Chemical Starting Matter for HNF4α Ligand Discovery and Chemogenomics


 and
and
Abstract
:1. Introduction
2. Results
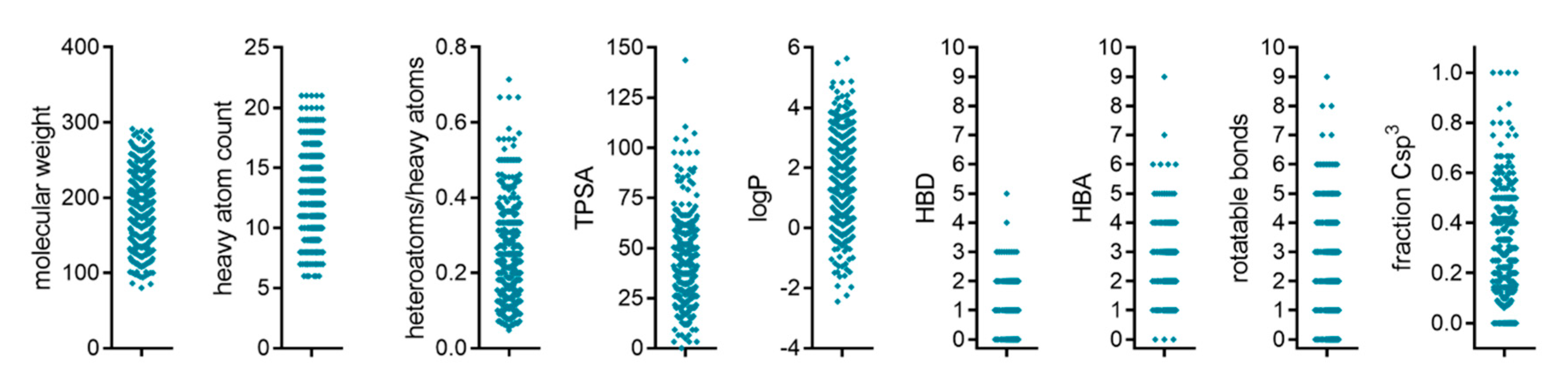
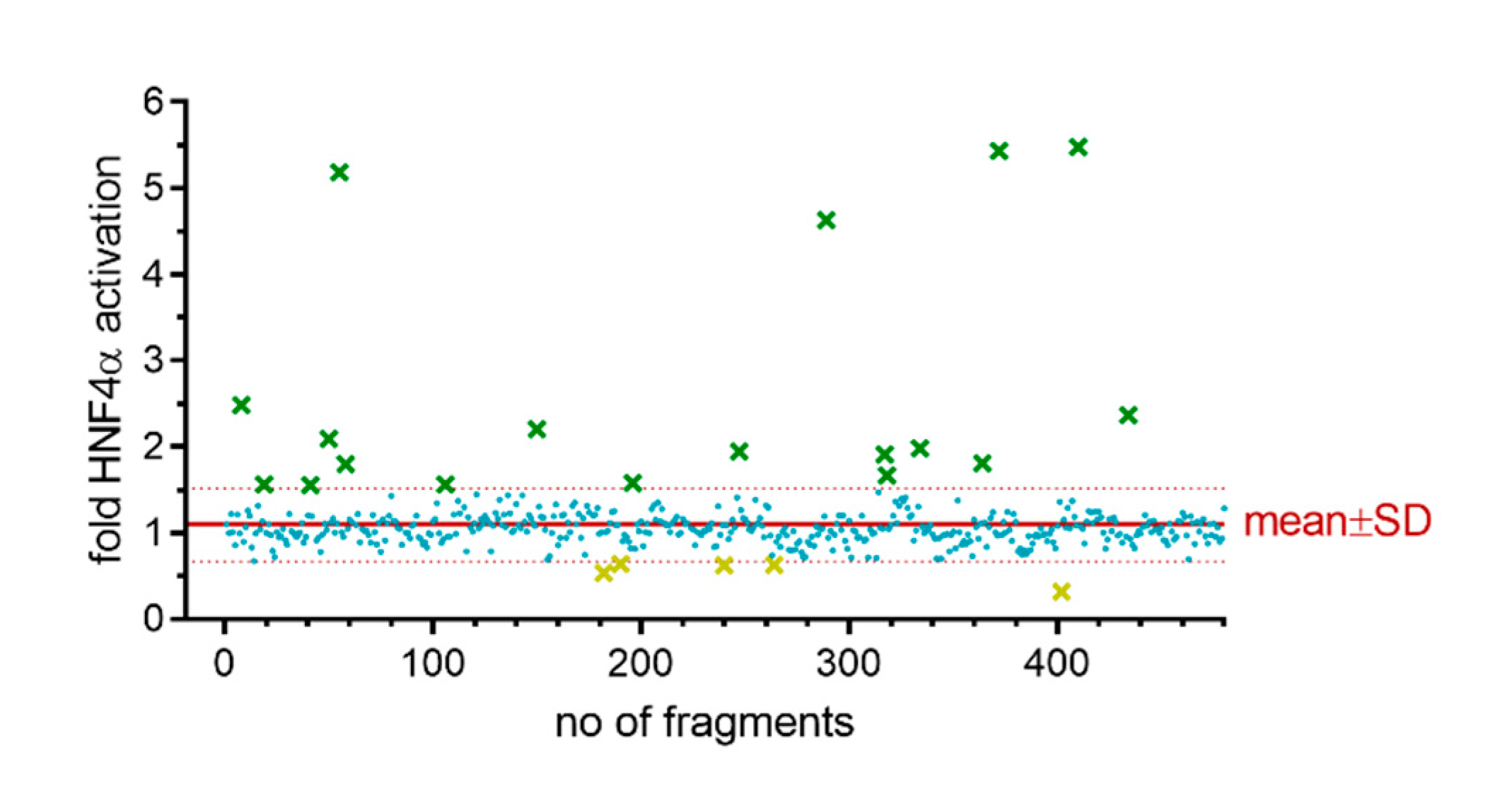
2.1. Primary Screening
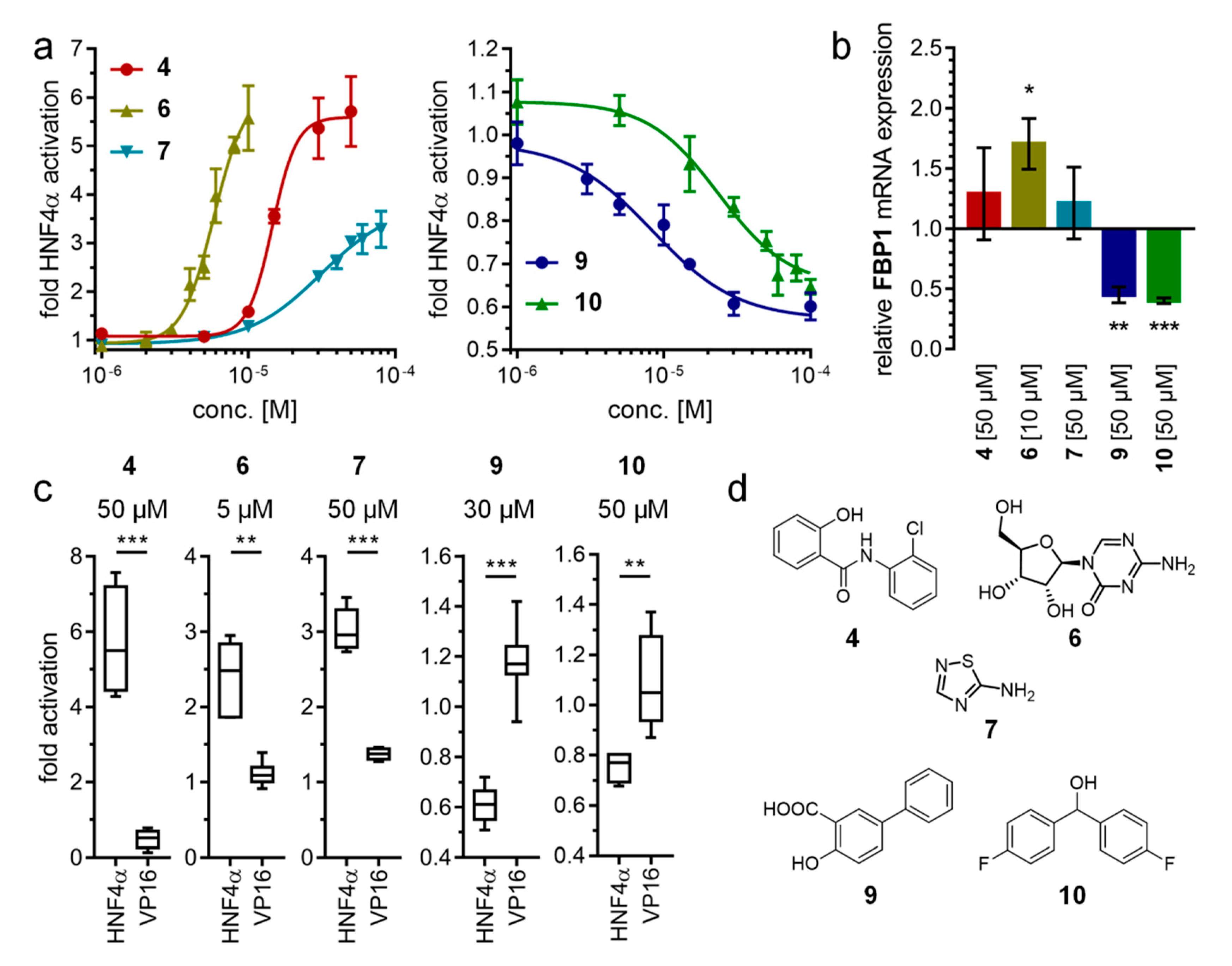
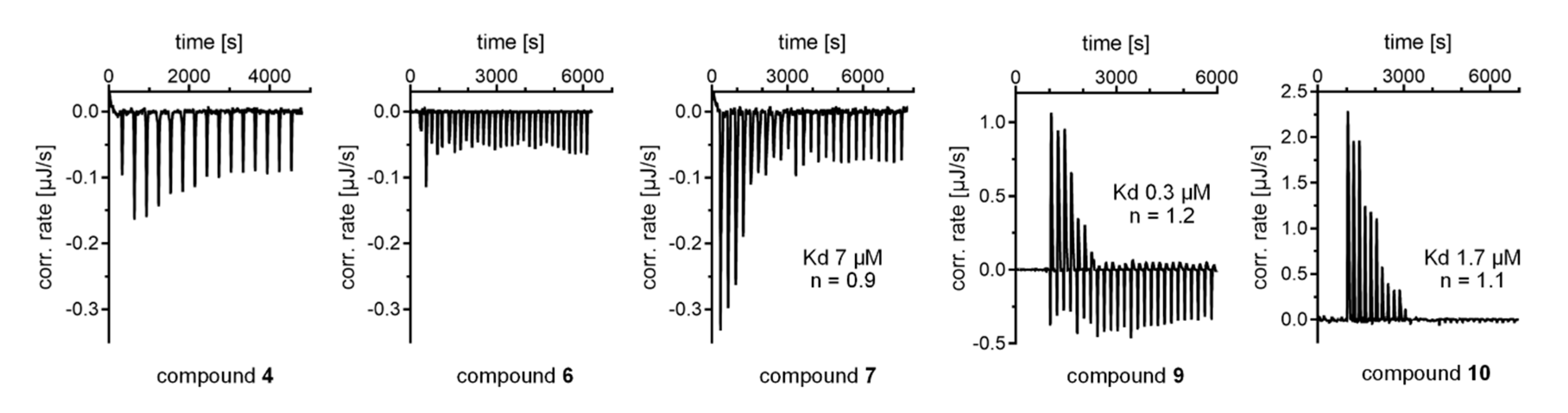
2.2. Hit Validation
3. Discussion
4. Materials and Methods
4.1. Hybrid Gal4-HNF4α Reporter Gene Assay
4.2. Recombinant HNF4α LBD Expression and Purification
4.3. Quantification of Human FBP1 mRNA Expression in HepG2 Cells by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.4. Isothermal Titration Calorimetry
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FBP1 | fructose-1,6-bisphosphatase 1 |
| HNF4α | hepatocyte nuclear factor 4α |
| LBD | Ligand-binding domain |
| MODY | maturity-onset diabetes of the young |
| RLU | relative light units |
References
- Alexander, S.P.; Cidlowski, J.A.; Kelly, E.; Marrion, N.V.; Peters, J.A.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Sharman, J.L.; Southan, C.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: Nuclear hormone receptors. Br. J. Pharmacol. 2017, 174, S208–S224. [Google Scholar] [CrossRef] [Green Version]
- Benoit, G.; Cooney, A.; Giguere, V.; Ingraham, H.; Lazar, M.; Muscat, G.; Perlmann, T.; Renaud, J.P.; Schwabe, J.; Sladek, F.; et al. International union of pharmacology. LXVI. Orphan nuclear receptors. Pharmacol. Rev. 2006, 58, 798–836. [Google Scholar] [CrossRef] [Green Version]
- Chandra, V.; Huang, P.; Potluri, N.; Wu, D.; Kim, Y.; Rastinejad, F. Multidomain integration in the structure of the HNF-4α nuclear receptor complex. Nature 2013, 495, 394–398. