The Grapevine Calmodulin-Like Protein Gene CML21 Is Regulated by Alternative Splicing and Involved in Abiotic Stress Response



Abstract
:1. Introduction
2. Results
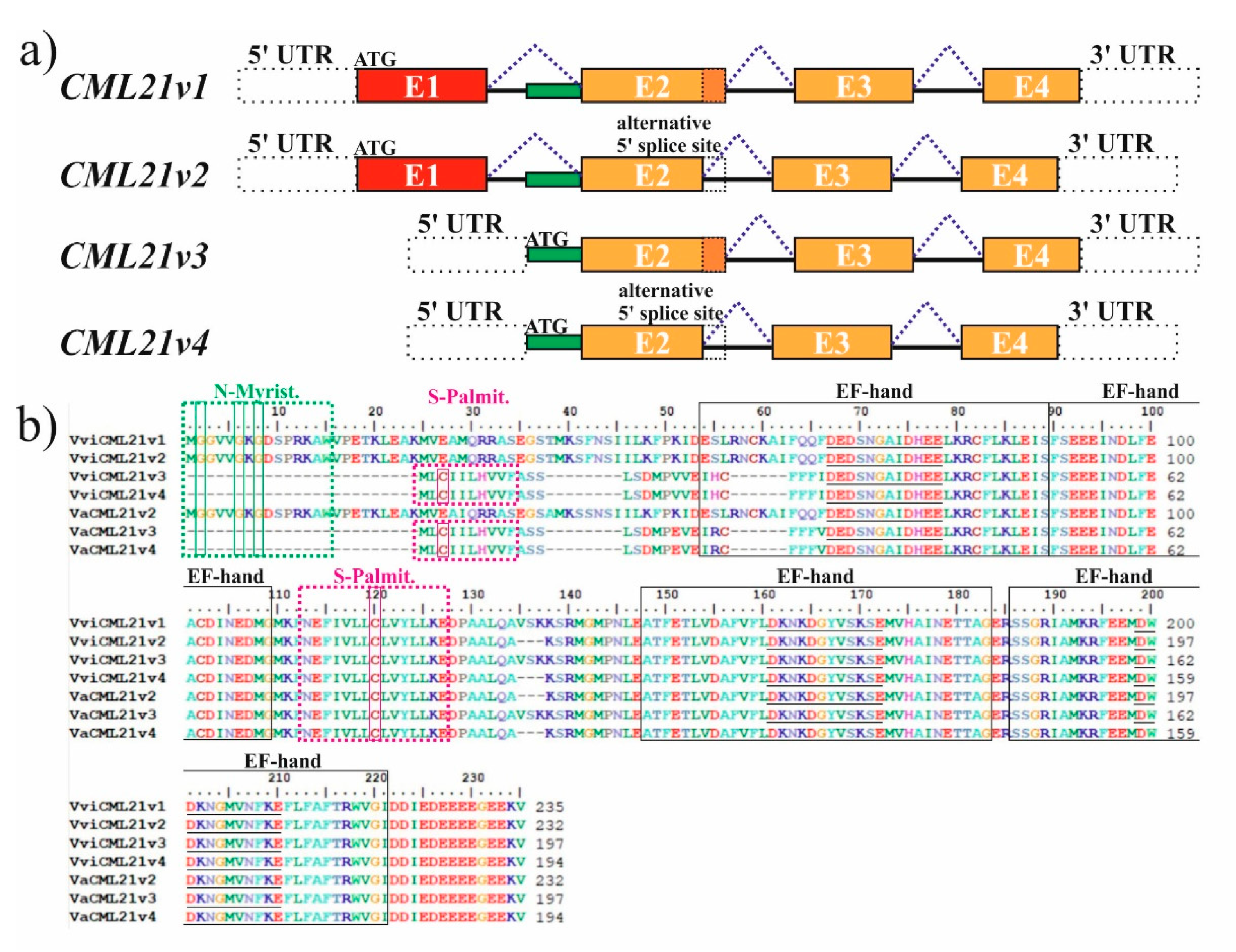
2.1. VaCML21 Is Regulated by Alternative Splicing
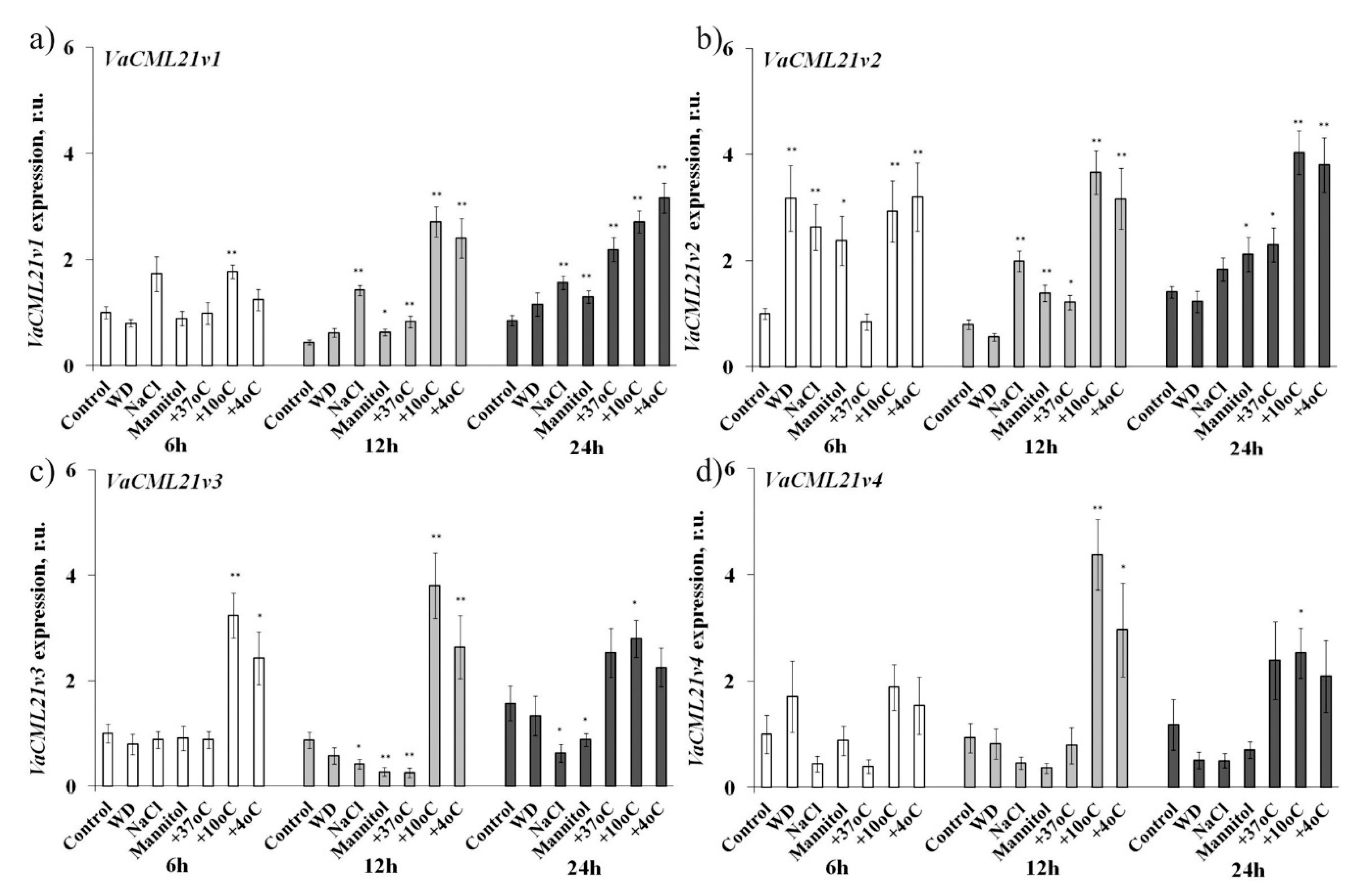
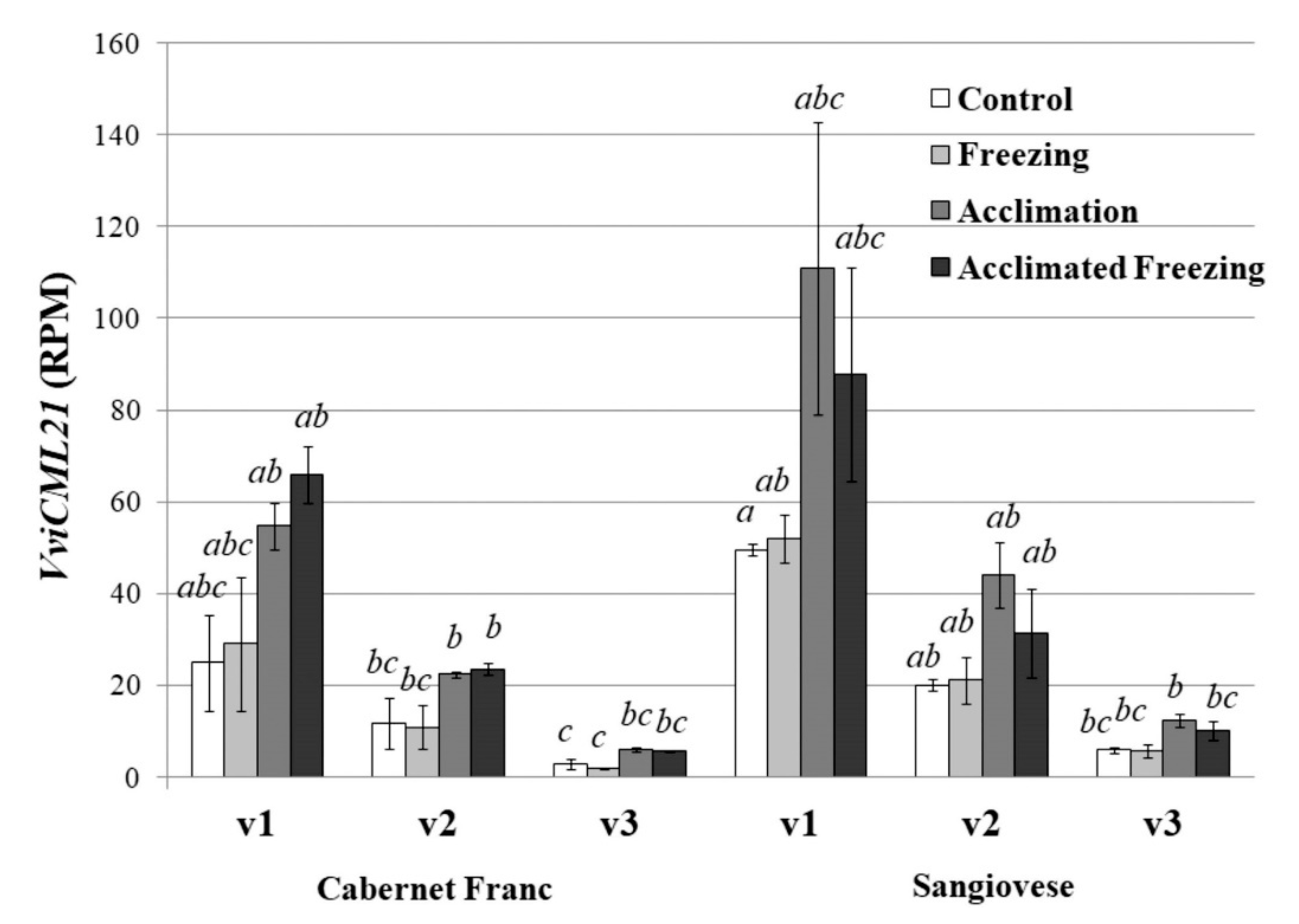
2.2. CML21 Expression Profile under Different Abiotic Stresses
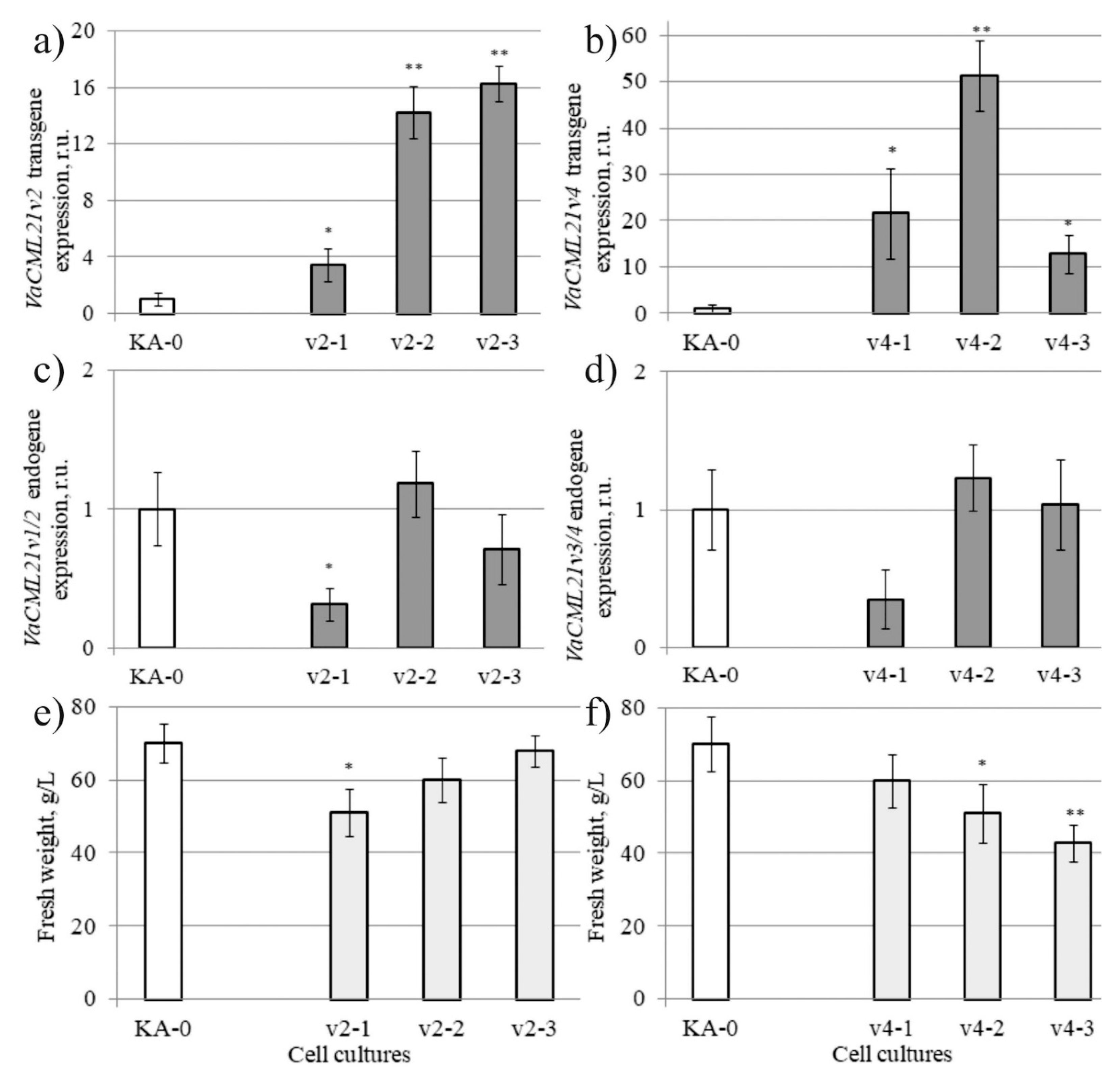
2.3. Genetic Plant Transformation with the VaCML21v2 and VaCML21v4 Splice Variants and Selection of the Transformants
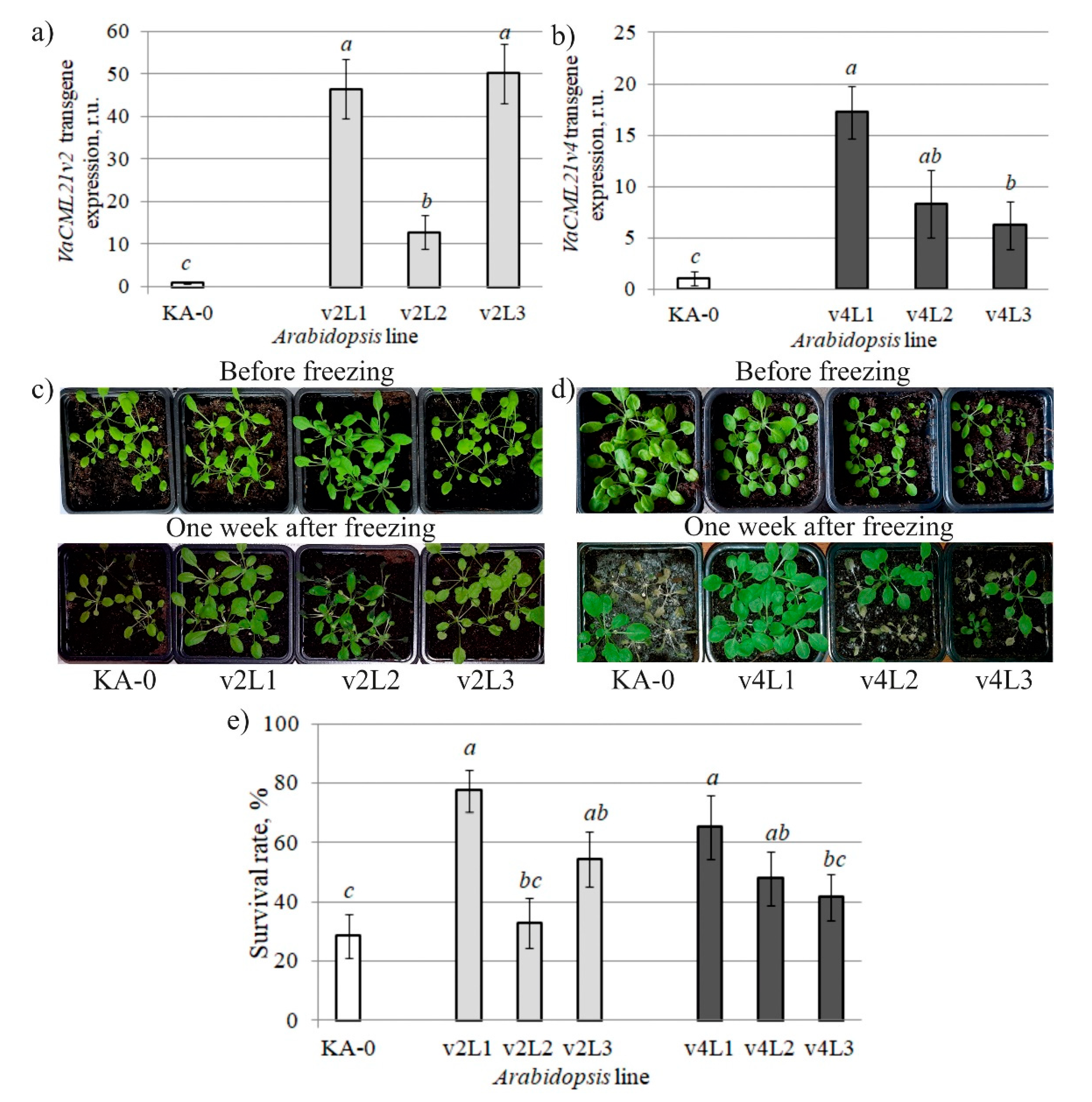
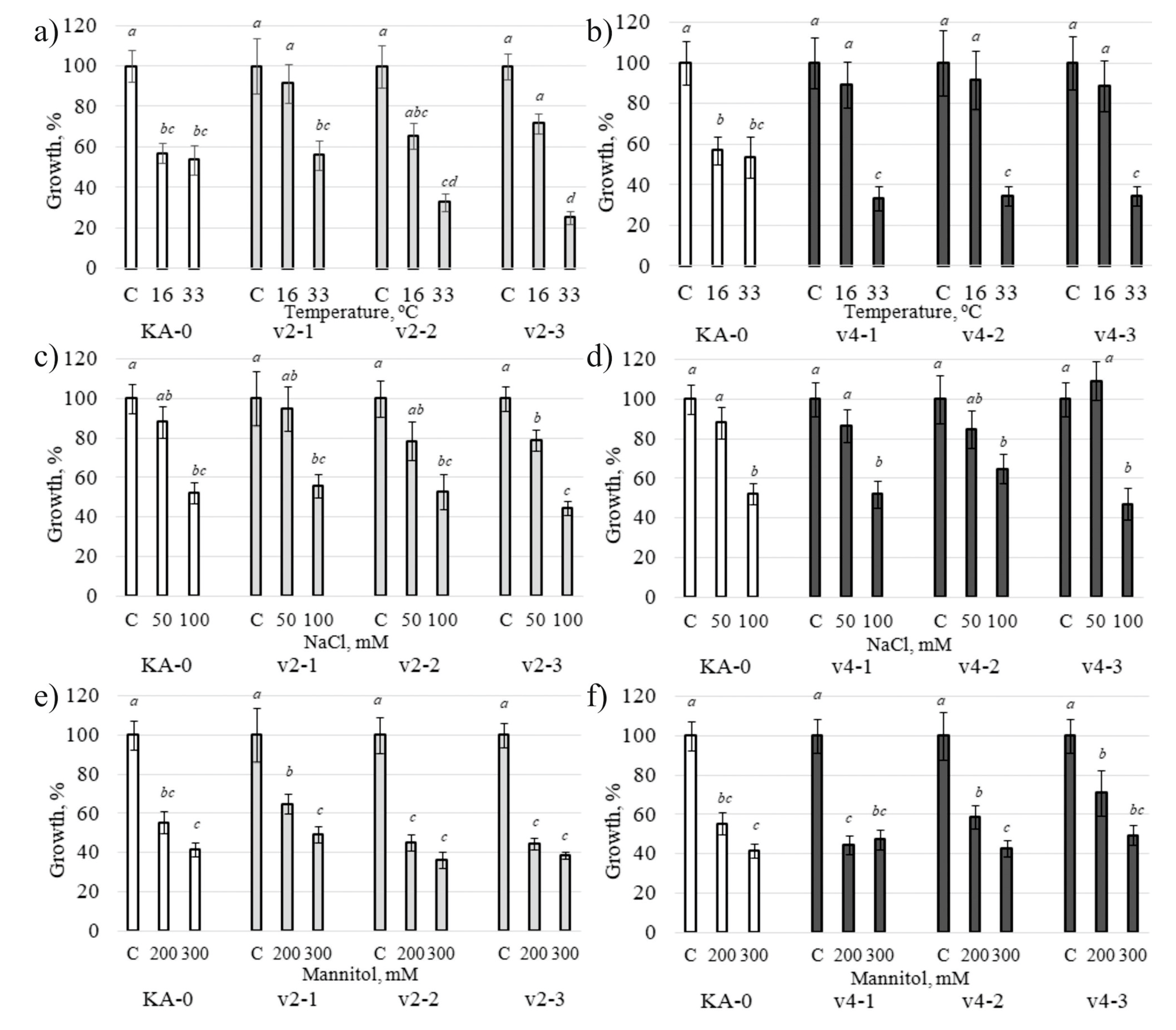
2.4. Stress Tolerance of VaCML21-Overexpressing V. amurensis Cell Cultures and A. thaliana Plants
2.5. Transformation with VaCML21 Enhanced Transcription of Cold-Inducible Regulatory Genes in Stressed Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Isolation and Sequencing of VaCML21 Spliced Transcript Variants
4.3. Generation of Transgenic Grapevine Cell Cultures
4.4. Generation of Transgenic Arabidopsis Plants
4.5. RNA Isolation and cDNA Synthesis
4.6. Gene Expression Analysis by qRT-PCR
4.7. V. vinifera RNAseq Library Analsis
4.8. Abiotic Stress Treatments of Transgenic Cell Cultures and Plants
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signaling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta 2012, 1820, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Yadav, D.; Khan, A.L.; Hashem, A.; Abd Allah, E.F.; Al-Harrasi, A. Molecular players of EF-hand containing