Tracing Autism Traits in Large Multiplex Families to Identify Endophenotypes of the Broader Autism Phenotype

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
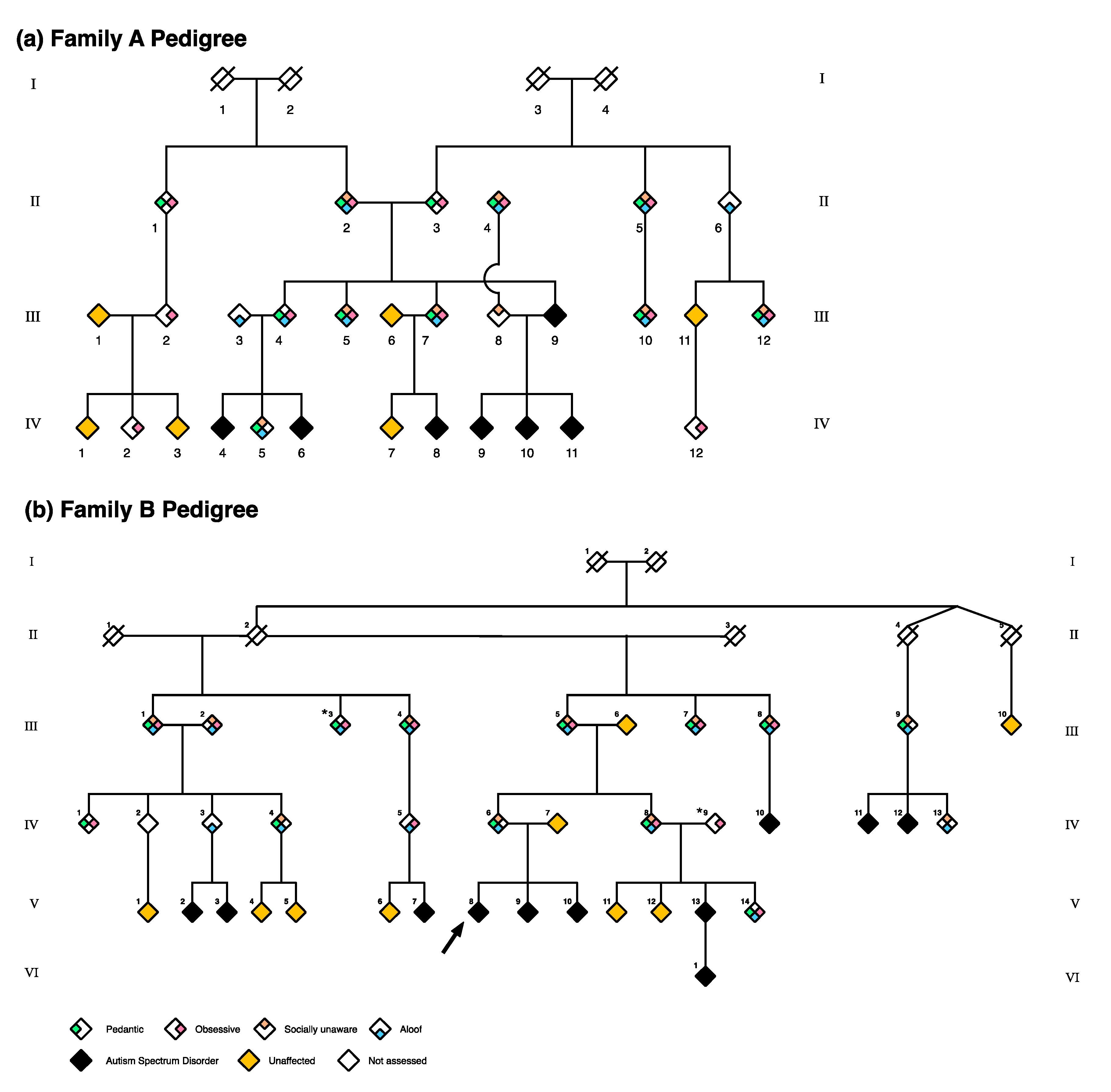
2.1. Hypothesis 1: Multiple Individuals in Large Families Demonstrate the BAP
2.2. Hypothesis 2: Specific BAP Endophenotypes Exist Across BAP Domains
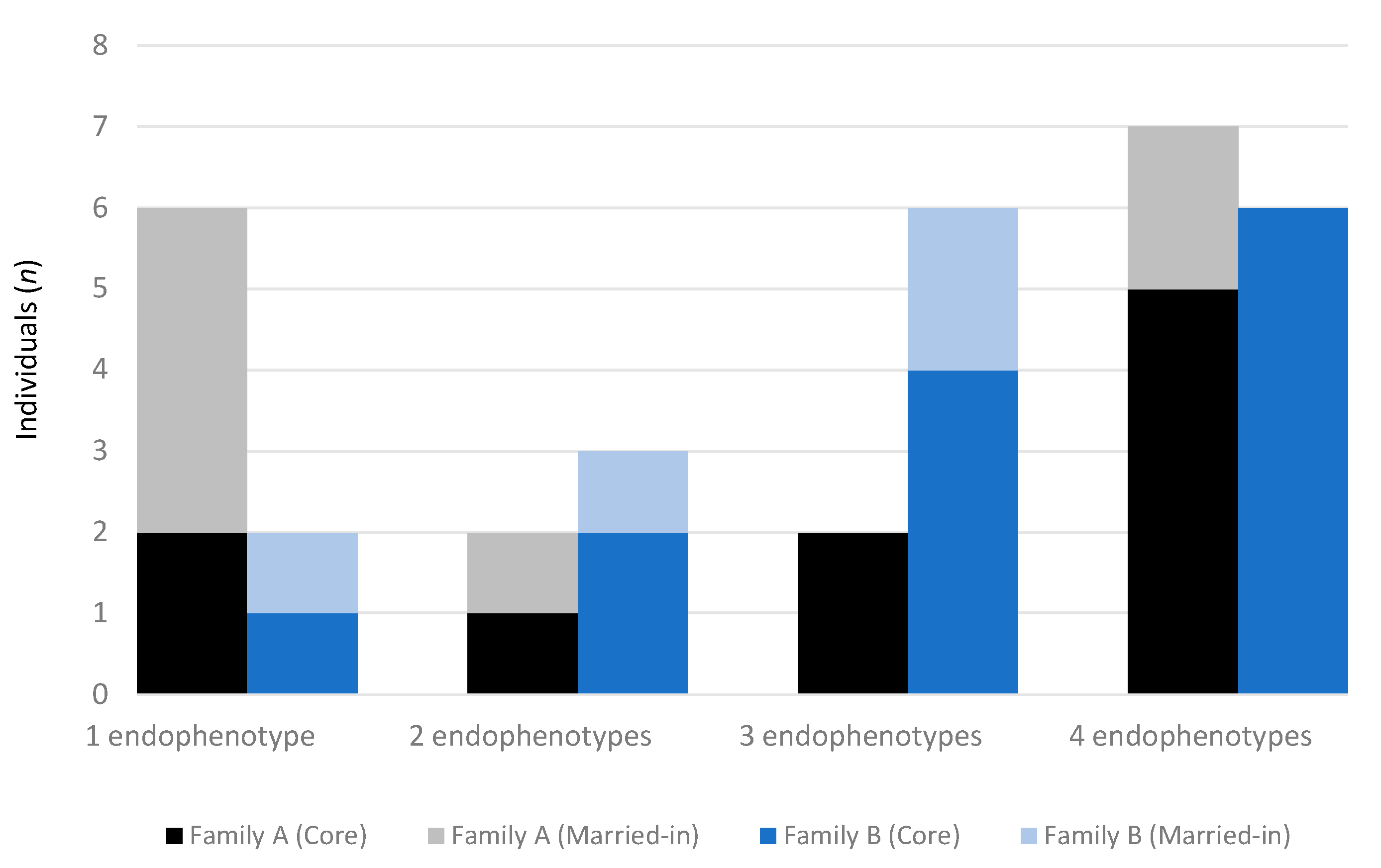
2.3. Hypothesis 3: BAP Endophenotypes Vary in Large Multiplex Families
2.4. Correlates of the BAP Endophenotypes
3. Discussion
4. Materials and Methods
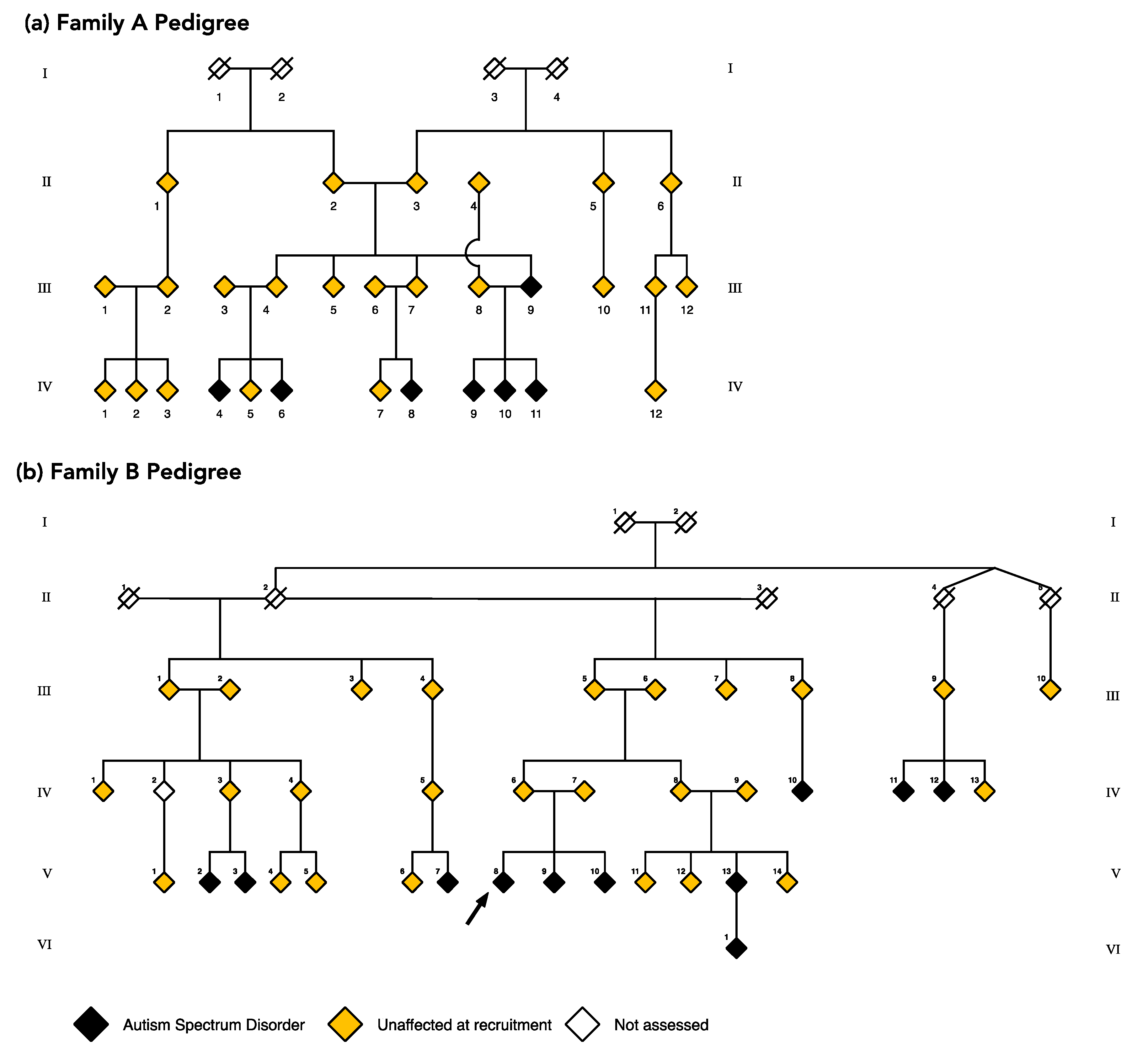
4.1. Large Multiplex Families
4.2. Protocol for Diagnosing ASD in Large Multiplex Families
4.3. Protocol for Phenotyping the BAP in Large Multiplex Families
4.4. Small Families
4.5. Endophenotyping Procedure
4.5.1. Step 1: Identification of Potential BAP Endophenotypes in Large Multiplex Families
4.5.2. Step 2: Validation of BAP Endophenotypes in Small Families
4.5.3. Step 3: Assessment of BAP Endophenotypes in Family A and B
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Maenner, M.J.; Shaw, K.A.; Baio, J. Prevalence of autism spectrum disorder among children aged 8 years—Autism and developmental disabilities monitoring network, 11 sites, United States, 2016. Mmwr Surveill. Summ. 2020, 69, 1. [Google Scholar] [CrossRef] [PubMed]
- Yuen, R.K.; Merico, D.; Bookman, M.; Howe, J.L.; Thiruvahindrapuram, B.; Patel, R.V.; Whitney, J.; Deflaux, N.; Bingham, J.; Wang, Z. Whole genome sequencing resource identifies 18 new candidate genes for autism spectrum disorder. Nat. Neurosci. 2017, 20, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Trost, B.; Engchuan, W.; Nguyen, C.M.; Thiruvahindrapuram, B.; Dolzhenko, E.; Backstrom, I.; Mirceta, M.; Mojarad, B.A.; Yin, Y.; Dov, A. Genome-wide detection of tandem DNA repeats that are expanded in autism. Nature 2020, 586, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Bolton, P.; Macdonald, H.; Pickles, A.; Rios, P.; Goode, S.; Crowson, M.; Bailey, A.; Rutter, M. A case-control family history study of autism. J. Child Psychol. Psychiatry 1994, 35, 877–900. [Google Scholar] [CrossRef] [PubMed]
- Kanner, L. Autistic Disturbances of Affective Contact. Nerv. Child 1943, 2, 34. [Google Scholar]
- Barak, B.; Feng, G. Neurobiology of social behavior abnormalities in autism and Williams syndrome. Nat. Neurosci. 2016, 19, 647–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantino, J.N.; Todd, R.D. Intergenerational transmission of subthreshold autistic traits in the general population. Biol. Psychiatry 2005, 57, 655–660. [Google Scholar] [CrossRef]