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Ta, T.C.; Lin, M.; Evans, J.R.; Dong, Y.; Bolotin, E.; Sherman, M.A.; Forman, B.M.; Sladek, F.M. Identification of an endogenous ligand bound to a native orphan nuclear receptor. PLoS ONE 2009, 4, e5609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiselyuk, A.; Lee, S.H.; Farber-Katz, S.; Zhang, M.; Athavankar, S.; Cohen, T.; Pinkerton, A.B.; Ye, M.; Bushway, P.; Richardson, A.D.; et al. HNF4α antagonists discovered by a high-throughput screen for modulators of the human insulin promoter. Chem. Biol. 2012, 19, 806–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Tanaka, T.; Iwanari, H.; Hotta, H.; Yamashita, H.; Kumakura, J.; Wanatabe, Y.; Uchiyama, Y.; Aburatani, H.; Hamakubo, T.; et al. Expression and localization of P1 promoter-driven hepatocyte nuclear factor-4α (HNF4α) isoforms in human and rats. Nucl. Recept. 2003, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Vasoya, R.P.; Toke, N.H.; Parthasarathy, A.; Luo, S.; Chiles, E.; Flores, J.; Gao, N.; Bonder, E.M.; Su, X.; et al. HNF4 Regulates Fatty Acid Oxidation and Is Required for Renewal of Intestinal Stem Cells in Mice. Gastroenterology 2020, 158, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Hayhurst, G.P.; Lee, Y.-H.; Lambert, G.; Ward, J.M.; Gonzalez, F.J. Hepatocyte Nuclear Factor 4α (Nuclear Receptor 2A1) Is Essential for Maintenance of Hepatic Gene Expression and Lipid Homeostasis. Mol. Cell. Biol. 2001, 21, 1393–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.K.; Gao, N.; Gorski, R.K.; White, P.; Hardy, O.T.; Rafiq, K.; Brestelli, J.E.; Chen, G.; Stoeckert, C.J.; Kaestner, K.H. Expansion of adult β-cell mass in response to increased metabolic demand is dependent on HNF-4α. Genes Dev. 2007, 21, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Warncke, K.; Kummer, S.; Raile, K.; Grulich-Henn, J.; Woelfle, J.; Steichen, E.; Prinz, N.; Holl, R.W. Frequency and Characteristics of MODY 1 (HNF4A Mutation) and MODY 5 (HNF1B Mutation): Analysis from the DPV Database. J. Clin. Endocrinol. Metab. 