Identification of Peritrophins and Antiviral Effect of Bm01504 against BmNPV in the Silkworm, Bombyx mori

Abstract
:1. Introduction
2. Results
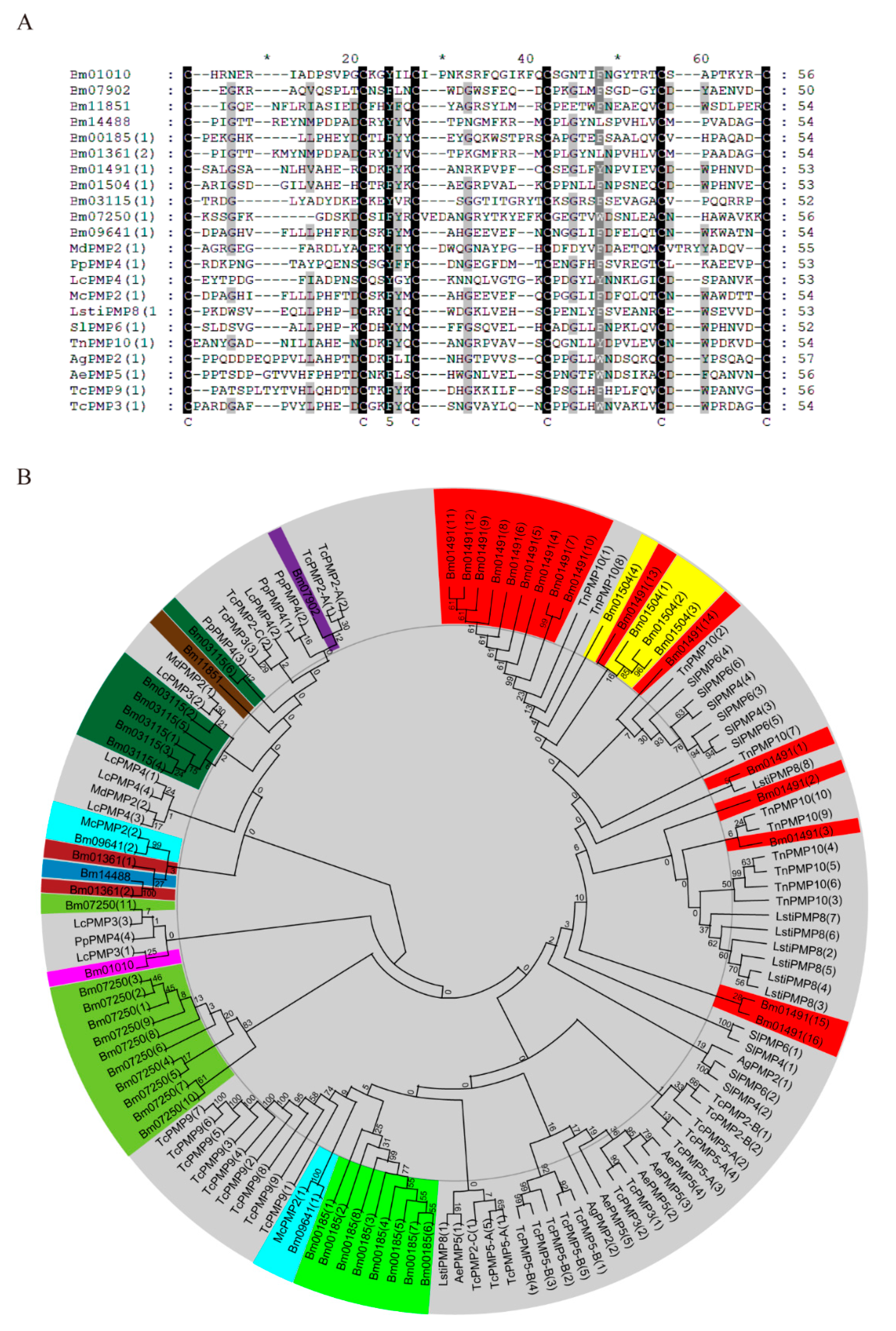
2.1. Identification and Bioinformatics Analysis of Peritrophins in Bombyx mori
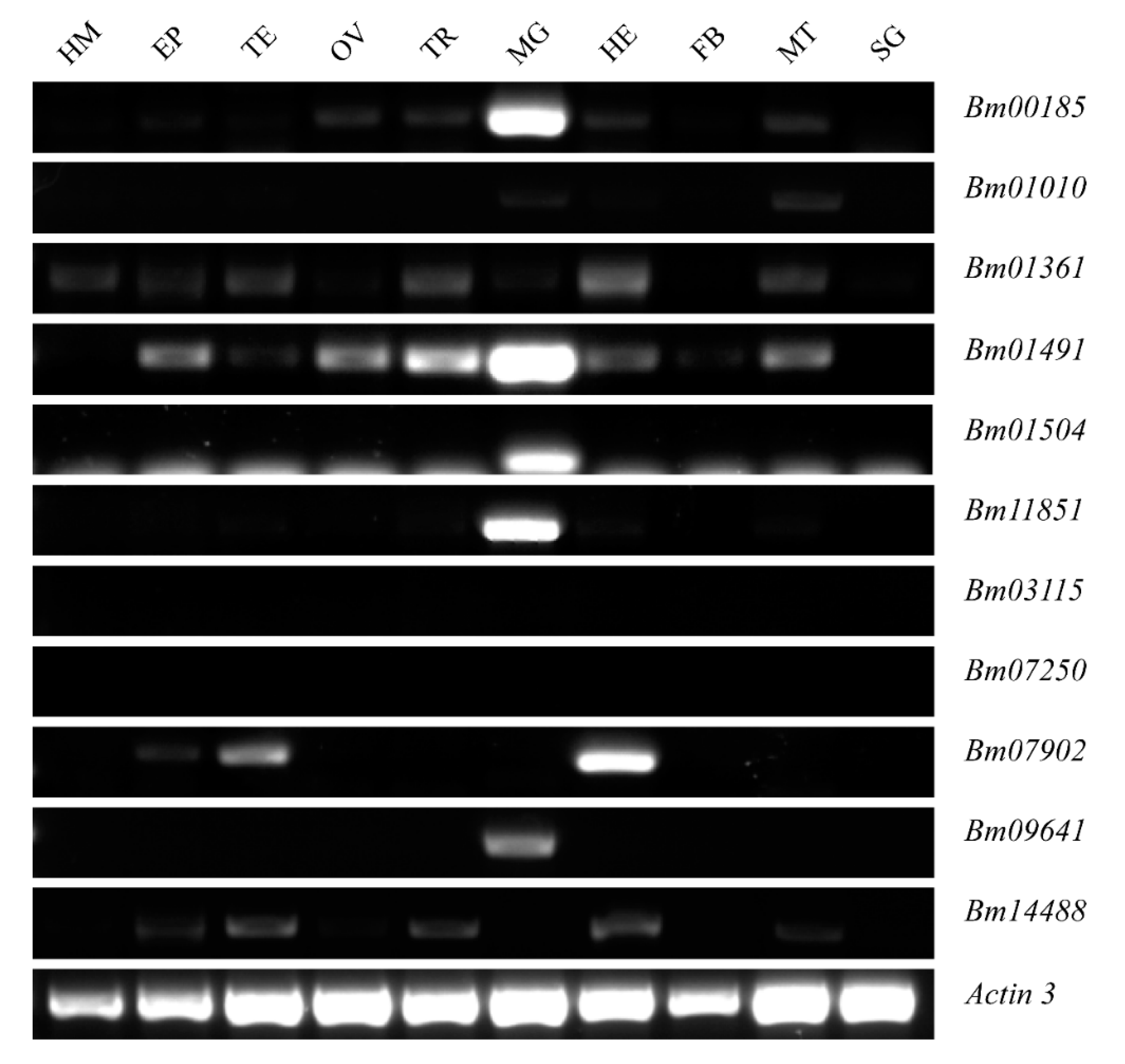
2.2. Expression Patterns of Peritrophins in Silkworm, Bombyx mori
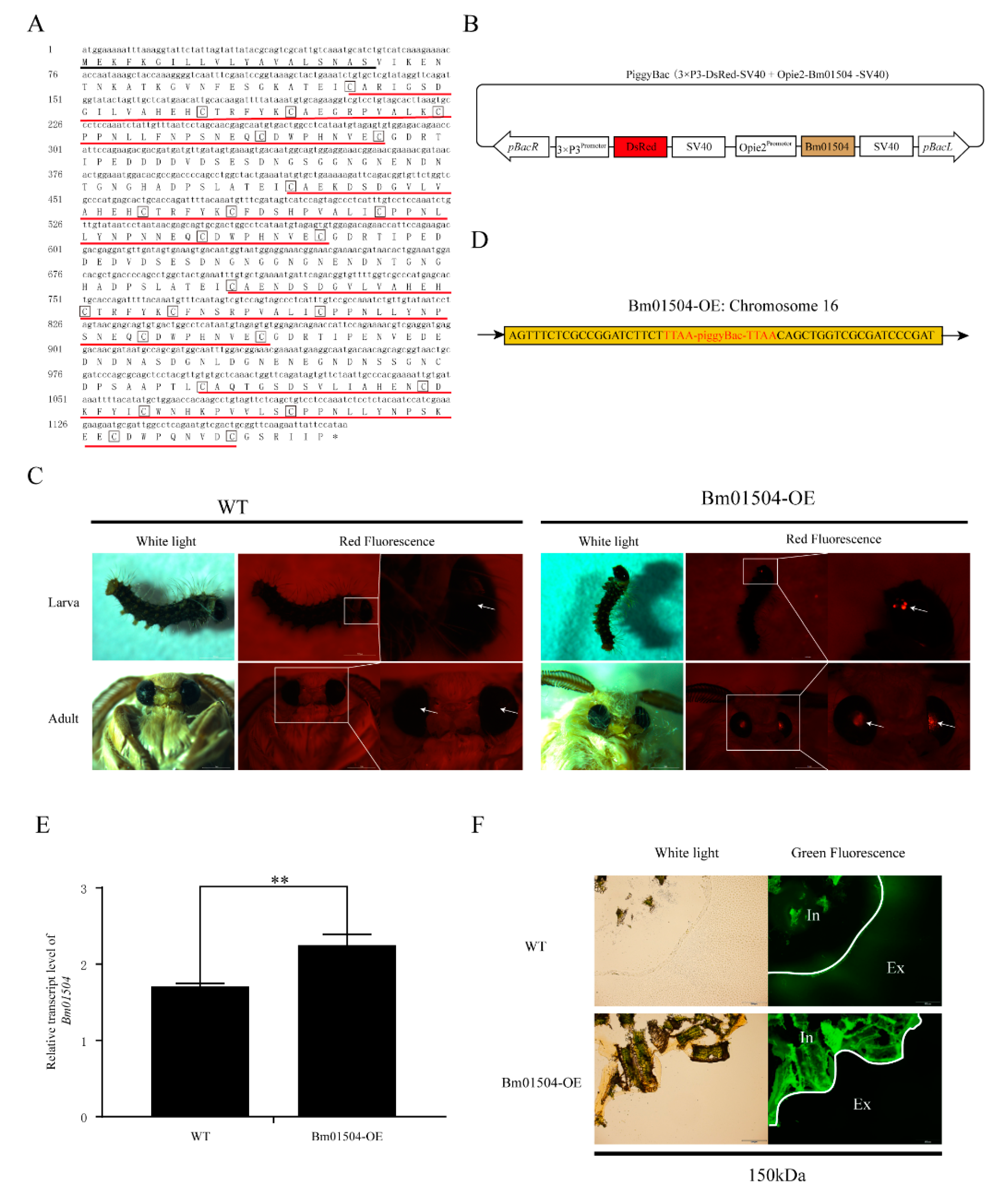
2.3. Generation of Bm01504 Overexpression (Bm01504-OE) in Transgenic Silkworms
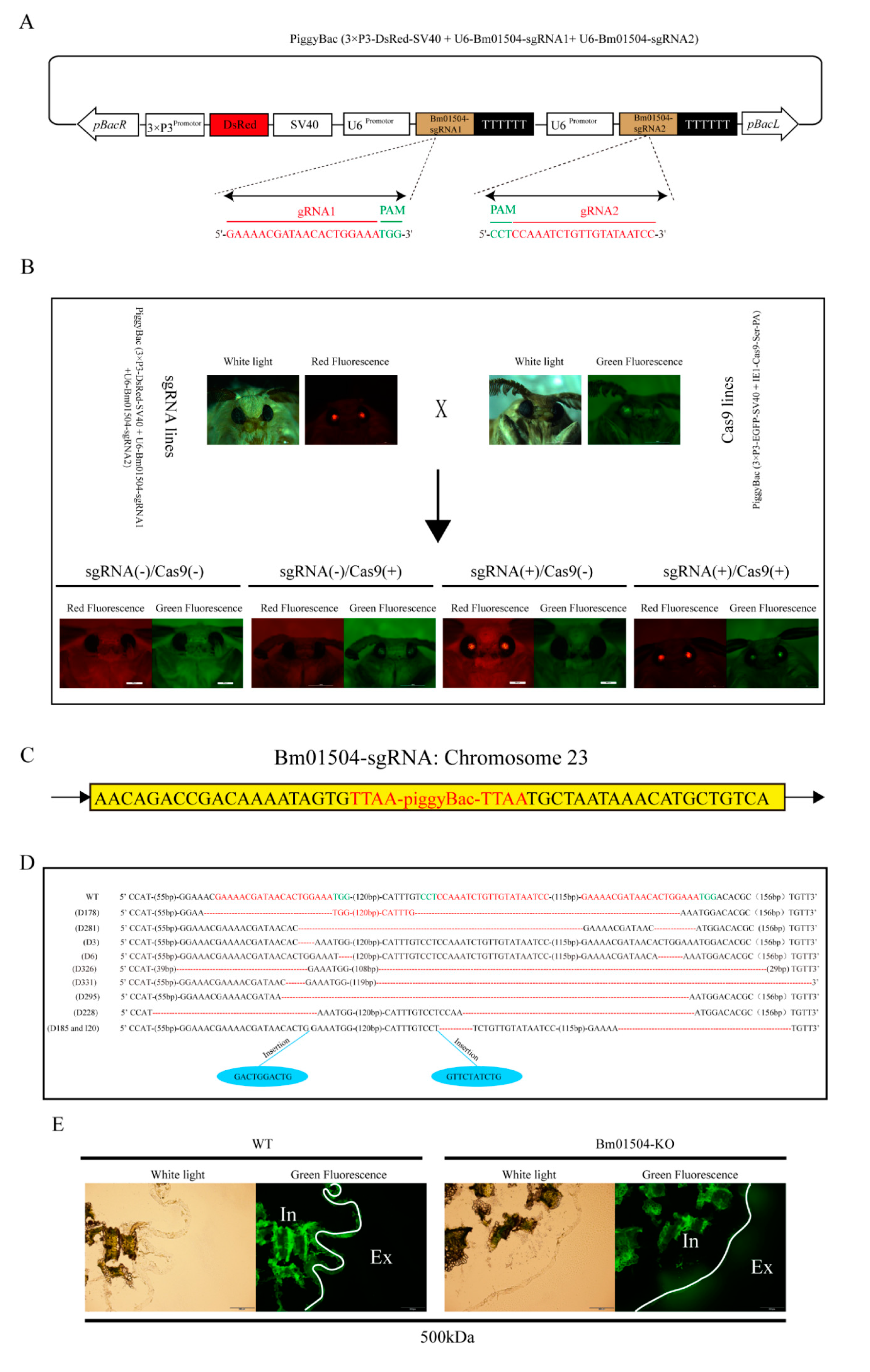
2.4. Generation of Bm01504 Knockout (Bm01504-KO) in Transgenic Silkworms
2.5. Economic Characteristics of Transgenic Silkworm Lines
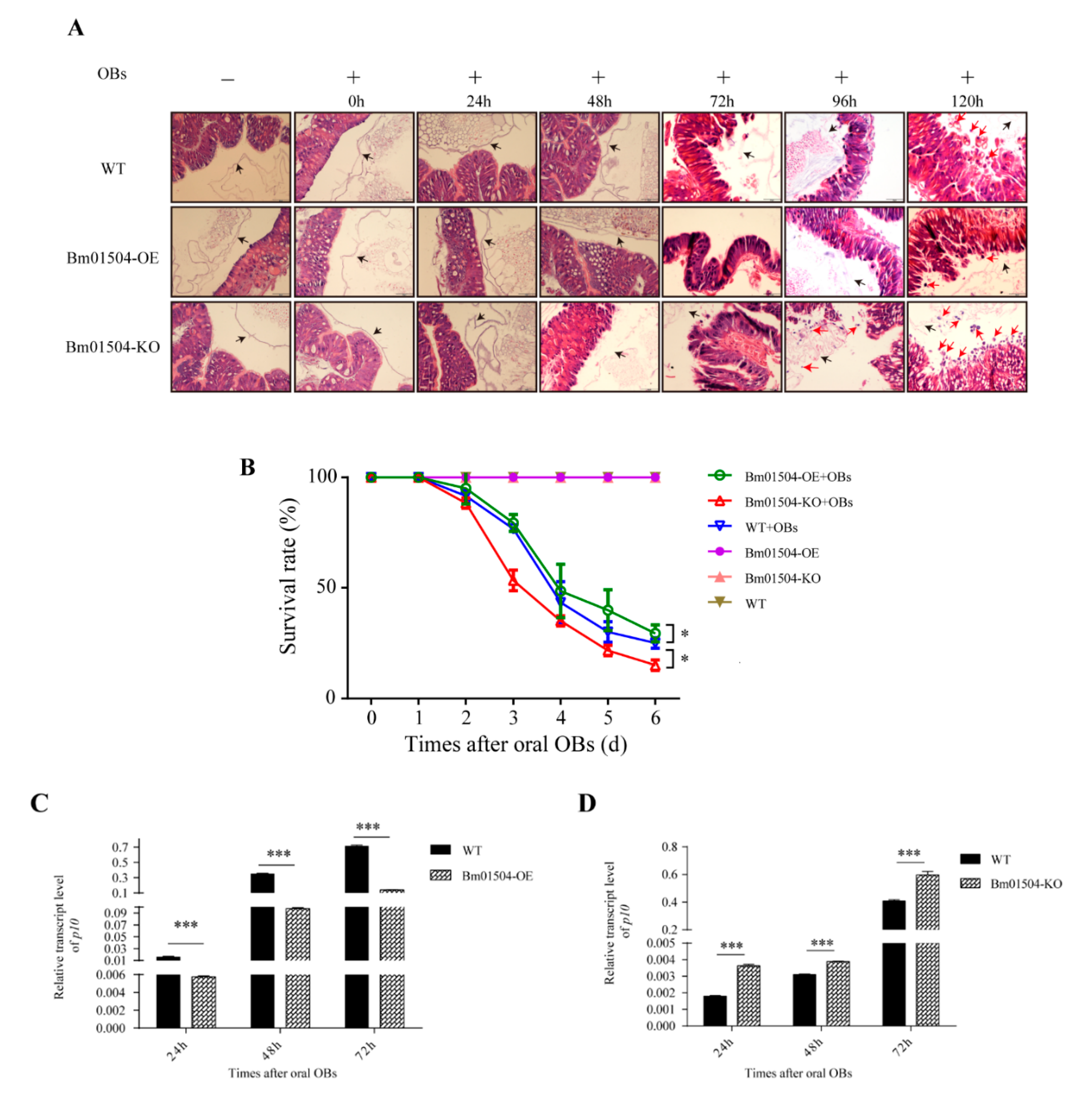
2.6. Transgenic Larvae have Different Levels of Resistance to BmNPV
3. Discussion
4. Materials and Methods
4.1. Silkworm Strain and Virus
4.2. Identification of Peritrophins in the Bombyx mori Genome Database
4.3. Phylogenetic Analysis and Multiple Sequence Alignment
4.4. Oral Inoculation of OBs
4.5. Semi-Quantitative RT-PCR and qRT-PCR
4.6. Vector Construction
4.7. Microinjection and sCreening
4.8. Analysis of Insertion Site
4.9. FITC-Dextran Permeability Assays
4.10. Analysis of Economic Characteristics
4.11. Paraffin Sectioning
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Bm01504-OE | Bm01504 overexpression |
| Bm01504-KO | Bm01504 knockout |
| WT | Wild type |
| CBD | Chitin binding domain |
| BmNPV | B. mori nucleopolyhedrovirus |
| PM | Peritrophic membrane |
| PMP | Peritrophic membrane protein |
| CBM_14 | Carbohydrate Binding Module family14 |
| ChtBD2 | Type 2 CBD |
| CPAP | Cuticle proteins analogous to peritrophin |
| CHT | Chitinase |
| CDA | Chitin deacetylase |
| MUC | Mucin |
| MD | Mucin domain |
| HMM | Hidden Markov model |
| mRNA | Messenger RNA |
| RT-PCR | Reverse transcription polymerase chain reaction |
| qRT-PCR | Quantitative RT-PCR |
| FITC | Fluoresceine isothiocyanate |
| DsRed | Discosoma sp. red fluorescent protein |
| Opie2 | Orgyia pseudotsugata multicapsid nucleopolyhedrovirus (OpMNPV) immediate-early 2 (ie2) |
| SV40 | Terminator of Simian virus 40 |
| PAM | Protospacer adjacent motif |
| ODV | Occlusion-derived viruses |
| BV | Budded virus |
| OB | Occlusion body |
| WSSV | White spot syndrome virus |
| MUSCLE | Multiple sequence comparison by log-expectation |
| CDS | Coding sequence |