- Piven, J.; Palmer, P.; Jacobi, D.; Childress, D.; Arndt, S. Broader autism phenotype: Evidence from a family history study of multiple-incidence autism families. Am. J. Psychiatry 1997, 154, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Landa, R.; Piven, J.; Wzorek, M.M.; Gayle, J.O.; Chase, G.A.; Folstein, S.E. Social language use in parents of autistic individuals. Psychol. Med. 1992, 22, 245–254. [Google Scholar] [CrossRef]
- Piven, J.; Wzorek, M.; Landa, R.; Lainhart, J.; Bolton, P.; Chase, G.A.; Folstein, S. Personality characteristics of the parents of autistic individuals. Psychol. Med. 1994, 24, 783–795. [Google Scholar] [CrossRef]
- Eyuboglu, M.; Baykara, B.; Eyuboglu, D. Broad autism phenotype: Theory of mind and empathy skills in unaffected siblings of children with autism spectrum disorder. Psychiatry Clin. Psychopharmacol. 2018, 28, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Folstein, S.E.; Santangelo, S.L.; Gilman, S.E.; Piven, J.; Landa, R.; Lainhart, J.; Hein, J.; Wzorek, M. Predictors of cognitive test patterns in autism families. J. Child Psychol. Psychiatry 1999, 40, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.; Russell, J.; Robbins, T.W. Evidence for executive dysfunction in autism. Neuropsychologia 1994, 32, 477–492. [Google Scholar] [CrossRef]
- Hughes, C.; Plumet, M.H.; Leboyer, M. Towards a cognitive phenotype for autism: Increased prevalence of executive dysfunction and superior spatial span amongst siblings of children with autism. J. Child Psychol. Psychiatry 1999, 40, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Ronald, A.; Hoekstra, R.A. Autism spectrum disorders and autistic traits: A decade of new twin studies. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2011, 156, 255–274. [Google Scholar] [CrossRef] [PubMed]
- Constantino, J.N. Recurrence rates in autism spectrum disorders. JAMA 2014, 312, 1154–1155. [Google Scholar] [CrossRef] [PubMed]
- Castelbaum, L.; Sylvester, C.M.; Zhang, Y.; Yu, Q.; Constantino, J.N. On the nature of monozygotic twin concordance and discordance for autistic trait severity: A quantitative analysis. Behav. Genet. 2019, 50, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Colvert, E.; Tick, B.; McEwen, F.; Stewart, C.; Curran, S.R.; Woodhouse, E.; Gillan, N.; Hallett, V.; Lietz, S.; Garnett, T. Heritability of autism spectrum disorder in a UK population-based twin sample. JAMA Psychiatry 2015, 72, 415–423. [Google Scholar] [CrossRef]
- Frazier, T.W.; Thompson, L.; Youngstrom, E.A.; Law, P.; Hardan, A.Y.; Eng, C.; Morris, N. A twin study of heritable and shared environmental contributions to autism. J. Autism Dev. Disord. 2014, 44, 2013–2025. [Google Scholar] [CrossRef] [Green Version]
