Genome-Wide Identification of the Gossypium hirsutum NHX Genes Reveals That the Endosomal-Type GhNHX4A Is Critical for the Salt Tolerance of Cotton

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
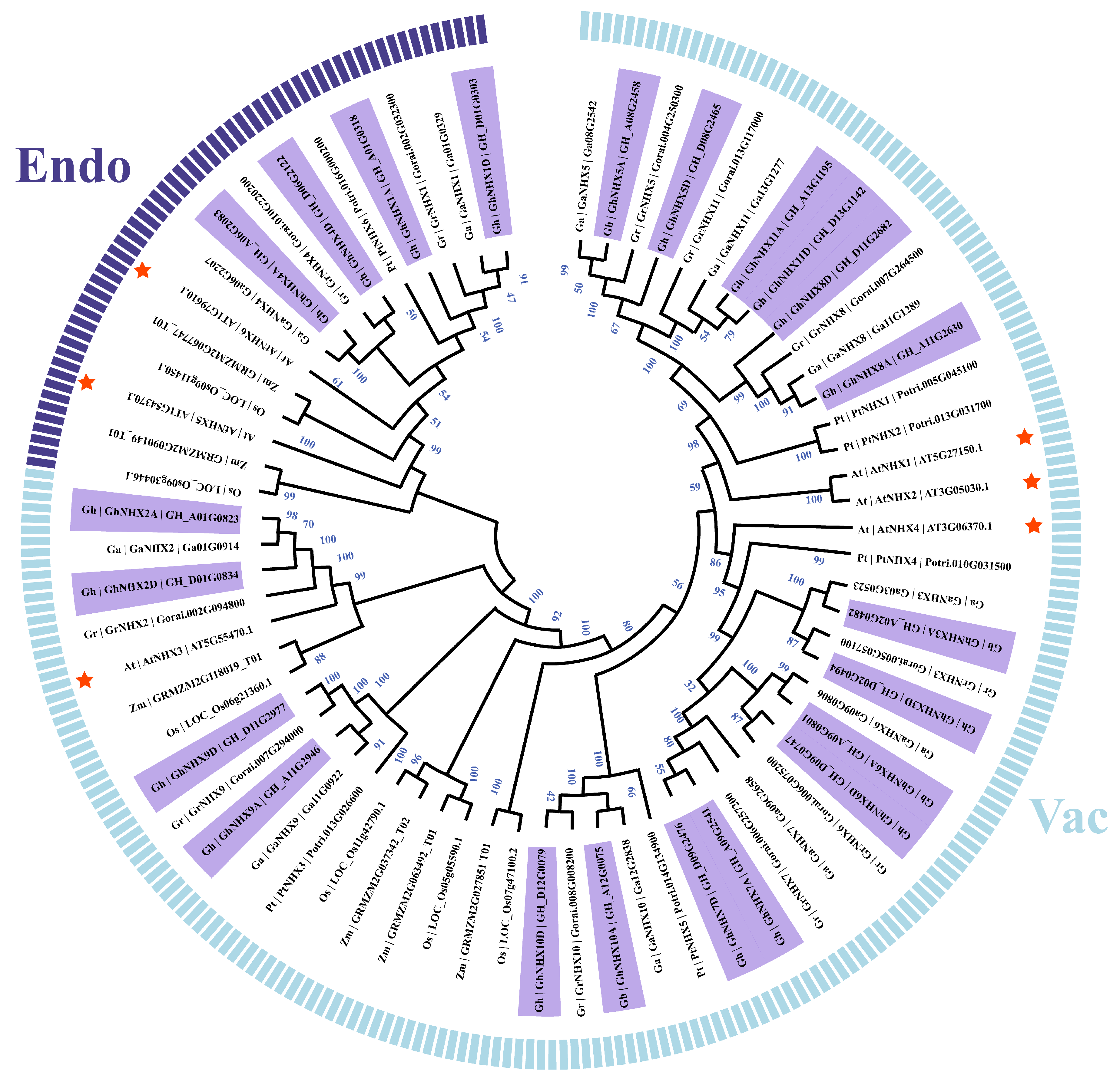
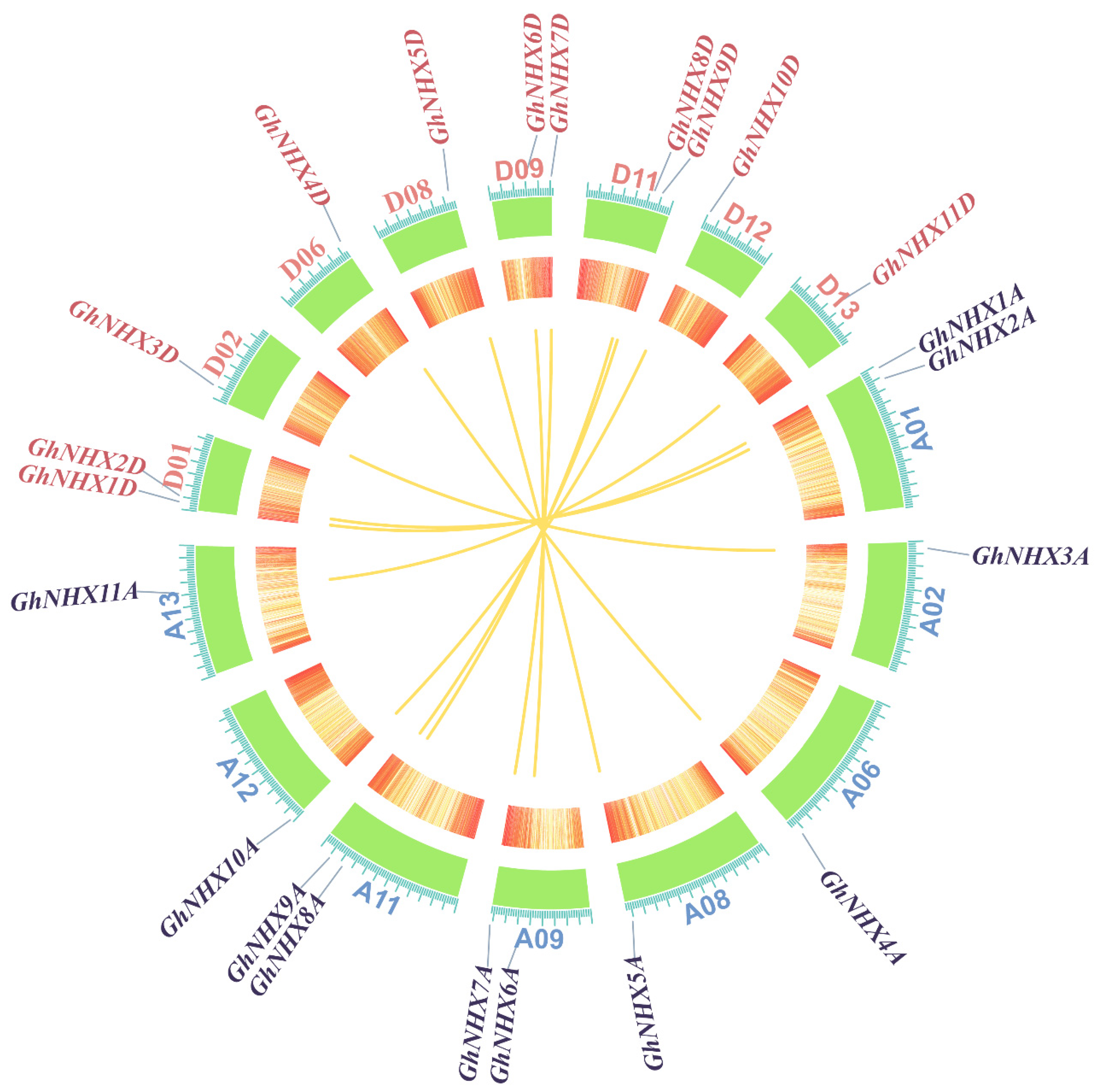
2.1. Genome-Wide Identification and Phylogenetic Relationships of NHX Genes
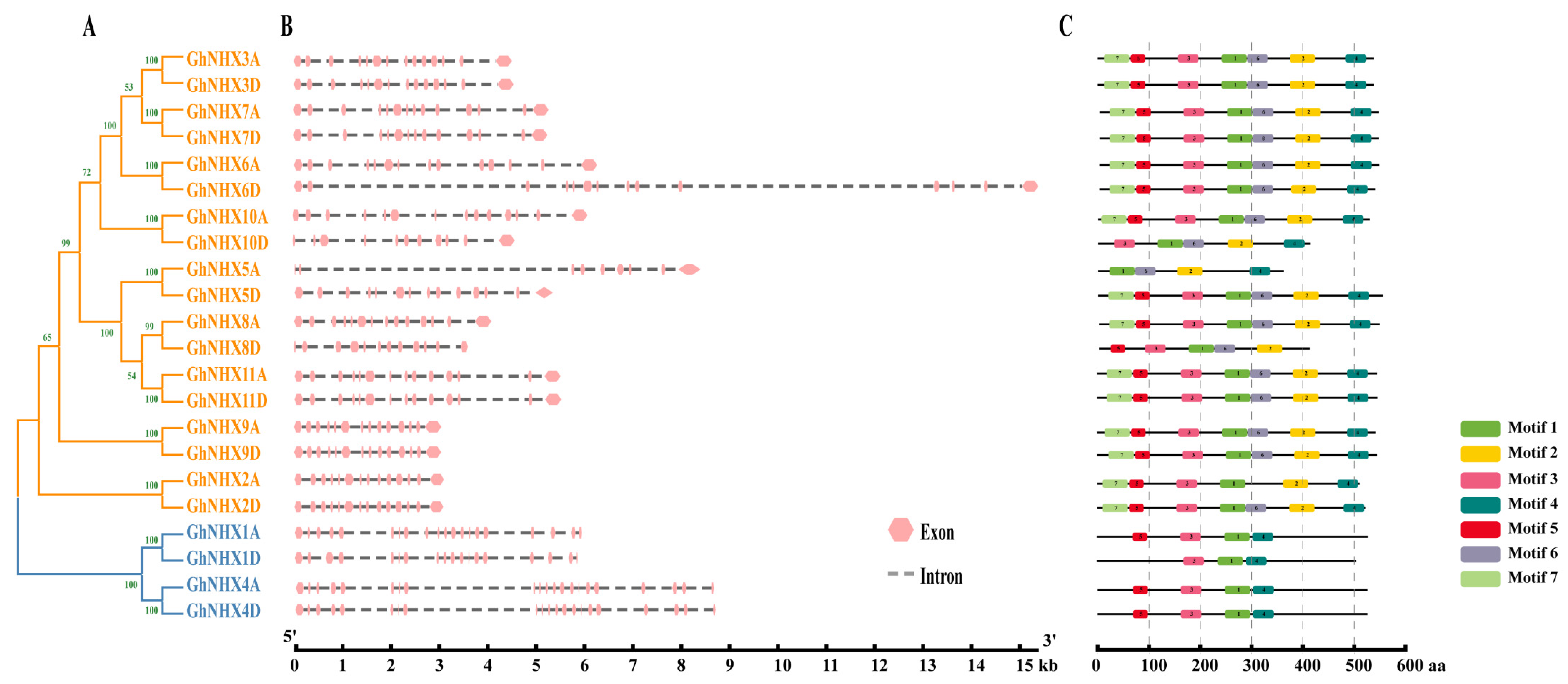
2.2. Gene Structures and Conserved Motifs of NHXs in G. hirsutum
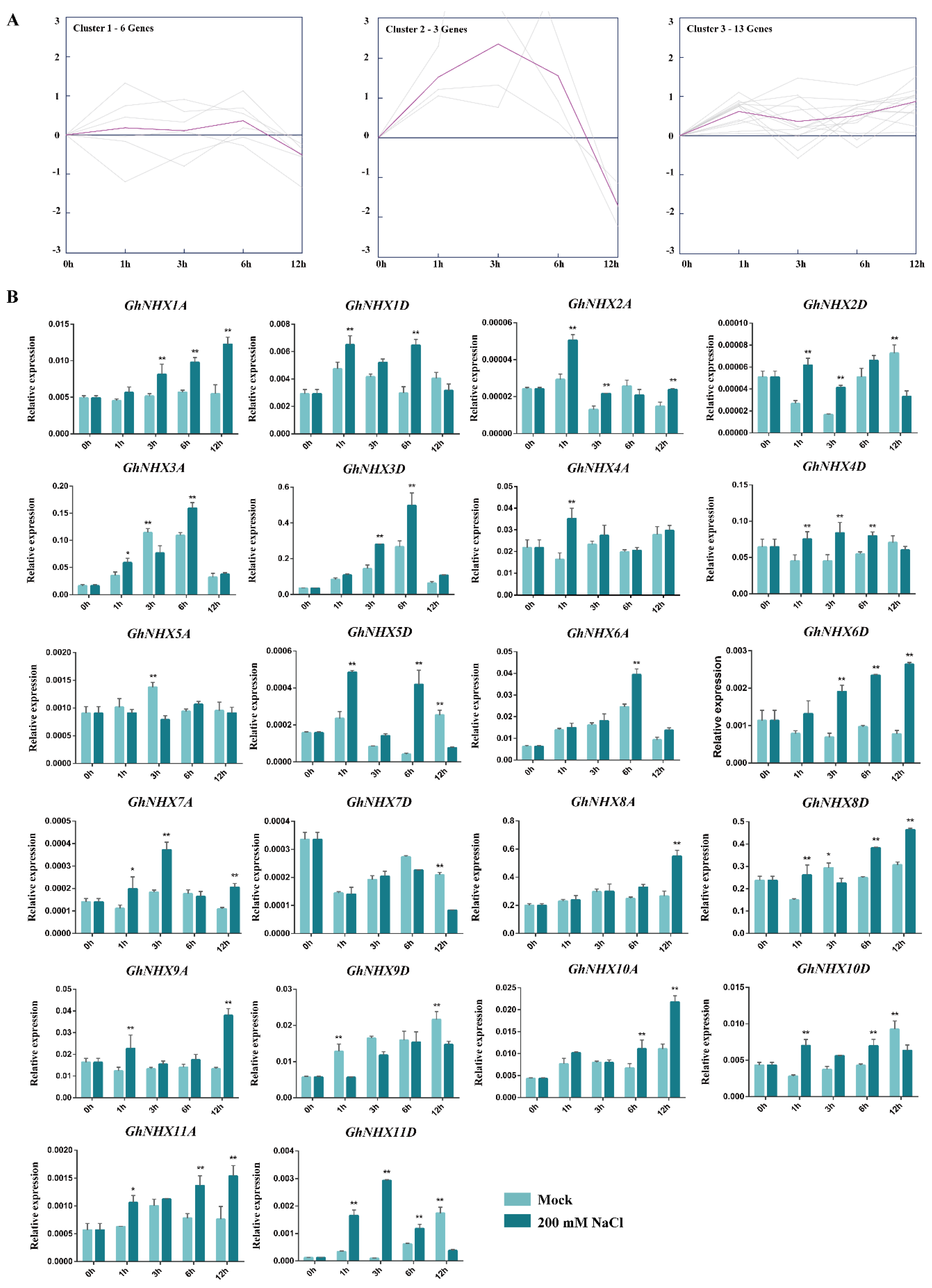
2.3. Expression Patterns of GhNHX Genes under a Salt Treament
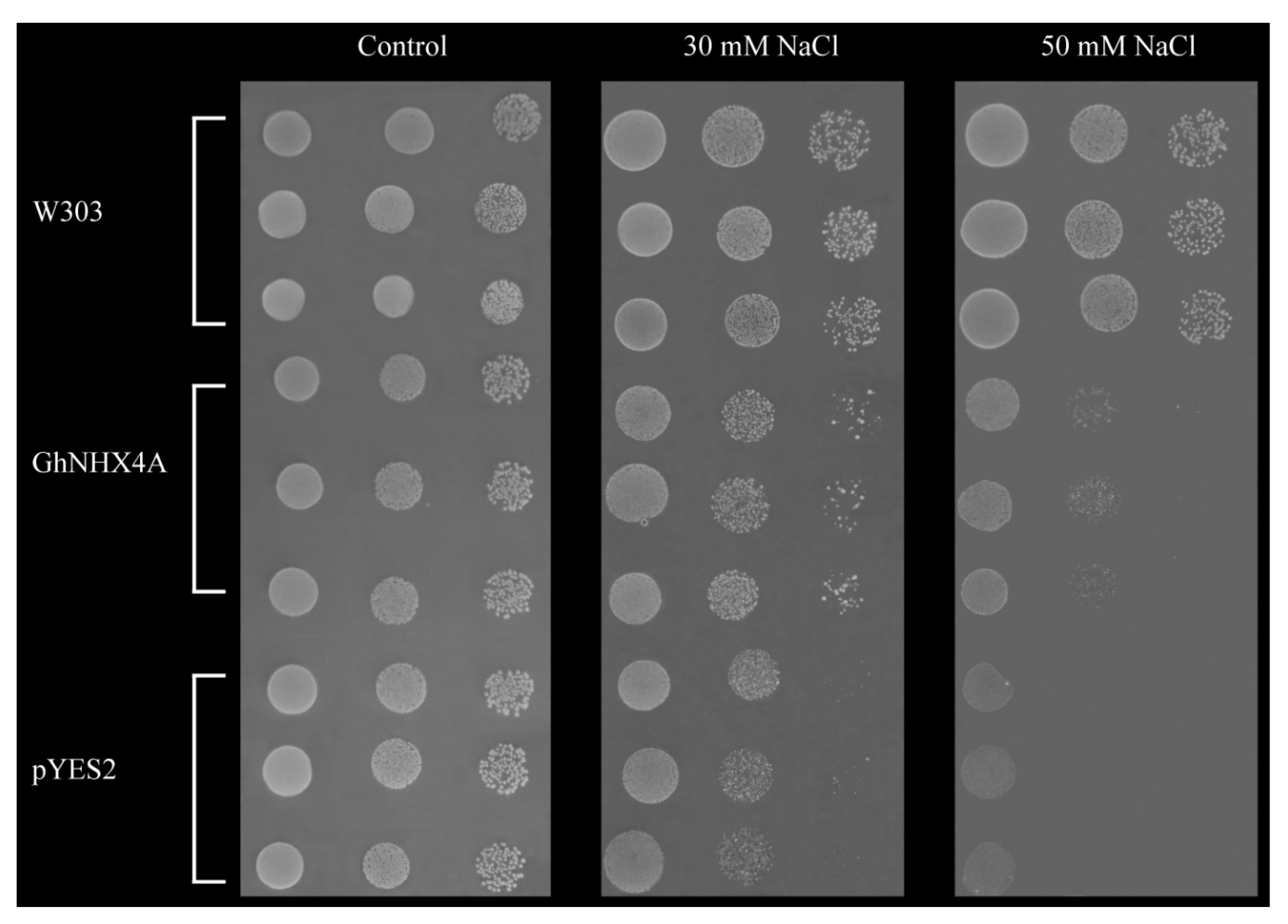
2.4. Functional Complementation of GhNHX4A in Yeast Mutants
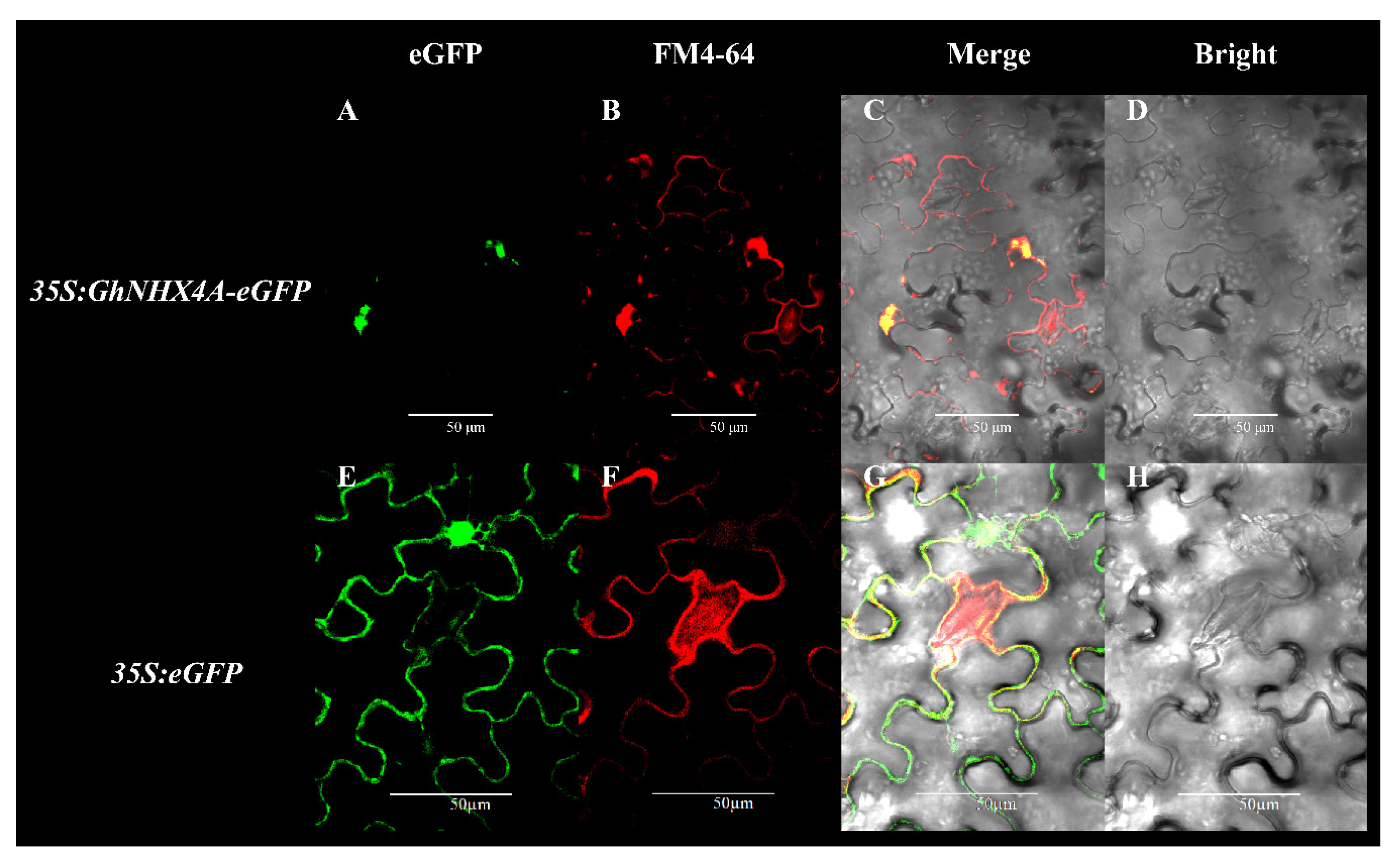
2.5. Subcellular Localization of GhNHX4A
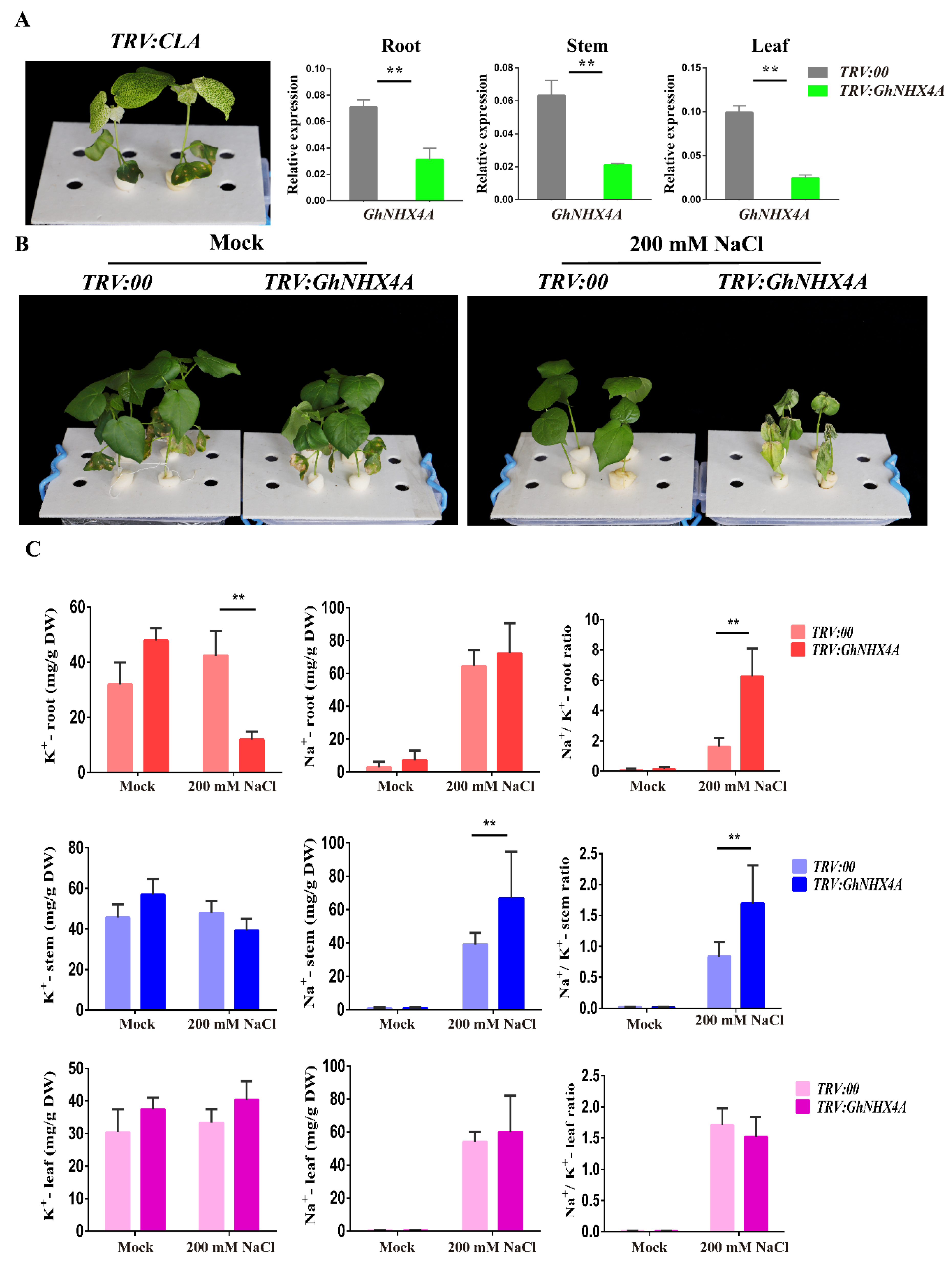
2.6. Virus-Induced Gene Silencing of GhNHX4A in G. hirsutum
3. Discussion
3.1. Cotton NHX Gene Family
3.2. Expression Profiles of GhNHX Genes under the Salt Treatment
3.3. GhNHX4A-Mediated Salt Tolerance
4. Materials and Methods
4.1. Identification of NHX Genes in Gossypium Species
4.2. Phylogenetic Relationship, Chromosomal Localization, Gene Structure, and Conserved Motif Analyses
4.3. Plant Materials and Treaments
4.4. RNA Isolation and Quantitative Real-Time PCR Analysis
4.5. Functional Analysis of GhNHXs Using Yeast Mutants
4.6. Transient Expression of Enhanced Green Fluorescent Protein (eGFP) Constructs in Tobacco Leaves
4.7. Virus-Induced Gene Silencing Analyses in Cotton
4.8. Determination of Intracellular K+ and Na+ Contents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CPA1 | cation/proton antiporter-1 |
| NHX | Na+/H+ antiporter |
| qPCR | quantitative real-time PCR |
| VIGS | virus-induced gene silencing |
References
- Filiz, E.; Ozyigit, I.I.; Saracoglu, I.A.; Uras, M.E.; Sen, U.; Yalcin, B. Abiotic stress-induced regulation of antioxidant genes in different Arabidopsis ecotypes: Microarray data evaluation. Biotechnol. Biotechnol. Equip. 2019, 33, 128–143. [Google Scholar] [CrossRef] [Green Version]