2018, 104, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.H.; Shah, Y.M.; Inoue, J.; Morimura, K.; Kim, I.; Yim, S.H.; Lambert, G.; Kurotani, R.; Nagashima, K.; Gonzalez, F.J.; et al. Hepatocyte nuclear factor 4α in the intestinal epithelial cells protects against inflammatory bowel disease. Inflamm. Bowel Dis. 2008, 14, 908–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Wei, X.; Wu, Z. HNF-4α promotes multidrug resistance of gastric cancer cells through the modulation of cell apoptosis. Oncol. Lett. 2017, 14, 6477–6484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.M.; Jung, Y.Y.; Hwang, C.J.; Park, M.H.; Yoon, N.Y.; Kim, T.M.; Yu, J.M.; Kim, D.H.; Seo, D.W.; Youn, H.S.; et al. Anti-cancer effect of N-(3,5-bis(trifluoromethyl)phenyl)-5-chloro-2,3-dihydronaphtho[1,2-b]furan-2-carboxamide, a novel synthetic compound. Mol. Carcinog. 2016, 55, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Kaestner, K.H. HNF-4α: From MODY to late-onset type 2 diabetes. Trends Mol. Med. 2004, 10, 521–524. [Google Scholar] [CrossRef]
- Anik, A.; Çatli, G.; Abaci, A.; Böber, E. Maturity-onset diabetes of the young (MODY): An update. J. Pediatr. Endocrinol. Metab. 2015, 28, 251–263. [Google Scholar] [CrossRef]
- Colclough, K.; Bellanne-Chantelot, C.; Saint-Martin, C.; Flanagan, S.E.; Ellard, S. Mutations in the Genes Encoding the Transcription Factors Hepatocyte Nuclear Factor 1 Alpha and 4 Alpha in Maturity-Onset Diabetes of the Young and Hyperinsulinemic Hypoglycemia. Hum. Mutat. 2013, 34, 669–685. [Google Scholar] [CrossRef]
- Le Guével, R.; Oger, F.; Lecorgne, A.; Dudasova, Z.; Chevance, S.; Bondon, A.; Barath, P.; Simonneaux, G.; Salbert, G. Identification of small molecule regulators of the nuclear receptor HNF4α based on naphthofuran scaffolds. Bioorg. Med. Chem. 2009, 17, 7021–7030. [Google Scholar] [CrossRef]
