The Stimulation of Neurogenesis Improves the Cognitive Status of Aging Rats Subjected to Gestational and Perinatal Deficiency of B9–12 Vitamins

 , , and
, , and 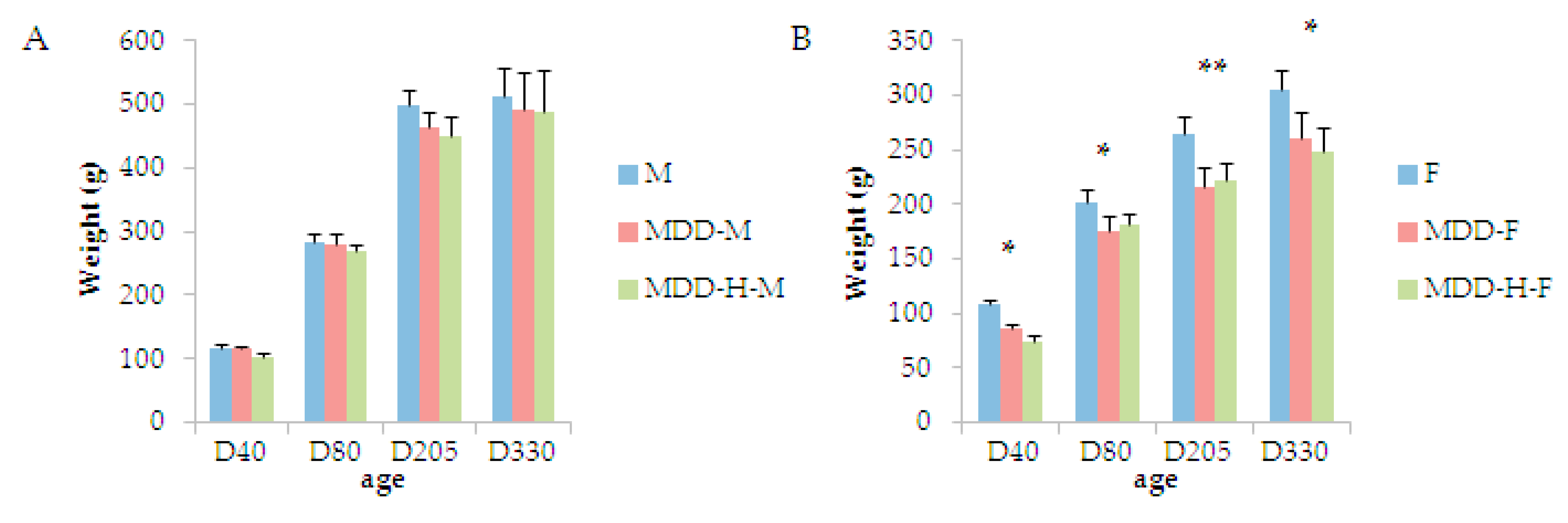{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
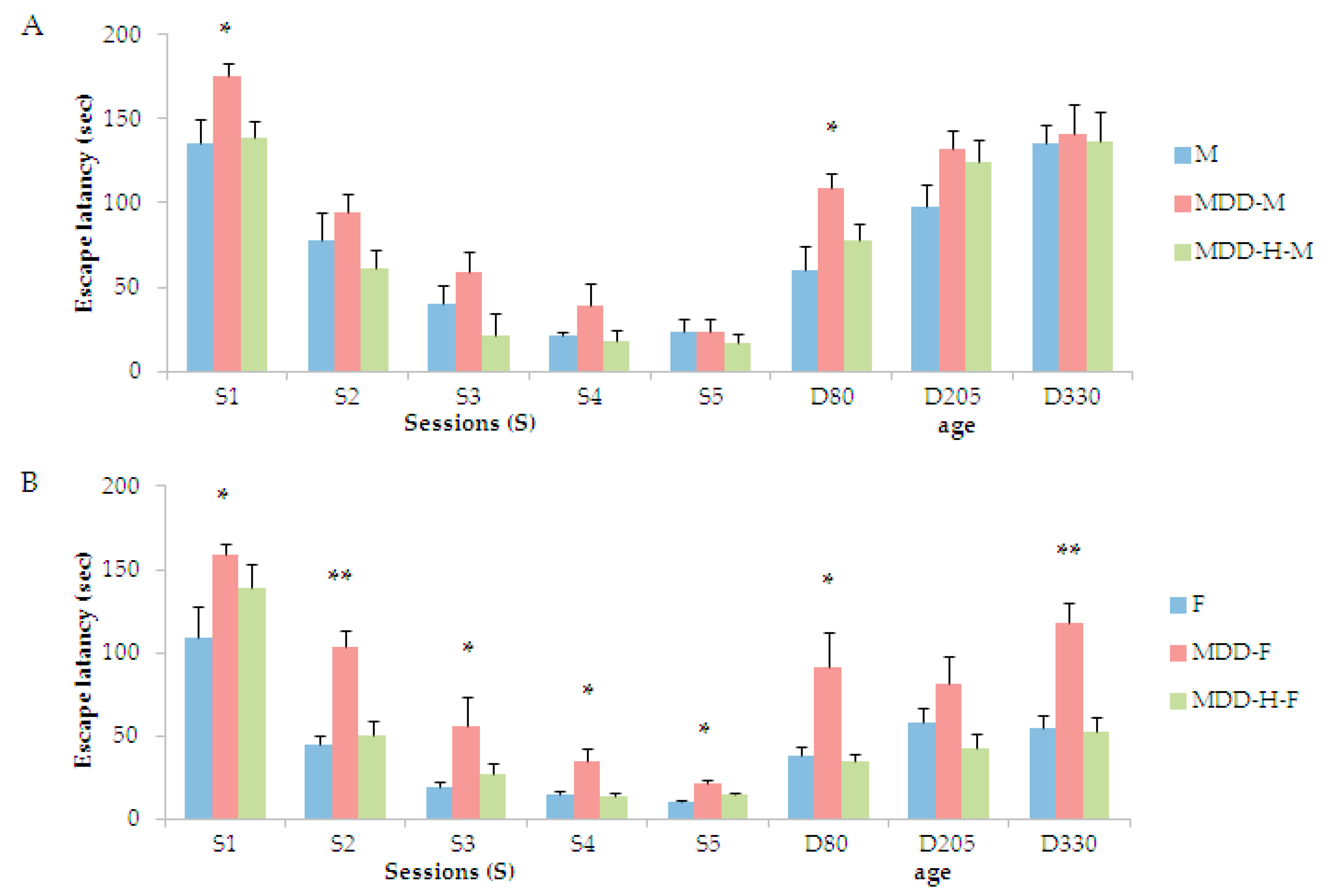
2. Results
3. Discussion
3.1. The Correction of B-Vitamin Deficiency Did Not Lead to the Rescue of All Body Parameters
3.2. The Initial B-Vitamin Deficiency Provokes Persistent Cognitive Deficits Corrected by the Interventional Stimulation of Neurogenesis
3.3. Various Tissue Parameters Attested of a Rescue Mechanism
4. Methods
4.1. Animal Treatments
4.2. Exposure to Hypoxia
4.3. Biochemical Analyses
4.4. Behavioral Test
4.5. Histopathological Measurements
4.6. Quantification of Proteins
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kellogg, R.; Lee, P.; Deibert, C.P.; Tempel, Z.; Zwagerman, N.T.; Bonfield, C.M.; Johnson, S.; Greene, S. Twenty years’ experience with myelomeningocele management at a single institution: Lessons learned. J. Neurosurg. Pediatr. 2018, 224, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Deora, H.; Srinivas, D.; Shukla, D.; Devi, B.I.; Mishra, A.; Beniwal, M.; Kannepalli, N.R.; Somanna, S. Multiple-site neural tube defects: Embryogenesis with complete review of existing literature. Neurosurg Focus 2019, 47, E18. [Google Scholar] [CrossRef] [PubMed]
- Fofou-Caillierez, M.B.; Guéant-Rodriguez, R.M.; Alberto, J.M.; Chéry, C.; Josse, T.; Gérard, P.; Forges, T.; Foliguet, B.; Feillet, F.; Guéant, J.L. Vitamin B-12 and liver activity and expression of methionine synthase are decreased in fetuses with neural tube defects. Am. J. Clin. Nutr. 2019, 1096, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Atta, C.A.; Fiest, K.M.; Frolkis, A.D.; Jette, N.; Pringsheim, T.; St Germaine-Smith, C.; Rajapakse, T.; Kaplan, G.G.; Metcalfe, A. Global Birth Prevalence of Spina Bifida by Folic Acid Fortification Status: A Systematic Review and Meta-Analysis. Am. J. Public Health 2016, 106, e24–e34. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Gleeson, J.G. Closing in on Mechanisms of Open Neural Tube Defects. Trends Neurosci. 2020, 435, 519–532. [Google Scholar] [CrossRef]
- Huemer, M.; Simma, B.; Fowler, B.; Suormala, T.; Bodamer, O.A.; Sass, J.O. Prenatal and postnatal treatment in cobalamin C defect. J. Pediatr. 2005, 1474, 469–472. [Google Scholar] [CrossRef]
