Distinct Osteogenic Potentials of BMP-2 and FGF-2 in Extramedullary and Medullary Microenvironments

 , and
, and
Abstract
:1. Introduction
2. Results
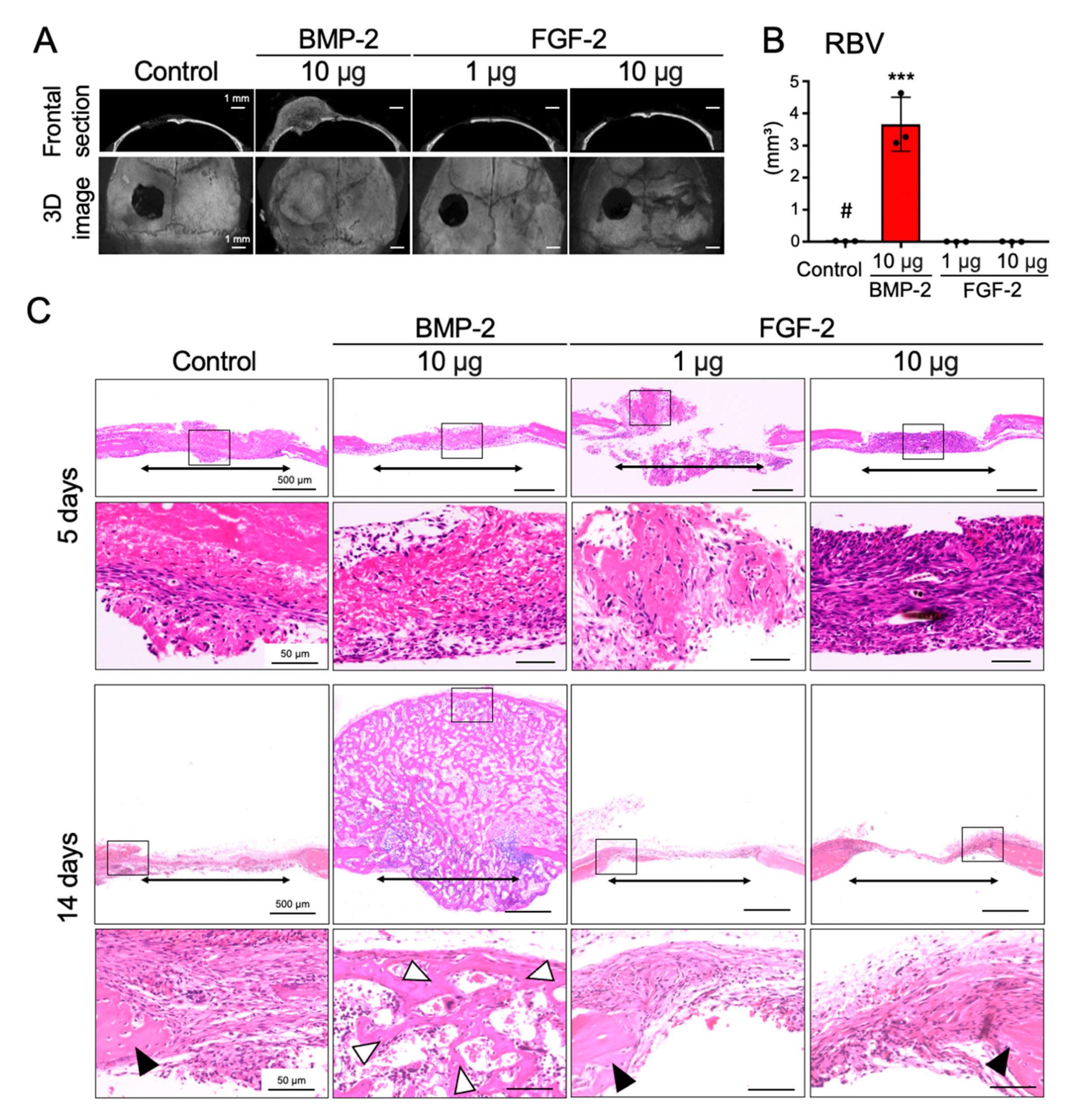
2.1. BMP-2, but Not FGF-2, Promotes the Repair of Mouse Calvarial Defect
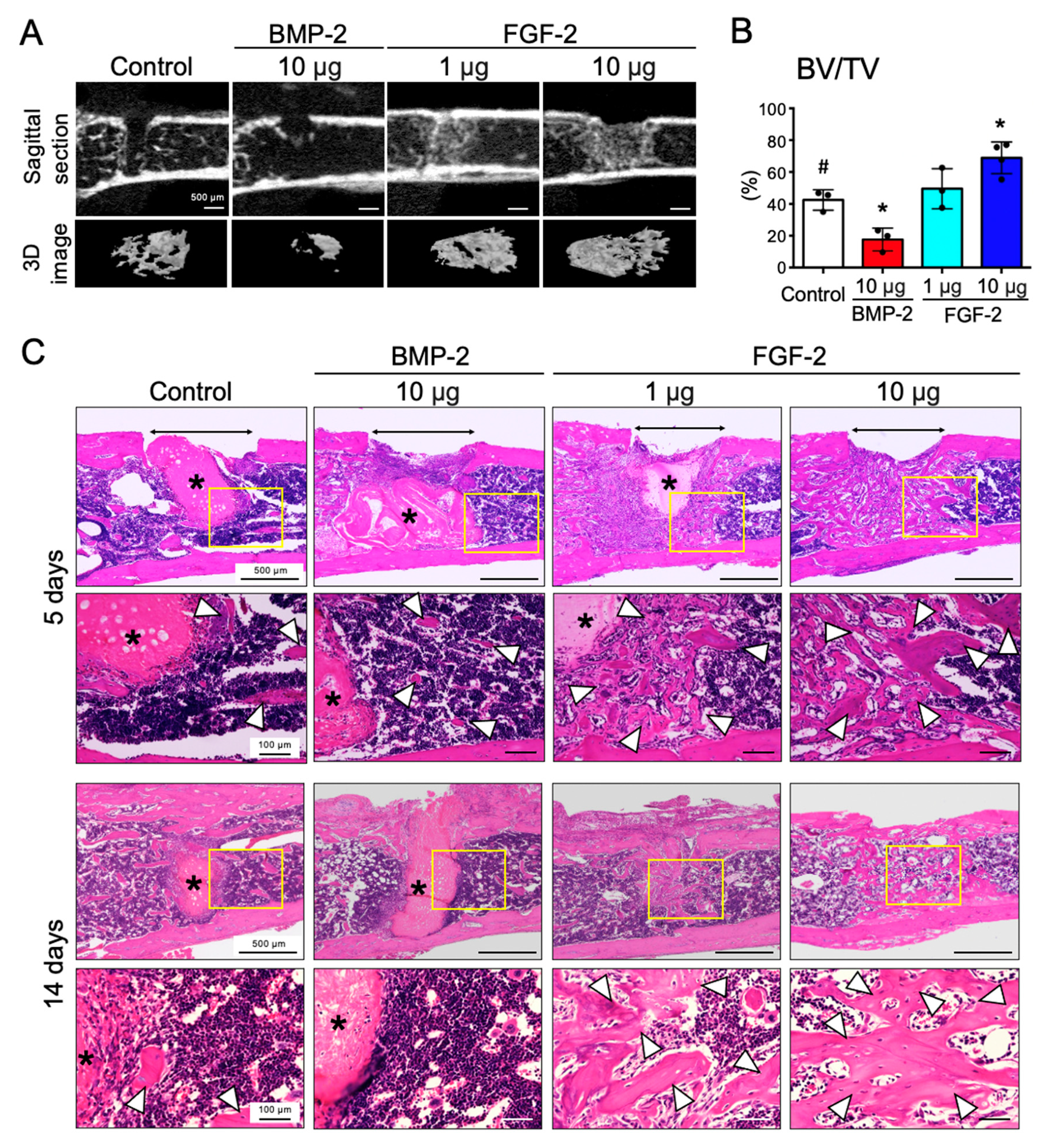
2.2. FGF-2, but Not BMP-2, Promotes the Repair of Mouse Femoral Defect
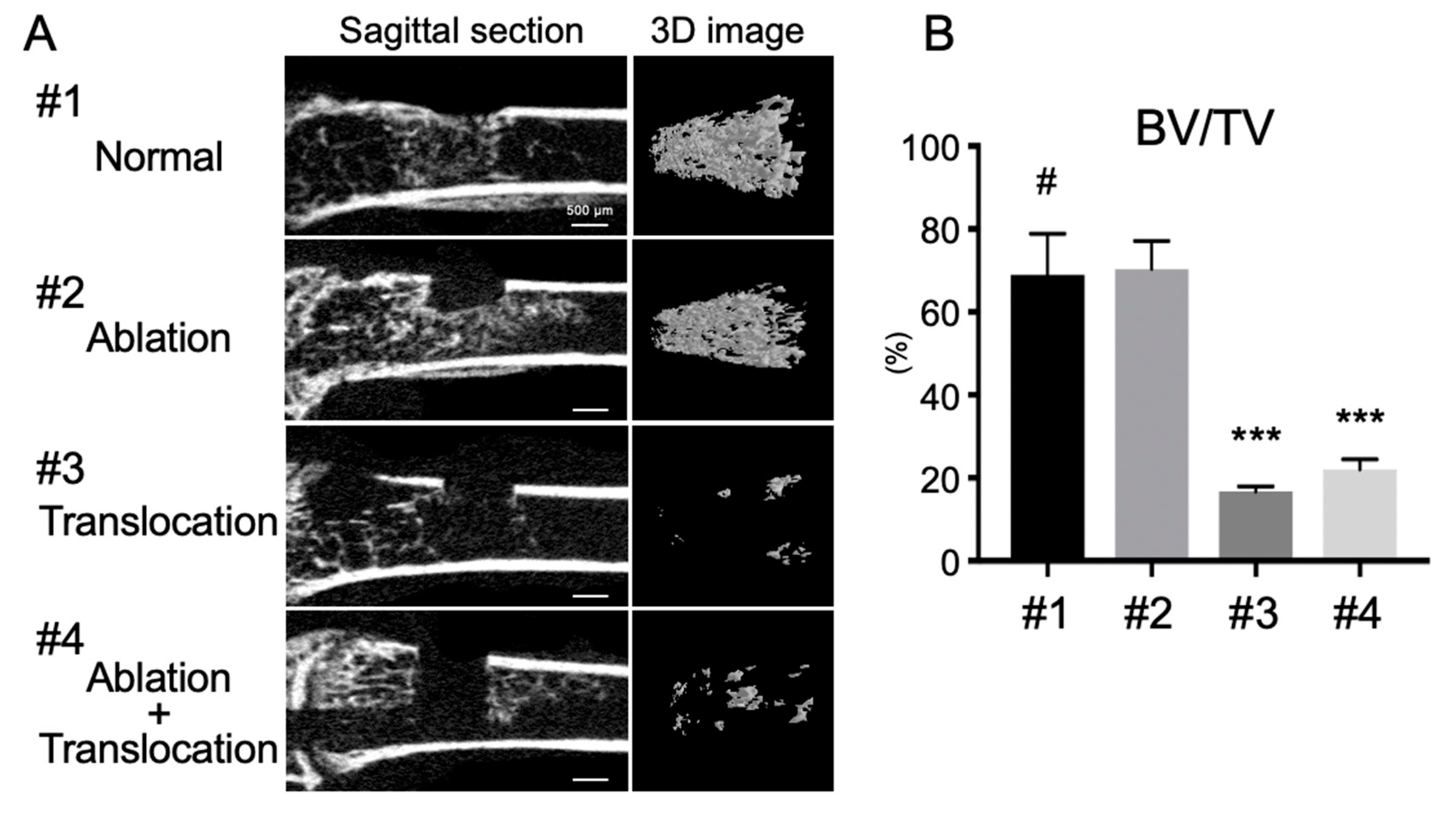
2.3. Depletion of Bone Marrow Cells Inhibits FGF-2-Induced Bone Formation in the Marrow Cavity
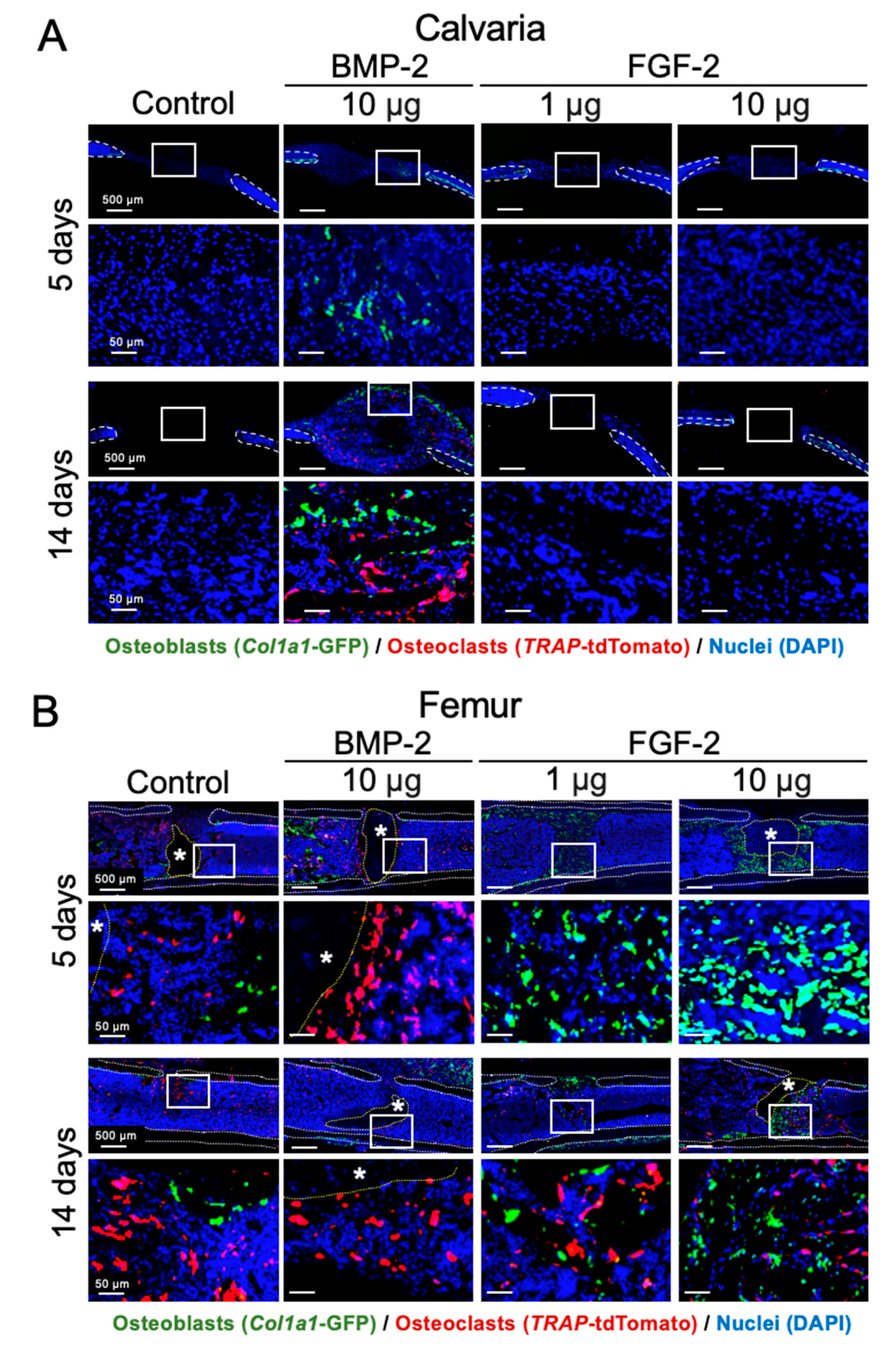
2.4. Effects of BMP-2 and FGF-2 on Osteoblast and Osteoclast in Mouse Calvarial and Femoral Defects
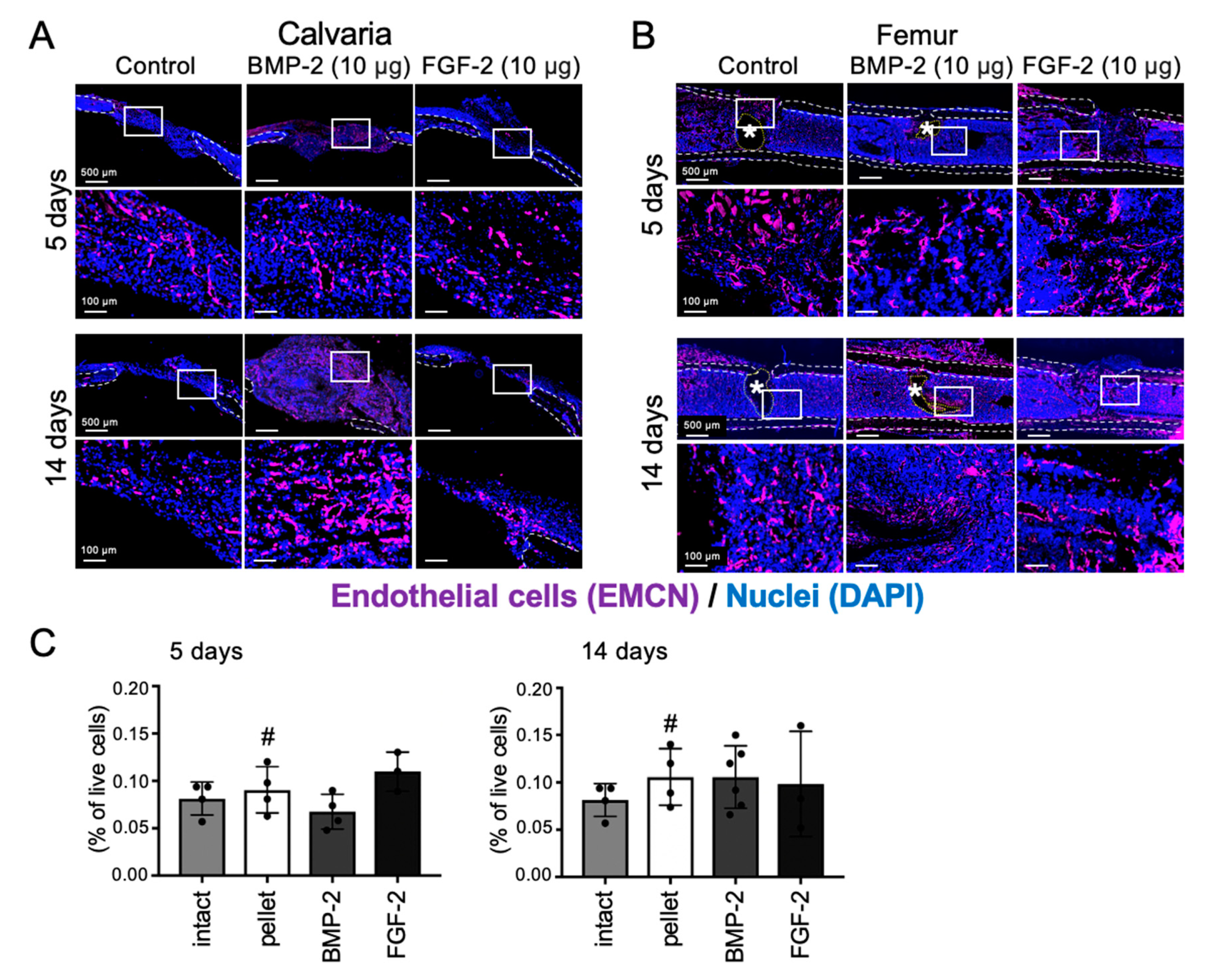
2.5. Effects of BMP-2 and FGF-2 on Angiogenesis in Mouse Calvarial and Femoral Defects
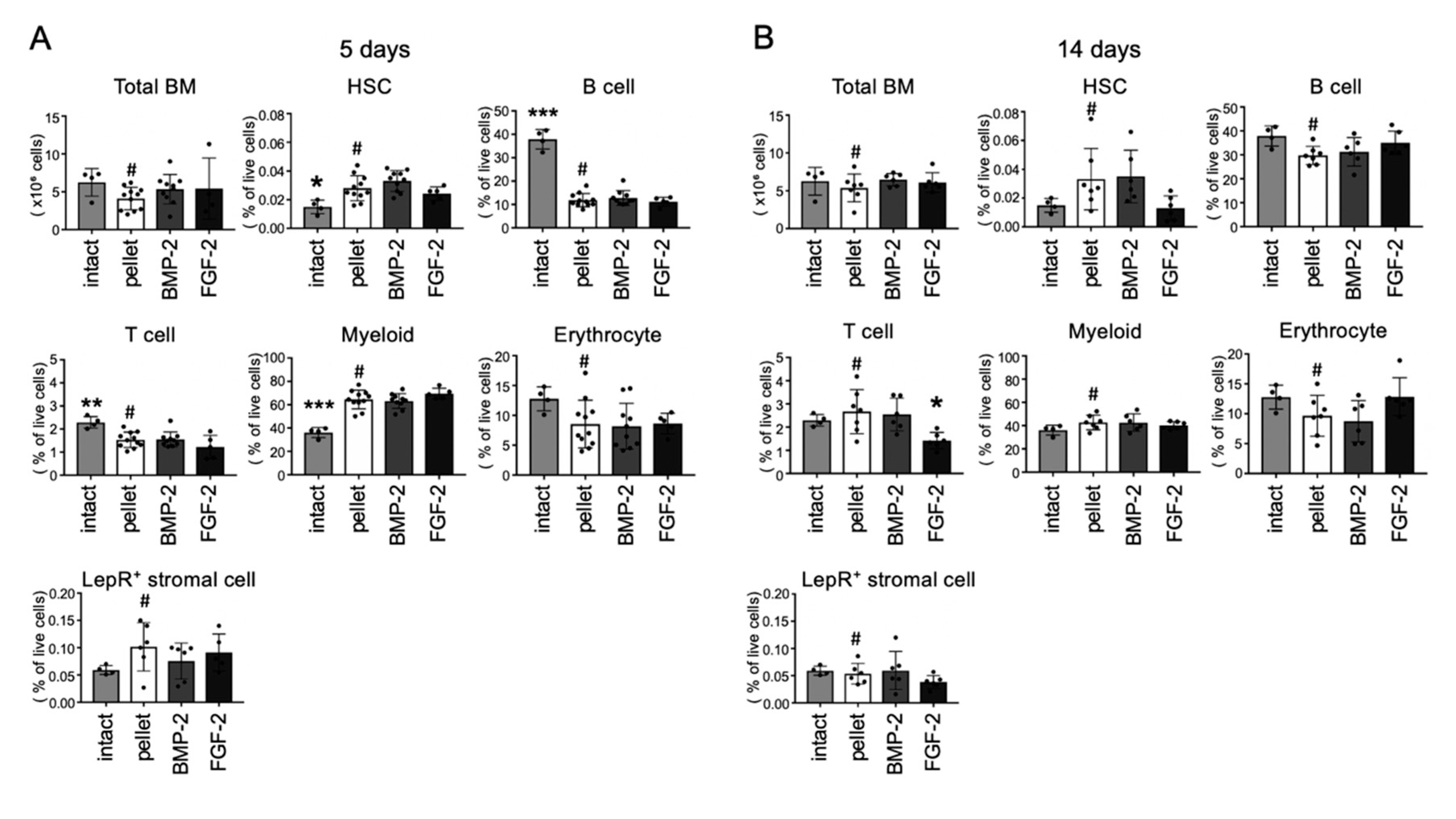
2.6. Effects of BMP-2 and FGF-2 on Bone Marrow Cell Populations
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animal Experiments
4.3. Micro-CT Analysis
4.4. Histological Analysis
4.5. Flow Cytometry
5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sakkas, A.; Wilde, F.; Heufelder, M.; Winter, K.; Schramm, A. Autogenous bone grafts in oral implantology-is it still a “gold standard”? A consecutive review of 279 patients with 456 clinical procedures. Int. J. Implant Dent. 2017, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Moses, O.; Nemcovsky, C.E.; Langer, Y.; Tal, H. Severely resorbed mandible treated with iliac crest autogenous bone graft and dental implants: 17-year follow-up. Int. J. Oral Maxillofac. Implants 2007, 22, 1017–1021. [Google Scholar] [PubMed]