calcium signaling event in plants. Int. J. Mol. Sci. 2019, 20, 1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perochon, A.; Aldon, D.; Galaud, J.P.; Ranty, B. Calmodulin and calmodulin-like proteins in plant calcium signaling. Biochimie 2011, 93, 2048–2053. [Google Scholar] [CrossRef]
- La Verde, V.; Dominici, P.; Astegno, A. Towards understanding plant calcium signaling through calmodulin-like proteins: A biochemical and structural perspective. Int. J. Mol. Sci. 2018, 19, 1331. [Google Scholar] [CrossRef] [Green Version]
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the crossroads of signaling. Plant Cell 2002, 14, S401–S417. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Kudla, J. Calcium decoding mechanisms in plants. Biochimie 2011, 93, 2054–2059. [Google Scholar] [CrossRef]
- Zhu, X.; Dunand, C.; Snedden, W.; Galaud, J.P. CaM and CML emergence in the green lineage. Trends Plant Sci. 2015, 20, 483–489. [Google Scholar] [CrossRef]
- Zeng, H.; Xu, L.; Singh, A.; Wang, H.; Du, L.; Poovaiah, B.W. Involvement of calmodulin and calmodulin-like proteins in plant responses to abiotic stresses. Front. Plant Sci. 2015, 6, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheval, C.; Aldon, D.; Galaud, J.P.; Ranty, B. Calcium/calmodulin-mediated regulation of plant immunity. Biochim. Biophys. Acta 2013, 1833, 1766–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midhat, U.; Ting, M.K.Y.; Teresinski, H.J.; Snedden, W.A. The calmodulin-like protein, CML39, is involved in regulating seed development, germination, and