- Huemer, M.; Diodato, D.; Schwahn, B.; Schiff, M.; Bandeira, A.; Benoist, J.F.; Burlina, A.; Cerone, R.; Couce, M.L.; Garcia-Cazorla, A.; et al. Guidelines for diagnosis and management of the cobalamin-related remethylation disorders cblC, cblD, cblE, cblF, cblG, cblJ and MTHFR deficiency. J. Inherit. Metab. Dis. 2017, 402, 21–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, S.; Huemer, M.; Baumgartner, M.; Deodato, F.; Ballhausen, D.; Boneh, A.; Burlina, A.B.; Cerone, R.; Garcia, P.; Gökçay, G.; et al. Clinical presentation and outcome in a series of 88 patients with the cblC defect. J. Inherit. Metab. Dis. 2014, 378, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Copp, A.J.; Stanier, P.; Greene, N.D. Neural tube defects: Recent advances, unsolved questions, and controversies. Lancet Neurol. 2013, 127, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Berrocal-Zaragoza, M.I.; Sequeira, J.M.; Murphy, M.M.; Fernandez-Ballart, J.D.; Abdel Baki, S.G.; Bergold, P.J.; Quadros, E.V. Folate deficiency in rat pups during weaning causes learning and memory deficits. Br. J. Nutr. 2014, 1121, 1323–1332. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Cahill, C.M.; Huang, X.; Roffman, J.L.; Lamon-Fava, S.; Fava, M.; Mischoulon, D.; Rogers, J.T. S-Adenosyl Methionine and Transmethylation Pathways in Neuropsychiatric Diseases Throughout Life. Neurotherapeutics 2018, 151, 156–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J. The fetal origins of adult hypertension. J. Hypertens. 1992, 10, S39–S44. [Google Scholar] [CrossRef]
- Barker, D.J. The developmental origins of adult disease. J. Am. Coll. Nutr. 2004, 235, 588S–595S. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Rogers, J.M. Embryonic and fetal programming of physiological disorders in adulthood. Birth Defects Res. C Embryo Today 2004, 723, 300–312. [Google Scholar] [CrossRef]
- Bruce, K.D.; Hanson, M.A. The developmental origins, mechanisms, and implications of metabolic syndrome. J. Nutr. 2010, 1406, 48–652. [Google Scholar] [CrossRef] [Green Version]
- Blaise, S.A.; Alberto, J.M.; Audonnet-Blaise, S.; Guéant, J.L.; Daval, J.L. Gestational vitamin B deficiency leads to homocysteine-associated brain apoptosis and alters neurobehavioral development in rats. Am. J. Pathol. 2007, 170, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Battaglia-Hsu, S.-F.; Akchiche, N.; Noel, N.; Alberto, J.-M.; Jeannesson, E.; Orozco-Barrios, C.E.; Martinez-Fong, D.; Daval, J.-L.; Guéant, J.-L. Vitamin B12 deficiency reduces proliferation and promotes differentiation of neuroblastoma cells and up-regulates PP2A, proNGF, and TACE. Proc. Natl. Acad. Sci. USA 2009, 1062, 1930–21935. [Google Scholar] [CrossRef] [Green Version]
- Daval, J.L.; Blaise, S.; Guéant, J.L. Vitamin B deficiency causes neural cell loss and cognitive impairment in the developing rat. Proc. Natl. Acad. Sci. USA 2009, 106, E1. [Google Scholar] [CrossRef] [Green Version]
- Sable, P.; Kale, A.; Joshi, A.; Joshi, S. Maternal micronutrient imbalance alters gene expression of BDNF, NGF, TrkB and CREB in the offspring brain at an adult age. Int. J. Dev. Neurosci. 2014, 342, 4–32. [Google Scholar] [CrossRef] [PubMed]