- Kopycińska, M.; Lipa, P.; Cieśla, J.; Kozieł, M.; Janczarek, M. Extracellular polysaccharide protects Rhizobium leguminosarum cells against zinc stress in vitro and during symbiosis with clover. Environ. Microbiol. Rep. 2017, 10, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Parida, A.; Das, A.; Mittra, B. Effects of salt on growth, ion accumulation, photosynthesis and leaf anatomy of the mangrove, Bruguiera parviflora. Trees 2004, 18, 167–174. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuteja, N. Mechanisms of high salinity tolerance in plants. Methods Enzymol. 2007, 428, 419–438. [Google Scholar] [PubMed]
- Sykes, S.R. The inheritance of salt exclusion in woody perennial fruit species. Plant Soil 1992, 50, 123–129. [Google Scholar] [CrossRef]
- Volkmar, K.M.; Hu, Y.; Steppuhn, H. Physiological responses of plants to salinity: A review. Can. J. Plant Sci. 1998, 78, 19–27. [Google Scholar] [CrossRef]
- Michelet, B.; Boutry, M. The plasma membrane H⁺-ATPase: A highly regulated enzyme with multiple physiological functions. Plant Physiol. 1995, 108, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Tang, Z. Enhanced H+ transport activity of tonoplast vesicles isolated from roots of salt-tolerant mutant of wheat under NaCl stress. Chin. Sci. Bull. 1999, 1198–1201. [Google Scholar] [CrossRef]
- Zhu, J.K. Cell signaling under salt, water and cold stresses. Curr. Opin. Plant Biol. 2001, 4, 401–406. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Jensen, R.G. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol. 1996, 14, 89–97. [Google Scholar] [CrossRef]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Fukada-Tanaka, S.; Inagaki, Y.; Saito, N.; Yonekura-Sakakibara, K.; Tanaka, Y.; Kusumi, T.; Iida, S. Genes encoding the vacuolar Na+/H+ exchanger and flower coloration. Plant. Cell Physiol. 2001, 42, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Apse, P.M. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef]
- Brett, C.L.; Donowitz, M.; Rao, R. Evolutionary origins of eukaryotic sodium/proton exchangers. Am. J. Physiol. Cell Physiol. 2005, 288, C223–C239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Rosales, M.P.; Gálvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant. Signaling Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanroj, S.; Wang, G.; Venema, K.; Zhang, M.W.; Delwiche, C.F.; Sze, H. Conserved and diversified gene families of monovalent cation/H+ antiporters from algae to flowering plants. Front. Plant. Sci. 2012, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, S.; Quintero, F.J.; Cubero, B.; Ruiz, M.T.; Pardo, J.M. Differential expression and function of Arabidopsis thaliana NHX Na+/H+ antiporters in the salt stress response. Plant. J. 2002, 30, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Blumwald, E.; Aharon, G.S.; Apse, M.P. Sodium transport in plant cells. Biochim. Biophys. Acta, Biomembr. 2000, 1465, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Rausch, T.; Kirsch, M.; Löw, R.; Lehr, A.; Viereck, R.; Zhigang, A. Salt stress responses of higher plants: The role of proton pumps and Na+/H+ antiporters. J. Plant. Physiol. 1996, 148, 425–433. [Google Scholar] [CrossRef]
- Gaxiola, R.A.; RAO, R.; Sherman, A. The Arabidopsis thaliana proton transporters, AtNltxl and Avpl, can function in cation detoxification in yeast. Proc. Natl. Acad. Sci. USA. 1999, 96, 1480–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.H.; Ma, Q.J.; Liu, X.; Zhu, X.P.; Hu, D.G.; Hao, Y.J. Molecular cloning and functional characterization of MdNHX1 reveals its involvement in salt tolerance in apple calli and Arabidopsis. Sci. Hortic. 2017, 215, 126–133. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, Y.; Hong, S.; Xia, Z.; Cui, D.; Guo, J.; Xu, H.; Jiang, X. Functional characterization of a Wheat NHX antiporter gene TaNHX2 that encodes a K+/H+ exchanger. PLoS ONE 2013, 8, e78098. [Google Scholar] [CrossRef]
- Liu, H.U.A.; Wang, Q.; Yu, M.; Zhang, Y.; Wu, Y.; Zhang, H. Transgenic salt-tolerant sugar beet (Beta vulgaris L.) constitutively expressing an Arabidopsis thaliana vacuolar Na+/H+ antiporter gene, AtNHX3, accumulates more soluble sugar but less salt in storage roots. Plant. Cell Environ. 2008, 31, 1325–1334. [Google Scholar] [CrossRef]
- Tian, N.; Wang, J.; Xu, Z.Q. Overexpression of Na+/H+ antiporter gene AtNHX1 from Arabidopsis thaliana improves the salt tolerance of kiwifruit (Actinidia deliciosa). S. Afr. J. Bot. 2011, 77, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhai, H.; He, S.; Zhang, H.; Ren, Z.; Zhang, D.; Liu, Q. A vacuolar Na+/H+ antiporter gene, IbNHX2, enhances salt and drought tolerance in transgenic sweetpotato. Sci. Hortic. 2016, 201, 153–166. [Google Scholar] [CrossRef]