fruit development in Arabidopsis. Plant Mol. Biol. 2018, 96, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Popescu, S.C.; Popescu, G.V.; Bachan, S.; Zhang, Z.; Seay, M.; Gerstein, M.; Snyder, M.; Dinesh-Kumar, S.P. Differential binding of calmodulin-related proteins to their targets revealed through high-density Arabidopsis protein microarrays. Proc. Natl. Acad. Sci. USA 2007, 104, 4730–4735. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Aharon, G.S.; Sottosanto, J.B.; Blumwald, E. Vacuolar Na+/H+ antiporter cation selectivity is regulated by calmodulin from within the vacuole in a Ca2+-and pH-dependent manner. Proc. Natl. Acad. Sci. USA 2005, 102, 16107–16112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobney, S.; Chiasson, D.; Lam, P.; Smith, S.P.; Snedden, W.A. The calmodulin-related calcium sensor CML42 plays a role in trichome branching. J. Biol. Chem. 2009, 284, 31647–31657. [Google Scholar] [CrossRef] [Green Version]
- Munir, S.; Liu, H.; Xing, Y.; Hussain, S.; Ouyang, B.; Zhang, Y.; Li, H.; Ye, Z. Overexpression of calmodulin-like (ShCML44) stress-responsive gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses. Sci. Rep. 2016, 6, 31772. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xiong, X.; Arif, S.; Gao, L.; Zhao, L.; Shah, I.H.; Zhang, Y. A calmodulin-like CmCML13 from Cucumis melo improved transgenic Arabidopsis salt tolerance through reduced shoot’s Na(+), and also improved drought resistance. Plant Physiol. Biochem. 