- Crouzier, L.; Couly, S.; Roques, C.; Peter, C.; Belkhiter, R.; Arguel Jacquemin, M.; Bonetto, A.; Delprat, B.; Maurice, T. Sigma-1 (σ(1)) receptor activity is necessary for physiological brain plasticity in mice. Eur. Neuropsychopharmacol. 2020, 39, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Niklison-Chirou, M.V.; Agostini, M.; Amelio, I.; Melino, G. Regulation of adult neurogenesis in mammalian brain. Int. J. Mol. Sci. 2020, 21, 4869. [Google Scholar] [CrossRef]
- Mahalakshmi, B.; Maurya, N.; Lee, S.D.; Bharath Kumar, V. Possible neuroprotective mechanisms of physical exercise in neurodegeneration. Int. J. Mol. Sci. 2020, 215, 895. [Google Scholar]
- Blaise, S.A.; Nédélec, E.; Alberto, J.M.; Schroeder, H.; Audonnet, S.; Bossenmeyer-Pourié, C.; Guéant, J.L.; Daval, J.L. Short hypoxia could attenuate the adverse effects of hyperhomocysteinemia on the developing rat brain by inducing neurogenesis. Exp. Neurol. 2009, 2162, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.; Pourié, G.; Bossenmeyer-Pourié, C.; Jazi, R.; Guéant, J.L.; Vert, P.; Daval, J.L. Conditioning-like brief neonatal hypoxia improves cognitive function and brain tissue properties with marked gender dimorphism in adult rats. Semin. Perinatol. 2010, 341, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.; Bossenmeyer-Pourié, C.; Koziel, V.; Jazi, R.; Audonnet, S.; Vert, P.; Guéant, J.L.; Daval, J.L.; Pourié, G. Non-injurious neonatal hypoxia confers resistance to brain senescence in aged male rats. PLoS ONE 2012, 7, e48828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Fu, J.; Qu, Z.; Li, D.; Si, P.; Qiao, Q.; Zhang, W.; Xue, Y.; Zhen, J.; Wang, W. Chronic mild hypoxia promotes hippocampal neurogenesis involving Notch1 signaling in epileptic rats. Brain Res. 2019, 17148, 88–98. [Google Scholar] [CrossRef]
- Wakhloo, D.; Scharkowski, F.; Curto, Y.; Javed Butt, U.; Bansal, V.; Steixner-Kumar, A.A.; Wüstefeld, L.; Rajput, A.; Arinrad, S.; Zillmann, M.R.; et al. Functional hypoxia drives neuroplasticity and neurogenesis via brain erythropoietin. Nat. Commun. 2020, 111, 313. [Google Scholar] [CrossRef] [Green Version]
- Geoffroy, A.; Saber-Cherif, L.; Pourié, G.; Helle, D.; Umoret, R.; Guéant, J.L.; Bossenmeyer-Pourié, C.; Daval, J.L. Developmental Impairments in a Rat Model of Methyl Donor Deficiency: Effects of a Late Maternal Supplementation with Folic Acid. Int. J. Mol. Sci. 2019, 20, 973. [Google Scholar] [CrossRef] [Green Version]
- Malouf, M.; Grimley, E.J.; Areosa, S.A. Folic acid with or without vitamin B12 for cognition and dementia. Cochrane Database Syst. Rev. 2003, CD004514. [Google Scholar] [CrossRef]
