Inhibiting P2Y12 in Macrophages Induces Endoplasmic Reticulum Stress and Promotes an Anti-Tumoral Phenotype


Abstract
:1. Introduction
2. Results
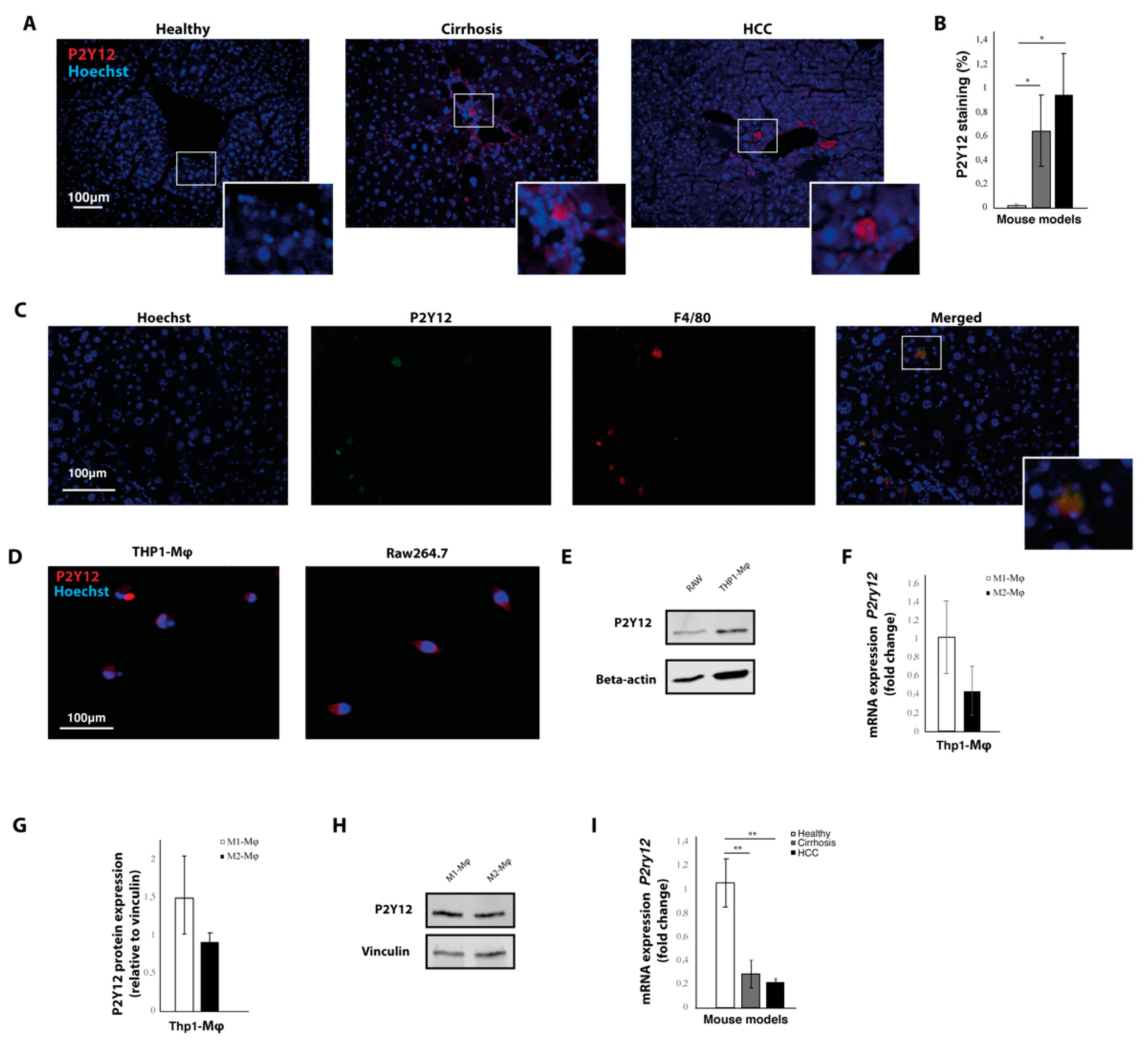
2.1. The P2Y12 Receptor Is Expressed in Mouse Models for Liver Cirrhosis and Hepatocellular Carcinoma
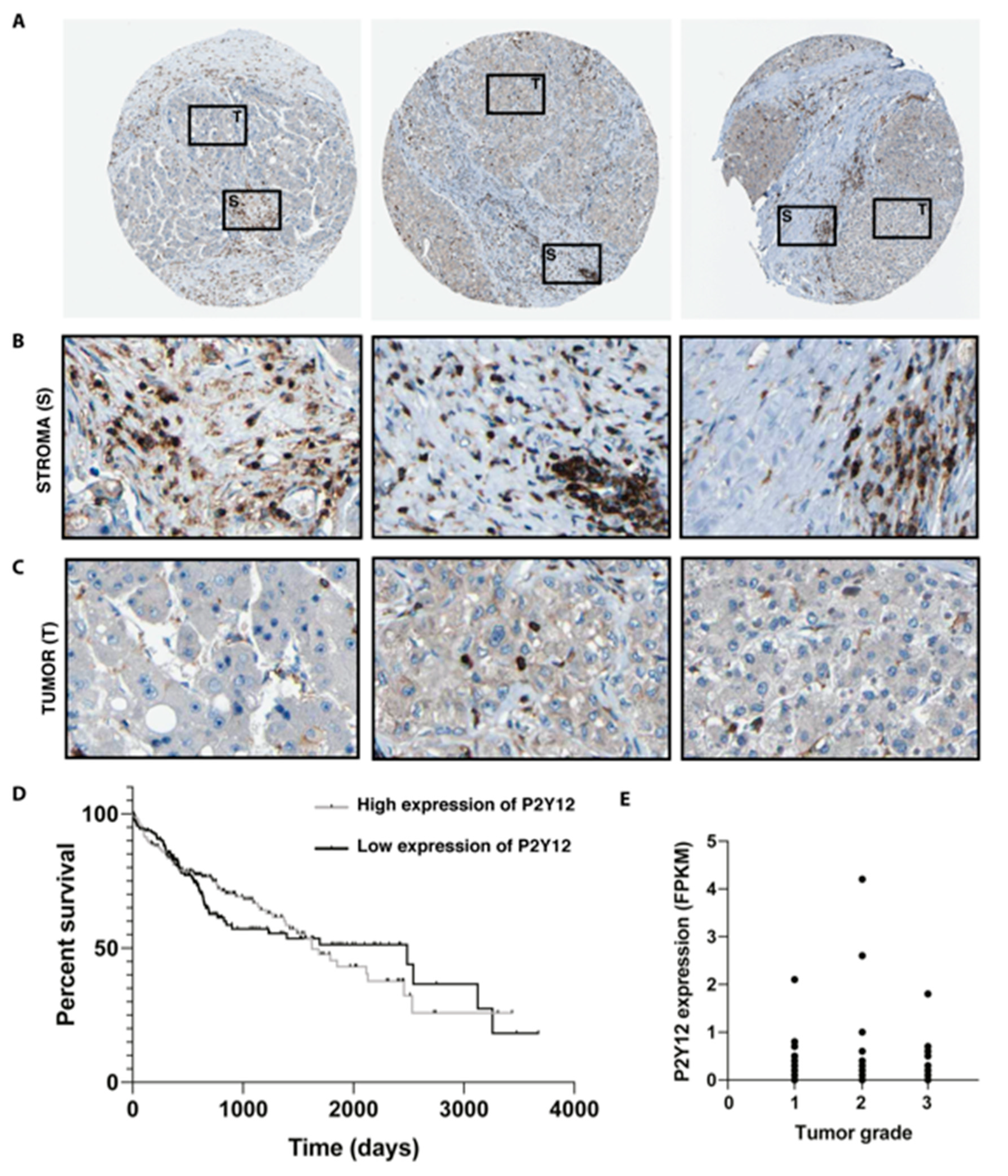
2.2. Expression of P2Y12 Is Located in the Stroma of Patients with Hepatocellular Carcinoma
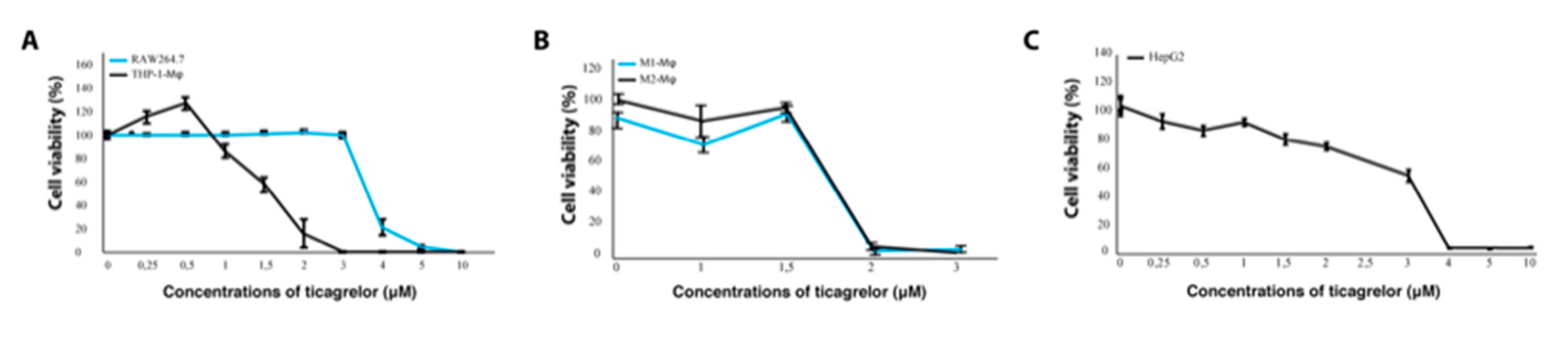
2.3. Inhibition of the P2Y12 Receptor Decreases Viability of Macrophages
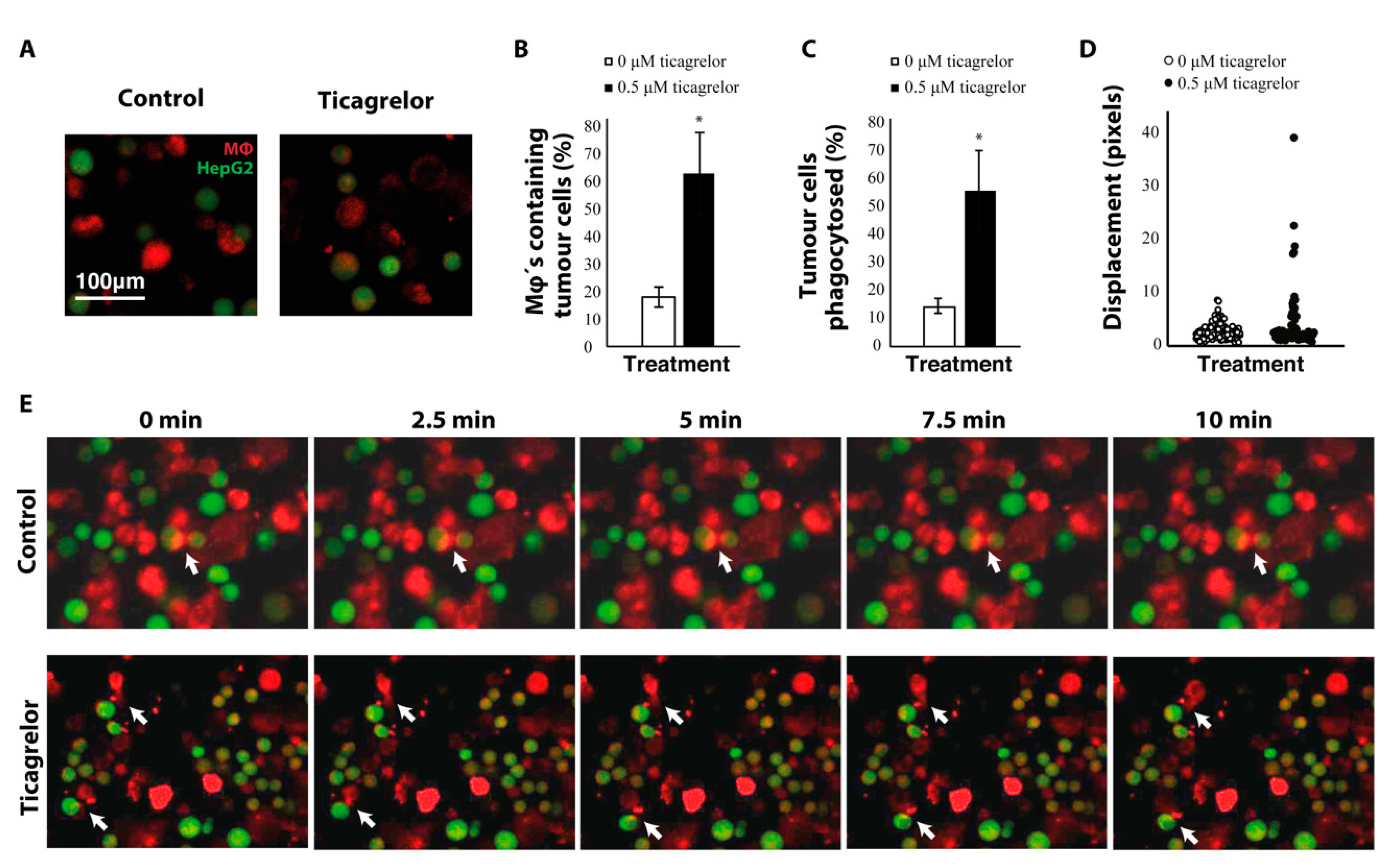
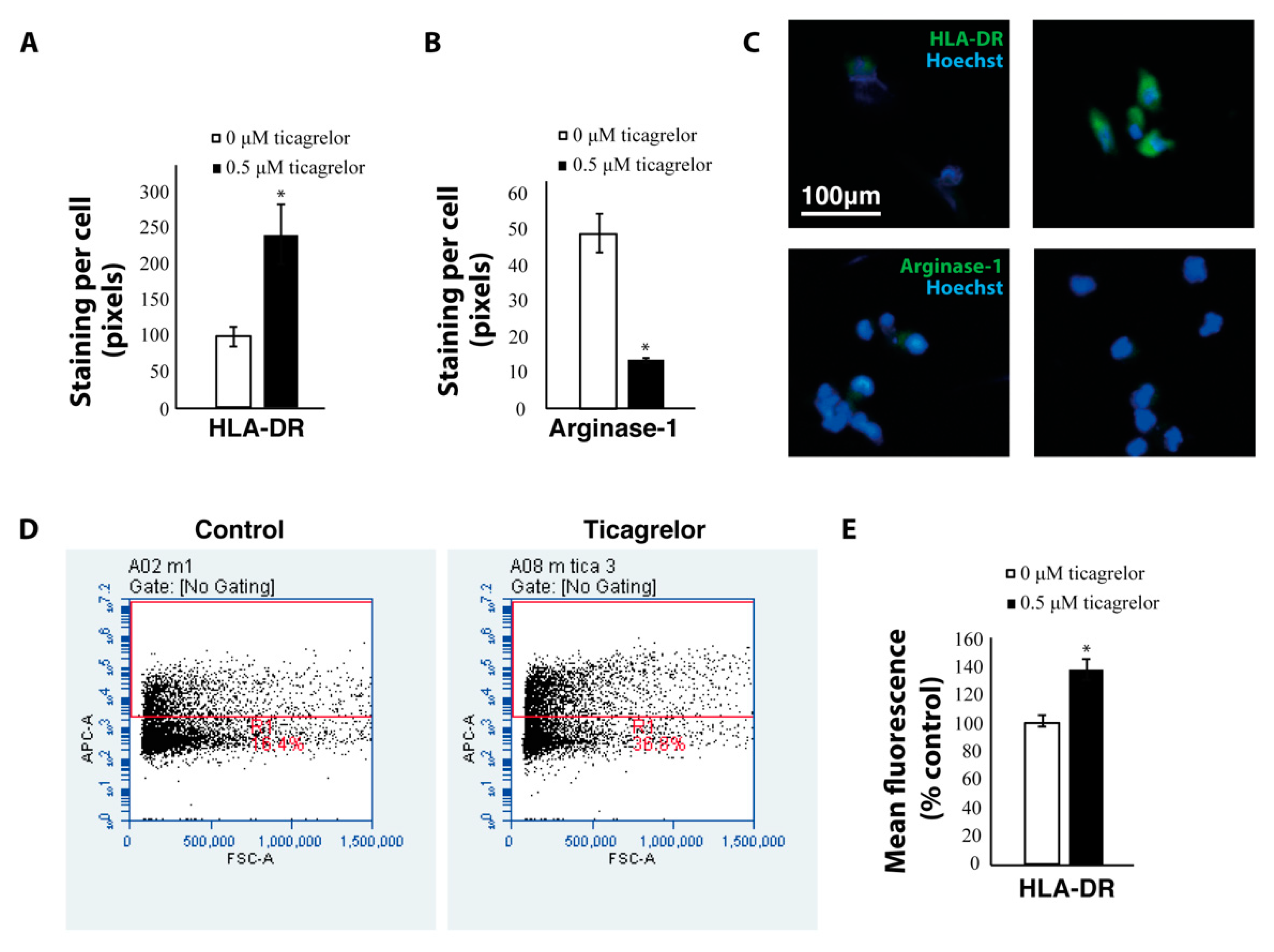
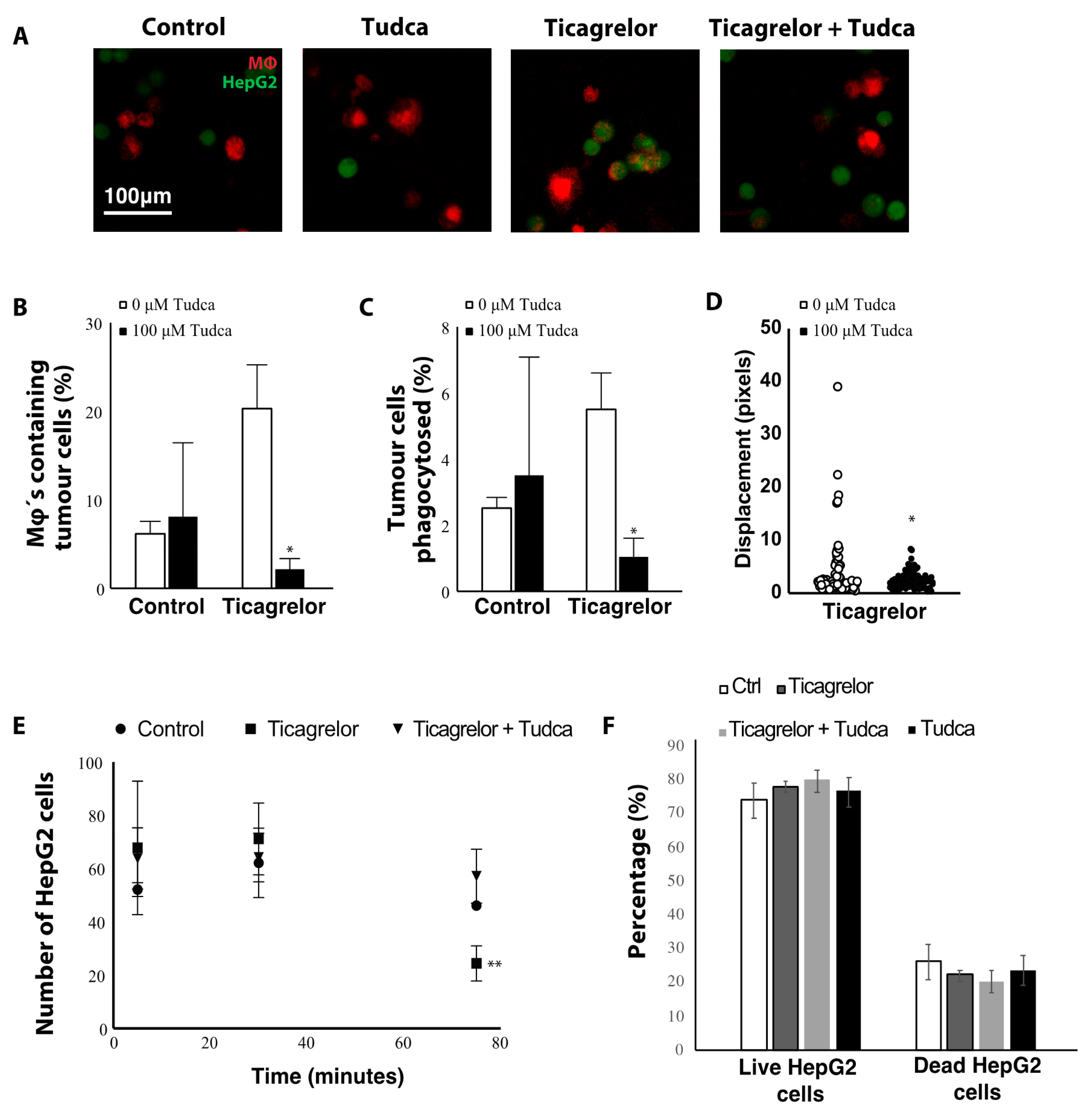
2.4. Ticagrelor Enhances Phagocytosis of Tumor Cells
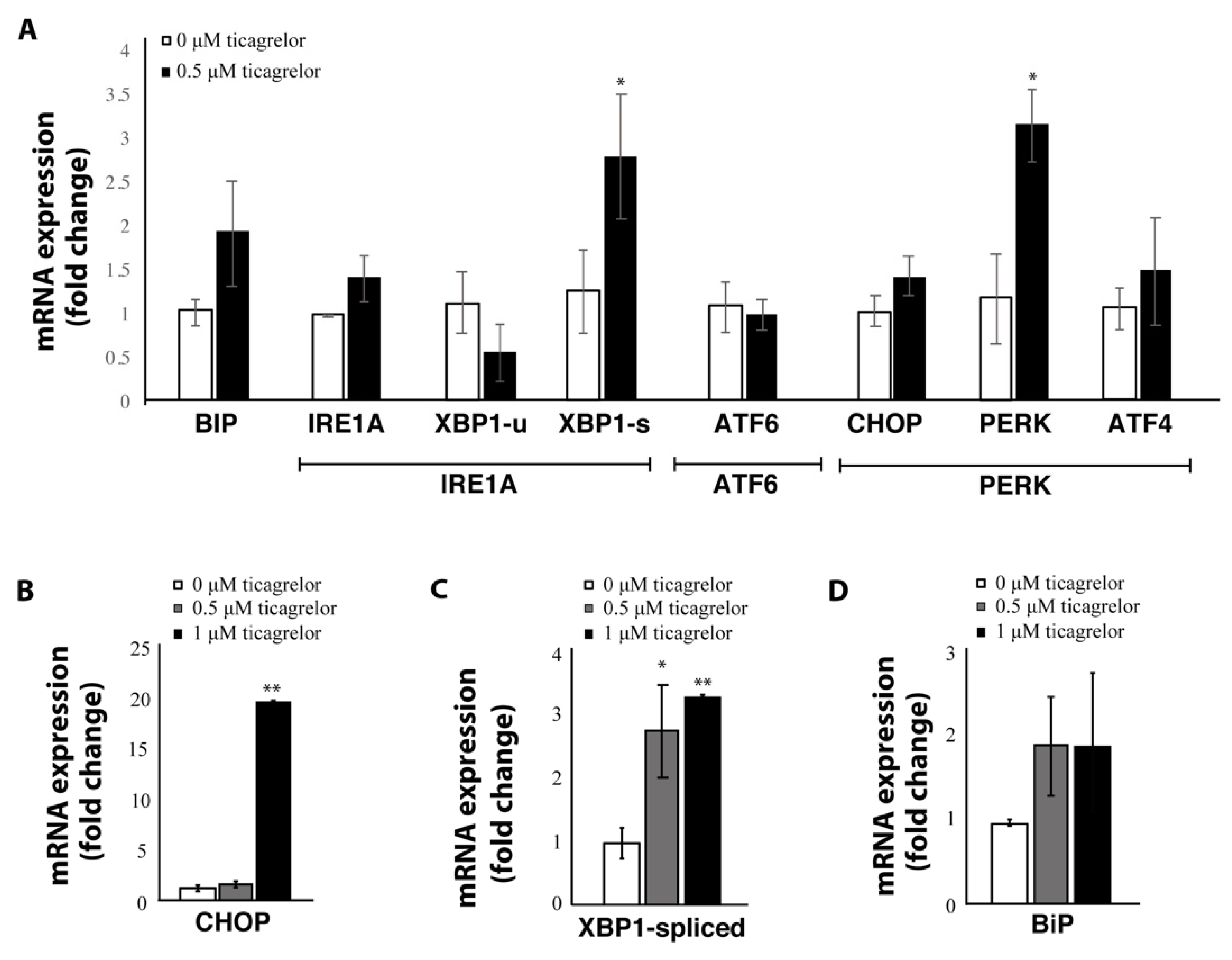
2.5. Expression of ER Stress Genes Is Increased after P2Y12 Inhibition with Ticagrelor
3. Discussion
4. Materials and Methods
4.1. Animal Models
4.2. Cell Culture and Reagents
4.3. Differentiation of THP1 Cells
4.4. Proliferation
4.5. Phagocytosis Assay
4.6. Immunohistochemistry
4.7. SDS-PAGE and Western Blot
4.8. Flow Cytometry
4.9. RT-qPCR
4.10. Human Protein Atlas
4.11. Live/Dead Cell Double Staining Kit
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADP | Adenosine diphosphate |
| ATF6 | Activating transcription factor 6 |
| ATP | Adenosine triphosphate |
| BIP | Binding immunoglobulin protein |
| CCL4 | Carbon tetrachloride |
| DEN | Diethylnitrosamine |
| DMEM | Dulbecco modified eagle medium |