References
- Hegedus, D.D.; Toprak, U.; Erlandson, M. Peritrophic matrix formation. J. Insect Physiol. 2019, 117, 103898. [Google Scholar] [CrossRef]
- Erlandson, M.A.; Toprak, U.; Hegedus, D.D. Role of the peritrophic matrix in insect-pathogen interactions. J. Insect Physiol. 2019, 117, 103894. [Google Scholar] [CrossRef]
- Traub-Csekö, Y.M.; Sadlova, J.; Homola, M.; Myskova, J.; Jancarova, M.; Volf, P. Refractoriness of Sergentomyia schwetzi to Leishmania spp. is mediated by the peritrophic matrix. PloS Negl. Trop. Dis. 2018, 12, e0006382. [Google Scholar]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Tellam, R.L.; Wijffels, G.; Willadsen, P. Peritrophic matrix proteins. Insect Biochem. Mol. Biol. 1999, 29, 87–101. [Google Scholar] [CrossRef]
- Elvin, C.M.; Vuocolo, T.; Pearson, R.D.; East, I.J.; Riding, G.A.; Eisemann, C.H.; Tellam, R.L. Characterization of a major peritrophic membrane protein, peritrophin-44, from the larvae of Lucilia cuprina cDNA and deduced amino acid sequences. J. Biol. Chem. 1996, 271, 8925–8935. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, J.; Yao, L.; Li, S.; Chen, Y.; Yang, H.; Fan, D. Chitin deacetylase: A potential target for Mythimna separata (Walker) control. Arch. Insect Biochem. Physiol. 2020, 104, e21666. [Google Scholar] [CrossRef]
- Rodríguez-de la Noval, C.; Rodríguez-Cabrera, L.; Izquierdo, L.; Espinosa, L.A.; Hernandez, D.; Ponce, M.; Moran-Bertot, I.; Tellez-Rodríguez, P.; Borras-Hidalgo, O.; Huang, S.; et al. Functional expression of a peritrophin A-like SfPER protein is required for larval development in Spodoptera frugiperda (Lepidoptera: Noctuidae). Sci. Rep. 2019, 9, 2630. [Google Scholar] [CrossRef]
- Liu, X.; Cooper, A.M.; Yu, Z.; Silver, K.; Zhang, J.; Zhu, K.Y. Progress and prospects of arthropod chitin pathways and structures as targets for pest management. Pestic. Biochem. Physiol. 2019, 161, 33–46. [Google Scholar] [CrossRef]
- Konno, K.; Mitsuhashi, W. The peritrophic membrane as a target of proteins that play important roles in plant defense and microbial attack. J. Insect Physiol. 2019, 117, 103912. [Google Scholar] [CrossRef]
- Sandoval-Mojica, A.F.; Scharf, M.E. Silencing gut genes associated with the peritrophic matrix of Reticulitermes flavipes (Blattodea: Rhinotermitidae) increases susceptibility to termiticides. Insect Mol. Biol. 2016, 25, 734–744. [Google Scholar] [CrossRef]