- Codina-Sola, M.; Rodriguez-Santiago, B.; Homs, A.; Santoyo, J.; Rigau, M.; Aznar-Lain, G.; Del Campo, M.; Gener, B.; Gabau, E.; Botella, M.P.; et al. Integrated analysis of whole-exome sequencing and transcriptome profiling in males with autism spectrum disorders. Mol. Autism 2015, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Tammimies, K.; Marshall, C.R.; Walker, S.; Kaur, G.; Thiruvahindrapuram, B.; Lionel, A.C.; Yuen, R.K.; Uddin, M.; Roberts, W.; Weksberg, R.; et al. Molecular Diagnostic Yield of Chromosomal Microarray Analysis and Whole-Exome Sequencing in Children With Autism Spectrum Disorder. JAMA 2015, 314, 895–903. [Google Scholar] [CrossRef]
- Fernandez, B.A.; Scherer, S.W. Syndromic autism spectrum disorders: Moving from a clinically defined to a molecularly defined approach. Dialogues Clin. Neurosci. 2017, 19, 353. [Google Scholar] [PubMed]
- Waye, M.M.; Cheng, H.Y. Genetics and epigenetics of autism: A Review. Psychiatry Clin. Neurosci. 2018, 72, 228–244. [Google Scholar] [CrossRef]
- O’Roak, B.J.; Vives, L.; Girirajan, S.; Karakoc, E.; Krumm, N.; Coe, B.P.; Levy, R.; Ko, A.; Lee, C.; Smith, J.D. Sporadic autism exomes reveal a highly interconnected protein network of de novo mutations. Nature 2012, 485, 246–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, J.M.; Geschwind, D.H. Autism genetics: Searching for specificity and convergence. Genome Biol. 2012, 13, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaugler, T.; Klei, L.; Sanders, S.J.; Bodea, C.A.; Goldberg, A.P.; Lee, A.B.; Mahajan, M.; Manaa, D.; Pawitan, Y.; Reichert, J. Most genetic risk for autism resides with common variation. Nat. Genet. 2014, 46, 881–885. [Google Scholar] [CrossRef] [Green Version]
- Helbig, I.; Scheffer, I.E.; Mulley, J.C.; Berkovic, S.F. Navigating the channels and beyond: Unravelling the genetics of the epilepsies. Lancet Neurol. 2008, 7, 231–245. [Google Scholar] [CrossRef]
- Bora, E.; Aydin, A.; Sarac, T.; Kadak, M.T.; Kose, S. Heterogeneity of subclinical autistic traits among parents of children with autism spectrum disorder: Identifying the broader autism phenotype with a data-driven method. Autism Res. 2017, 10, 321–326. [Google Scholar] [CrossRef]
- Woodbury-Smith, M.; Paterson, A.D.; Thiruvahindrapduram, B.; Lionel, A.C.; Marshall, C.R.; Merico, D.; Fernandez, B.A.; Duku, E.; Sutcliffe, J.S.; O’Conner, I.; et al. Using extended pedigrees to identify novel autism spectrum disorder (ASD) candidate genes. Hum. Genet. 2015, 134, 191–201. [Google Scholar] [CrossRef]
