cell Res. 2007, 313, 4130–4144. [Google Scholar] [CrossRef] [PubMed]
- Brunson, C.E.; Abbud, E.; Osman, K.; Skelton, T.N.; Markov, A.K. Osborn (J) wave appearance on the electrocardiogram in relation to potassium transfer and myocardial metabolism during hypothermia. J. Investig. Med. 2005, 53, 434–437. [Google Scholar] [CrossRef] [PubMed]
- James, A.B.; Sullivan, S.; Nimmo, H.G. Global spatial analysis of Arabidopsis natural variants implicates 5′UTR splicing of late elongated hypocotyl in responses to temperature. Plant Cell Environ. 2018, 41, 1524–1538. [Google Scholar] [CrossRef] [Green Version]
- Shimaoka, H.; Kawaguchi, T.; Morikawa, K.; Sano, Y.; Naitou, K.; Nakamori, H.; Shiina, T.; Shimizu, Y. Induction of hibernation-like hypothermia by central activation of the A1 adenosine receptor in a non-hibernator, the rat. J. Physiol. Sci. 2018, 68, 425–430. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |


| Hamsters | ||||
| Method of Inducing Deep Hypothermia | Decline Rate of Body Temperature | Shift in Splicing Pattern of CIRP | Cardiac Arrhythmias and/or Cardiac Damage | References |
| Natural hibernation | Slow | Yes | No | [13,128] |
| Central administration of adenosine A1-receptor agonist combined with cooling | Slow | Yes | No | [128] |
| Cooling under pentobarbital anesthesia | Rapid | No | Yes | [128] |
| Cooling under isoflurane anesthesia | Rapid | No | Yes | [13] |
| Cooling under isoflurane anesthesia after keeping 30 °C for few hours | Slow | Yes (during keeping 30 ºC) | No | [13] |
| Non-Hibernators (Mice and Rats) | ||||
| Method of Inducing Deep Hypothermia | Decline Rate of Body Temperature | Shift in Splicing Pattern of CIRP | Cardiac Arrhythmias and/or Cardiac Damage | References |
| Central administration of adenosine A1-receptor agonist combined with cooling (Rats) | Slow | Yes | No | [142] |
| Cooling under isoflurane anesthesia (Mice and Rats) | Rapid (until ~ 20 ºC) | No | Lethal | [15] |
| Cooling under isoflurane anesthesia after keeping 30 °C for few hours (Mice and Rats) | Slow | Yes | No | [15] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiina, T.; Shimizu, Y. Temperature-Dependent Alternative Splicing of Precursor mRNAs and Its Biological Significance: A Review Focused on Post-Transcriptional Regulation of a Cold Shock Protein Gene in Hibernating Mammals. Int. J. Mol. Sci. 2020, 21, 7599. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207599
Shiina T, Shimizu Y. Temperature-Dependent Alternative Splicing of Precursor mRNAs and Its Biological Significance: A Review Focused on Post-Transcriptional Regulation of a Cold Shock Protein Gene in Hibernating Mammals. International Journal of Molecular Sciences. 2020; 21(20):7599. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207599
Chicago/Turabian StyleShiina, Takahiko, and Yasutake Shimizu. 2020. "Temperature-Dependent Alternative Splicing of Precursor mRNAs and Its Biological Significance: A Review Focused on Post-Transcriptional Regulation of a Cold Shock Protein Gene in Hibernating Mammals" International Journal of Molecular Sciences 21, no. 20: 7599. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207599