2020, 155, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Qiao, Z.; Liu, H.; Acharya, B.R.; Li, C.; Zhang, W. CML20, an Arabidopsis calmodulin-like protein, negatively regulates guard cell ABA signaling and drought stress tolerance. Front. Plant Sci. 2017, 8, 824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.X.; Wang, T.Z.; Liu, M.; Sun, W.; Zhang, W.H. Calmodulin-like gene MtCML40 is involved in salt tolerance by regulating MtHKTs transporters in Medicago truncatula. Environ. Exp. Bot. 2019, 157, 79–90. [Google Scholar] [CrossRef]
- Vandelle, E.; Vannozzi, A.; Wong, D.; Danzi, D.; Digby, A.M.; Dal Santo, S.; Astegno, A. Identification, characterization, and expression analysis of calmodulin and calmodulin-like genes in grapevine (Vitis vinifera) reveal likely roles in stress responses. Plant Physiol. Biochem. 2018, 129, 221–237. [Google Scholar] [CrossRef]
- Li, C.; Meng, D.; Zhang, J.; Cheng, L. Genome-wide identification and expression analysis of calmodulin and calmodulin-like genes in apple (Malus × domestica). Plant Physiol. Biochem. 2019, 139, 600–612. [Google Scholar] [CrossRef]
- Ma, Q.P.; Zhou, Q.Q.; Chen, C.M.; Cui, Q.Y.; Zhao, Y.X.; Wang, K.; Arkorful, E.; Chen, X.; Sun, K.; Li, X.H. Isolation and expression analysis of CsCML genes in response to abiotic stresses in the tea plant (Camellia sinensis). Sci. Rep. 2019, 9, 8211. [Google Scholar] [CrossRef] [PubMed]
