Integrative Analysis of Multi-Omics Data Based on Blockwise Sparse Principal Components


1
Department of Preventive Medicine, Eulji University, Daejeon 34824, Korea
2
Department of Statistics, Korea University, Seoul 02841, Korea
3
Department of Statistics, Seoul National University, Seoul 08826, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(21), 8202; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218202
Submission received: 22 September 2020
/
Revised: 27 October 2020
/
Accepted: 31 October 2020
/
Published: 2 November 2020
(This article belongs to the Special Issue OMICs, Data Integration, and Applications in Personalized Medicine)
Abstract
:The recent development of high-throughput technology has allowed us to accumulate vast amounts of multi-omics data. Because even single omics data have a large number of variables, integrated analysis of multi-omics data suffers from problems such as computational instability and variable redundancy. Most multi-omics data analyses apply single supervised analysis, repeatedly, for dimensional reduction and variable selection. However, these approaches cannot avoid the problems of redundancy and collinearity of variables. In this study, we propose a novel approach using blockwise component analysis. This would solve the limitations of current methods by applying variable clustering and sparse principal component (sPC) analysis. Our approach consists of two stages. The first stage identifies homogeneous variable blocks, and then extracts sPCs, for each omics dataset. The second stage merges sPCs from each omics dataset, and then constructs a prediction model. We also propose a graphical method showing the results of sparse PCA and model fitting, simultaneously. We applied the proposed methodology to glioblastoma multiforme data from The Cancer Genome Atlas. The comparison with other existing approaches showed that our proposed methodology is more easily interpretable than other approaches, and has comparable predictive power, with a much smaller number of variables.
1. Introduction
Recent advances in high-throughput technologies have generated massive amounts of various types of biological data. Moreover, multiple “omics” data, such as genomics, epigenomics, transcriptomics, proteomics, and metabolomics, have been collected from various sources [1,2,3]. Analysis of these omics data sets may provide insight into mechanisms of disease, or the identification of new biomarkers [4]. However, because omics data is high-dimensional, there are many difficulties in using all the variables, to establish a statistical model for diagnostic or prognostic purposes. For example, computational instability, variable redundancy, and difficulties in interpretation can all occur. Moreover, integration of multiple omics data is also a challenge. In this paper, we propose an approach for constructing an efficient prediction model, for multi-omics data, considering three aspects: dimensionality reduction, variable selection, and data integration.
Dimension reduction plays a crucial role in data exploration, downstream pattern recognition, classification, and clustering for high-dimensional data that contains a large number of variables [4,5,6,7]. With regard to unsupervised approaches, principal component analysis (PCA) is a well-known dimensional reduction method, providing principal components (PCs) that are derived by linearly combining the original variables. The coefficients of the original variables are called “loadings” [8]. Instead of using individual variables, several PCs can be used as predictors for disease modeling.
However, each PC still needs all the original variables, since the coefficients are typically nonzero [9], and it is difficult to interpret the derived PCs. Therefore, it is desirable to select variables, as well as to reduce the dimensionality. An ad hoc approach is to ignore variables with small absolute values of loadings, when interpreting the PCs, or to artificially set the loading of variables, that are less than a certain threshold, to zero [10]. Though frequently used in practice, this approach is quite unreliable, since it does not take into account the variances, and the patterns of correlations of the variables [11]. The second possible approach is to use regularization penalties, to modify principal components, with sparse loading. Sparse principal component analysis (sPCA), which imposes elastic net penalties, can be used to overcome the drawback of PCA, and be formulated for variable selection [10]. To apply sPCA for ultra-high dimensional data, such as omics data, a screening procedure is still required to perform a penalized selection. One approach is to filter significant variables first, by repeating single supervised analysis. However, this type of variable selection can repeatedly select highly correlated variables, although the problems of variable redundancy and collinearity between variables, remain [12,13].
On the other hand, it is common to analyze single omics data sets, from a single study; however, to better understand the mechanisms for complex diseases, integration of the same types of omics data, across multiple studies, and/or integration of multi-omics data for the same samples, is desirable [2,14,15,16]. For horizontal integration of the same type of omics data, typical meta-analysis can be used for biomarker detection [17,18]. For vertical integration of multi-omics data, parallel integration, which treats each type of omics measurements equivalently, has been studied [19]. Usually, more attention is focused on vertical integration than horizontal integration. As for supervised integration, Zhao et al. [20] integrated four types of multi-omics datasets from The Cancer Genome Atlas (TCGA), including gene mRNA expression, DNA methylation, microRNA expression, and copy number alterations, for predicting cancer prognosis of invasive breast carcinoma, glioblastoma multiforme, acute myeloid leukemia, and lung squamous cell carcinoma. They then extracted important features by applying PCA, partial least squares (PLS), and least absolute shrinkage and selection operator (LASSO) methods, for individual genomic data sets, and then fitted them to a Cox model [20]. For the prediction of cutaneous melanoma prognosis, Jiang et al. [21] used elastic net, sparse PCA, and sparse PLS, to extract important variables from each type of omics dataset, and then integrated the important variables. Please note that in both studies, supervised screening for single genes was conducted, for each omics data set, in advance, to select a small number of genes. Therefore, they did not avoid the problems of redundancy and collinearity, of selected variables.
Various methodologies have been developed for dimensional reduction, variable selection, and integration of multi-omics data. Before conducting an integrative analysis of multi-omics data, variable selection in each single omics dataset is usually done first, whether it is explicitly reported or not [19]. Supervised techniques involving penalized variable selection such as LASSO, SCAD, and MCP can be applied [22,23,24,25]. Methods based on Bayesian approaches can also be applied for biological data [26]. In addition, some machine learning techniques, including the random forest and boosting methods, are applicable for variable selection in integrative analysis [27,28]. Unsupervised techniques, such as PCA and canonical correlation analysis, have also been considered. Several variations of PCA, such as consensus PCA (CPCA), multiple-block PCA (MBPCA), and nonnegative matrix factorization (NMF) have been developed [29,30,31]. By adding sparsity properties to these methods, sPCA, sparse CCA, and the joint and individual variation explained (JIVE) method were applied for genomic data [32,33,34]. There are various tools and methodologies for multi-omics data integration [16,19,35]. Wu et al. [19] classified integrated analyses into parallel and hierarchical integration. Parallel integration treats each omics dataset equally, whereas hierarchical integration incorporates the prior knowledge of regulatory relationships among each omics dataset. For example, iCluster, an integrative clustering method using a joint latent variable model, and its variants exemplify the parallel integration approach [36], while iBAG, an integrative Bayesian analysis of genomics data, is a hierarchical integration method [37]. Subramanian et al. [38] recently introduced a total of 31 analysis tools classified into similarity, correlation, network, Bayesian, fusion and multivariate approaches. Although many methodologies have been developed, each has its own advantages and disadvantages, and none of them dominates the others.
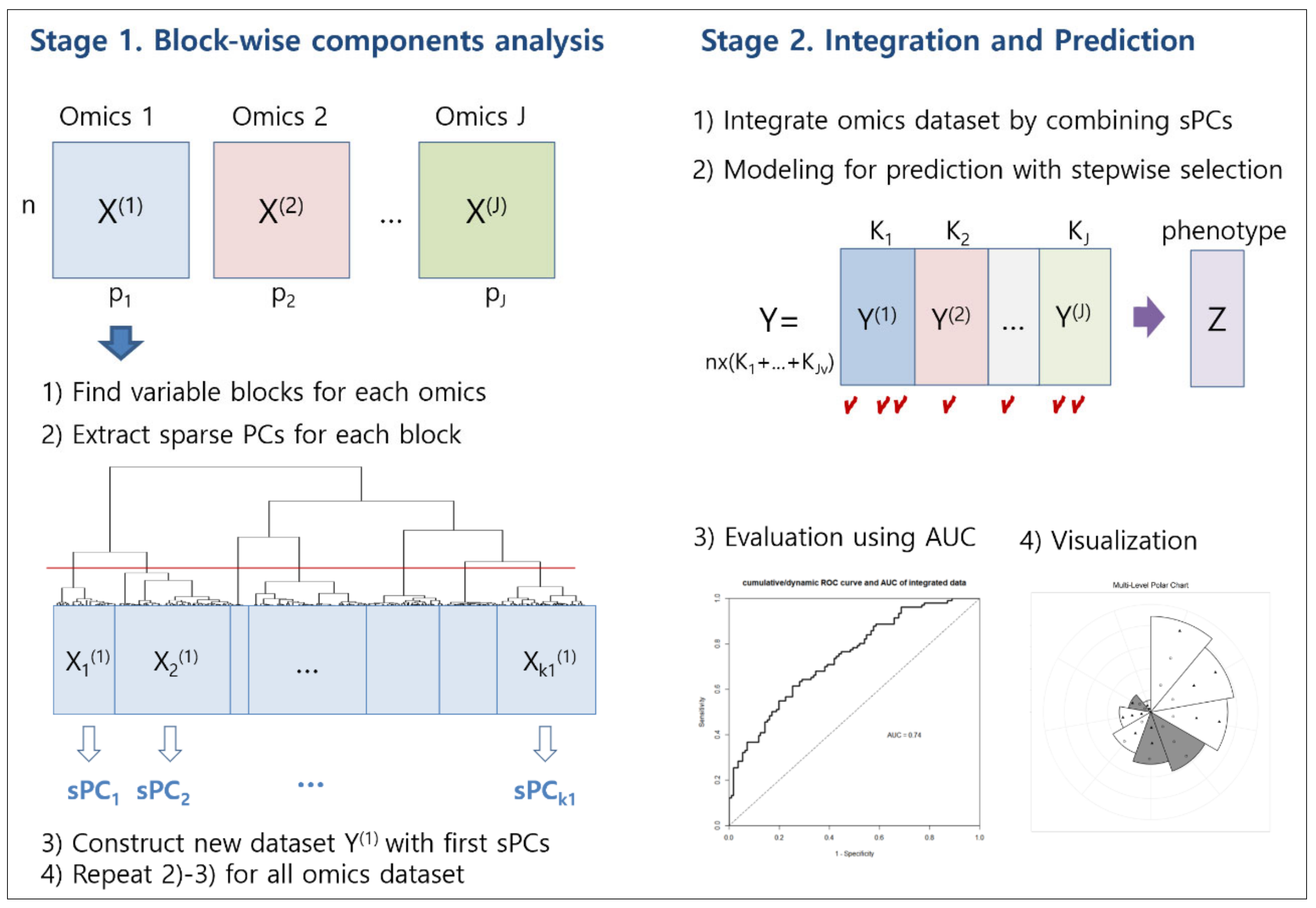
In this study, we propose an integrative analysis of multi-omics data that reduces multi-collinearity and redundancy, while reducing dimensionality and selecting variables of importance. It is based on blockwise sparse principal components. The proposed methodology is called iMO-BSPC (Integrative analysis of Multi-Omics data based on Block-wise sparse Principal Component Analysis). Unlike other multi-omics integration methods, our approach first performs variable clustering for each omics data. This process yields a set of closely related variables, and provides the same information contained in the initial variables. Herein, we call this a variable cluster “block”. For each block, we select the first sparse principal components (sPCs), as surrogate variables for dimensional reduction. Applying sPCA, instead of PCA, can reduce the number of variables. Then, sPCs from each omics data are merged to form an integrated multi-omics dataset. Finally, a prediction model using these sPCs as explanatory variables is constructed. We also suggest a new graphical method based on polar charts, that represents the results from sPCA and Cox regression. The proposed graph can simultaneously represent important sPCs for survival time prediction and the variables that make up the sPCs.