- Jensen, A.T.; Jensen, S.S.; Worsaae, N. Complications related to bone augmentation procedures of localized defects in the alveolar ridge. A retrospective clinical study. Oral Maxillofac. Surg. 2016, 20, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Nkenke, E.; Neukam, F.W. Autogenous bone harvesting and grafting in advanced jaw resorption: Morbidity, resorption and implant survival. Eur. J. Oral Implantol. 2014, 7 (Suppl. 2), S203–S217. [Google Scholar] [PubMed]
- Ho-Shui-Ling, A.; Bolander, J.; Rustom, L.E.; Johnson, A.W.; Luyten, F.P.; Picart, C. Bone regeneration strategies: Engineered scaffolds, bioactive molecules and stem cells current stage and future perspectives. Biomaterials 2018, 180, 143–162. [Google Scholar] [CrossRef]
- Szpalski, C.; Wetterau, M.; Barr, J.; Warren, S.M. Bone tissue engineering: Current strategies and techniques--part I: Scaffolds. Tissue Eng. Part B Rev. 2012, 18, 246–257. [Google Scholar] [CrossRef]
- Amini, A.R.; Adams, D.J.; Laurencin, C.T.; Nukavarapu, S.P. Optimally porous and biomechanically compatible scaffolds for large-area bone regeneration. Tissue Eng. Part A 2012, 18, 1376–1388. [Google Scholar] [CrossRef] [Green Version]
- Einhorn, T.A.; Gerstenfeld, L.C. Fracture healing: Mechanisms and interventions. Nat. Rev. Rheumatol. 2015, 11, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Burkus, J.K.; Heim, S.E.; Gornet, M.F.; Zdeblick, T.A. Is INFUSE bone graft superior to autograft bone? An integrated analysis of clinical trials using the LT-CAGE lumbar tapered fusion device. J. Spinal Disord. Tech. 2003, 16, 113–122. [Google Scholar] [CrossRef]