- Dubrovina, A.S.; Aleynova, O.A.; Ogneva, Z.V.; Suprun, A.R.; Ananev, A.A.; Kiselev, K.V. The effect of abiotic stress conditions on expression of calmodulin (CaM) and calmodulin-like (CML) genes in wild-growing grapevine Vitis amurensis. Plants 2019, 8, 602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasani, E.; DalCorso, G.; Costa, A.; Zenoni, S.; Furini, A. The Arabidopsis thaliana transcription factor MYB59 regulates calcium signalling during plant growth and stress response. Plant Mol. Biol. 2019, 99, 517–534. [Google Scholar] [CrossRef]
- La Verde, V.; Trande, M.; D’Onofrio, M.; Dominici, P.; Astegno, A. Binding of calcium and target peptide to calmodulin-like protein CML19, the centrin 2 of Arabidopsis thaliana. Int. J. Biol. Macromol. 2018, 108, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.D.; Cao, Y.; Ma, L.G. Alternative splicing in plant genes: A means of regulating the environmental fitness of plants. Int. J. Mol. Sci. 2017, 18, 432. [Google Scholar] [CrossRef]
- Nishiyama, R.; Mizuno, H.; Okada, S.; Yamaguchi, T.; Takenaka, M.; Fukuzawa, H.; Ohyama, K. Two mRNA species encoding calcium-dependent protein kinases are differentially expressed in sexual organs of Marchantia polymorpha through alternative splicing. Plant Cell Physiol. 1999, 40, 205–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubrovina, A.S.; Aleynova, O.A.; Kiselev, K.V.; Novikova, G.V. True and false alternative transcripts of calcium-dependent protein kinase CPK9 and CPK3a genes in Vitis amurensis. Acta Physiol Plant. 2014, 36, 1727–1737. [Google Scholar] [CrossRef]