- Mohan, S.; Ma, P.W.K.; Pechan, T.; Bassford, E.R.; Williams, W.P.; Luthe, D.S. Degradation of the S. frugiperda peritrophic matrix by an inducible maize cysteine protease. J. Insect Physiol. 2006, 52, 21–28. [Google Scholar] [CrossRef]
- Liu, X.; Cooper, A.M.; Zhang, J.; Zhu, K.Y. Biosynthesis, modifications and degradation of chitin in the formation and turnover of peritrophic matrix in insects. J. Insect Physiol. 2019, 114, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; De Schutter, K.; Smargiasso, N.; De Pauw, E.; Van Damme, E.J.; Smagghe, G. The N-glycan profile of the peritrophic membrane in the Colorado potato beetle larva (Leptinotarsa decemlineata). J. Insect Physiol. 2019, 115, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chamankhah, M.; Visal-Shah, S.; Hemmingsen, S.M.; Erlandson, M.; Braun, L.; Alting-Mees, M.; Khachatourians, G.G.; O’Grady, M.; Hegedus, D.D. Modeling the structure of the Type I peritrophic matrix: Characterization of a Mamestra configurata intestinal mucin and a novel peritrophin containing 19 chitin binding domains. Insect Biochem. Mol. Biol. 2004, 34, 1101–1115. [Google Scholar] [CrossRef] [PubMed]
- Toprak, U.; Erlandson, M.; Hegedus, D.D. Peritrophic matrix proteins. Trends Entomol. 2010, 6, 23–51. [Google Scholar]
- Tetreau, G.; Dittmer, N.T.; Cao, X.; Agrawal, S.; Chen, Y.-R.; Muthukrishnan, S.; Haobo, J.; Blissard, G.W.; Kanost, M.R.; Wang, P. Analysis of chitin-binding proteins from Manduca sexta provides new insights into evolution of peritrophin A-type chitin-binding domains in insects. Insect Biochem. Mol. Biol. 2015, 62, 127–141. [Google Scholar] [CrossRef] [Green Version]
- Jasrapuria, S.; Arakane, Y.; Osman, G.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Genes encoding proteins with peritrophin A-type chitin-binding domains in Tribolium castaneum are grouped into three distinct families based on phylogeny, expression and function. Insect Biochem. Mol. Biol. 2010, 40, 214–227. [Google Scholar] [CrossRef]