- Heering, J.; Merk, D. Hybrid Reporter Gene Assays: Versatile In Vitro Tools to Characterize Nuclear Receptor Modulators. In Methods in Molecular Biology (Clifton, N.J.); Humana: New York, NY, USA, 2019; Volume 1966, pp. 175–192. [Google Scholar]
- Wattanavanitchakorn, S.; Rojvirat, P.; Chavalit, T.; MacDonald, M.J.; Jitrapakdee, S. CCAAT-enhancer binding protein-α (C/EBPα) and hepatocyte nuclear factor 4α (HNF4α) regulate expression of the human fructose-1,6-bisphosphatase 1 (FBP1) gene in human hepatocellular carcinoma HepG2 cells. PLoS ONE 2018, 13, e0194252. [Google Scholar] [CrossRef]
- Tetko, I.V.; Tanchuk, V.Y. Application of associative neural networks for prediction of lipophilicity in ALOGPS 2.1 program. J. Chem. Inf. Comput. Sci. 2002, 42, 1136–1145. [Google Scholar] [CrossRef]
- Hopkins, A.L.; Keserü, G.M.; Leeson, P.D.; Rees, D.C.; Reynolds, C.H. The role of ligand efficiency metrics in drug discovery. Nat. Rev. Drug Discov. 2014, 13, 105–121. [Google Scholar] [CrossRef]
- Flesch, D.; Cheung, S.-Y.; Schmidt, J.; Gabler, M.; Heitel, P.; Kramer, J.S.; Kaiser, A.; Hartmann, M.; Lindner, M.; Lüddens-Dämgen, K.; et al. Non-acidic farnesoid X receptor modulators. J. Med. Chem. 2017, 60, 7199–7205. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Rotter, M.; Weiser, T.; Wittmann, S.; Weizel, L.; Kaiser, A.; Heering, J.; Goebel, T.; Angioni, C.; Wurglics, M.; et al. A dual modulator of farnesoid X receptor and soluble epoxide hydrolase to counter nonalcoholic steatohepatitis. J. Med. Chem. 2017, 60, 7703–7724. [Google Scholar] [CrossRef]
- Willems, S.; Kilu, W.; Ni, X.; Chaikuad, A.; Knapp, S.; Heering, J.; Merk, D. The orphan nuclear receptor Nurr1 is responsive to non-steroidal anti-inflammatory drugs. Commun. Chem. 2020, 3, 85. [Google Scholar] [CrossRef]