- Almadanim, M.C.; Gonçalves, N.M.; Rosa, M.T.G.; Alexandre, B.M.; Cordeiro, A.M.; Rodrigues, M.; Saibo, N.J.M.; Soares, C.M.; Romão, C.V.; Oliveira, M.M.; et al. The rice cold-responsive calcium-dependent protein kinase OsCPK17 is regulated by alternative splicing and post-translational modifications. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, S.K.; Kanwar, P.; Samtani, H.; Kaur, K.; Jha, S.K.; Pandey, G.K. Alternative splicing of CIPK3 results in distinct target selection to propagate ABA signaling in Arabidopsis. Front Plant Sci. 2017, 8, 1924. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Li, Q.H.; Yu, Y.N.; Qiao, Y.M.; Haq, S.U.; Gong, Z.H. The CBL-CIPK pathway in plant response to stress signals. Int. J. Mol. Sci. 2020, 21, 5668. [Google Scholar] [CrossRef]
- Falquet, L.; Pagni, M.; Bucher, P.; Hulo, N.; Sigrist, C.J.; Hofmann, K.; Bairoch, A. The PROSITE database, its status in 2002. Nucleic Acids Res. 2002, 30, 235–238. [Google Scholar] [CrossRef]
- Database of Protein Domains, Families and Functional Sites. Available online: http://prosite.expasy.org/ (accessed on 18 September 2020).
- Xie, Y.; Zheng, Y.; Li, H.; Luo, X.; He, Z.; Cao, S.; Shi, Y.; Zhao, Q.; Xue, Y.; Zuo, Z.; et al. GPS-Lipid: A robust tool for the prediction of multiple lipid modification sites. Sci. Rep. 2016, 16, 28249. [Google Scholar] [CrossRef] [PubMed]
- Prediction of Lipid Modification Sites. Available online: http://lipid.biocuckoo.org/webserver.php (accessed on 18 September 2020).
- Alzohairy, A.M. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Compute pI/Mw Tool. Available online: http://web.expasy.org/compute_pi/ (accessed on 18 September 2020).