In Section 2, we apply the iMO-BSPC to glioblastoma multiforme (GBM) data sets from the TCGA. Since the response variable is survival time, with censoring, we perform Cox regression analysis for prognosis. Comparisons of the results from two alternative approaches for dimensional reduction are considered. We summarize and discuss the results in Section 3. The step-by-step procedure of the proposed iMO-BSPC approach is introduced in Section 4.
2. Results
We applied our framework to glioblastoma multiforme (GBM) data from The Cancer Genome Atlas (TCGA). The data set consists of 215 GBM patients, and three omics data: DNA methylation, with 1305 genes, mRNA expression with 12,042 genes, and miRNA expression, with 534 miRNAs. Among the 12,042 genes’ mRNA expression data, we sorted the genes in alphabetical order and selected the first gene for every 10 genes, i.e., 1205 genes in total. As a response variable, we used survival times, and their corresponding censoring statuses. We then used data downloaded from http://compbio.cs.toronto.edu/SNF/SNF/Software.html.
2.1. Analysis Using iMO-BSPC
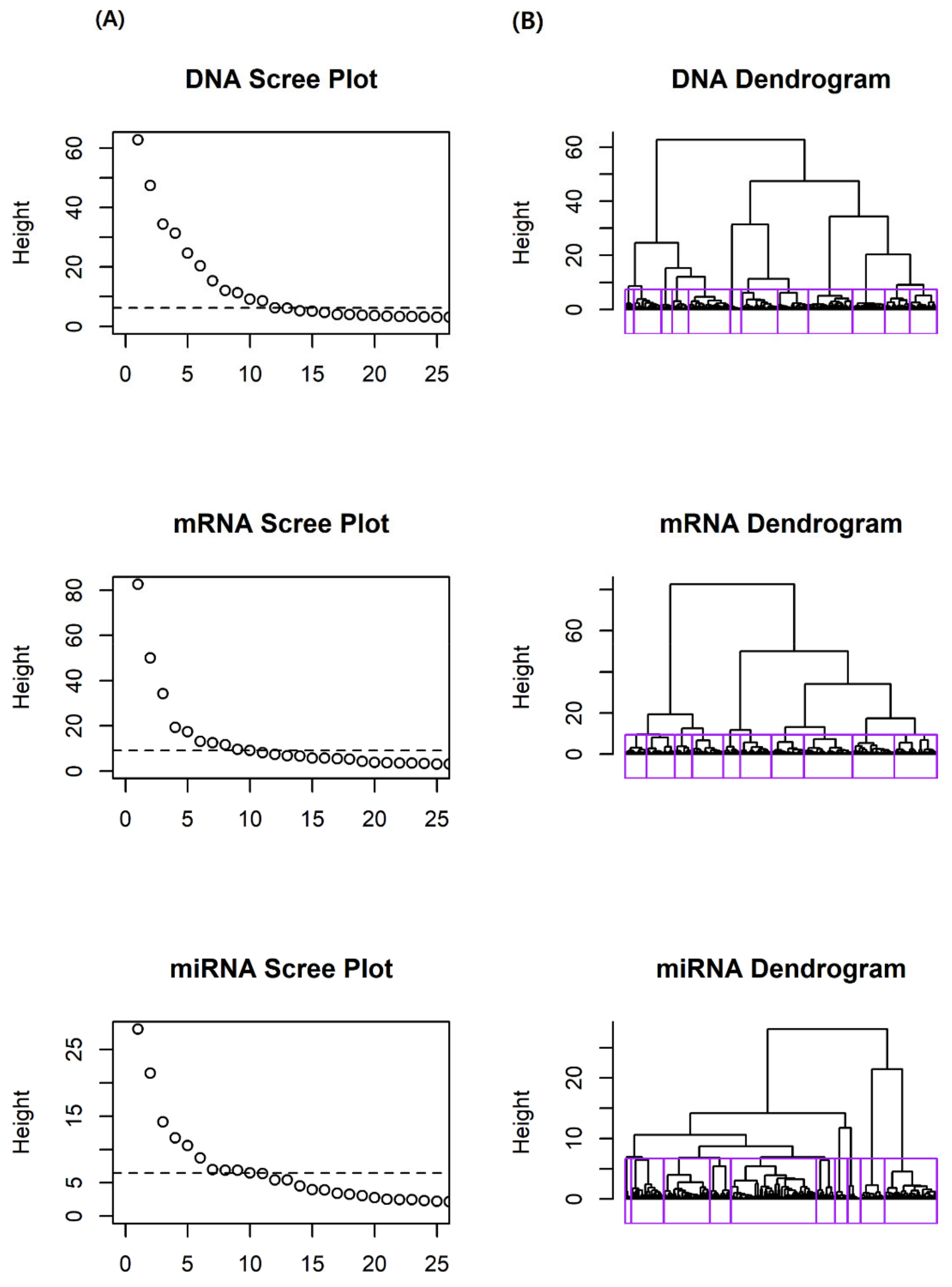
We first conducted variable clustering, using the hclustvar() function in the R package ‘ClustOfVar’(ver. 1.1). Based on the dendrograms and scree plots, of each omics data set, we determined 12, 10, and 10 blocks for DNA methylation, mRNA expression, and miRNA expression, respectively. Figure 1 shows the dendrograms and scree plots for each omics data set. In dendrograms, each block is depicted by a rectangle. As a result of variable clustering, we obtained 32 homogeneous variable blocks.
Next, sPCA was performed on each block. The first sPC was obtained from 32 blocks, resulting in a total of 32 sPCs. We used 10% of the number of variables as sparsity threshold. Thus, each sPC was made up of 10% of its original variables. This procedure was performed using the spca() function in the R package ‘elasticnet’. Table S1 shows the obtained sparse loadings. For convenience, only the results corresponding to the block selected in the next stage are listed. For example, the sPC in the fourth block, DNA methylation (D4), consisted of five genes of ALK_E183_R, KRAS_P651_F, IFNGR2_E164_F, RASA1_E107_F, and CCKBR_P361_R. Among them, ALK_E183_R showed the highest influence from sparse principal components, among the variables in the block. KRAS_P651_F, the variable with the second largest loading, represented the variable of DNA methylation block 4. All the selected variables from each block are listed in Table S2.
For the integrated dataset with 32 sPCs for 215 patients we used the Cox regression model. Stepwise variable selection based on Akaike information criterion (AIC) values was performed. Thirteen sPCs were finally selected that had significant effects on survival time. The AIC was 1731.66. Table 1 summarizes the selected sPCs, and their coefficients and hazard ratios (HRs). SPC of mRNA’s 9th block (MR9) showed the biggest absolute coefficient among 34 blocks, followed by the 2nd block of DNA methylation (D6), the 3rd block of miRNA (MI3), and so on. For example, sPC of mRNA’s 9th block (MR9) had a hazard ratio of 0.67, showing a significant effect on survival time. The larger the value of sPC of MR9, the lower the HR.