- Burkus, J.K.; Sandhu, H.S.; Gornet, M.F.; Longley, M.C. Use of rhBMP-2 in combination with structural cortical allografts: Clinical and radiographic outcomes in anterior lumbar spinal surgery. J. Bone Joint Surg. Am. 2005, 87, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, M.; Akamatsu, M.; Machigashira, M.; Hara, Y.; Sakagami, R.; Hirofuji, T.; Hamachi, T.; Maeda, K.; Yokota, M.; Kido, J.; et al. FGF-2 stimulates periodontal regeneration: Results of a multi-center randomized clinical trial. J. Dent. Res. 2011, 90, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Fiorellini, J.P.; Howell, T.H.; Cochran, D.; Malmquist, J.; Lilly, L.C.; Spagnoli, D.; Toljanic, J.; Jones, A.; Nevins, M. Randomized study evaluating recombinant human bone morphogenetic protein-2 for extraction socket augmentation. J. Periodontol. 2005, 76, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.; Grose, R. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116–129. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Marie, P.J. Fibroblast growth factor signaling in skeletal development and disease. Genes Dev. 2015, 29, 1463–1486. [Google Scholar] [CrossRef] [Green Version]
- Itoh, N.; Ornitz, D.M. Fibroblast growth factors: From molecular evolution to roles in development, metabolism and disease. J. Biochem. 2011, 149, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Charoenlarp, P.; Rajendran, A.K.; Iseki, S. Role of fibroblast growth factors in bone regeneration. Inflamm. Regen. 2017, 37, 10. [Google Scholar] [CrossRef] [Green Version]
- Lowery, J.W.; Rosen, V. The BMP Pathway and Its Inhibitors in the Skeleton. Physiol. Rev. 2018, 98, 2431–2452. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar] [CrossRef]
- Urist, M.R. Bone: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Urist, M.R.; Sato, K.; Brownell, A.G.; Malinin, T.I.; Lietze, A.; Huo, Y.K.; Prolo, D.J.; Oklund, S.; Finerman, G.A.; DeLange, R.J. Human bone morphogenetic protein (hBMP). Proc. Soc. Exp. Biol. Med. 1983, 173, 194–199. [Google Scholar] [CrossRef]