| FPKM | Fragments Per Kilobase of exon per Million |
| HCC | Hepatocellular carcinoma |
| IRE1α | Inositol-requiring enzyme alpha |
| IL | Interleukin |
| MAPK | Mitogen-activated protein kinase |
| PBS | Phosphate buffered saline |
| PERK | Protein kinase R-like endoplasmic reticulum kinase |
| Tudca | Tauroursodeoxycholic acid |
| UPR | Unfolded protein response |
References
- Guillot, A.; Tacke, F. Liver Macrophages: Old Dogmas and New Insights. Hepatol. Commun. 2019, 3, 730–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillot, A.; Kohlhepp, M.S.; Bruneau, A.; Heymann, F.; Tacke, F. Deciphering the Immune Microenvironment on a Single Archival Formalin-Fixed Paraffin-Embedded Tissue Section by An Immediately Implementable Multiplex Fluorescence Immunostaining Protocol. Cancers 2020, 12, 2449. [Google Scholar] [CrossRef] [PubMed]
- Best, J.; Verhulst, S.; Syn, W.K.; Lagaisse, K.; van Hul, N.; Heindryckx, F.; Sowa, J.P.; Peeters, L.; Van Vlierberghe, H.; Leclercq, I.A.; et al. Macrophage Depletion Attenuates Extracellular Matrix Deposition and Ductular Reaction in a Mouse Model of Chronic Cholangiopathies. PLoS ONE 2016, 11, e0162286. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, M.; Rubis, B.; Frydrychowicz, M.; Nowicka, A.; Brajer-Luftmann, B.; Kozlowska, M.; Lagiedo, M.; Batura-Gabryel, H.; Sikora, J. Pleural Macrophages can Promote or Inhibit Apoptosis of Malignant Cells via Humoral Mediators Depending on Intracellular Signaling Pathways. Cancer Investig. 