- Bassil, E.; Ohto, M.A.; Esumi, T.; Tajima, H.; Zhu, Z.; Cagnac, O.; Belmonte, M.; Peleg, Z.; Yamaguchi, T.; Blumwald, E. The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant. Cell 2011, 23, 224–239. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wu, X.; Liu, Y.; Qiu, Q.S.; Van, V.H.W. AtNHX5 and AtNHX6 control cellular K+ and pH homeostasis in Arabidopsis: Three conserved acidic residues are essential for K+ transport. PLoS ONE 2015, 10, e0144716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.R.; Lin, X.J.; Li, H.Q.; Pan, X.P.; Wu, G.J. Overexpression of AtNHX5 improves tolerance to both saltand water stress in rice (Oryza sativa L.). Plant. Cell Tissue Organ. Cult. 2011, 107, 283–293. [Google Scholar] [CrossRef]
- Rodríguez-Rosales, M.P.; Jiang, X.; Gálvez, F.J.; Aranda, M.N.; Cubero, B.; Venema, K. Overexpression of the tomato K+/H+ antiporter LeNHX2 confers salt tolerance by improving potassium compartmentalization. New Phytol. 2008, 179, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Sunilkumar, G.; Campbell, L.; Puckhaber, L.; Stipanovic, R.; Rathore, K. Engineering cottonseed for use in human nutrition by tissue-specific reduction of toxic gossypol. Proc. Natl. Acad. Sci. USA 2006, 103, 18054–18059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grover, C.E.; Gallagher, J.P.; Jareczek, J.J.; Page, J.T.; Wendel, J.F. Re-evaluating the phylogeny of allopolyploid Gossypium, L. Mol. Phylogenet. Evol. 2015, 92, 45–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Azhar, F.M.; Khan, I.A.; EhsanUllah. Variability for drought tolerance in cotton (Gossypium hirsutum) and its genetic basis. Int. J. Agric. Biol. 2011, 13, 61–66. [Google Scholar]
- Fu, X.; Lu, Z.; Wei, H.; Zhang, J.; Yang, X.; Wu, A.; Ma, L.; Kang, M.; Lu, J.; Wang, H.; et al. Genome-wide identification and expression analysis of the NHX (sodium/hydrogen antiporter) gene family in cotton. Front. Genet. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.A.; Yang, G.D.; Meng, Q.W.; Zheng, C.C. The cotton GhNHX1 gene encoding a novel putative tonoplast Na+/H+ antiporter plays an important role in salt stress. Plant. Cell Physiol. 2004, 45, 600–607. [Google Scholar] [CrossRef] [Green Version]