- Dawson, G.; Webb, S.; Schellenberg, G.D.; Dager, S.; Friedman, S.; Aylward, E.; Richards, T. Defining the broader phenotype of autism: Genetic, brain, and behavioral perspectives. Dev. Psychopathol. 2002, 14, 581–611. [Google Scholar] [CrossRef] [Green Version]
- Gould, T.D.; Gottesman, I.I. Psychiatric endophenotypes and the development of valid animal models. Genes Brain Behav. 2006, 5, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Lenzenweger, M.F. Endophenotype, intermediate phenotype, biomarker: Definitions, concept comparisons, clarifications. Depress. Anxiety 2013, 30, 185–189. [Google Scholar] [CrossRef]
- Beauchaine, T.P.; Constantino, J.N. Redefining the endophenotype concept to accommodate transdiagnostic vulnerabilities and etiological complexity. Biomark. Med. 2017, 11, 769–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glahn, D.C.; Knowles, E.E.; McKay, D.R.; Sprooten, E.; Raventós, H.; Blangero, J.; Gottesman, I.I.; Almasy, L. Arguments for the sake of endophenotypes: Examining common misconceptions about the use of endophenotypes in psychiatric genetics. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2014, 165, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duong, L.; Klitten, L.L.; Moller, R.S.; Ingason, A.; Jakobsen, K.D.; Skjodt, C.; Didriksen, M.; Hjalgrim, H.; Werge, T.; Tommerup, N. Mutations in NRXN1 in a family multiply affected with brain disorders: NRXN1 mutations and brain disorders. Am. J. Med. Genet B Neuropsychiatr. Genet. 2012, 159, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, X.; Golhar, R.; Otieno, F.G.; He, M.; Hou, C.; Kim, C.; Keating, B.; Lyon, G.J.; Wang, K.; et al. Whole-genome sequencing in an autism multiplex family. Mol. Autism 2013, 4, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, N.H.; Nato, A.Q., Jr.; Bernier, R.; Ankenman, K.; Sohi, H.; Munson, J.; Patowary, A.; Archer, M.; Blue, E.M.; Webb, S.J.; et al. Whole exome sequencing in extended families with autism spectrum disorder implicates four candidate genes. Hum. Genet. 2015, 134, 1055–1068. [Google Scholar] [CrossRef] [Green Version]
- Salyakina, D.; Cukier, H.N.; Lee, J.M.; Sacharow, S.; Nations, L.D.; Ma, D.; Jaworski, J.M.; Konidari, I.; Whitehead, P.L.; Wright, H.H.; et al. Copy number variants in extended autism spectrum disorder families reveal candidates potentially involved in autism risk. PLoS ONE 2011, 6, e26049. [Google Scholar] [CrossRef]
- Losh, M.; Adolphs, R.; Poe, M.D.; Couture, S.; Penn, D.; Baranek, G.T.; Piven, J. Neuropsychological profile of autism and the broad autism phenotype. Arch. Gen. Psychiatry 2009, 66, 518–526. [Google Scholar] [CrossRef] [Green Version]