- Wang, E.A.; Rosen, V.; Cordes, P.; Hewick, R.M.; Kriz, M.J.; Luxenberg, D.P.; Sibley, B.S.; Wozney, J.M. Purification and characterization of other distinct bone-inducing factors. Proc. Natl. Acad. Sci. USA 1988, 85, 9484–9488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asahina, I. Bone Morphogenetic Proteins: Their History and Characteristics. J. Hard Tissue Biol. 2014, 23, 283–286. [Google Scholar] [CrossRef] [Green Version]
- James, A.W.; LaChaud, G.; Shen, J.; Asatrian, G.; Nguyen, V.; Zhang, X.; Ting, K.; Soo, C. A Review of the Clinical Side Effects of Bone Morphogenetic Protein-2. Tissue Eng. Part B Rev. 2016, 22, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Ono, M.; Oida, Y.; Hara, E.S.; Komori, T.; Akiyama, K.; Nguyen, H.T.T.; Aung, K.T.; Pham, H.T.; Tosa, I.; et al. Bone Marrow Cells Inhibit BMP-2-Induced Osteoblast Activity in the Marrow Environment. J. Bone Miner Res. 2019, 34, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Nagayasu-Tanaka, T.; Nozaki, T.; Miki, K.; Sawada, K.; Kitamura, M.; Murakami, S. FGF-2 promotes initial osseointegration and enhances stability of implants with low primary stability. Clin. Oral Implant. Res. 2017, 28, 291–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Globus, R.K.; Patterson-Buckendahl, P.; Gospodarowicz, D. Regulation of bovine bone cell proliferation by fibroblast growth factor and transforming growth factor beta. Endocrinology 1988, 123, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Collin-Osdoby, P.; Rothe, L.; Bekker, S.; Anderson, F.; Huang, Y.; Osdoby, P. Basic fibroblast growth factor stimulates osteoclast recruitment, development, and bone pit resorption in association with angiogenesis in vivo on the chick chorioallantoic membrane and activates isolated avian osteoclast resorption in vitro. J. Bone Miner Res. 2002, 17, 1859–1871. [Google Scholar] [CrossRef] [PubMed]