- Toprak, U.; Erlandson, M.; Baldwin, D.; Karcz, S.; Wan, L.; Coutu, C.; Gillott, C.; Hegedus, D.D. Identification of the Mamestra configurata(Lepidoptera: Noctuidae) peritrophic matrix proteins and enzymes involved in peritrophic matrix chitin metabolism. Insect Sci. 2016, 23, 656–674. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.L.; Tian, S.; Yang, H.; Zhou, X.; Xu, S.P.; Zhang, Z.Y.; Gong, J.; Hou, Y.; Xia, Q.Y. Genome-wide identification of chitin-binding proteins and characterization of BmCBP1 in the silkworm. Bombyx mori. Insect Sci. 2019, 26, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Park, M.J.; Kim, B.Y.; Jin, B.R. Molecular characterization of a chitin-binding protein with the peritrophin-A domain from the Asiatic honeybee Apis cerana. J. Asia-Pac. Entomol. 2016, 19, 963–968. [Google Scholar] [CrossRef]
- Yin, J.; Yang, S.; Li, K.; Guo, W.; Cao, Y. Identification and molecular characterization of a chitin-binding protein from the Beet Webworm, Loxostege sticticalis L. Int. J. Mol. Sci. 2014, 15, 19147–19161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.J.; Huang, L.X.; Hu, D.; Liu, L.Y.; Gu, J.; Huang, L.H.; Feng, Q.L. Cloning, expression and chitin-binding activity of two peritrophin-like protein genes in the common cutworm, Spodoptera litura. Insect Sci. 2014, 21, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Toprak, U.; Baldwin, D.; Erlandson, M.; Gillott, C.; Harris, S.; Hegedus, D.D. Expression patterns of genes encoding proteins with peritrophin A domains and protein localization in Mamestra configurata. J. Insect Physiol. 2010, 56, 1711–1720. [Google Scholar] [CrossRef]
- Mittapalli, O.; Sardesai, N.; Shukle, R.H. cDNA cloning and transcriptional expression of a peritrophin-like gene in the Hessian fly, Mayetiola destructor [Say]. Arch. Insect Biochem. Physiol. 2007, 64, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Devenport, M.; Alvarenga, P.H.; Shao, L.; Fujioka, H.; Jacobs-Lorena, M. Identification of the Aedes aegypti peritrophic matrix protein AeIMUCI as a heme-binding protein. Biochemistry 2006, 45, 9540–9549. [Google Scholar] [CrossRef]
- Shao, L.; Devenport, M.; Fujioka, H.; Ghosh, A.; Jacobs-Lorena, M. Identification and characterization of a novel peritrophic matrix protein, Ae-Aper50, and the microvillar membrane protein, AEG12, from the mosquito. Aedes aegypti. Insect Biochem. Mol. Biol. 2005, 35, 947–959. [Google Scholar] [CrossRef]
- Wang, P.; Li, G.; Granados, R.R. Identification of two new peritrophic membrane proteins from larval Trichoplusia ni: Structural characteristics and their functions in the protease rich insect gut. Insect Biochem. Mol. Biol. 2004, 34, 215–227. [Google Scholar] [CrossRef]