- Miller, J.W. Assessing the association between vitamin B-12 status and cognitive function in older adults. Am. J. Clin. Nutr. 2006, 841, 259–1260. [Google Scholar] [CrossRef] [Green Version]
- Clarke, R. Vitamin B12, folic acid, and the prevention of dementia. N. Engl. J. Med. 2006, 3542, 817–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoffroy, A.; Kerek, R.; Pourié, G.; Helle, D.; Guéant, J.L.; Daval, J.L.; Bossenmeyer-Pourié, C. Late Maternal Folate Supplementation Rescues from Methyl Donor Deficiency-Associated Brain Defects by Restoring Let-7 and miR-34 Pathways. Mol. Neurobiol. 2017, 545, 017–5033. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, Z.; Li, S.; Wang, X.; Wilson, J.X.; Huang, G. Periconceptional Folic Acid Supplementation Benefit to Development of Early Sensory-Motor Function through Increase DNA Methylation in Rat Offspring. Nutrients 2018, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- McCarty, M.F.; O’Keefe, J.H.; DiNicolantonio, J.J. A diet rich in taurine, cysteine, folate, B(12) and betaine may lessen risk for Alzheimer’s disease by boosting brain synthesis of hydrogen sulfide. Med. Hypotheses 2019, 1321, 09356. [Google Scholar]
- Deniz, B.F.; Confortim, H.D.; Deckmann, I.; Miguel, P.M.; Bronauth, L.; de Oliveira, B.C.; Barbosa, S.; Cechinel, L.R.; Siqueira, I.R.; Pereira, L.O. Folic acid supplementation during pregnancy prevents cognitive impairments and BDNF imbalance in the hippocampus of the offspring after neonatal hypoxia-ischemia. J. Nutr. Biochem. 2018, 603, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Huot, P.S.; Ly, A.; Szeto, I.M.; Reza-López, S.A.; Cho, D.; Kim, Y.I.; Anderson, G.H. Maternal and postweaning folic acid supplementation interact to influence body weight, insulin resistance, and food intake regulatory gene expression in rat offspring in a sex-specific manner. Appl. Physiol. Nutr. Metab. 2016, 414, 411–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nohara, K.; Baba, T.; Murai, H.; Kobayashi, Y.; Suzuki, T.; Tateishi, Y.; Matsumoto, M.; Nishimura, N.; Sano, T. Global DNA methylation in the mouse liver is affected by methyl deficiency and arsenic in a sex-dependent manner. Arch. Toxicol. 2011, 856, 653–661. [Google Scholar] [CrossRef]
- Pourié, G.; Martin, N.; Bossenmeyer-Pourié, C.; Akchiche, N.; Guéant-Rodriguez, R.M.; Geoffroy, A.; Jeannesson, E.; El Hajj Chehadeh, S.; Mimoun, K.; Brachet, P.; et al. Folate- and vitamin B12-deficient diet during gestation and lactation alters cerebellar synapsin expression via impaired influence of estrogen nuclear receptor α. FASEB J. 2015, 293, 3713–3725. [Google Scholar] [CrossRef]