- Sadowski, I.; Ma, J.; Triezenberg, S.; Ptashne, M. GAL4-VP16 is an unusually potent transcriptional activator. Nature 1988, 335, 563–564. [Google Scholar] [CrossRef] [PubMed]
- Budzyński, M.A.; Puustinen, M.C.; Joutsen, J.; Sistonen, L. Uncoupling Stress-Inducible Phosphorylation of Heat Shock Factor 1 from Its Activation. Mol. Cell. Biol. 2015, 35, 2530–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, J.; Klingler, F.-M.; Proschak, E.; Steinhilber, D.; Schubert-Zsilavecz, M.; Merk, D. NSAIDs Ibuprofen, Indometacin, and Diclofenac do not interact with Farnesoid X Receptor. Sci. Rep. 2015, 5, 14782. [Google Scholar] [CrossRef]
- Merk, D.; Lamers, C.; Ahmad, K.; Carrasco Gomez, R.; Schneider, G.; Steinhilber, D.; Schubert-Zsilavecz, M. Extending the structure-activity relationship of anthranilic acid derivatives as farnesoid X receptor modulators: Development of a highly potent partial farnesoid X receptor agonist. J. Med. Chem. 2014, 57, 8035–8055. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Structure | Primary Screen (Fold Act., 50 µM) | Follow-up HNF4α | VP16 Control | HNF4α vs. VP16 |
|---|---|---|---|---|---|
| 1 |  | 2.48 | 1.1 ± 0.2 (50 µM) | 0.80 ± 0.07 (50 µM) | n.s. |
| 2 |  | 1.55 | 1.3 ± 0.2 (50 µM) | 1.3 ± 0.3 (50 µM) | n.s. |
| 3 |  | 2.09 | 1.5 ± 0.2 (50 µM) | 1.7 ± 0.3 (50 µM) | n.s. |
| 4 |  | 5.18 | 4 ± 2 (50 µM) | 0.5 ± 0.2 (50 µM) | p = 0.0014 (**) |
| 5 |  | 1.80 | 2.5 ± 0.2 (100 µM) | 1.1 ± 0.1 (100 µM) | p < 0.0001 (***) |
| 6 |  | 5.48 | 2.4 ± 0.5 (5 µM) | 1.1 ± 0.1 (5 µM) | p = 0.0034 (**) |
| 7 |  | 2.36 | 3.0 ± 0.3 (50 µM) | 1.4 ± 0.1 (50 µM) | p < 0.0001 (***) |
| 8 |  | 2.09 | 1.1 ± 0.2 (50 µM) | 0.92 ± 0.07 (50 µM) | n.s. |
| 9 |  | 0.54 | 0.6 ± 0.1 (30 µM) | 1.2 ± 0.2 (30 µM) | p < 0.0001 (***) |
| 10 |  | 0.63 | 0.75 ± 0.05 (50 µM) | 1.1 ± 0.2 (50 µM) | p = 0.0057 (**) |