- He, C. Expression of an Arabidopsis vacuolar sodium/proton antiporter gene in cotton improves photosynthetic performance under salt conditions and increases fiber yield in the field. Plant. Cell Physiol. 2005, 46, 1848–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, U.; Song, Y.; Liang, C.; Abid, M.A.; Askari, M.; Myat, A.A.; Abbas, M.; Malik, W.; Ali, Z.; Guo, S.; et al. Genome-wide characterization and expression analysis of NHX gene family under salinity stress in Gossypium barbadense and its comparison with Gossypium hirsutum. Genes 2020, 11, 803. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K.; Siu, C.L.; Hillmer, S.; Jang, S.; An, G.; Robinson, D.G.; Jiang, L. Rice SCAMP1 defines clathrin-coated, trans-golgi-located tubular-vesicular structures as an early endosome in tobacco BY-2 cells. Plant. Cell 2007, 19, 296–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enrico, M.; Masayoshi, M.; Ekkehard, N.H. Vacuolar transporters and their essential role in plant metabolism. J. Exp. Bot. 2007, 58, 83–102. [Google Scholar]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta 2011, 233, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Noll, A.; Karl, S.; Leib, K.; Yan, F.; Schubert, S. Molecular characterization of Na+/H+ antiporters (ZmNHX) of maize (Zea mays L.) and their expression under salt stress. J. Plant. Physiol. 2005, 162, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Chang, E.; Li, Y.; Sun, P.; Hu, J.; Zhang, J. Expression and integrated network analyses revealed functional divergence of NHX-type Na+/H+ exchanger genes in poplar. Sci. Rep. 2017, 7, 2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendel, J.F. Genome evolution in polyploids. Plant. Mol. Biol. 2000, 42, 225–249. [Google Scholar] [CrossRef]
- Kondrashov, F.A.; Rogozin, I.B.; Wolf, Y.I.; Koonin, E.V. Selection on the evolution of gene duplications. Genome Biol. 2002, 3, research0008.0001. [Google Scholar] [CrossRef] [Green Version]
- Conant, G.C.; Wolfe, K.H. Turning a hobby into a job: How duplicated genes find new functions. Nat. Rev. Genet. 2008, 9, 938–950. [Google Scholar] [CrossRef]
- Ramsey, J.; Schemske, D.W. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef] [Green Version]