- Tellam, R.L.; Vuocolo, T.; Eisemann, C.; Briscoe, S.; Riding, G.; Elvin, C.; Pearson, R. Identification of an immuno-protective mucin-like protein, peritrophin-55, from the peritrophic matrix of Lucilia cuprina larvae. Insect Biochem. Mol. Biol. 2003, 33, 239–252. [Google Scholar] [CrossRef]
- Vuocolo, T.; Eisemann, C.H.; Pearson, R.D.; Willadsen, P.; Tellam, R.L. Identification and molecular characterisation of a peritrophin gene, peritrophin-48, from the myiasis fly Chrysomya bezziana. Insect Biochem. Mol. Biol. 2001, 31, 919–932. [Google Scholar] [CrossRef]
- Tellam, R.L.; Eisemann, C.; Casu, R.; Pearson, R. The intrinsic peritrophic matrix protein peritrophin-95 from larvae of Lucilia cuprina is synthesised in the cardia and regurgitated or excreted as a highly immunogenic protein. Insect Biochem. Mol. Biol. 2000, 30, 9–17. [Google Scholar] [CrossRef]
- Shen, Z.; Jacobs-Lorena, M.A. Type I peritrophic matrix protein from the malaria vector Anopheles gambiae binds to chitin. J. Biol. Chem. 1998, 273, 17665–17670. [Google Scholar] [CrossRef] [Green Version]
- Schorderet, S.; Pearson, R.D.; Vuocolo, T.; Eisemann, C.; Tellam, R.L. cDNA and deduced amino acid sequences of a peritrophic membrane glycoprotein, ‘peritrophin-48′, from the larvae of Lucilia cuprina. Insect Biochem. Mol. Biol. 1998, 28, 99–111. [Google Scholar] [CrossRef]
- Sandoval-Mojica, A.F.; Scharf, M.E. Gut genes associated with the peritrophic matrix in Reticulitermes flavipes (Blattodea: Rhinotermitidae): Identification and characterization. Arch. Insect Biochem. Physiol. 2016, 92, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Devenport, M.; Fujioka, H.; Jacobs-Lorena, M. Storage and secretion of the peritrophic matrix protein Ag-Aper1 and trypsin in the midgut of Anopheles gambiae. Insect Mol. Biol. 2004, 13, 349–358. [Google Scholar] [CrossRef]
- Russell, C.J.; Xie, S.; Zhang, X.; Zhang, J.; Li, F.; Xiang, J. Envelope proteins of white spot syndrome virus (WSSV) Interact with Litopenaeus vannamei peritrophin-like protein (LvPT). PLoS ONE 2015, 10, e0144922. [Google Scholar]
- Huang, Y.; Ma, F.; Wang, W.; Ren, Q. Identification and molecular characterization of a peritrophin-like gene, involved in the antibacterial response in Chinese mitten crab, Eriocheir sinensis. Dev. Comp. Immunol. 2015, 50, 129–138. [Google Scholar] [CrossRef]