- Paternain, L.; Martisova, E.; Campión, J.; Martínez, J.A.; Ramírez, M.J.; Milagro, F.I. Methyl donor supplementation in rats reverses the deleterious effect of maternal separation on depression-like behaviour. Behav. Brain Res. 2016, 2995, 51–58. [Google Scholar] [CrossRef]
- Canever, L.; Freire, T.G.; Mastella, G.A.; Damázio, L.; Gomes, S.; Fachim, I.; Michels, C.; Carvalho, G.; Godói, A.K.; Peterle, B.R.; et al. Changes in behavioural parameters, oxidative stress and neurotrophins in the brain of adult offspring induced to an animal model of schizophrenia: The effects of FA deficient or FA supplemented diet during the neurodevelopmental phase. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 865, 52–64. [Google Scholar] [CrossRef]
- Smith, A.D.; Refsum, H.; Bottiglieri, T.; Fenech, M.; Hooshmand, B.; McCaddon, A.; Miller, J.W.; Rosenberg, I.H.; Obeid, R. Homocysteine and Dementia: An International Consensus Statement. J. Alzheimers Dis. 2018, 625, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauvais-Jarvis, F.; Bairey Merz, N.; Barnes, P.J.; Brinton, R.D.; Carrero, J.J.; DeMeo, D.L.; De Vries, G.J.; Epperson, C.N.; Govindan, R.; Klein, S.L.; et al. Sex and gender: Modifiers of health, disease, and medicine. Lancet 2020, 3965, 565–582. [Google Scholar] [CrossRef]
- Talboom, J.S.; West, S.G.; Engler-Chiurazzi, E.B.; Enders, C.K.; Crain, I.; Bimonte-Nelson, H.A. Learning to remember: Cognitive training-induced attenuation of age-related memory decline depends on sex and cognitive demand, and can transfer to untrained cognitive domains. Neurobiol. Aging 2014, 352, 2791–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalska, M.; Hnilicova, P.; Kalenska, D.; Tomascova, A.; Adamkov, M.; Lehotsky, J. Effect of Methionine Diet on Time-Related Metabolic and Histopathological Changes of Rat Hippocampus in the Model of Global Brain Ischemia. Biomolecules 2020, 101, 128. [Google Scholar]
- Gatta, E.; Mairesse, J.; Deruyter, L.; Marrocco, J.; Van Camp, G.; Bouwalerh, H.; Lo Guidice, J.M.; Morley-Fletcher, S.; Nicoletti, F.; Maccari, S. Reduced maternal behavior caused by gestational stress is predictive of life span changes in risk-taking behavior and gene expression due to altering of the stress/anti-stress balance. Neurotoxicology 2018, 661, 138–149. [Google Scholar] [CrossRef]
- Hassan, Z.; Coelho, D.; Kokten, T.; Alberto, J.M.; Umoret, R.; Daval, J.L.; Guéant, J.L.; Bossenmeyer-Pourié, C.; Pourié, G. Brain Susceptibility to Methyl Donor Deficiency: From Fetal Programming to Aging Outcome in Rats. Int. J. Mol. Sci. 2019, 20, 5692. [Google Scholar] [CrossRef] [Green Version]