- Hayek, A.; Culler, F.L.; Beattie, G.M.; Lopez, A.D.; Cuevas, P.; Baird, A. An in vivo model for study of the angiogenic effects of basic fibroblast growth factor. Biochem. Biophys. Res. Commun. 1987, 147, 876–880. [Google Scholar] [CrossRef]
- Pearson, H.B.; Mason, D.E.; Kegelman, C.D.; Zhao, L.; Dawahare, J.H.; Kacena, M.A.; Boerckel, J.D. Effects of Bone Morphogenetic Protein-2 on Neovascularization During Large Bone Defect Regeneration. Tissue Eng. Part A 2019, 25, 1623–1634. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, H.; Nakamura, K.; Tabata, Y.; Ikada, Y.; Aoyama, I.; Anzai, J.; Nakamura, T.; Hiyama, Y.; Tamura, M. Acceleration of fracture healing in nonhuman primates by fibroblast growth factor-2. J. Clin. Endocrinol. Metab. 2001, 86, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Hughes-Fulford, M.; Li, C.F. The role of FGF-2 and BMP-2 in regulation of gene induction, cell proliferation and mineralization. J. Orthop. Surg. Res. 2011, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Oliver, L.J.; Rifkin, D.B.; Gabrilove, J.; Hannocks, M.J.; Wilson, E.L. Long-term culture of human bone marrow stromal cells in the presence of basic fibroblast growth factor. Growth Factors 1990, 3, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Tensho, K.; Nakaya, H.; Nawata, M.; Okabe, T.; Wakitani, S. Low dose fibroblast growth factor-2 (FGF-2) enhances bone morphogenetic protein-2 (BMP-2)-induced ectopic bone formation in mice. Bone 2005, 36, 399–407. [Google Scholar] [CrossRef]