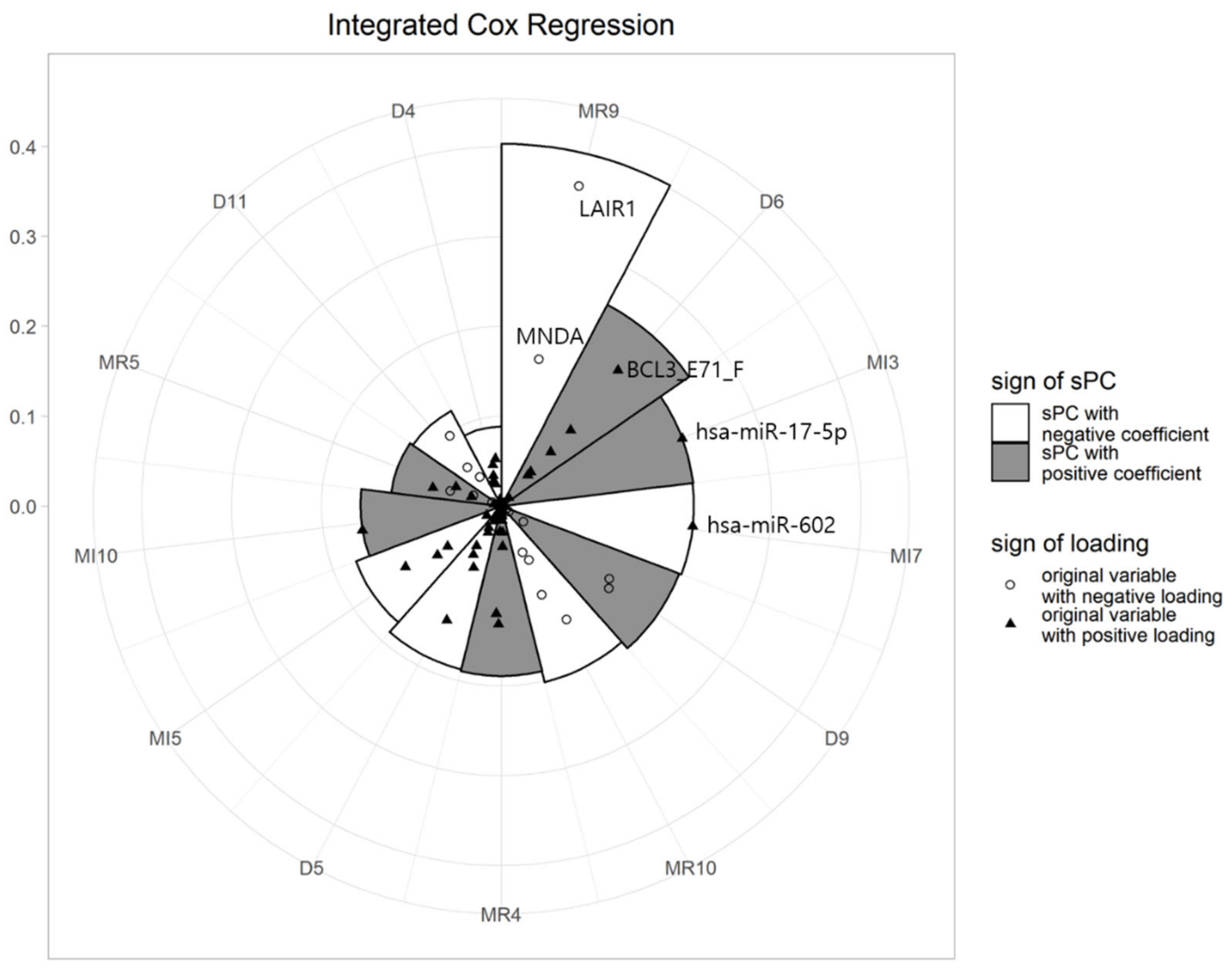
After fitting the Cox regression model, we drew a multi-level polar chart (MP chart), and identified biomarkers related to GBM. Figure 2 shows the MP chart of the results from the integrated dataset. Using this graph, we can easily assess the effects of sPCs on survival time. The illustrative plot depicts that the sPC of the 9th block of mRNA (MR9) had an outstanding effect on survival, followed by sPC of the 6th block of DNA (D6), the 3rd block of miRNA (MI3), and so on. It also shows that the sPCs of MR9 are mainly composed of LAIR1 and MNDA. The coefficients of the two genes, also being negative, shows that the larger the value of these genes, the smaller the value of the sPC. On the other hand, the sign of the sPC for the MR9 coefficient was negative, meaning that the HR decreases, as the value of the sPC for MR9 increases. Therefore, we can interpret that when the values of LAIR1 and MNDA are large, HRs tend to increase and survival times tend to decrease. SPC of D6 was composed of various variables, including BCL3_E71_F. Please note that hsa-miR-602 and hsa-miR-17-5p dominated the sPCs for the 3rd and 7th blocks of miRNA data, respectively.
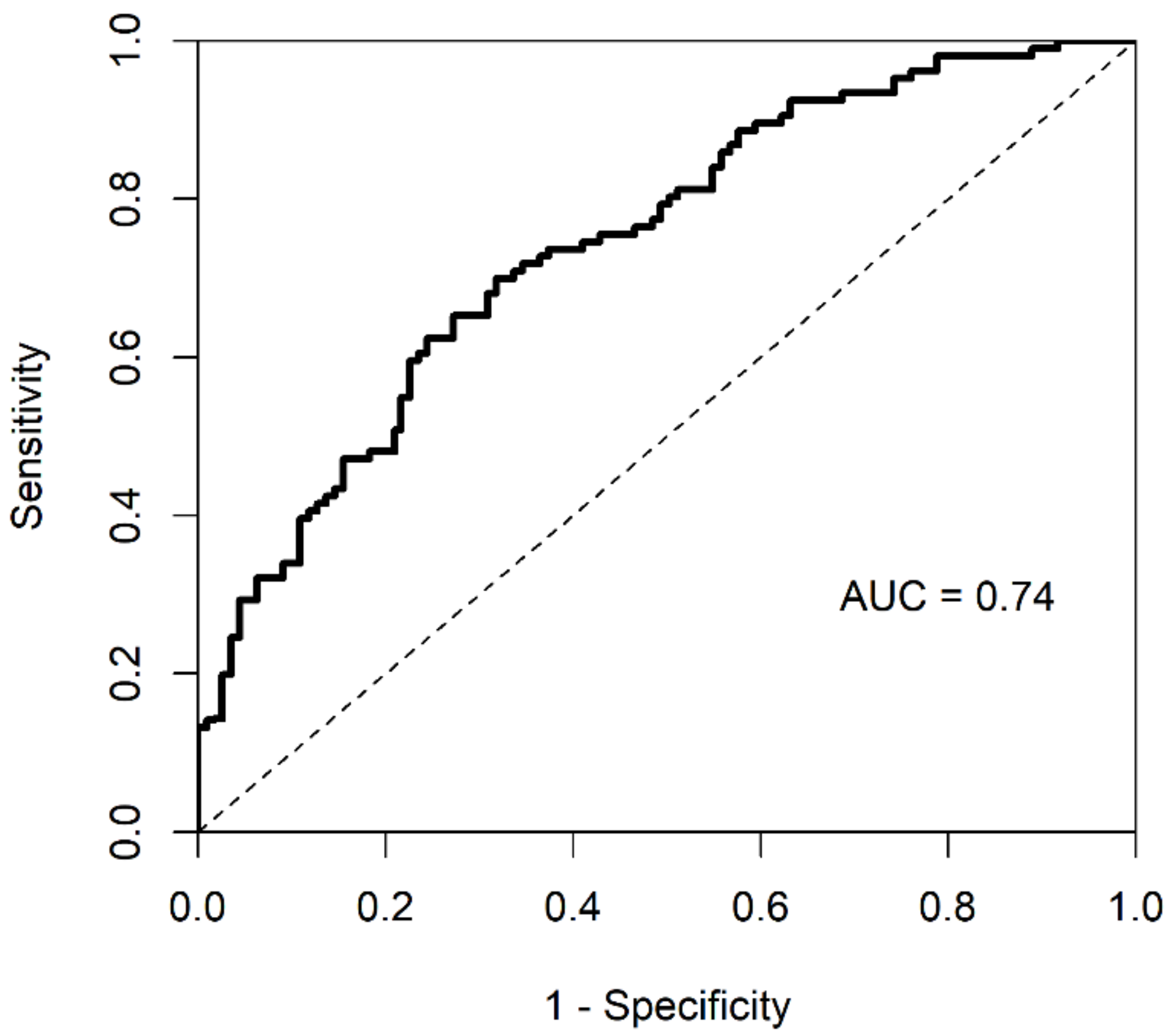
Using the Cox regression model, we predicted whether any specific GBM patient’s survival time is above or below the median survival time. To that end, we assessed the predictive power of the proposed methodology, using a cumulative/dynamic ROC curve and its corresponding AUC value. The cumulative/dynamic ROC curve derived from Cox regression of integrated data is shown in Figure 3. The corresponding AUC value was 0.74 and C-index was 0.67.
2.2. Comparison with Other Approaches
We compared the predictive power of our framework with other methodologies. We focused on comparing our method with others using similar approaches. In particular, we considered (i) PCA analyses without variable clustering, (ii) traditional PCA with clustering, and (iii) iMO-BSPC for each set of omics data. For the first approach, we performed traditional PCA for each omics dataset. That is, neither variable clustering to form homogeneous blocks nor shrinkage to reduce the number of variables was performed. Then, we conducted a Cox regression model with PCs. For the second approach, after variable clustering, we performed PCA for each block. Here, the difference lies in applying PCA instead of sPCA. Finally, in order to compare the results from multi-omics dataset analysis against those from single omics datasets, we applied iMO-BSPC to each omics dataset. For these existing methods, we constructed a Cox model (a) for each omics dataset, and (b) for the entire multi-omics dataset. Therefore, a total of 6 (= 3 × 2) methods, including the proposed method, were compared. Table 2 summarizes the schemes and results of the comparison.