- Flagel, L.E.; Wendel, J.F. Gene duplication and evolutionary novelty in plants. New Phytol. 2009, 183, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Long, D.; Zhang, M.; Liu, C.; Xiang, Z.; Zhao, A. Molecular characterization and expression analysis of the mulberry Na+/H+ exchanger gene family. Plant. Physiol. Biochem. 2016, 99, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, D.; Pudussery, M.V.; Kaundal, R.; Suarez, D.L.; Kaundal, A.; Sekhon, R.S. Molecular characterization and expression analysis of the Na+/H+ exchanger gene family in Medicago truncatula. Funct. Integr. Genom. 2018, 18, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Q.; Wang, J.L.; Li, S.J. Genome-wide identification of Na+/H+ antiporter (NHX) genes in Sugar Beet (Beta vulgaris L.) and their regulated expression under salt stress. Genes 2019, 10, 401. [Google Scholar] [CrossRef] [Green Version]
- Jia, Q.; Zheng, C.; Sun, S.; Amjad, H.; Liang, K.; Lin, W. The role of plant cation/proton antiporter gene family in salt tolerance. Biol. Plant 2018, 62, 617–629. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Z.; Liu, Q.; Tang, J.; Huang, S.; Dhankher, O.P.; Yuan, H. Growth, physiological adaptation, and NHX gene expression analysis of Iris halophila under salt stress. Environ. Sci. Pollut. Res. 2018, 25, 25207–25216. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, E.; Blumwald, E.; Pedro Donaire, J.; Belver, A. Na+/H+ antiport activity in tonoplast vesicles isolated from sunflower roots induced by NaCl stress. Physiol. Plant 1997, 99, 328–334. [Google Scholar] [CrossRef]
- Ye, C.Y.; Zhang, H.C.; Chen, J.H.; Xia, X.L.; Yin, W.L. Molecular characterization of putative vacuolar NHX-type Na+/H+ exchanger genes from the salt-resistant tree Populus euphratica. Physiol. Plant 2009, 137, 166–174. [Google Scholar] [CrossRef]
- Wang, J.; Qiu, N.; Wang, P.; Zhang, W.; Yang, X.; Chen, M.; Wang, B.; Sun, J. Na+ compartmentation strategy of Chinese cabbage in response to salt stress. Plant. Physiol. Biochem. 2019, 140, 151–157. [Google Scholar] [CrossRef]
- Razzouk, S.; Whittington, W.J. Effects of salinity on cotton yield and quality. Field Crops Res. 1991, 26, 305–314. [Google Scholar] [CrossRef]