- Wang, L.; Li, F.; Wang, B.; Xiang, J. A new shrimp peritrophin-like gene from Exopalaemon carinicauda involved in white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2013, 35, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Hsu, T.C.; Huang, P.Y.; Kang, S.T.; Lo, C.F.; Huang, W.P.; Chen, L.L. Penaeus monodon chitin-binding protein (PmCBP) is involved in white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2009, 27, 460–465. [Google Scholar] [CrossRef]
- Chen, L.; Lu, L.; Wu, W.; Lo, C.; Huang, W. White spot syndrome virus envelope protein VP53A interacts with Penaeus monodon chitin-binding protein (PmCBP). Dis. Aquat. Organ. 2007, 74, 171–178. [Google Scholar] [CrossRef]
- Agrawal, S.; Kelkenberg, M.; Begum, K.; Steinfeld, L.; Williams, C.E.; Kramer, K.J.; Beeman, R.W.; Park, Y.; Muthukrishnan, S.; Merzendorfer, H. Two essential peritrophic matrix proteins mediate matrix barrier functions in the insect midgut. Insect Biochem. Mol. Biol. 2014, 49, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, P.F.; Coutinho-Abreu, I.V.; Sharma, N.K.; Robles-Murguia, M.; Ramalho-Ortigao, M. Characterization of Phlebotomus papatasi peritrophins, and the role of PpPer1 in Leishmania major survival in its natural vector. PLoS Negl. Trop. Dis. 2013, 7, e2132. [Google Scholar]
- Cheng, T.; Lin, P.; Huang, L.; Wu, Y.; Jin, S.; Liu, C.; Xia, Q. Genome-wide analysis of host responses to four different types of microorganisms in Bombyx mori (Lepidoptera: Bombycidae). J. Insect Sci. 2016, 16, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.H.; Fernandes, K.M.; Goncalves, W.G.; Zanuncio, J.C.; Serrao, J.E. A peritrophin mediates the peritrophic matrix permeability in the workers of the bees Melipona quadrifasciata and Apis mellifera. Arthropod Struct. Dev. 2019, 53, 100885. [Google Scholar] [CrossRef]
- Toprak, U.; Hegedus, D.D.; Baldwin, D.; Coutu, C.; Erlandson, M. Spatial and temporal synthesis of Mamestra configurata peritrophic matrix through a larval stadium. Insect Biochem. Mol. Biol. 2014, 54, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, W.; Miyamoto, K. Disintegration of the peritrophic membrane of silkworm larvae due to spindles of an entomopoxvirus. J. Invertebr. Pathol. 2003, 82, 34–40. [Google Scholar] [CrossRef]