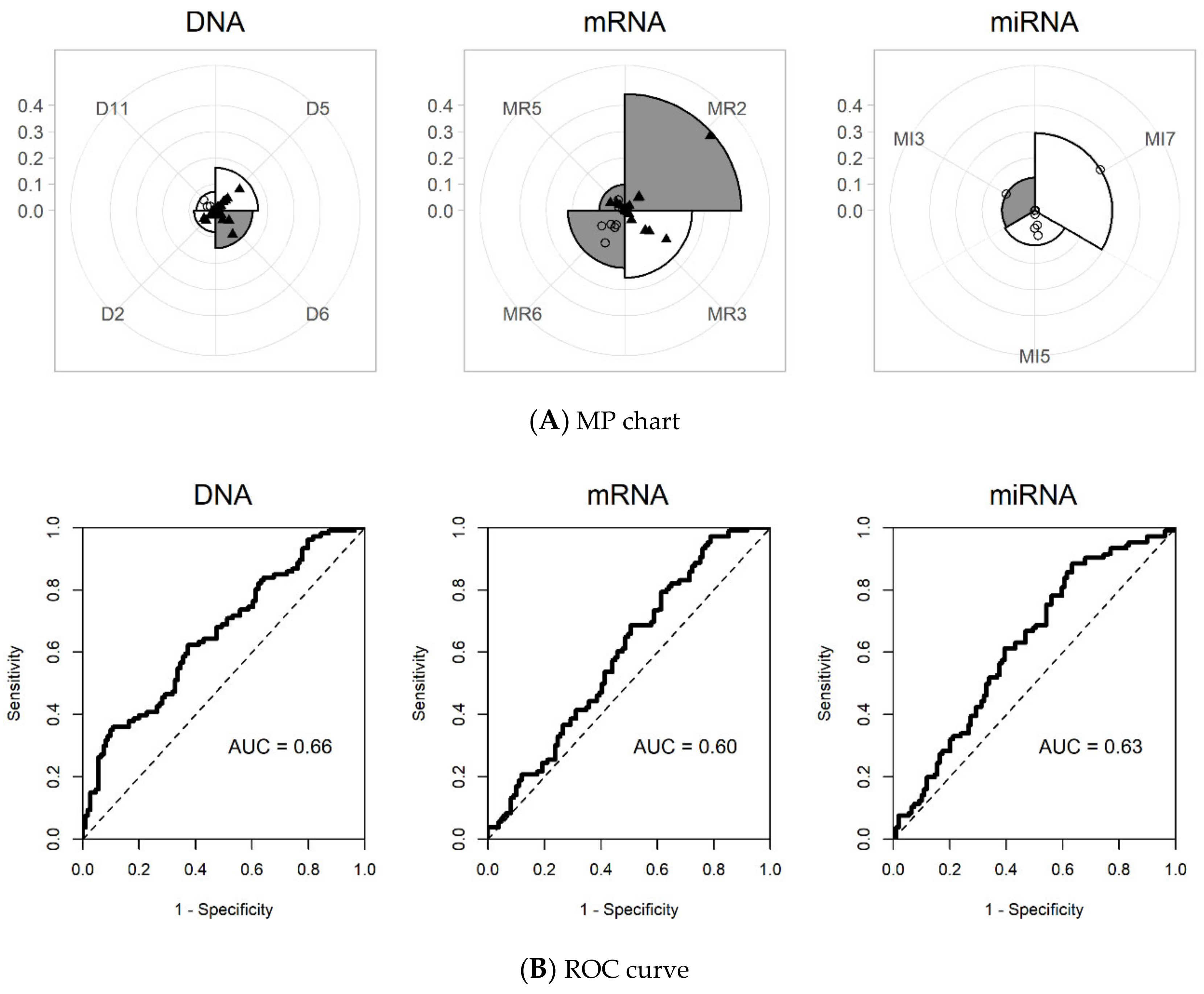
Single omics dataset analysis was also performed, for the purpose of comparison. Since the iMO-BSPC methodology created variable blocks for each omics dataset, there were 12, 10, and 10 blocks for DNA methylation, mRNA expression, and miRNA expression, respectively. During the variable selection process, four sPCs from DNA methylation, four sPCs from mRNA expression, and three sPCs from miRNA expression were finally chosen for building a prediction model. The MP chart generated by iMO-BSPC, from three single omics dataset analyses, is depicted in Figure 4. For DNA data, the sPC of D5 had the greatest influence, followed by the sPCs of D6, D2, and D11. Compared to the multi-omics results, the sPCs of D4 and D9 were removed, while the sPC of D2 was added for single omics. D5 and D6 had coefficients of similar magnitude, but the signs were opposite. For mRNA, sPC of M2 had the greatest influence, followed by the sPCs of M3, M6, and M5. Single omics dataset analysis of mRNA chose sPCs that were all different from those chosen in the multi-omics data analysis. In contrast, for the miRNA datasets, the same sPCs were chosen, except for MI10. Here, the effect of MI7 was shown to be larger than that of the multi-omics case. The AUCs for the DNA, mRNA, and miRNA datasets were 0.66, 0.60, and 0.63, respectively, much lower than the AUC value of 0.74 from the multi-omics data analysis. The C-indices were 0.61, 0.59, and 0.60 for the DNA methylation, mRNA, and miRNA datasets, respectively (Table 2).
When applying traditional PCA without variable clustering, 188 PCs had an eigenvalue of 1 or higher, and the first 78 PCs accounted for more than 80% of the total variance for DNA methylation. For mRNA expression and miRNA expression, 120 and 77 PCs had eigenvalues of 1 or higher, and the first 32 and 39 PCs explained more than 80% of total variance, respectively. For a fair comparison, we used the same number of PCs with the number of sPCs selected by iMO-BSPC. Therefore, the first 12, 10, and 10 PCs were used to build the prediction model. During the variable selection process, 5, 2, and 2 PCs remained for the DNA methylation, mRNA expression, and miRNA expression dataset, respectively. The final PCs in the prediction model were linear combinations of 563, 188, and 225 variables for each omics dataset, respectively, and the number of variables used was much more than that of iMO-BSPC, as seen in Table 2. However, the AUCs and C-indices were rather lower than those of iMO-BSPC, except for the DNA methylation data.
When applying traditional PCA to the blocks after variable clustering, total of 15 PCs with 1661 variables were selected for the prediction model. Compared to iMO-BSPC, the AUC and C-index values were higher (AUC = 0.71, C-index = 0.64) for DNA methylation, from which twice as many PCs were extracted, but the AUC and C-index values were lower than those of iMO-BSPC for both mRNA and miRNA.
For the multi-omics data analysis, we used the first 32 PCs (sPCs) for each methodology. As seen in Table 2, in all methodologies, the AUC and C-index were higher than the values of the single omics datasets, indicating that it is reasonable to use multi-omics data. The comparison study showed that iMO-BSPC had similar predictive performance (AUC = 0.74, C-index = 0.67) to existing PCA-based methods with a much smaller number of variables.
3. Discussion
In this study, we proposed a novel iMO-BSPC (Integrative analysis of Multi-Omics data based on Block-wise sparse Principal Component Analysis) approach for analyzing multi-omics data. Our proposed iMO-BSPC has several advantages. To reduce variable redundancy, iMO-BSPC adopted a shrinkage approach, sPCA, which enabled reductions in dimensionality and the number of explicitly used variables. In addition, the selected variable, the corresponding block, and the results of model fit were expressed in a single graph to enable more intuitive interpretation. The comparison of prediction performance with those of other existing approaches showed that our proposed methodology had comparable AUCs to those of other existing PCA-based methods, with a much smaller number of variables.
However, there are some limitations of our approach. Despite its usefulness in practical applications, sPCA is limited in terms of lack of orthogonality in the loadings of different principal components, the existence of correlations in the principal components, and the expensive computation needed [39]. Also, we used 10% of variables as the threshold. However, more sophisticated methods of choosing the sparsity parameter can be applied. For example, another method chooses the optimal parameter explaining a large proportion of variance with a small number of nonzero loadings among multiple sparsity parameters [10,40]. A sequential method (sPCA-rSVD) using regularized singular value decomposition [41] and an iterative method using penalized matrix decomposition [42] could be used to choose the sparsity parameter. To determine the appropriate number of blocks, we used a simple method of inspecting dendrograms and scree plots. Instead, one may adopt a bootstrap-based approach or use the dissimilarity value or rand index. Another limitation of this study is that it only compared iMO-BSPC with the existing PCA-based method.
iMO-BSPC can be easily extended to various other contexts. If there is a nonlinear relationship, one may apply nonlinear methods, such as kernel PCA, rather than sPCA. Popular kernels, such as Gaussian, polynomial, and hyperbolic tangent kernels can be used [43]. Although we selected Cox regression as a prediction model, our approach can be easily extended to other prediction models and data mining methods such as logistic regression, support vector machine, and random forest. This kind of extension adds high flexibility to our iMO-BSPC and facilitates integrative analyses of multi-omics data.
iMO-BSPC analysis identified several significant genes reported in other studies. The sPCA of 9th clock for mRNA mainly consisted of linear combinations of leukocyte-associated immunoglobulin-like receptor 1 (LAIR1) and human myeloid nuclear differentiation antigen (MNDA). LAIR1 showed the most significant influence among all variables and is known to be broadly expressed in the majority of immune cells [44]. When LAIR-1 binds to its ligands, immune function in the tumor microenvironment is lost, and T cell function and the immune responses of antigen-presenting cells are reduced [45]. Several studies have reported that LAIR1 is significantly upregulated in multiple types of solid tumors such as ovarian cancer, human cervical cancer, and GBM [44,45,46,47,48]. MNDA was the variable with the second largest loading, suggesting that it plays a significant role. MNDA appears to regulate the activity of transcription factors and, in some cases, serves to mediate cell death [49]. Several studies reported that NMDA plays a role as a suppressor of cancer in pancreatic cancer, osteosarcoma, and GBM [50,51,52]. The variables with the largest loadings in DNA methylation cluster 6, DNA cluster 5, miRNA cluster 7, mRNA cluster 10, and miRNA cluster 3 also had significant influences. The representative variable of DNA cluster 6 is BCL3_E71_F. It should be noted that B cell CLL/lymphoma 3 (BCL-3) is a proto-oncogene candidate. BCL3 was reported to be an informative indicator of glioma response to alkylating chemotherapy [53]. It was also identified as a target gene for hepatocellular carcinoma [53,54]. The hsa-miR-602 gene of micro-RNA, homo sapiens miR-602 stem-loop, represented miRNA cluster 3. It has been reported as a high-risk gene for mesenchymal subtype GBM, liver cancer, and hepatocellular carcinoma [55,56,57]. The gene with the largest loading in miRNA cluster 7 was hsa-miR-17–5p, and previous studies have found miR17 to be associated with polycystic kidney disease and B-cell lymphomas. It was identified as a high-risk gene for renal cell carcinoma and colorectal cancer [58,59].