2018, 36, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kawaguchi, T.; Yan, L.; Peng, X.; Qi, Q.; Morris, L.G.T.; Chan, T.A.; Tsung, A.; Otsuji, E.; Takabe, K. Immune Cytolytic Activity for Comprehensive Understanding of Immune Landscape in Hepatocellular Carcinoma. Cancers 2020, 12, 1221. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, Z.B.; Liao, C.J.; Hu, X.W.; Li, S.L.; Qi, M.; Fan, X.G.; Huang, Y. LncRNA TP73-AS1/miR-539/MMP-8 axis modulates M2 macrophage polarization in hepatocellular carcinoma via TGF-beta1 signaling. Cell. Signal. 2020, 75, 109738. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Gao, W.; Tang, Q.; Yu, Y.; You, W.; Wu, Z.; Fan, Y.; Zhang, L.; Wu, C.; Han, G.; et al. M2 macrophage-derived exosomes facilitate hepatocarcinoma metastasis by transferring alphaM beta2 integrin to tumor cells. Hepatology 2020. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.A.; Rutkowski, D.T. Experimental reconstitution of chronic ER stress in the liver reveals feedback suppression of BiP mRNA expression. Elife 2016, 5. [Google Scholar] [CrossRef]
- Yang, F.; Liu, Y.; Ren, H.; Zhou, G.; Yuan, X.; Shi, X. ER-stress regulates macrophage polarization through pancreatic EIF-2alpha kinase. Cell. Immunol. 2019, 336, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jiang, Y.; Chen, L.; Zheng, X.; Zhu, J.; Song, X.; Shi, J.; Li, Y.; He, W. Inhibition of the endoplasmic reticulum (ER) stress-associated IRE-1/XBP-1 pathway alleviates acute lung injury via modulation of macrophage activation. J. Thorac. Dis. 2020, 12, 284–295. [Google Scholar] [CrossRef]