- Kuraishi, T.; Binggeli, O.; Opota, O.; Buchon, N.; Lemaitre, B. Genetic evidence for a protective role of the peritrophic matrix against intestinal bacterial infection in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2011, 108, 15966–15971. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulos, G.; Rodgers, F.H.; Gendrin, M.; Wyer, C.A.; Christophides, G.K. Microbiota-induced peritrophic matrix regulates midgut homeostasis and prevents systemic infection of malaria vector mosquitoes. PLoS Pathog. 2017, 13, e1006391. [Google Scholar]
- Liu, T.H.; Dong, X.L.; Pan, C.X.; Du, G.Y.; Wu, Y.F.; Yang, J.G.; Chen, P.; Lu, C.; Pan, M.H. A newly discovered member of the Atlastin family, BmAtlastin-n, has an antiviral effect against BmNPV in Bombyx mori. Sci. Rep. 2016, 6, 28946. [Google Scholar] [CrossRef] [Green Version]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus Insecticides in Latin America: Historical Overview, Current Status and Future Perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef]
- Sudhir, K.; Glen, S.; Michael, L.; Christina, K.; Koichiro, T. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID 1 | Gene ID 2 | Chr. | Protein Length (Number of aa) | Number of CBDs |
|---|---|---|---|---|---|
| Bm01504 | BGIBMGA001504 | KWMTBOMO12505 | 21 | 392 | 4 |
| Bm00185 | BGIBMGA000185 | KWMTBOMO00962 | 2 | 1261 | 8 |
| Bm03115 | BGIBMGA003115 | KWMTBOMO02225 | 4 | 561 | 6 |
| Bm07250 | BGIBMGA007250 | KWMTBOMO10355 | 17 | 1527 | 11 |
| Bm01491 | BGIBMGA001491 | KWMTBOMO12463 | 21 | 1878 | 16 |
| Bm07902 | BGIBMGA007902 | KWMTBOMO08750 | 15 | 190 | 1 |
| Bm09641 | BGIBMGA009641 | KWMTBOMO00941 | 2 | 225 | 2 |
| Bm01361 | BGIBMGA001361 | KWMTBOMO00967 | 2 | 340 | 2 |
| Bm14488 | BGIBMGA014488 | KWMTBOMO00966 | 2 | 282 | 1 |
| Bm11851 | BGIBMGA011851 | KWMTBOMO06502 | 11 | 539 | 1 |
| Bm01010 | BGIBMGA001010 | —— | 13 | 241 | 1 |
| Name | Forward (5′→3′) | Reverse (5′→3′) | |
|---|---|---|---|
| Primers for semi-quantitative RT-PCR | Bm01504 | TGGCCTCAGAATGTCGACT | CAATAATCTAAAATCCATAATGCTAC |
| Bm00185 | CATCCTCCCCTGGGCTCAC | CGTAATCAAGGTCATTTGTTCGC | |
| Bm03115 | ACCTGTTATGAACCCGTTTGTGC | CGTTCACATTCTGGACCGCC | |
| Bm07250 | TAACAGAGCAATCTACAAATCAAGC | TGGTGGTAGAAGATTCAGTGCC | |
| Bm01491 | CGTCAATACTGGTCCTTGTAACTGT | GTAGTGTCTGATGTTTTGTCGTGC | |
| Bm07902 | TCGGACCTCGCATAGCAGC | TTTCAGCGTAATCGCAGTAGCC | |