Using 338 pathways in the KEGG database, we performed Fisher’s exact test to identify pathways significantly associated with genes. The Benjamin–Hochberg method was used to adjust the p-values. The Fisher’s exact test for the genes in D9 yielded 8 significant pathways with q-values smaller than 5%. Among them, the PI3K-Akt signaling pathway, TNF signaling pathway, and small cell lung cancer have been reported to be related to cancer [60]. In addition, the ECM–receptor interaction, focal adhesion, and cell adhesion molecules pathways were also significant, and could be related to invasive behavior of cancer cells [61]. The Fisher’s exact test for the genes in MR9 yielded Staphylococcus aureus infection as a significant pathway, with a q-value smaller than 5%. It was reported to be related to infection with S. aureus.
4. Materials and Methods
Our iMO-BSPC approach consists of two stages. In the first stage, we divide whole variables into non-overlapping, homogeneous blocks, using a variable clustering technique, and then construct shrunken components for each block by applying sPCA. This procedure is repeated for each single omics data set. In the second stage, we generate a new data set, by parallel combination of sPCs, from each omics data set. Then, we conduct Cox regression analysis, to construct a prognostic model. The detailed algorithm is given as follows (Figure 5).
4.1. Stage 1: Finding Blockwise Components for Each Omics Data
The aim of the first stage is to find a few representative components, for each omics dataset. To accomplish this, we identify homogeneous variable blocks, and then extract sparse PCs.
4.1.1. Finding Homogeneous Variable Blocks
To find homogeneous blocks, we conduct variable clustering, for each omics dataset. There are several methods for clustering, including the VARCLUS procedure of SAS software [62], clustering around latent variables (CLV), and diametrical clustering [42,63]. In this study, we use the ClustOfVar algorithm, which is based on PCAMIX [43,64]. Though it can handle a mixture of qualitative and quantitative variables, we assume that there are only quantitative variables. The algorithm of ClustOfVar packages are as follows.
Consider we have a J omics data set, and that the jth omics dataset consists of samples and variables. In our example, we used three omics datasets of DNA methylation, RNA expression and miRNA expression. The DNA methylation data set consisted of 215 samples of GBM patients and 1305 genetic variables. Similarly, the RNA expression and miRNA expression dataset consisted of 1205 and 534 genetic variables for 215 patients, respectively.
Let be a partition into clusters of variables. For each cluster, , define a synthetic variable which is “most linked” to all the variables in the cluster. That is,
where denotes the squared Pearson correlation coefficient [64]. The homogeneity of a cluster is defined as follows.
Please note that is the first principal component of PCAMIX, and is the first eigenvalue obtained, for cluster [64]. Now, the aim is to find a partition,which maximizes the homogeneity function .
In this study, we consider the agglomerative hierarchical clustering algorithm, and thus we start with the partition into clusters. Then, we agglomerate two clusters, and , with the smallest dissimilarity measure, , which is defined as:
This aggregation is then repeated until all variables belong to a single cluster.
To determine the appropriate number of clusters, we use a simple method for inspecting dendrograms and scree plots. We call the final clusters “blocks” This procedure is conducted, for each omics dataset, to find its homogeneous variable blocks. In the GBM data analysis, we obtained 12 homogeneous gene blocks for DNA methylation dataset, and RNA expression and miRNA expression dataset are divided into 10 homogeneous gene blocks each.
4.1.2. Extracting Sparse Principal Components
To find surrogate variables for each block, we apply sparse principal component analysis (sPCA). SPCA aims to find a set of sparse weight vectors; that is, weight vectors with only a few nonzero values [10,65]. Let be a kth block, of the jth omics data set. Then, the first sparse principal components, , for a data matrix, can be obtained by optimization, as follows:
for any . Then, is derived as , where denotes the ith row vector of the matrix [10]. Among the generated series of sPCs, we used the first sPC as a representative variable.
We then apply this procedure to every block, in each omics dataset. Once the variable is divided into blocks, we can then construct a new dataset for the jth omics dataset, where represents the first sPC for the kth block of the jth omics data set.
4.2. Stage 2: Integrative Analysis for Multi-Omics Data
In this stage, we firstly integrate the information from the omics datasets in parallel, and then construct a prediction model using the sPCs obtained from stage 1. We also assess the performance of the model, using AUC, and visualize the results by a graph.
4.2.1. Parallel Integration of Omics Data, and Construction of a Prediction Model
Here, we combine the sPCs, from every omics data set and construct a new dataset . Then, the new dataset has rows and columns. For example, in the GBM data analysis, the new dataset consisted of 215 rows of patients and 32 columns of sPCs from three omics datasets. Set an matrix,, consisting of two variables of survival time , and a censoring indicator, . We can then consider Cox regression with the hazard function as:
where is a baseline hazard function, and a stepwise selection procedure is applied to select variables. Among various stepwise procedures, we used Akaike information criterion (AIC) in this study [66].
4.2.2. Assessing Prediction Performance
To evaluate predictive power, we use evaluation measures such as the sensitivity and specificity for time-to-event data [67]. We then divide the observations into two classes, above- and below-median survival times. Then, we evaluate whether our model classifies individuals properly. This classification was evaluated using cumulative/dynamic, time-dependent receiver operator characteristic (ROC) curves, corresponding to AUC values [68]. The sensitivity and specificity are then defined, at each time point, and are denoted as cumulative sensitivity and dynamic specificity. Given the cumulative sensitivity and dynamic specificity, the cumulative/dynamic ROC curve was plotted with 1-specificity, at the x-axis, and sensitivity at the y-axis, as in an ordinary ROC curve. The cumulative/dynamic ROC curve, and corresponding AUC value, can be derived using the cdROC() function of the R package nsROC.
Let denote the diagnostic marker and denote the survival time. Then, the cumulative sensitivity and dynamic specificity are defined as follows [68].
Given the cumulative sensitivity and dynamic specificity, the cumulative/dynamic ROC curve was plotted with 1-specificity, at the x-axis, and sensitivity at the y-axis, as in an ordinary ROC curve. The cumulative/dynamic ROC curve, and corresponding AUC value, can be derived using the cdROC() function of the R package nsROC.
We also compute Harrell’s C-index, also known as concordance index, which has been commonly used to assess and compare the discriminative power of risk prediction models [69]. It can be expressed in a formula
where and are risk score and time to event for the ith patient, respectively; denotes event indicator for the ith patient, with 0 for censoring and 1 for observed event; and is an indicator function [70]. The C-index can be interpreted as the fraction of all pairs of subjects whose predicted survival times are correctly ordered among all subjects who can actually be ordered [71]. We used the coxph() function of the R package ’Survival’ to calculate the C-index.
4.2.3. Visualization
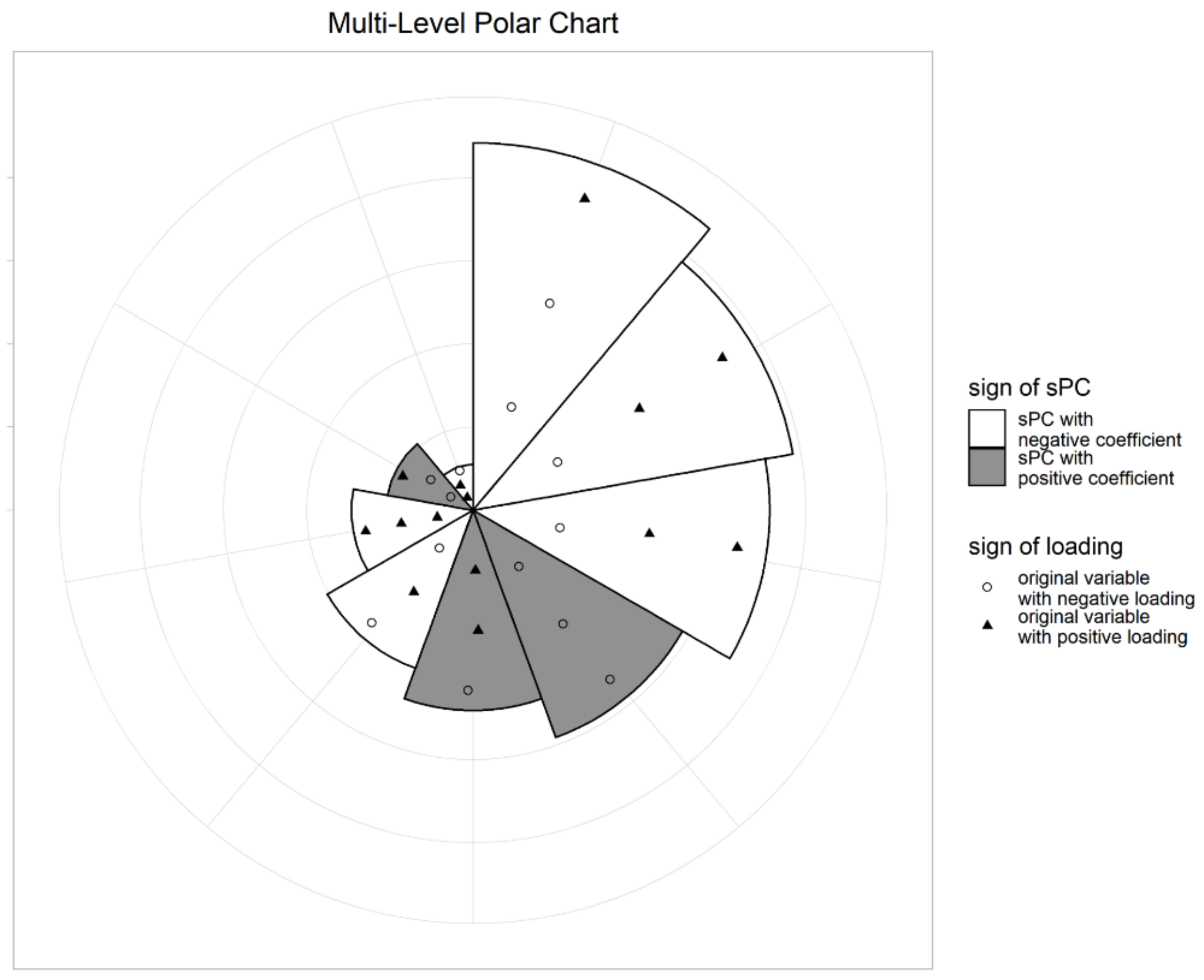
For visualization of the results, from the two stages together, we propose a new graph representing the estimated coefficients of selected sPCs, and loadings of the original variables, consisting of sPCs. Since this shows two levels of coefficients, from the Cox regression model and sPCA analysis, in a polar chart, we call it a multi-level polar chart (MP chart).