- Van Campenhout, S.; Tilleman, L.; Lefere, S.; Vandierendonck, A.; Raevens, S.; Verhelst, X.; Geerts, A.; Van Nieuwerburgh, F.; Van Vlierberghe, H.; Devisscher, L. Myeloid-specific IRE1alpha deletion reduces tumour development in a diabetic, non-alcoholic steatohepatitis-induced hepatocellular carcinoma mouse model. Metabolism 2020, 107, 154220. [Google Scholar] [CrossRef]
- Heindryckx, F.; Binet, F.; Ponticos, M.; Rombouts, K.; Lau, J.; Kreuger, J.; Gerwins, P. Endoplasmic reticulum stress enhances fibrosis through IRE1alpha-mediated degradation of miR-150 and XBP-1 splicing. EMBO Mol. Med. 2016, 8, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Borkham-Kamphorst, E.; Steffen, B.T.; van de Leur, E.; Haas, U.; Weiskirchen, R. Portal myofibroblasts are sensitive to CCN-mediated endoplasmic reticulum stress-related apoptosis with potential to attenuate biliary fibrogenesis. Cell. Signal. 2018, 51, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, D.; Nakao, Y.; Mauer, A.S.; Thompson, J.M.; Sehrawat, T.S.; Liao, C.Y.; Krishnan, A.; Lucien, F.; Guo, Q.; Liu, M.; et al. IRE1A Stimulates Hepatocyte-derived Extracellular Vesicles That Promote Inflammation in Mice With Steatohepatitis. Gastroenterology 2020, 159, 1487–1503. [Google Scholar] [CrossRef]