- Londo, J.P.; Kovaleski, A.P.; Lillis, J.A. Divergence in the transcriptional landscape between low temperature and freeze shock in cultivated grapevine (Vitis vinifera). Hortic Res. 2018, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, K.; Takeda, M.; Kidokoro, S.; Yamada, K.; Sakuma, Y.; Urano, K.; Fujita, M.; Yoshiwara, K.; Matsukura, S.; Morishita, Y.; et al. Metabolic pathways involved in cold acclimation identified by integrated analysis of metabolites and transcripts regulated by DREB1A and DREB2A. Plant Physiol. 2009, 150, 1972–1980. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Furumoto, T. Cold Signaling and Cold Response in Plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, X.; Zhang, L. Structural and functional dynamics of dehydrins: A plant protector protein under abiotic stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef] [Green Version]
- Kurkela, S.; Franck, M. Cloning and characterization of a cold-and ABA-inducible Arabidopsis gene. Plant Mol. Biol. 1990, 15, 137–144. [Google Scholar] [CrossRef]
- Chung, E.; Park, J.M.; Oh, S.K.; Joung, Y.H.; Lee, S.; Choi, D. Molecular and biochemical characterization of the Capsicum annuum calcium-dependent protein kinase 3 (CaCDPK3) gene induced by abiotic and biotic stresses. Planta 2004, 220, 286–295. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S. Expression of calcium-dependent protein kinase (CDPK) genes under abiotic stress conditions in wild-growing grapevine Vitis amurensis. J. Plant Physiol. 2013, 170, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Aleynova-Shumakova, O.A.; Dubrovina, A.S.; Manyakhin, A.Y.; Karetin, Y.A.; Kiselev, K.V. VaCPK20 gene overexpression significantly increased resveratrol content and expression of stilbene synthase genes in cell cultures of Vitis amurensis Rupr. Appl. Microbiol. Biotechnol. 2014, 98, 5541–5549. [Google Scholar] [CrossRef] [PubMed]
- Tzfira, T.; Tian, G.W.; Lacroix, B.; Vyas, S.; Li, J.; Leitner-Dagan, Y.; Krichevsky, A.; Taylor, T.; Vainstein, A.; Citovsky, V. pSAT vectors: A modular series of plasmids for autofluorescent protein tagging and expression of multiple genes in plants. Plant Mol. Biol. 2005, 57, 503–516. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Dubrovina, A.S.; Shumakova, O.A.; Karetin, Y.A.; Manyakhin, A.Y. Structure and expression profiling of a novel calcium-dependent protein kinase gene, CDPK3a, in leaves, stems, grapes, and cell cultures of wild-growing grapevine Vitis amurensis Rupr. Plant Cell Rep. 2013, 32, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Tyunin, A.P.; Suprun, A.R.; Nityagovsky, N.N.; Manyakhin, A.Y.; Karetin, Y.A.; Dubrovina, A.S.; Kiselev, K.V. The effect of explant origin and collection season on stilbene biosynthesis in cell cultures of Vitis amurensis Rupr. Plant Cell Tiss. Organ Cult. 2019, 136, 189–196. [Google Scholar] [CrossRef]
- Zhang, X.R.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. VaCPK20, a calcium-dependent protein kinase gene of wild grapevine Vitis amurensis Rupr, mediates cold and drought stress tolerance. J. Plant Physiol. 2015, 185, 1–12. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Shumakova, O.A.; Manyakhin, A.Y.; Mazeika, A.N. Influence of calcium influx induced by the calcium ionophore, A23187, on resveratrol content and the expression of CDPK and STS genes in the cell cultures of Vitis amurensis. Plant Growth Regul. 2012, 68, 371–381. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. The calcium-dependent protein kinase gene VaCPK29 is involved in grapevine responses to heat and osmotic stresses. Plant Growth Regul. 2017, 82, 79–89. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Aleynova, O.A.; Manyakhin, A.Y.; Kiselev, K.V. The role of calcium-dependent protein kinase genes CPK16, CPK25, CPK30, and CPK32 in stilbene biosynthesis and the stress resistance of grapevine Vitis amurensis Rupr. Appl. Biochem. Microbiol. 2018, 54, 410–417. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.V.; Dubrovina, A.S.; Bulgakov, V.P. Phenylalanine ammonia-lyase and stilbene synthase gene expression in rolB transgenic cell cultures of Vitis amurensis. Appl. Microbiol. Biotechnol. 2009, 82, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Dubrovina, A.S.; Kiselev, K.V.; Aleynova, O.A. Influence of overexpression of the true and false alternative transcripts of calcium-dependent protein kinase CPK9 and CPK3a genes on the growth, stress tolerance, and resveratrol content in Vitis amurensis cell cultures. Acta Physiol. Plant. 2016, 38, 78. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vitis amurensis (cDNA Clone) | No. of EF Hands | Predicted Lipid Modification Sites (Position) | MW (kDA) | Vitis vinifera (mRNA Prediction) | No. of EF Hands | Predicted Lipid Modification Sites (Position) | MW (kDA) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Transcript Variant | ID (NCBI GenBank) | CDS (Amino Acids) | Transcript Variant | ID | CDS (Amino Acids) | ||||||