Suppose that sPCs are selected from stage 2. Draw a circle and divide the angle equally into pieces. For the first level chart, sort sPCs by the absolute value of the coefficients in the Cox regression model, and determine the corresponding sector clockwise. Adjust the radius of each sector to be proportional to the absolute value of the coefficient of the corresponding sPCs, . The signs of coefficients are distinguished by the color of each sector. For example, the white sector indicates that the coefficient of the corresponding sPC is negative. At the second level of the chart, to represent how the sPCs are constructed, we plot the associated original variables with sPCs in the sector. Then, the distance from the origin of each variable is proportional to the corresponding loading of sPC. The sign is represented by the color and/or shape of the point. For example, white circles in a sector indicate that the variable loading is negative, while black triangles indicate that the variable loading is positive. Jittering would be done to avoid duplicates, and to clearly distinguish between variable points. Thus, the MP chart can simultaneously represent the important sPCs for survival time prediction, and what variables make up those sPCs. An illustrative example of our plots is presented in Figure 6.
Supplementary Materials
Supplementary materials can be found at https://0-www-mdpi-com.brum.beds.ac.uk/1422-0067/21/21/8202/s1.
Author Contributions
Conceptualization, M.P.; methodology, M.P. and T.P.; software, D.K. and K.M.; formal analysis, D.K. and K.M.; writing—original draft preparation, M.P.; writing—review and editing, T.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Basic Science Research Program, through the National Research Foundation of Korea (NRF), funded by the Ministry of Science, ICT & Future Planning (NRF-2017R1A2B4011504). This research was also supported by the Bio & Medical Technology Development Program of the NRF, funded by the Korean government, MSIP (No. 2016M3A9B6904244, 2013M3A9C4078158).
Acknowledgments
We would like to thank Myung-Hoe Huh who gave much valuable advice. We also thank Kyulhee Han for helping with the pathway analysis.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AIC | Akaike information criterion |
| AUC | Area under the curve |
| GBM | Glioblastoma multiforme |
| HR | Hazard ratio |
| iMO-BSPC | Integrative analysis of multi-omics data based on block-wise sparse principal component analysis |
| LASSO | least absolute shrinkage and selection operator |
| MP chart | Multi-level polar chart |
| PCA | Principal component analysis |
| PLS | Partial least squares |
| ROC curve | Receiver operator characteristic curve |
| SAS | Statistical analysis system |
| sPC | Sparse principal component |
| TCGA | The Cancer Genome Atlas |
References
- Shafi, A.; Nguyen, T.; Peyvandipour, A.; Nguyen, H.; Draghici, S. A Multi-Cohort and Multi-Omics Meta-Analysis Framework to Identify Network–Based Gene Signatures. Front. Genet. 2019, 10, 159. [Google Scholar] [CrossRef]
- Greenawalt, D.M.; Sieberts, S.K.; Cornelis, M.C.; Girman, C.J.; Zhong, H.; Yang, X.; Guinney, J.; Qi, L.; Hu, F.B. Integrating genetic association, genetics of gene expression, and single nucleotide polymorphism set analysis to identify susceptibility Loci for type 2 diabetes mellitus. Am. J. Epidemiol. 2012, 176, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karczewski, K.J.; Snyder, M.P. Integrative omics for health and disease. Nat. Rev. Genet. 2018, 19, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Zeleznik, O.A.; Thallinger, G.G.; Kuster, B.; Gholami, A.M.; Culhane, A.C. Dimension reduction techniques for the integrative analysis of multi-omics data. Brief. Bioinform. 2016, 17, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Abraham, G.; Inouye, M. Fast principal component analysis of large-scale genome-wide data. PLoS ONE 2014, 9, e93766. [Google Scholar] [CrossRef] [Green Version]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef]
- Alonso–Gutierrez, J.; Kim, E.-M.; Batth, T.S.; Cho, N.; Hu, Q.; Chan, L.J.G.; Petzold, C.J.; Hillson, N.J.; Adams, P.D.; Keasling, J.D.; et al. Principal component analysis of proteomics (PCAP) as a tool to direct metabolic engineering. Metab. Eng. 2015, 28, 123–133. [Google Scholar]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Bougeard, S.; Abdi, H.; Saporta, G.; Niang, N. Clusterwise analysis for multiblock component methods. Adv. Data Anal. Classif. 2018, 12, 285–313. [Google Scholar] [CrossRef]
- Zou, H.; Hastie, T.; Tibshirani, R. Sparse Principal Component Analysis. J. Comput. Graph. Stat. 2006, 15, 265–286. [Google Scholar] [CrossRef] [Green Version]
- Cadima, J.; Jolliffe, I.T. Loading and correlations in the interpretation of principle compenents. J. Appl. Stat. 1995, 22, 203–214. [Google Scholar] [CrossRef]
- Yu, L.; Liu, H. Efficient feature selection via analysis of relevance and redundancy. J. Mach. Learn. Res. 2004, 5, 1205–1224. [Google Scholar]
- Venkatesh, B.; Anuradha, J. A review of feature selection and its methods. Cybern. Inf. Technol. 2019, 19, 3–26. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, V.N.; Lingjærde, O.C.; Russnes, H.G.; Vollan, H.K.M.; Frigessi, A.; Børresen–Dale, A. –L. Principles and methods of integrative genomic analyses in cancer. Nat. Rev. Cancer 2014, 14, 299. [Google Scholar] [CrossRef] [PubMed]
- Pineda, S.; Real, F.X.; Kogevinas, M.; Carrato, A.; Chanock, S.J.; Malats, N.; Van Steen, K. Integration Analysis of Three Omics Data Using Penalized Regression Methods: An Application to Bladder Cancer. PLoS Genet. 2015, 11, e1005689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.; Tseng, G.C.; Sun, W. Statistical Methods in Integrative Genomics. Annu. Rev. Stat. Appl. 2016, 3, 181–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.R.; Attia, J.; Minelli, C. The meta-analysis of genome-wide association studies. Brief. Bioinform. 2011, 12, 259–269. [Google Scholar] [CrossRef]
- Begum, F.; Ghosh, D.; Tseng, G.C.; Feingold, E. Comprehensive literature review and statistical considerations for GWAS meta-analysis. Nucleic Acids Res. 2012, 40, 3777–3784. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Zhou, F.; Ren, J.; Li, X.; Jiang, Y.; Ma, S. A Selective Review of Multi-Level Omics Data Integration Using Variable Selection. High Throughput 2019, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Shi, X.; Xie, Y.; Huang, J.; Shia, B.; Ma, S. Combining multidimensional genomic measurements for predicting cancer prognosis: Observations from TCGA. Brief. Bioinform. 2014, 16, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Shi, X.; Zhao, Q.; Krauthammer, M.; Rothberg, B.E.G.; Ma, S. Integrated analysis of multidimensional omics data on cutaneous melanoma prognosis. Genomics 2016, 107, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Zou, H. The Adaptive Lasso and Its Oracle Properties. J. Am. Stat. Assoc. 2006, 101, 1418–1429. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Li, R. Variable Selection via Nonconcave Penalized Likelihood and its Oracle Properties. J. Am. Stat. Assoc. 2001, 96, 1348–1360. [Google Scholar] [CrossRef]
- Zhang, C.-H. Nearly unbiased variable selection under minimax concave penalty. Ann. Stat. 2010, 38, 894–942. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Ma, S. A selective review of robust variable selection with applications in bioinformatics. Brief Bioinform. 2015, 16, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Ickstadt, K.; Schäfer, M.; Zucknick, M. Toward Integrative Bayesian Analysis in Molecular Biology. Annu. Rev. Stat. Its Appl. 2018, 5, 141–167. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.H. Greedy function approximation: A gradient boosting machine. Ann. Stat. 2001, 29, 1189–1232. [Google Scholar] [CrossRef]
- Hassani, S.; Martens, H.; Qannari, E.M.; Hanafi, M.; Borge, G.I.; Kohler, A. Analysis of -omics data: Graphical interpretation– and validation tools in multi–block methods. Chemom. Intell. Lab. Syst. 2010, 104, 140–153. [Google Scholar] [CrossRef]
- Hassani, S.; Hanafi, M.; Qannari, E.M.; Kohler, A. Deflation strategies for multi-block principal component analysis revisited. Chemom. Intell. Lab. Syst. 2013, 120, 154–168. [Google Scholar] [CrossRef]