- Pavlović, N.; Calitz, C.; Thanapirom, K.; Mazza, G.; Rombouts, K.; Gerwins, P.; Heindryckx, F. Inhibiting IRE1α-endonuclease activity decreases tumor burden in a mouse model for hepatocellular carcinoma. eLife 2020, 9, e55865. [Google Scholar] [CrossRef]
- O’Connor, S.; Montalescot, G.; Collet, J.P. The P2Y(12) receptor as a target of antithrombotic drugs. Purinergic Signal. 2011, 7, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Mansour, A.; Bachelot-Loza, C.; Nesseler, N.; Gaussem, P.; Gouin-Thibault, I. P2Y12 Inhibition beyond Thrombosis: Effects on Inflammation. Int. J. Mol. Sci. 2020, 21, 1391. [Google Scholar] [CrossRef] [Green Version]
- Zanin, R.F.; Braganhol, E.; Bergamin, L.S.; Campesato, L.F.; Filho, A.Z.; Moreira, J.C.; Morrone, F.B.; Sevigny, J.; Schetinger, M.R.; de Souza Wyse, A.T.; et al. Differential macrophage activation alters the expression profile of NTPDase and ecto-5’-nucleotidase. PLoS ONE 2012, 7, e31205. [Google Scholar] [CrossRef] [Green Version]
- Haynes, S.E.; Hollopeter, G.; Yang, G.; Kurpius, D.; Dailey, M.E.; Gan, W.B.; Julius, D. The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 2006, 9, 1512–1519. [Google Scholar] [CrossRef]
- Moore, C.S.; Ase, A.R.; Kinsara, A.; Rao, V.T.; Michell-Robinson, M.; Leong, S.Y.; Butovsky, O.; Ludwin, S.K.; Seguela, P.; Bar-Or, A.; et al. P2Y12 expression and function in alternatively activated human microglia. Neurol. Neuroimmunol. Neuroinflamm. 