| nd | nd | nd | nd | nd | nd | VviCML21v.1 | VIT_219s0015g01200.1 | 708 (235) | 4 | N-Myrist(2,6,8) | 26.9 |
| VaCML21v.2 | MN540599 | 699 (232) | 4 | N-Myrist(2,6,8) N-Palmit(120) | 26.4 | VviCML21v.2 | VIT_219s0015g01200.2 | 699 (232) | 4 | N-Myrist(2,6,8) N-Palmit(120) | 26.5 |
| VaCML21v.3 | MW033207 | 594 (197) | 4 | N-Palmit(3) | 22.7 | VviCML21v.3 | VIT_219s0015g01200.7 | 594 (197) | 4 | N-Palmit(3) | 22.7 |
| VaCML21v.4 | MW033208 | 585 (194) | 4 | N-Palmit(3,82) | 22.4 | VviCML21v.4 | VIT_219s0015g01200.8 | 585 (194) | 4 | N-Palmit(3,82) | 22.4 |
| Transcript Name (GenBankID) | Primers, 5′–3′ |
|---|---|
| Cloning and sequencing full-length cDNA coding sequences of VaCML21, 5′–3′ | |
| CML21v1/v2 (no sequence/MN540599) | F1:ATGGGAGGCGTGGTGGG R:TCAAACTTTCTCTTCACCTTC |
| CML21v3/v4 (MW033207/ MW033208) | F2:ATGCTGTGTATCATCCTTCATG R:TCAAACTTTCTCTTCACCTTC |
| Cloning VaCML21v2 and VaCML21v4 for plant transformation by the BglII and SalI sites (underlined), 5’–3’ | |
| VaCML21v2 (MN540599) | F:GCTCAGATCTATGGGAGGCGTGGTGGGAAAA R:TCGAGTCGACTTAAACTTTCTCTTCACCTTC |
| VaCML21v4 (MW033208) | F:GCTCAGATCTATGCTGTGTATCATCCTTCAT R:TCGAGTCGACTCAAACTTTCTCTTCACCTT |
| Primers for real-time PCR, 5′–3′ | |
| VaCML21v1 (no sequence) | GAACTGCAAAGCTATTTTTCAGCA CCCATCCGTGATTTTTTGGATAC |
| VaCML21v2 (MN540599) | GAACTGCAAAGCTATTTTTCAGCA ATCCGTGATTTGGCCTGAAGG |
| VaCML21v3 (MW033207) | TGCCAGAGGTTGAAATTCGTTGTT CCCATCCGTGATTTTTTGGATAC |
| VaCML21v4 (MW033208) | TGCCAGAGGTTGAAATTCGTTGTT ATCCGTGATTTGGCCTGAAGG |
| VaActin1 (DQ517935) | GTATTGTGCTGGATTCTGGTGAT AGCAAGGTCAAGACGAAGGATAG |
| VaGAPDH (XM_002263109) | CACTGAAGATGATGTTGTTTCC GCTATTCCAGCCTTGGCAT |
| AtGAPDH (NM_111283.4) | TTGGTGACAACAGGTCAAGCA AAACTTGTCGCTCAATGCAAT |
| AtEF1a (XM_002864638) | TGAGCACGCTCTTCTTGCTTTCA GGTGGTGGCATCCATCTTGTTACA |
| Primers for cDNA check-up on DNA contamination, 5′–3′ | |
| VaActin1 (DQ517935) | TTGCCATTCAGGCTGTTCTTTCT AGGAGCTGCTCTTTGCAGTTTCC |
| AtActin1 (NM_112764) | GATTCAGATGCCCAGAAGTC TCTGTGAACGATTCCTGGA |
| Primers for qRT-PCR quantification of the VaCML21v2 and VaCML21v4 transgene and endogene mRNAs in transformed plant cells, 5′–3′ | |
| VaCML21v2 transgene (MN540599) | CAAGCATTCTACTTCTATTG ACCATTTTGGCCTCAAGC |
| VaCML21v1/v2 endogene (no sequence/MN540599) | CATACTTAGCACTTGTCCCTTTTCC ACCATTTTGGCCTCAAGC |
| VaCML21v4 transgene (MW033208) | CAAGCATTCTACTTCTATTG TCTTCATAGATGAGGATTCAAATG |
| VaCML21v3/v4 endogene (MW033207/ MW033208) | TGAACCAGTTTTCATTTTAATTCAACGTG TCTTCATAGATGAGGATTCAAATG |
| Sangiovese | Cabernet Franc | Cultivation Conditions a | ||
|---|---|---|---|---|
| SRR6026721 | SRR6026711 | SRR6026737 | SRR6026699 | Control |
| SRR6026722 | SRR6026712 | SRR6026736 | SRR6026700 | Freezing (−3) for 45 min |
| SRR6026716 | SRR6026735 | SRR6026738 | SRR6026698 | Chilling stress (+4) for 48 h |
| SRR6026715 | SRR6026734 | SRR6026739 | SRR6026697 | Acclimated freezing (+4 → −3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleynova, O.A.; Kiselev, K.V.; Ogneva, Z.V.; Dubrovina, A.S. The Grapevine Calmodulin-Like Protein Gene CML21 Is Regulated by Alternative Splicing and Involved in Abiotic Stress Response. Int. J. Mol. Sci. 2020, 21, 7939. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217939
Aleynova OA, Kiselev KV, Ogneva ZV, Dubrovina AS. The Grapevine Calmodulin-Like Protein Gene CML21 Is Regulated by Alternative Splicing and Involved in Abiotic Stress Response. International Journal of Molecular Sciences. 2020; 21(21):7939. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217939
Chicago/Turabian StyleAleynova, Olga A., Konstantin V. Kiselev, Zlata V. Ogneva, and Alexandra S. Dubrovina. 2020. "The Grapevine Calmodulin-Like Protein Gene CML21 Is Regulated by Alternative Splicing and Involved in Abiotic Stress Response" International Journal of Molecular Sciences 21, no. 21: 7939. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217939