- Yang, Z.; Michailidis, G. A non-negative matrix factorization method for detecting modules in heterogeneous omics multi-modal data. Bioinformatics 2016, 32, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waaijenborg, S.; Zwinderman, A.H. Sparse canonical correlation analysis for identifying, connecting and completing gene–expression networks. Bmc Bioinform. 2009, 10, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Safo, S.E.; Long, Q. Incorporating biological information in sparse principal component analysis with application to genomic data. BMC Bioinform. 2017, 18, 332. [Google Scholar] [CrossRef]
- Lock, E.F.; Hoadley, K.A.; Marron, J.S.; Nobel, A.B. Joint and individual variation explained (jive) for integrated analysis of multiple data types. Ann. Appl. Stat. 2013, 7, 523–542. [Google Scholar] [CrossRef]
- Li, Y.; Wu, F.X.; Ngom, A. A review on machine learning principles for multi–view biological data integration. Brief Bioinform. 2018, 19, 325–340. [Google Scholar] [CrossRef]
- Shen, R.; Olshen, A.B.; Ladanyi, M. Integrative clustering of multiple genomic data types using a joint latent variable model with application to breast and lung cancer subtype analysis. Bioinformatics 2009, 25, 2906–2912. [Google Scholar] [CrossRef]
- Wang, W.; Baladandayuthapani, V.; Morris, J.S.; Broom, B.M.; Manyam, G.; Do, K.-A. iBAG: Integrative Bayesian analysis of high–dimensional multiplatform genomics data. Bioinformatics 2013, 29, 149–159. [Google Scholar] [CrossRef]
- Subramanian, I.; Verma, S.; Kumar, S.; Jere, A.; Anamika, K. Multi-omics Data Integration, Interpretation, and Its Application. Bioinform. Biol. Insights 2020, 14, 1177932219899051. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Luo, R.; Zhao, H. Sparse principal component analysis by choice of norm. J. Multivar. Anal. 2013, 114, 127–160. [Google Scholar] [CrossRef]
- Huh, M.-H. Representing variables in the latent space. Korean J. Appl. Stat. 2017, 30, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, I.S.; Marcotte, E.M.; Roshan, U. Diametrical clustering for identifying anti-correlated gene clusters. Bioinformatics 2003, 19, 1612–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiers, H.A. Simple structure in component analysis techniques for mixtures of qualitative and quantitative variables. Psychometrika 1991, 56, 197–212. [Google Scholar] [CrossRef]
- Wang, Q. Kernel principal component analysis and its applications in face recognition and active shape models. arXiv 2012, preprint. arXiv:1207.3538. [Google Scholar]
- Wang, Y.; Zhang, X.; Miao, F.; Cao, Y.; Xue, J.; Cao, Q.; Zhang, X. Clinical significance of leukocyte-associated immunoglobulin-like receptor-1 expression in human cervical cancer. Exp. Ther. Med. 2016, 12, 3699–3705. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wang, S.; Li, J.; Li, J.; Li, B. Cancer immunotherapy based on blocking immune suppression mediated by an immune modulator LAIR–1. OncoImmunology 2020, 9, 1740477. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Guan, G.F.; Cao, J.Y.; Zou, C.Y.; Zhu, C.; Cheng, W.; Xu, X.Y.; Lin, Z.G.; Cheng, P.; Wu, A.H. Overexpression of oncostatin M receptor regulates local immune response in glioblastoma. J. Cell. Physiol. 2019, 234, 15496–15509. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, L.; Shangguan, F.; Zhao, X.; Wang, W.; Gao, Z.; Zhou, H.; Qu, G.; Huang, Y.; An, J. LAIR–1 suppresses cell growth of ovarian cancer cell via the PI3K–AKT–mTOR pathway. Aging 2020, 12, 16142. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, L.; Zhou, J.; Liu, L.; Fu, Q.; Fu, A.; Feng, X.; Xin, R.; Liu, H.; Gao, Y. Clinicopathologic significance of LAIR–1 expression in hepatocellular carcinoma. Curr. Probl. Cancer 2019, 43, 18–26. [Google Scholar] [CrossRef]
- Briggs, R.C.; Atkinson, J.B.; Miranda, R.N. Variable expression of human myeloid specific nuclear antigen MNDA in monocyte lineage cells in atherosclerosis. J. Cell. Biochem. 2005, 95, 293–301. [Google Scholar] [CrossRef]
- North, W.G.; Liu, F.; Lin, L.Z.; Tian, R.; Akerman, B. NMDA receptors are important regulators of pancreatic cancer and are potential targets for treatment. Clin. Pharmacol. Adv. Appl. 2017, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Liu, C.; Dong, J.; Li, D.; Li, W. Effects of the myeloid cell nuclear differentiation antigen on the proliferation, apoptosis and migration of osteosarcoma cells. Oncol. Lett. 2014, 7, 815–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Q.; Li, R.; Li, Y.; Xu, X.; Ni, W.; Lin, H.; Ning, L. Prediction of a competing endogenous RNA co-expression network as a prognostic marker in glioblastoma. J. Cell. Mol. Med. 1–10. [CrossRef] [PubMed]
- Wu, L.; Bernal, G.M.; Cahill, K.E.; Pytel, P.; Fitzpatrick, C.A.; Mashek, H.; Weichselbaum, R.R.; Yamini, B. BCL3 expression promotes resistance to alkylating chemotherapy in gliomas. Sci. Transl. Med. 2018, 10, eaar2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Cui, X.; Wang, Q.; Fang, C.; Tan, Y.; Wang, Y.; Yi, K.; Yang, C.; You, H.; Shang, R. RUNX1 contributes to the mesenchymal subtype of glioblastoma in a TGFβ pathway–dependent manner. Cell Death Dis. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Huang, Y.; Chen, Y.; Xie, Y.; Wen, H.; Tan, W.; Wang, C. miR–602 Mediates the RASSF1A/JNK Pathway, Thereby Promoting Postoperative Recurrence in Nude Mice with Liver Cancer. Oncotargets Ther. 2020, 13, 6767. [Google Scholar] [CrossRef]
- Li, R.; Gao, K.; Luo, H.; Wang, X.; Shi, Y.; Dong, Q.; Luan, W.; You, Y. Identification of intrinsic subtype–specific prognostic microRNAs in primary glioblastoma. J. Exp. Clin. Cancer Res. 2014, 33, 9. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Ma, Z.; Wang, D.; Zhao, W.; Chen, L.; Wang, G. MicroRNA–602 regulating tumor suppressive gene RASSF1A is over–expressed in hepatitis B virus–infected liver and hepatocellular carcinoma. Cancer Biol. Ther. 2010, 9, 803–808. [Google Scholar] [CrossRef] [Green Version]
- Tsz–fung, F.C.; Mankaruos, M.; Scorilas, A.; Youssef, Y.; Girgis, A.; Mossad, S.; Metias, S.; Rofael, Y.; Honey, R.J.; Stewart, R. The miR–17–92 cluster is over expressed in and has an oncogenic effect on renal cell carcinoma. J. Urol. 2010, 183, 743–751. [Google Scholar]
- Fu, F.; Jiang, W.; Zhou, L.; Chen, Z. Circulating exosomal miR–17–5p and miR–92a–3p predict pathologic stage and grade of colorectal cancer. Transl. Oncol. 2018, 11, 221–232. [Google Scholar] [CrossRef]
- Agnihotri, S.; Burrell, K.E.; Wolf, A.; Jalali, S.; Hawkins, C.; Rutka, J.T.; Zadeh, G. Glioblastoma, a brief review of history, molecular genetics, animal models and novel therapeutic strategies. Arch. Immunol. Ther. Exp. 2013, 61, 25–41. [Google Scholar] [CrossRef]
- Sayegh, E.T.; Kaur, G.; Bloch, O.; Parsa, A.T. Systematic review of protein biomarkers of invasive behavior in glioblastoma. Mol. Neurobiol. 2014, 49, 1212–1244. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT12.1; SAS Institute, Inc.: Cary, NC, USA, 2012; pp. 8497–8529. [Google Scholar]
- Vigneau, E.; Qannari, E. Clustering of variables around latent components. Commun. Stat. Simul. Comput. 2003, 32, 1131–1150. [Google Scholar] [CrossRef]
- Chavent, M.; Kuentz–Simonet, V.; Liquet, B.; Saracco, J. ClustOfVar: An R Package for the Clustering of Variables. arXiv 2012, arXiv:1112.0295. [Google Scholar]
- Feng, C.-M.; Gao, Y.-L.; Liu, J.-X.; Zheng, C.-H.; Li, S.-J.; Wang, D. A Simple Review of Sparse Principal Components Analysis, Proceedings of the International Conference on Intelligent Computing, Lanzhou, China, 2–5 August 2016; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Wolf, P.; Schmidt, G.; Ulm, K. The use of ROC for defining the validity of the prognostic index in censored data. Stat. Probab. Lett. 2011, 81, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Heagerty, P.J.; Zheng, Y. Survival model predictive accuracy and ROC curves. Biometrics 2005, 61, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrell, F.E., Jr.; Califf, R.M.; Pryor, D.B.; Lee, K.L.; Rosati, R.A. Evaluating the Yield of Medical Tests. JAMA 1982, 247, 2543–2546. [Google Scholar] [CrossRef]
- Schmid, M.; Wright, M.N.; Ziegler, A. On the use of Harrell’s C for clinical risk prediction via random survival forests. Expert Syst. Appl. 2016, 63, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Raykar, V.C.; Steck, H.; Krishnapuram, B.; Dehing–Oberije, C.; Lambin, P. On Ranking in Survival Analysis: Bounds on the Concordance Index. In Proceedings of the 20th International Conference on Neural Information Processing Systems, Daegu, Korea, 3–7 November 2013; Curran Associates Inc.: Vancouver, BC, Canada, 2007; pp. 1209–1216. [Google Scholar]
Figure 1.