2015, 2, e80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrottmaier, W.C.; Kral, J.B.; Badrnya, S.; Assinger, A. Aspirin and P2Y12 Inhibitors in platelet-mediated activation of neutrophils and monocytes. Thromb. Haemost. 2015, 114, 478–489. [Google Scholar] [CrossRef]
- Van Kolen, K.; Gilany, K.; Moens, L.; Esmans, E.L.; Slegers, H. P2Y12 receptor signalling towards PKB proceeds through IGF-I receptor cross-talk and requires activation of Src, Pyk2 and Rap1. Cell. Signal. 2006, 18, 1169–1181. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xu, D.; Hou, K.; Gou, X.; Li, Y. The role of P2Y12 receptor inhibition in ischemic stroke on microglia, platelets and vascular smooth muscle cells. J. Thromb. Thrombolysis 2020, 50, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Rauch, B.H.; Rosenkranz, A.C.; Ermler, S.; Bohm, A.; Driessen, J.; Fischer, J.W.; Sugidachi, A.; Jakubowski, J.A.; Schror, K. Regulation of functionally active P2Y12 ADP receptors by thrombin in human smooth muscle cells and the presence of P2Y12 in carotid artery lesions. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2434–2442. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Proteinatlas Images. Available online: https://www.proteinatlas.org/ENSG00000169313-P2RY12/pathology/liver+cancer#img (accessed on 5 September 2020).
- Proteinatlas Data. Available online: https://www.proteinatlas.org/ENSG00000169313-P2RY12/pathology/liver+cancer (accessed on 5 September 2020).