- Morello, M.; Landel, V.; Lacassagne, E.; Baranger, K.; Annweiler, C.; Féron, F.; Millet, P. Vitamin D Improves Neurogenesis and Cognition in a Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2018, 556, 6463–6479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, W.; Li, Z.; Ma, Y.; Yan, J.; Wilson, J.X.; Huang, G. Maternal Folic Acid Supplementation During Pregnancy Promotes Neurogenesis and Synaptogenesis in Neonatal Rat Offspring. Cereb. Cortex 2019, 293, 3390–3397. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, J.; Feng, S.; Huang, C.; Liu, Z.; Sun, Y.E.; Liu, H. Metformin improves cognition of aged mice by promoting cerebral angiogenesis and neurogenesis. Aging 2020, 12, 17845–17862. [Google Scholar] [CrossRef]
- Qin, Y.; An, D.; Xu, W.; Qi, X.; Wang, X.; Chen, L.; Chen, L.; Sha, S. Estradiol Replacement at the Critical Period Protects Hippocampal Neural Stem Cells to Improve Cognition in APP/PS1 Mice. Front. Aging Neurosci. 2020, 122, 40. [Google Scholar]
- Fan, C.; Fu, H.; Dong, H.; Lu, Y.; Lu, Y.; Qi, K. Maternal n-3 polyunsaturated fatty acid deprivation during pregnancy and lactation affects neurogenesis and apoptosis in adult offspring: Associated with DNA methylation of brain-derived neurotrophic factor transcripts. Nutr. Res. 2016, 361, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, Z.; He, R.; Li, H.; Ding, S. GLAST-CreERT2 mediated deletion of GDNF increases brain damage and exacerbates long-term stroke outcomes after focal ischemic stroke in mouse model. Glia 2020, 682, 2395–2414. [Google Scholar] [CrossRef] [PubMed]
- Li, W.J.; Mao, F.X.; Chen, H.J.; Qian, L.H.; Buzby, J.S. Treatment with UDP-glucose, GDNF, and memantine promotes SVZ and white matter self-repair by endogenous glial progenitor cells in neonatal rats with ischemic PVL. Neuroscience 2015, 2844, 444–458. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pourié, G.; Martin, N.; Daval, J.-L.; Alberto, J.-M.; Umoret, R.; Guéant, J.-L.; Bossenmeyer-Pourié, C. The Stimulation of Neurogenesis Improves the Cognitive Status of Aging Rats Subjected to Gestational and Perinatal Deficiency of B9–12 Vitamins. Int. J. Mol. Sci. 2020, 21, 8008. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218008
Pourié G, Martin N, Daval J-L, Alberto J-M, Umoret R, Guéant J-L, Bossenmeyer-Pourié C. The Stimulation of Neurogenesis Improves the Cognitive Status of Aging Rats Subjected to Gestational and Perinatal Deficiency of B9–12 Vitamins. International Journal of Molecular Sciences. 2020; 21(21):8008. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218008
Chicago/Turabian StylePourié, Grégory, Nicolas Martin, Jean-Luc Daval, Jean-Marc Alberto, Rémy Umoret, Jean-Louis Guéant, and Carine Bossenmeyer-Pourié. 2020. "The Stimulation of Neurogenesis Improves the Cognitive Status of Aging Rats Subjected to Gestational and Perinatal Deficiency of B9–12 Vitamins" International Journal of Molecular Sciences 21, no. 21: 8008. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218008