- Schliermann, A.; Nickel, J. Unraveling the Connection between Fibroblast Growth Factor and Bone Morphogenetic Protein Signaling. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel Meguid, E.; Ke, Y.; Ji, J.; El-Hashash, A.H.K. Stem cells applications in bone and tooth repair and regeneration: New insights, tools, and hopes. J. Cell Physiol. 2018, 233, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Hattori, H.; Yasue, K.; Mizutani, H.; Ueda, M.; Kaneda, T.; Hoshino, T. Development of hemopoietic bone marrow within the ectopic bone induced by bone morphogenetic protein. Blood Cells 1994, 20, 191–199. [Google Scholar]
- Kusumoto, K.; Bessho, K.; Fujimura, K.; Konishi, Y.; Ogawa, Y.; Iizuka, T. Comparative study of bone marrow induced by purified BMP and recombinant human BMP-2. Biochem. Biophys. Res. Commun. 1995, 215, 205–211. [Google Scholar] [CrossRef]
- Goldman, D.C.; Bailey, A.S.; Pfaffle, D.L.; Al Masri, A.; Christian, J.L.; Fleming, W.H. BMP4 regulates the hematopoietic stem cell niche. Blood 2009, 114, 4393–4401. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Saunders, T.L.; Enikolopov, G.; Morrison, S.J. Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 2012, 481, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Itkin, T.; Ludin, A.; Gradus, B.; Gur-Cohen, S.; Kalinkovich, A.; Schajnovitz, A.; Ovadya, Y.; Kollet, O.; Canaani, J.; Shezen, E.; et al. FGF-2 expands murine hematopoietic stem and progenitor cells via proliferation of stromal cells, c-Kit activation, and CXCL12 down-regulation. Blood 2012, 120, 1843–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, M.; Sonoyama, W.; Nema, K.; Hara, E.S.; Oida, Y.; Pham, H.T.; Yamamoto, K.; Hirota, K.; Sugama, K.; Sebald, W.; et al. Regeneration of calvarial defects with