- Mira-Pascual, L.; Tran, A.N.; Andersson, G.; Nareoja, T.; Lang, P. A Sub-Clone of RAW264.7-Cells Form Osteoclast-Like Cells Capable of Bone Resorption Faster than Parental RAW264.7 through Increased De Novo Expression and Nuclear Translocation of NFATc1. Int. J. Mol. Sci. 2020, 21, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Hu, H.; Tian, M.; Ding, C.; Yu, S. The C/EBP Homologous Protein (CHOP) Transcription Factor Functions in Endoplasmic Reticulum Stress-Induced Apoptosis and Microbial Infection. Front. Immunol. 2018, 9, 3083. [Google Scholar] [CrossRef] [Green Version]
- Malehmir, M.; Pfister, D.; Gallage, S.; Szydlowska, M.; Inverso, D.; Kotsiliti, E.; Leone, V.; Peiseler, M.; Surewaard, B.G.J.; Rath, D.; et al. Platelet GPIbalpha is a mediator and potential interventional target for NASH and subsequent liver cancer. Nat. Med. 2019, 25, 641–655. [Google Scholar] [CrossRef] [Green Version]
- Sitia, G.; Iannacone, M.; Guidotti, L.G. Anti-platelet therapy in the prevention of hepatitis B virus-associated hepatocellular carcinoma. J. Hepatol. 2013, 59, 1135–1138. [Google Scholar] [CrossRef] [Green Version]
- Tan-Shalaby, J.L. Reproducible complete remission of advanced hepatocellular carcinoma with sorafenib in combination with clopidogrel. BMJ Case Rep. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlovic, N.; Rani, B.; Gerwins, P.; Heindryckx, F. Platelets as Key Factors in Hepatocellular Carcinoma. Cancers 2019, 11, 1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amadio, S.; Parisi, C.; Montilli, C.; Carrubba, A.S.; Apolloni, S.; Volonte, C. P2Y(12) receptor on the verge of a neuroinflammatory breakdown. Mediat. Inflamm. 2014, 2014, 975849. [Google Scholar] [CrossRef] [Green Version]
- Irino, Y.; Nakamura, Y.; Inoue, K.; Kohsaka, S.; Ohsawa, K. Akt activation is involved in P2Y12 receptor-mediated chemotaxis of microglia. J. Neurosci. Res. 2008, 86, 1511–1519. [Google Scholar] [CrossRef]
- Chen, W.; Sandoval, H.; Kubiak, J.Z.; Li, X.C.; Ghobrial, R.M.; Kloc, M. The phenotype of peritoneal mouse macrophages depends on the mitochondria and ATP/ADP homeostasis. Cell. Immunol. 2018, 324, 1–7. [Google Scholar] [CrossRef]
- Suzuki, T.; Gao, J.; Ishigaki, Y.; Kondo, K.; Sawada, S.; Izumi, T.; Uno, K.; Kaneko, K.; Tsukita, S.; Takahashi, K.; et al. ER Stress Protein CHOP Mediates Insulin Resistance by Modulating Adipose Tissue Macrophage Polarity. Cell Rep. 2017, 18, 2045–2057. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.L.; Duan, Z.T.; Jiang, Z.D.; Cao, W.J.; Wang, Z.B.; Hu, K.W.; Gao, X.; Wang, S.K.; He, B.S.; Zhang, Z.Y.; et al. Increased endoplasmic reticulum stress response is involved in clopidogrel-induced apoptosis of gastric epithelial cells. PLoS ONE 2013, 8, e74381. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Yamanaka, H.; Fukuoka, T.; Dai, Y.; Obata, K.; Noguchi, K. P2Y12 receptor upregulation in activated microglia is a gateway of p38 signaling and neuropathic pain. J. Neurosci. 2008, 28, 2892–2902. [Google Scholar] [CrossRef]
- Chakraborty, P.; Chatterjee, S.; Ganguly, A.; Saha, P.; Adhikary, A.; Das, T.; Chatterjee, M.; Choudhuri, S.K. Reprogramming of TAM toward proimmunogenic type through regulation of MAP kinases using a redox-active copper chelate. J. Leukoc. Biol. 2012, 91, 609–619. [Google Scholar] [CrossRef]
- Cubillos-Ruiz, J.R.; Mohamed, E.; Rodriguez, P.C. Unfolding anti-tumor immunity: ER stress responses sculpt tolerogenic myeloid cells in cancer. J. Immunother. Cancer 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.; Riek, A.E.; Weng, S.; Petty, M.; Kim, D.; Colonna, M.; Cella, M.; Bernal-Mizrachi, C. Endoplasmic reticulum stress controls M2 macrophage differentiation and foam cell formation. J. Biol. Chem. 2012, 287, 11629–11641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Pantoja, D.R.; Wilson, A.S.; Clear, K.Y.; Westwood, B.; Triozzi, P.L.; Cook, K.L. Unfolded protein response signaling impacts macrophage polarity to modulate breast cancer cell clearance and melanoma immune checkpoint therapy responsiveness. Oncotarget 2017, 8, 80545–80559. [Google Scholar] [CrossRef]
- Husted, S.; van Giezen, J.J. Ticagrelor: The first reversibly binding oral P2Y12 receptor antagonist. Cardiovasc. Ther. 2009, 27, 259–274. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, K.; Lutz, D.A.; Strassburger, J.; Baqi, Y.; Muller, C.E.; von Kugelgen, I. Competitive mode and site of interaction of ticagrelor at the human platelet P2Y12 -receptor. J. Thromb. Haemost. 2014, 12, 1898–1905. [Google Scholar] [CrossRef] [PubMed]
- Maess, M.B.; Wittig, B.; Lorkowski, S. Highly efficient transfection of human THP-1 macrophages by nucleofection. J. Vis. Exp. 2014, e51960. [Google Scholar] [CrossRef] [Green Version]