- Gottesman, I.I.; Gould, T.D. The endophenotype concept in psychiatry: Etymology and strategic intentions. Am. J. Psychiatry 2003, 160, 636–645. [Google Scholar] [CrossRef]
- Billeci, L.; Calderoni, S.; Conti, E.; Gesi, C.; Carmassi, C.; Dell’Osso, L.; Cioni, G.; Muratori, F.; Guzzetta, A. The Broad Autism (Endo)Phenotype: Neurostructural and Neurofunctional Correlates in Parents of Individuals with Autism Spectrum Disorders. Front. Neurosci. 2016, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pua, E.P.K.; Bowden, S.C.; Seal, M.L. Autism spectrum disorders: Neuroimaging findings from systematic reviews. Res. Autism Spectr. Disord. 2017, 34, 28–33. [Google Scholar] [CrossRef]
- Pua, E.P.K.; Ball, G.; Adamson, C.; Bowden, S.; Seal, M.L. Quantifying individual differences in brain morphometry underlying symptom severity in Autism Spectrum Disorders. Sci. Rep. 2019, 9, 9898. [Google Scholar] [CrossRef] [PubMed]
- Pua, E.P.K.; Malpas, C.B.; Bowden, S.C.; Seal, M.L. Different brain networks underlying intelligence in autism spectrum disorders. Hum. Brain Mapp. 2018, 39, 3253–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pua, E.P.K.; Thomson, P.; Yang, J.Y.-M.; Craig, J.M.; Ball, G.; Seal, M. Individual Differences in Intrinsic Brain Networks Predict Symptom Severity in Autism Spectrum Disorders. Cereb. Cortex 2020. [Google Scholar] [CrossRef] [PubMed]
- Geschwind, D.H.; State, M.W. Gene hunting in autism spectrum disorder: On the path to precision medicine. Lancet Neurol. 2015, 14, 1109–1120. [Google Scholar] [CrossRef] [Green Version]
- Leppa, V.M.; Kravitz, S.N.; Martin, C.L.; Andrieux, J.; Le Caignec, C.; Martin-Coignard, D.; DyBuncio, C.; Sanders, S.J.; Lowe, J.K.; Cantor, R.M.; et al. Rare Inherited and De Novo CNVs Reveal Complex Contributions to ASD Risk in Multiplex Families. Am. J. Hum. Genet. 2016, 99, 540–554. [Google Scholar] [CrossRef] [Green Version]
- Icasiano, F.; Hewson, P.; Machet, P.; Cooper, C.; Marshall, A. Childhood autism spectrum disorder in the Barwon region: A community based study. J. Paediatr. Child Health 2004, 40, 696–701. [Google Scholar] [CrossRef]
- Lord, C.; Risi, S.; Lambrecht, L.; Cook, E.H., Jr.; Leventhal, B.L.; DiLavore, P.C.; Pickles, A.; Rutter, M. The autism diagnostic observation schedule-generic: A standard measure of social and communication deficits associated with the spectrum of autism. J. Autism Dev. Disord. 2000, 30, 205–223. [Google Scholar] [CrossRef]