Escherichia coli -derived rhBMP-2 adsorbed in PLGA membrane. Cells Tissues Organs 2013, 198, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Sonoyama, W.; Yamamoto, K.; Oida, Y.; Akiyama, K.; Shinkawa, S.; Nakajima, R.; Pham, H.T.; Hara, E.S.; Kuboki, T. Efficient bone formation in a swine socket lift model using Escherichia coli-derived recombinant human bone morphogenetic protein-2 adsorbed in beta-tricalcium phosphate. Cells Tissues Organs 2014, 199, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Kuroda, Y.; Nango, N.; Shimoda, K.; Kubota, Y.; Ema, M.; Bakiri, L.; Wagner, E.F.; Takeda, Y.; Yashiro, W.; et al. Osteogenic capillaries orchestrate growth plate-independent ossification of the malleus. Development 2015, 142, 3912–3920. [Google Scholar] [CrossRef] [Green Version]
- Kikuta, J.; Wada, Y.; Kowada, T.; Wang, Z.; Sun-Wada, G.H.; Nishiyama, I.; Mizukami, S.; Maiya, N.; Yasuda, H.; Kumanogoh, A.; et al. Dynamic visualization of RANKL and Th17-mediated osteoclast function. J. Clin. Investig. 2013, 123, 866–873. [Google Scholar] [CrossRef]
- Komori, T.; Ono, M.; Hara, E.S.; Ueda, J.; Nguyen, H.T.T.; Nguyen, H.T.; Yonezawa, T.; Maeba, T.; Kimura-Ono, A.; Takarada, T.; et al. Type IV collagen alpha6 chain is a regulator of keratin 10 in keratinization of oral mucosal epithelium. Sci. Rep. 2018, 8, 2612. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Conjugation | Clone | Source | Cat.# |
|---|---|---|---|---|
| CD3e | APC | 145-2C11 | BioLegend | 100312 |
| CD11b | APC | M1/70 | BioLegend | 101212 |
| CD16/32 | 2.4G2 | BD Biosciences | 553142 | |
| CD31 | APC | MEC13.3 | BioLegend | 102509 |
| CD31 | PE | MEC13.3 | BioLegend | 102507 |
| CD45 | APC | 30-F11 | BioLegend | 103111 |