- Sumaya, W.; Storey, R.F. Ticagrelor: Effects Beyond the P2Y12 Receptor. Interv. Cardiol. Clin. 2017, 6, 49–55. [Google Scholar]
- Armstrong, D.; Summers, C.; Ewart, L.; Nylander, S.; Sidaway, J.E.; van Giezen, J.J. Characterization of the adenosine pharmacology of ticagrelor reveals therapeutically relevant inhibition of equilibrative nucleoside transporter 1. J. Cardiovasc. Pharmacol. Ther. 2014, 19, 209–219. [Google Scholar] [CrossRef]
- Aungraheeta, R.; Conibear, A.; Butler, M.; Kelly, E.; Nylander, S.; Mumford, A.; Mundell, S.J. Inverse agonism at the P2Y12 receptor and ENT1 transporter blockade contribute to platelet inhibition by ticagrelor. Blood 2016, 128, 2717–2728. [Google Scholar] [CrossRef]
- Scholten, D.; Trebicka, J.; Liedtke, C.; Weiskirchen, R. The carbon tetrachloride model in mice. Lab. Anim. 2015, 49, 4–11. [Google Scholar] [CrossRef]
- Heindryckx, F.; Bogaerts, E.; Coulon, S.H.; Devlies, H.; Geerts, A.M.; Libbrecht, L.; Stassen, J.M.; Carmeliet, P.; Colle, I.O.; Van Vlierberghe, H.R. Inhibition of the placental growth factor decreases burden of cholangiocarcinoma and hepatocellular carcinoma in a transgenic mouse model. Eur. J. Gastroenterol. Hepatol. 2012, 24, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Heindryckx, F.; Mertens, K.; Charette, N.; Vandeghinste, B.; Casteleyn, C.; Van Steenkiste, C.; Slaets, D.; Libbrecht, L.; Staelens, S.; Starkel, P.; et al. Kinetics of angiogenic changes in a new mouse model for hepatocellular carcinoma. Mol. Cancer 2010, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Calitz, C.; Pavlović, N.; Rosenquist, J.; Zagami, C.; Samanta, A.; Heindryckx, F. A Biomimetic Model for Liver Cancer to Study Tumor-Stroma Interactions in a 3D Environment with Tunable Bio-Physical Properties. J. Vis. Exp. 2020. [Google Scholar] [CrossRef]
- Lin, R.C.; Yang, S.F.; Chiou, H.L.; Hsieh, S.C.; Wen, S.H.; Lu, K.H.; Hsieh, Y.H. Licochalcone A-Induced Apoptosis Through the Activation of p38MAPK Pathway Mediated Mitochondrial Pathways of Apoptosis in Human Osteosarcoma Cells In Vitro and In Vivo. Cells 2019, 8, 1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.R.; Park, H.B.; Kim, M.J.; Jung, B.D.; Lee, S.; Park, C.K.; Cheong, H.T. Effects of Endoplasmic Reticulum Stress Inhibitor Treatment during the Micromanipulation of Somatic Cell Nuclear Transfer in Porcine Oocytes. Dev. Reprod. 2019, 23, 43–54. [Google Scholar] [CrossRef]
- Tinevez, J.Y.; Perry, N.; Schindelin, J.; Hoopes, G.M.; Reynolds, G.D.; Laplantine, E.; Bednarek, S.Y.; Shorte, S.L.; Eliceiri, K.W. TrackMate: An open and extensible platform for single-particle tracking. Methods 2017, 115, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network; Wheeler, D.A.; Roberts, L.R. Comprehensive and Integrative Genomic Characterization of Hepatocellular Carcinoma. Cell 2017, 169, 1327–1341. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target (Species) | Forward | Reverse |
|---|---|---|
| P2ry12 (mouse) | ACGAGTTTGTGAAGGCACGA | AGACTCACGACTTTCTTCTTTGGA |
| P2ry12 (human) | TTTGCCCGAATTCCTTACAC | ATTGGGGCACTTCAGCATAC |
| 18S (human) | AGTCCCTGCCCTTTGTACACA | GATCCGAGGGCCTCACTAAAC |
| CHOP (human) | CATCACCACACCTGAAAGCA | TCAGCTGCCATCTCTGCA |
| XBP1-spliced (human) | AGACAGCGCTTGGGGATGGAT | CCTGCACCTGCTGCGGACTC |
| BIP (human) | GAAAGAAGGTTACCCATGCAGT | CAGGCCATAAGCAATAGCAGC |
| GAPDH (human) | GGAGCGAGATCCCTCCAAAAT | GGCTGTTGTCATACTTCTCATGG |
| GAPDH (mouse) | AATGAAGGGGTCGTTGATG | GGTGAAGGTCGGTGTGAAC |
| TBP1 (human) | AGTGACCCAGCATCACTGTTT | GGCAAACCAGAAACCCTTGC |
| ATF4 (human) | TCAGACAGTACCAACGCTTATGC | GTTGTACCACAGTAGGCTGAGA |
| ATF6 (human) | GTTGTACCACAGTAGGCTGAGA | TGGCCAAGCACTTCAAACCT |
| IRE1 (human) | TAGTCAGTTCTGCGTCCGCT | TTCCAAAAATCCCGAGGCCG |
| PERK (human) | ACGATGAGACAGAGTTGCGA | GCGCGTAAACAAGTTGCCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlović, N.; Kopsida, M.; Gerwins, P.; Heindryckx, F. Inhibiting P2Y12 in Macrophages Induces Endoplasmic Reticulum Stress and Promotes an Anti-Tumoral Phenotype. Int. J. Mol. Sci. 2020, 21, 8177. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218177
Pavlović N, Kopsida M, Gerwins P, Heindryckx F. Inhibiting P2Y12 in Macrophages Induces Endoplasmic Reticulum Stress and Promotes an Anti-Tumoral Phenotype. International Journal of Molecular Sciences. 2020; 21(21):8177. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218177
Chicago/Turabian StylePavlović, Nataša, Maria Kopsida, Pär Gerwins, and Femke Heindryckx. 2020. "Inhibiting P2Y12 in Macrophages Induces Endoplasmic Reticulum Stress and Promotes an Anti-Tumoral Phenotype" International Journal of Molecular Sciences 21, no. 21: 8177. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218177