- Lord, C.; Pickles, A.; McLennan, J.; Rutter, M.; Bregman, J.; Folstein, S.; Fombonne, E.; Leboyer, M.; Minshew, N. Diagnosing autism: Analyses of data from the Autism Diagnostic Interview. J. Autism Dev. Disord. 1997, 27, 501–517. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 4th ed.; Text Revision (DSM-IV-TR); American Psychiatric Association: Washington, DC, USA, 2000. [Google Scholar]
- Stone, V.E.; Baron-Cohen, S.; Knight, R.T. Frontal lobe contributions to theory of mind. J. Cogn. Neurosci. 1998, 10, 640–656. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; O’Riordan, M.; Stone, V.; Jones, R.; Plaisted, K. Recognition of faux pas by normally developing children and children with Asperger syndrome or high-functioning autism. J. Autism Dev. Disord. 1999, 29, 407–418. [Google Scholar] [CrossRef]
- Brown, N.J. Family and Community Study of the Genetics of Autism Spectrum Disorder; University of Melbourne: Melbourne, Australia, 2014. [Google Scholar]
- Goldman Eisler, F. Psycholinguistics: Experiments in Spontaneous Speech; Academic Press Inc: London, UK, 1968. [Google Scholar]
- Joanette, Y.; Goulet, P. Narrative Discourse in Right-Brain-Damaged Right-Handers. In Discourse Ability and Brain Damage; Joanette, Y., Brownell, H.H., Eds.; Springer: New York, NY, USA, 1990. [Google Scholar]
- Wechsler, D. Wechsler Abbreviated Scale of Intelligence; Harcourt Assessment Inc: San Antonio, TX, USA, 1999. [Google Scholar]
- Delis, D.C.; Kaplan, E.; Kramer, J.H. Delis-Kaplan Executive Function System (D-KEFS); Pearson Psychological Corporation: San Antonio, TX, USA, 2018. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P.L.; Oakland, T. Adaptive Behavior Assessment System—Second Edition (ABAS-II), 2nd ed.; The Psychological Corporation: San Antonio, TX, USA, 2003. [Google Scholar]
- Roth, R.M.; Gioia, G.A.; Guy, S.C.; Kenworthy, L.; Isquith, P.K. Behavior Rating Inventory of Executive Function: BRIEF; Psychological Assessment Resources: Odessa, FL, USA, 2000. [Google Scholar]
- Wechsler, D. Wechsler Abbreviated Scale of Intelligence-II; Harcourt Assessment Inc: San Antonio, TX, USA, 2011. [Google Scholar]
- Smith, J.A. Reflecting on the development of interpretative phenomenological analysis and its contribution to qualitative research in psychology. Qual. Res. Psychol. 2004, 1, 39–54. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Luo, J.; Xiong, C. Youden index and Associated Cut-points for Three Ordinal Diagnostic Groups. Commun. Stat. Simul. Comput. 2013, 42, 1213–1234. [Google Scholar] [CrossRef] [PubMed]
- Youden, W.J. Index for rating diagnostic tests. Cancer 1950, 3, 32–35. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participants (n) (Female) | Mean Age (Range) | Cognitive Data (n) | FSIQ Mean (SD) | VIQ Mean (SD) | PIQ Mean (SD) | |