Scree plots and dendrograms for GBM data. (A) Scree plot of dissimilarity according to number of clusters. Line in the scree plot represent cut-off lines. (B) Dendrogram from hierarchical clustering. Rectangles in dendrogram represent homogeneous variable blocks.
Figure 1.
Scree plots and dendrograms for GBM data. (A) Scree plot of dissimilarity according to number of clusters. Line in the scree plot represent cut-off lines. (B) Dendrogram from hierarchical clustering. Rectangles in dendrogram represent homogeneous variable blocks.

Figure 2.
MP chart of integrated dataset analysis (MR: mRNA, D:DNA methylation, MI:miRNA).
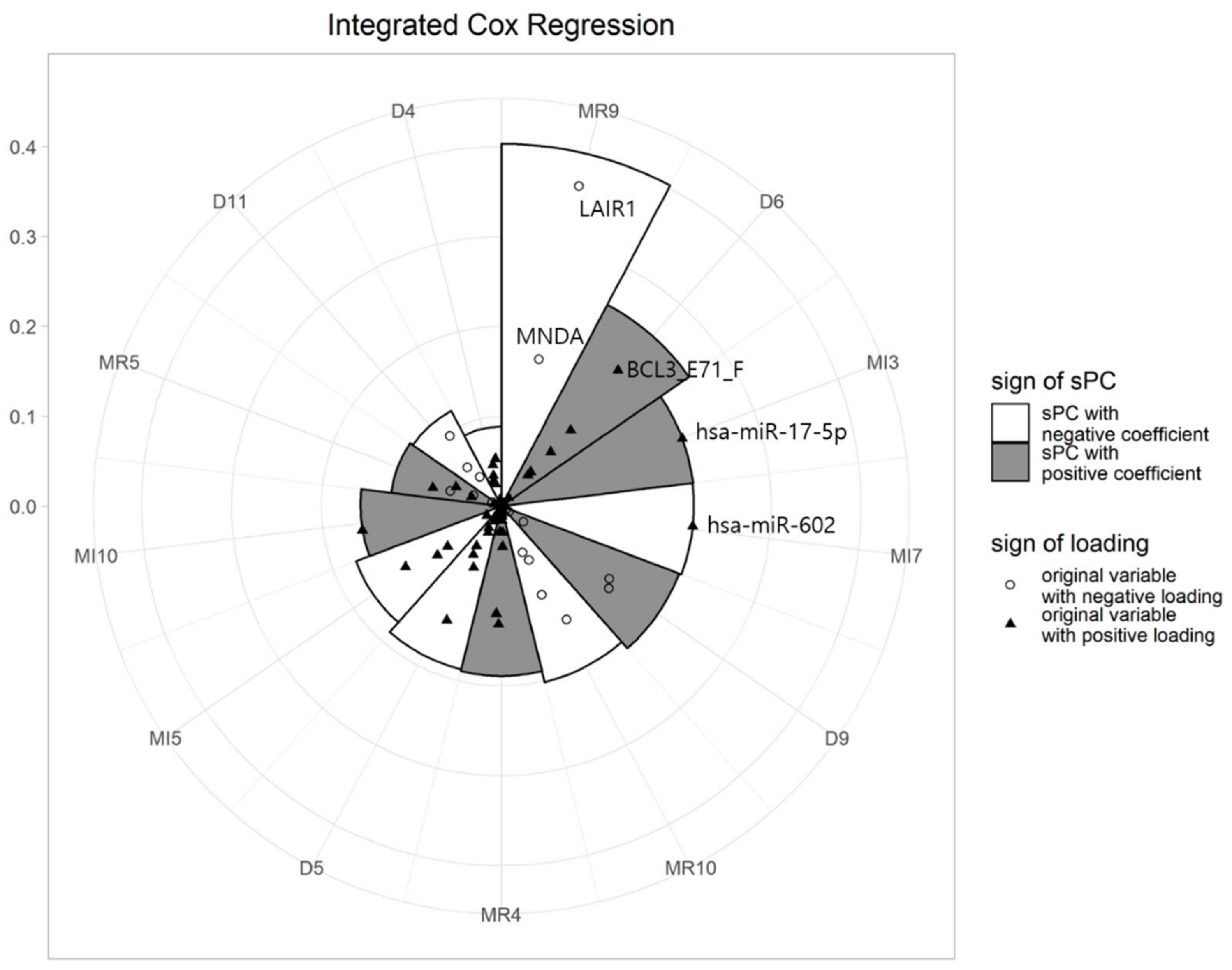
Figure 3.
ROC curve and AUC value of integrated dataset by iMO-BSPC.

Figure 4.
MP charts and ROC curves for each single omics dataset using iMO-BSPC. (A) MP chart. (B) ROC curve (DNA: DNA methylation, mRNA: mRNA expression, miRNA: miRNA expression).
Figure 4.
MP charts and ROC curves for each single omics dataset using iMO-BSPC. (A) MP chart. (B) ROC curve (DNA: DNA methylation, mRNA: mRNA expression, miRNA: miRNA expression).

Figure 5.
Procedure of iMO-BSPC.

Figure 6.
Example of a multi-level polar chart (MP chart): the radius of a sector is proportional to the coefficient of the sPC, and the distance from the origin to specific points is proportional to the variable loading.
Figure 6.
Example of a multi-level polar chart (MP chart): the radius of a sector is proportional to the coefficient of the sPC, and the distance from the origin to specific points is proportional to the variable loading.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Stepwise Cox regression results for the integrated datasets.
| Omics | Block | Number of Variables | Number of Variables Remained | Coefficient (Standard Error) | Hazard Ratio | 1p-Value |
|---|---|---|---|---|---|---|
| DNA | D4 | 128 | 13 | −0.09 (0.05) | 0.92 | 0.089 |
| D5 | 186 | 19 | −0.19 (0.05) | 0.83 | <0.001 | |
| D6 | 153 | 15 | 0.25 (0.06) | 1.29 | <0.001 | |
| D9 | 69 | 7 | 0.21 (0.07) | 1.24 | 0.004 | |
| D11 | 46 | 5 | −0.12 (0.05) | 0.89 | 0.011 | |
| mRNA | MR4 | 188 | 19 | 0.19 (0.07) | 1.21 | 0.009 |
| MR5 | 161 | 16 | 0.12 (0.06) | 1.13 | 0.033 | |
| MR9 | 65 | 6 | −0.40 (0.10) | 0.67 | <0.001 | |
| MR10 | 67 | 7 | −0.20 (0.07) | 0.82 | 0.011 | |
| miRNA | MI3 | 32 | 3 | 0.21 (0.08) | 1.24 | 0.005 |
| MI5 | 79 | 8 | −0.17 (0.05) | 0.84 | <0.001 | |
| MI7 | 56 | 6 | −0.21 (0.09) | 0.81 | 0.021 | |
| MI10 | 22 | 2 | 0.16 (0.06) | 1.17 | 0.016 |
1 Uncorrected p-value.
Table 2.
Comparisons of predictability with other approaches.
| Methodology | Single Omics | Multi-Omics | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (1) | (2) | (3) | (1) | (2) | (3) | |||||||
| Omics | (a) | (b) | (c) | (a) | (b) | (c) | (a) | (b) | (c) | all | all | all |
| 1 PC-before | 12 | 10 | 10 | 12 | 10 | 10 | 12 | 10 | 10 | 32 | 32 | 32 |
| 2 PC-after | 5 | 2 | 2 | 8 | 4 | 3 | 4 | 4 | 3 | 13 | 15 | 13 |
| 3 variable | 563 | 188 | 225 | 935 | 559 | 167 | 56 | 49 | 17 | 1580 | 1339 | 126 |
| AUC | 0.67 | 0.54 | 0.61 | 0.71 | 0.58 | 0.59 | 0.66 | 0.60 | 0.63 | 0.75 | 0.76 | 0.74 |
| C-index | 0.63 | 0.55 | 0.60 | 0.64 | 0.61 | 0.60 | 0.61 | 0.59 | 0.60 | 0.69 | 0.69 | 0.67 |
(1) PCA without variable clustering, (2) PCA with clustering, (3) iMO-BSPC; (a) DNA methylation, (b) mRNA expression, (c) miRNA expression; 1 Number of PCs (sPCs) before stepwise variable selection process; 2 Number of PCs (sPCs) selected by stepwise variable selection process; 3 Number of variables used for prediction.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, M.; Kim, D.; Moon, K.; Park, T. Integrative Analysis of Multi-Omics Data Based on Blockwise Sparse Principal Components. Int. J. Mol. Sci. 2020, 21, 8202. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218202
AMA Style
Park M, Kim D, Moon K, Park T. Integrative Analysis of Multi-Omics Data Based on Blockwise Sparse Principal Components. International Journal of Molecular Sciences. 2020; 21(21):8202. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218202
Chicago/Turabian StylePark, Mira, Doyoen Kim, Kwanyoung Moon, and Taesung Park. 2020. "Integrative Analysis of Multi-Omics Data Based on Blockwise Sparse Principal Components" International Journal of Molecular Sciences 21, no. 21: 8202. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218202
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.