- Saghir, A.; Noor-Ul-Islam, K.; Iqbal, M.Z.; Altaf, H.; Mahmudul, H. Salt tolerance of cotton ( Gossypium hirsutum L.). Asian, J. Plant Sci. 2002, 1, 715–719. [Google Scholar]
- Darley, C.P.; Wuytswinkel, O.C.M.V.; Woude, K.V.D.; Mager, W.H.; Boer, A.H.D. Arabidopsis thaliana and Saccharomyces cerevisiae NHX1 genes encode amiloride sensitive electroneutral Na+/H+ exchangers. Biochem. J. 2000, 351, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [Green Version]
- Venema, K.; Belver, A.; Marin-Manzano, M.C.; Rodriguez-Rosales, M.P.; Donaire, J.P. A novel intracellular K+/H+ antiporter related to Na+/H+ antiporters is important for K+ ion homeostasis in plants. J. Biol. Chem. 2003, 278, 22453–22459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Fu, J.; Yu, C.; Wang, X.; Jiang, Q.; Hong, J.; Lu, K.; Xue, G.; Yan, C.; James, A.; et al. Increasing cyclic electron flow is related to Na+ sequestration into vacuoles for salt tolerance in soybean. J. Exp. Bot. 2015, 66, 6877–6889. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Deng, G.; Wang, H.; Zhang, H.; Zhang, P. Elevated compartmentalization of Na+ into vacuoles improves salt and cold stress tolerance in sweet potato (Ipomoea batatas). Physiol Plant. 2015, 154, 560–571. [Google Scholar] [PubMed]
- De Luca, A.; Pardo, J.M.; Leidi, E.O. Pleiotropic effects of enhancing vacuolar K+/H+ exchange in tomato. Physiol. Plant. 2018, 163, 88–102. [Google Scholar] [CrossRef]
- Xue, Z.Y.; Zhi, D.Y.; Xue, G.P.; Zhang, H.; Zhao, Y.X.; Xia, G.M. Enhanced salt tolerance of transgenic wheat (Tritivum aestivum L.) expressing a vacuolar Na+/H+ antiporter gene with improved grain yields in saline soils in the field and a reduced level of leaf Na+. Plant. Sci. 2004, 167, 849–859. [Google Scholar] [CrossRef]
- Fukuda, A.; Chiba, K.; Maeda, M.; Nakamura, A.; Maeshima, M.; Tanaka, Y. Effect of salt and osmotic stresses on the expression of genes for the vacuolar H+-pyrophosphatase, H+-ATPase subunit A, and Na+/H+ antiporter from barley. J. Exp. Bot. 2004, 55, 585–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; Hou, L.; Kong, X.Q.; Zhao, Y.X.; Zhang, H. Overexpression of AtNHX5 increases salt tolerance of Arabidopsis thaliana. Xibei Zhiwu Xuebao 2012, 32, 1106–1111. [Google Scholar]