|---|---|---|---|---|---|---|
| Family A | ||||||
| ASD | 7 (1) | 11.43 (4–34) | 5 | 107 (16) | 97 (15) | 101 (30) |
| Unaffected | 6 (5) | 25.83 (2–50) | 5 | 127 (17) | 130 (17) | 117 (15) |
| BAP | 17 (8) | 49.18 (13–79) | 16 a | 119 (13) | 117 (13) | 115 (12) |
| Total | 30 (14) | 35.7 (2–79) | 26 | 118 (15) | 116 (17) | 113 (18) |
| Family B | ||||||
| ASD | 9 (3) | 15.00 (8–20) | 5 | 95 (20) | 89 (28) | 105 (16) |
| Unaffected | 9 (7) | 37.33 (10–73) | 9 | 110 (14) | 111 (14) | 108 (11) |
| BAP | 15 (10) | 47.40 (15–73) | 15 | 102 (17) | 105 (18) | 99 (17) |
| Total | 33 (20) | 35.06 (6–73) | 29 | 103 (17) | 104 (19) | 102 (15) |
| Total | 63 (34) | 35.37 (2–79) | 55 a | 110 (18) | 110 (19) | 107 (17) |
| Mean Proportional Score (SD) | Cut-off Score | BAP Traits | |
|---|---|---|---|
| Unaffected (n = 11) | BAP (n = 30) | ||
| ‘Socially unaware’: Poor self-regulation and reciprocity in conversation | |||
| 0.20 (0.18) | 0.69 (0.06) ** | >0.17 |
|
| ‘Pedantic’: Self-focused and technical in interactions | |||
| 0.04 (0.05) | 0.11 (0.12) * | >0.14 |
|
| ‘Aloof’: Difficulties relating to other’s emotions and expressing own emotions | |||
| 0.12 (0.07) | 0.31 (0.16) *** | >0.20 |
|
| ‘Obsessive’: Regimented approach to life and tendency to ruminate | |||
| 0.13 (0.10) | 0.27 (0.18) *** | >0.25 |
|
| Endophenotypes | ||||||
|---|---|---|---|---|---|---|
| Domain | Task | Socially Unaware | Pedantic | Aloof | Obsessive | Total Number |
| Social communication | PRS | 0.83 ** | 0.73 ** | 0.76 ** | 0.45 ** | 0.86 ** |
| FPT | −0.43 ** | −0.28 | −0.24 | −0.18 | −0.40 ** | |
| Intellect | FSIQ | −0.36 * | −0.10 | −0.31 * | −0.02 | −0.28 |
| VIQ | −0.29 | −0.05 | −0.31 * | −0.03 | −0.28 | |
| PIQ | −0.36 * | −0.13 | −0.19 | −0.02 | −0.26 | |
| Executive functions | Trails (numbers) a | −0.32 * | −0.20 | −0.29 | 0.17 | −0.19 |
| Trails (switch) a | −0.24 | −0.23 | −0.27 | −0.22 | −0.34 * | |
| Design fluency (switch) a | −0.27 | −0.25 | −0.15 | −0.09 | −0.34 * | |
| Design fluency (composite) a | −0.27 | −0.25 | −0.25 | −0.03 | −0.32 * | |
| Tower task (achievement) a | −0.40 ** | −0.19 | −0.27 | −0.22 | −0.26 | |
| Sorting (confirmed) | −0.33 * | −0.23 | −0.32 * | −0.22 | −0.33 * | |
| Sorting (free sort) | −0.31 | −0.15 | −0.24 | −0.37* | −0.37 * | |
| Adaptive function | Social index (self-report) | −0.46 * | −0.35 | −0.19 | −0.31 | −0.43 * |
| Endophenotype | Core Characteristic | Associated Functional Domains | |||
|---|---|---|---|---|---|
| Social | Intellect | Executive | Adaptive | ||