| CD45R (B220) | APC | RA3-6B2 | BioLegend | 103211 |
| CD48 | BV421 | HM48-1 | BioLegend | 103428 |
| CD117 (c-Kit) | FITC | 2B8 | BioLegend | 105806 |
| CD150 | PE | TC15-12F12.2 | BioLegend | 115903 |
| Leptin Receptor | Biotin | R&D systems | BAF497 | |
| Lineage marker cocktail | APC | 145-2C11(CD3e), M1/70(CD11b), RA3-6B2 (CD45R/B220), TER-119 (Ly76), RB6-8C5 (Ly6G/C) | BD Biosciences | 51-9003632 |
| Ly-6A/E (Sca-1) | PE-Cy7 | D7 | BioLegend | 108114 |
| Ly-6G/C (Gr-1) | BV421 | RB6-8C5 | BD Biosciences | 562709 |
| Streptavidin | BV421 | BioLegend | 405226 | |
| Ter119 | APC | TER-119 | BioLegend | 116211 |
| 7-AAD | BioLegend | 420404 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nosho, S.; Tosa, I.; Ono, M.; Hara, E.S.; Ishibashi, K.; Mikai, A.; Tanaka, Y.; Kimura-Ono, A.; Komori, T.; Maekawa, K.; et al. Distinct Osteogenic Potentials of BMP-2 and FGF-2 in Extramedullary and Medullary Microenvironments. Int. J. Mol. Sci. 2020, 21, 7967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217967
Nosho S, Tosa I, Ono M, Hara ES, Ishibashi K, Mikai A, Tanaka Y, Kimura-Ono A, Komori T, Maekawa K, et al. Distinct Osteogenic Potentials of BMP-2 and FGF-2 in Extramedullary and Medullary Microenvironments. International Journal of Molecular Sciences. 2020; 21(21):7967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217967
Chicago/Turabian StyleNosho, Shuji, Ikue Tosa, Mitsuaki Ono, Emilio Satoshi Hara, Kei Ishibashi, Akihiro Mikai, Yukie Tanaka, Aya Kimura-Ono, Taishi Komori, Kenji Maekawa, and et al. 2020. "Distinct Osteogenic Potentials of BMP-2 and FGF-2 in Extramedullary and Medullary Microenvironments" International Journal of Molecular Sciences 21, no. 21: 7967. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217967