| Bm09641 | ATGTTAGGTAAAGCCCTTAGTCTCTTG | CTAGTTTTTGTACACAATGAATTCGC | |
| Bm01361 | CATCGGCGAGACAAGAGGT | AGTGCGAAGGCGGTATCC | |
| Bm14488 | CATCGGCGAGACAAGAGGT | AGTGCGAAGGCGGTATCC | |
| Bm11851 | CCTTGTGGCTCCTGTGTTG | CAGTATCAGTGCCTTCTTCGTC | |
| Bm01010 | GGTCCGTATGGATTTATTTGCGA | ATGCCACTCCAGTTCTGTGTAAAAA | |
| Actin3 | AACACCCCGTCCTGCTCACTG | GGGCGAGACGTGTGATTTCCT | |
| Primers for qRT-PCR | Bm09641 | CTGAAGGTTCGGGCTTGGGT | TGTGCCTGCTGAGTCTGCTGTG |
| Bm01504 | TGGCCTCAGAATGTCGACT | CAATAATCTAAAATCCATAATGCTAC | |
| Bm00185 | CATCCTCCCCTGGGCTCAC | CGTAATCAAGGTCATTTGTTCGC | |
| Bm11851 | GCAGAACAGGTTTGCGACTG | GCTCAGGCTCTTGTTCTGGT | |
| Bm01491 | AAAGCTCCAGGGAGACAACG | TCCTCACCTGGAACGACTCT | |
| sw22934 | TTCGTACTGGCTCTTCTCGT | CAAAGTTGATAGCAATTCCCT | |
| p10 | TAGACGCCATTGCGGAAA | CGGGCAAACCGTCCAAA |
| Name | Primer (5′→3′) |
|---|---|
| Bm01504 (cds)-F | gatccATGGAAAAATTTAAAGGTATTCTATTAGTATTATAC |
| Bm01504 (cds)-R | gaattcCATGGAATAATTCTTGAACCGCAGT |
| U6-F1 | ggatccAGGTTATGTAGTACACATT |
| Bm01504-sgRNA1-R1 | GCTATTTCTAGCTCTAAAACTTTCCAGTGTTATCGTTTTCACTTGTAGAGCACGATATT |
| U6-R2 | CAAGTTGATAACGGACTAGCCTTATTTTAACTTGCTATTTCTAGCTCTAAAACA |
| U6-R31 | gaattcAAAAAAGCACCGACTCGGTGCCACTTTTTCAAGTTGATAACGGACTAG |
| U6-F2 | gcggccgcAGGTTATGTAGTACACATT |
| Bm01504-RNA2-R1 | GCTATTTCTAGCTCTAAAACCCAAATCTGTTGTATAATCCACTTGTAGAGCACGATATT |
| U6-R32 | tctagaAAAAAAGCACCGACTCGGTGCCACTTTTTCAAGTTGATAACGGACTAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zha, X.-L.; Yu, X.-B.; Zhang, H.-Y.; Wang, H.; Huang, X.-Z.; Shen, Y.-H.; Lu, C. Identification of Peritrophins and Antiviral Effect of Bm01504 against BmNPV in the Silkworm, Bombyx mori. Int. J. Mol. Sci. 2020, 21, 7973. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217973
Zha X-L, Yu X-B, Zhang H-Y, Wang H, Huang X-Z, Shen Y-H, Lu C. Identification of Peritrophins and Antiviral Effect of Bm01504 against BmNPV in the Silkworm, Bombyx mori. International Journal of Molecular Sciences. 2020; 21(21):7973. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217973
Chicago/Turabian StyleZha, Xu-Le, Xin-Bo Yu, Hong-Yan Zhang, Han Wang, Xian-Zhi Huang, Yi-Hong Shen, and Cheng Lu. 2020. "Identification of Peritrophins and Antiviral Effect of Bm01504 against BmNPV in the Silkworm, Bombyx mori" International Journal of Molecular Sciences 21, no. 21: 7973. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217973