- Shi, L.-Y.; Li, H.-Q.; Pan, X.-P.; Wu, G.-J.; Li, M.-R. Improvement of Torenia fournieri salinity tolerance by expression of Arabidopsis AtNHX5. Funct. Plant. Biol. 2008, 35, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Liang, C.; Meng, Z.; Sun, G.; Meng, Z.; Guo, S.; Zhang, R. CottonFGD: An integrated functional genomics database for cotton. BMC Plant. Biol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hima Kumari, P.; Anil Kumar, S.; Ramesh, K.; Sudhakar Reddy, P.; Nagaraju, M.; Bhanu Prakash, A.; Shah, T.; Henderson, A.; Srivastava, R.K.; Rajasheker, G.; et al. Genome-wide identification and analysis of Arabidopsis sodium proton antiporter (NHX) and human sodium proton exchanger (NHE) homologs in Sorghum bicolor. Genes 2018, 9, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.; Hotz, H.R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L. The Pfam protein families database. Nucleic Acids Res. 2008, 36, D281–D288. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018, 289660. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Nadya, W.; Chris, M.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, W.; Ren, Z.; Zhou, Y.; Zhao, J.; Zhang, F.; Feng, J.; Liu, W.; Ma, X. Genome-Wide Identification of the Gossypium hirsutum NHX Genes Reveals That the Endosomal-Type GhNHX4A Is Critical for the Salt Tolerance of Cotton. Int. J. Mol. Sci. 2020, 21, 7712. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207712
Ma W, Ren Z, Zhou Y, Zhao J, Zhang F, Feng J, Liu W, Ma X. Genome-Wide Identification of the Gossypium hirsutum NHX Genes Reveals That the Endosomal-Type GhNHX4A Is Critical for the Salt Tolerance of Cotton. International Journal of Molecular Sciences. 2020; 21(20):7712. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207712
Chicago/Turabian StyleMa, Wenyu, Zhongying Ren, Yang Zhou, Junjie Zhao, Fei Zhang, Junping Feng, Wei Liu, and Xiongfeng Ma. 2020. "Genome-Wide Identification of the Gossypium hirsutum NHX Genes Reveals That the Endosomal-Type GhNHX4A Is Critical for the Salt Tolerance of Cotton" International Journal of Molecular Sciences 21, no. 20: 7712. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207712