| 11 |  | 0.32 | 0.21 ± 0.03 (100 µM) | 0.21 ± 0.02 (100 µM) | n.s. |
| ID | Structure | HNF4α Ligand Type | Validated HNF4α Modulation | HNF4α LBD Binding |
|---|---|---|---|---|
| 4 |  | Agonist | EC50 15 ± 1 µM 5.6 ± 0.3-fold act. | no/weak binding |
| 5 |  | Agonist | EC50 > 100 µM | n.d. |
| 6 |  | Agonist | EC50 5.8 ± 0.6 µM 6.1 ± 0.7-fold act. | no binding |
| 7 |  | Agonist | EC50 31 ± 8 µM 3.8 ± 0.6-fold act. | Kd 7 µM |
| 9 |  | Inverse agonist | IC50 8 ± 2 µM 0.57 ± 0.04 rem. act. | Kd 0.3 µM |
| 10 |  | Inverse agonist | IC50 24 ± 5 µM 0.65 ± 0.05 rem. act. | Kd 1.7 µM |
| ID | HNF4α Activity | LogP | LogS | LE | LLE | SILE |
|---|---|---|---|---|---|---|
| 4 | pEC50 4.8 | 4.13 | −3.72 | 0.39 | 0.69 | 2.1 |
| 5 | pEC50 < 4 | 4.84 | −5.50 | <0.32 | <0 | <1.7 |
| 6 | pEC50 5.2 | −2.45 | −1.30 | 0.42 | 7.7 | 2.2 |
| 7 | pEC50 4.5 | −0.19 | −0.95 | 1.03 | 4.7 | 2.6 |
| 9 | pIC50 5.1 | 3.77 | −2.85 | 0.44 | 1.3 | 2.2 |
| 10 | pIC50 4.6 | 3.01 | −3.65 | 0.40 | 1.6 | 2.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meijer, I.; Willems, S.; Ni, X.; Heering, J.; Chaikuad, A.; Merk, D. Chemical Starting Matter for HNF4α Ligand Discovery and Chemogenomics. Int. J. Mol. Sci. 2020, 21, 7895. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217895
Meijer I, Willems S, Ni X, Heering J, Chaikuad A, Merk D. Chemical Starting Matter for HNF4α Ligand Discovery and Chemogenomics. International Journal of Molecular Sciences. 2020; 21(21):7895. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217895
Chicago/Turabian StyleMeijer, Isabelle, Sabine Willems, Xiaomin Ni, Jan Heering, Apirat Chaikuad, and Daniel Merk. 2020. "Chemical Starting Matter for HNF4α Ligand Discovery and Chemogenomics" International Journal of Molecular Sciences 21, no. 21: 7895. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217895