| Socially unaware | Poor self-regulation and reciprocity in conversation | ✓ | ✓ | ✓ | ✓ |
| Pedantic | Self-focused and technical in interactions | ✓ | |||
| Aloof | Difficulties expressing and relating to other’s emotions | ✓ | ✓ | ✓ | |
| Obsessive | Regimented approach to life and tendency to ruminate | ✓ | ✓ | ||
| Participants with ASD | Participants without ASD | ||||
|---|---|---|---|---|---|
| Protocol Item | Child or Adolescent ≥ 4.5–17 yr | Adult ≥ 18 yr | Child < 13 yr | Adolescent ≥ 13–17 yr | Adult ≥ 18 yr |
| ADI-R + ADOS-G or DSM-IV interview + ADOS-G | + | ± | − | − | − |
| Detailed developmental, medical, psychiatric and behavioural history | + | + | + | + | + |
| Family History Interview | − | + | − | − | ± |
| Standardised testing of cognition and executive function a | ± | + | + | + | ± |
| Questionnaires of adaptive behaviour b | + | + | + | + | ± |
| Broader Autism Phenotype Interview, the Faux Pas Task, Cartoon Task and Pragmatic Rating Scale | − | ± | − | + | + |
| Physical Examination | + | + | + | + | + |
| High resolution molecular karyotype, Fragile X testing, metabolic investigations | + | − | − | − | − |
| Unaffected | BAP | |
|---|---|---|
| Number of participants (female) | 11 (6) | 30 (19) |
| Mean age (range) | 41.09 (18–53) | 39.50 (14–53) |
| Mean BAPQ (SD) a | 2.43 (0.38) | 2.92 (0.92) |
| Mean FSIQ (SD) b | 108 (14) | 111 (13) |
| Mean VCI (SD) b | 106 (18) | 109 (15) |
| Mean PRI (SD) b | 109 (7) | 110 (15) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trevis, K.J.; Brown, N.J.; Green, C.C.; Lockhart, P.J.; Desai, T.; Vick, T.; Anderson, V.; Pua, E.P.K.; Bahlo, M.; Delatycki, M.B.; et al. Tracing Autism Traits in Large Multiplex Families to Identify Endophenotypes of the Broader Autism Phenotype. Int. J. Mol. Sci. 2020, 21, 7965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217965
Trevis KJ, Brown NJ, Green CC, Lockhart PJ, Desai T, Vick T, Anderson V, Pua EPK, Bahlo M, Delatycki MB, et al. Tracing Autism Traits in Large Multiplex Families to Identify Endophenotypes of the Broader Autism Phenotype. International Journal of Molecular Sciences. 2020; 21(21):7965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217965
Chicago/Turabian StyleTrevis, Krysta J., Natasha J. Brown, Cherie C. Green, Paul J. Lockhart, Tarishi Desai, Tanya Vick, Vicki Anderson, Emmanuel P. K. Pua, Melanie Bahlo, Martin B. Delatycki, and et al. 2020. "Tracing Autism Traits in Large Multiplex Families to Identify Endophenotypes of the Broader Autism Phenotype" International Journal of Molecular Sciences 21, no. 21: 7965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217965