Aspergillus flavus Exploits Maize Kernels Using an “Orphan” Secondary Metabolite Cluster


 , ,
, ,
Abstract
:1. Introduction
2. Results & Discussions
2.1. Phenotypic Characterization of A. flavus Mutant Strain
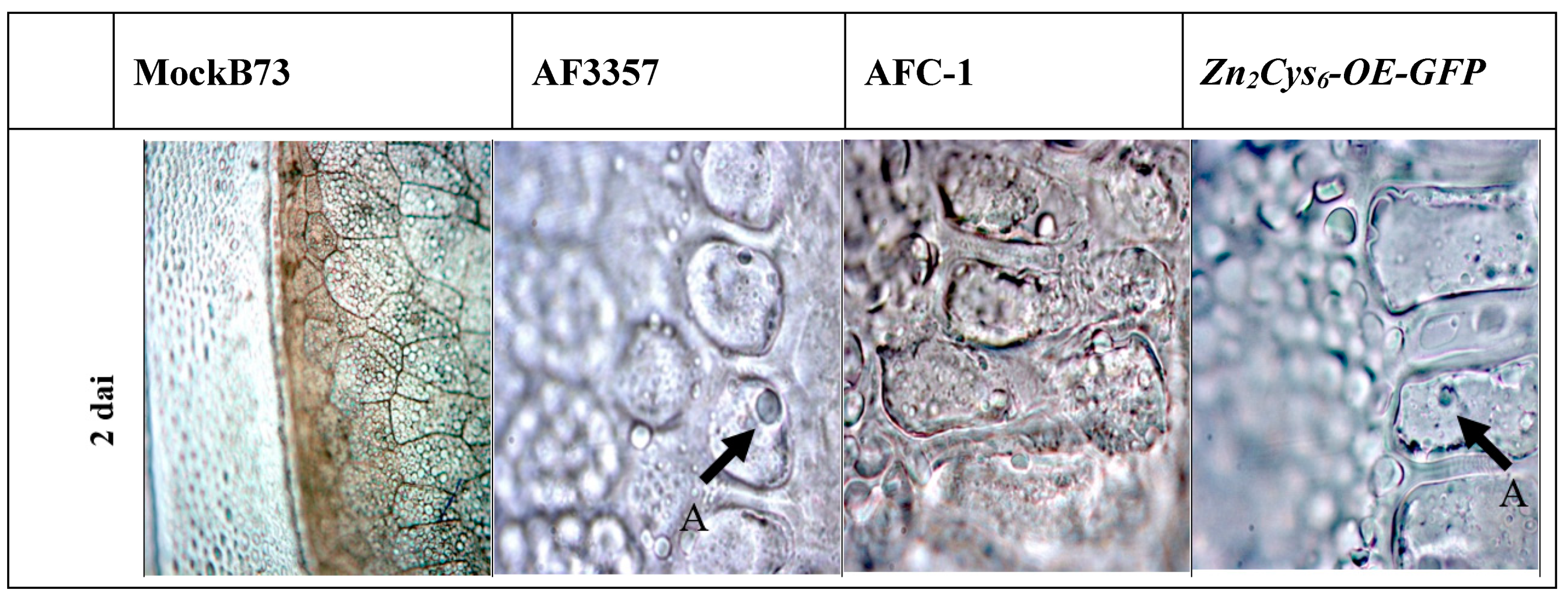
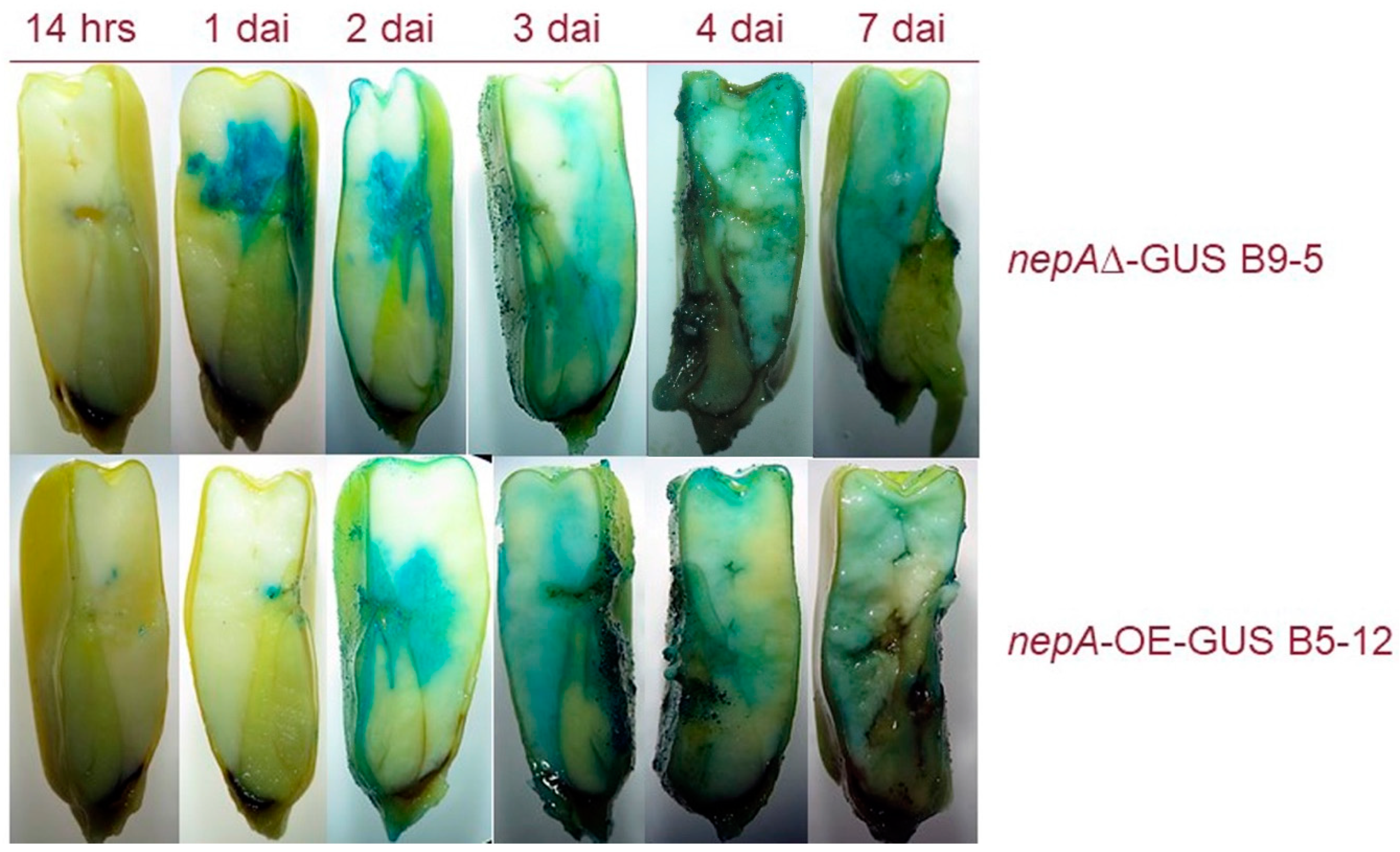
2.2. Colonisation of Maize Kernels by A. flavus AF3357, AFC-1 and Zn2Cys6-OE-GFP
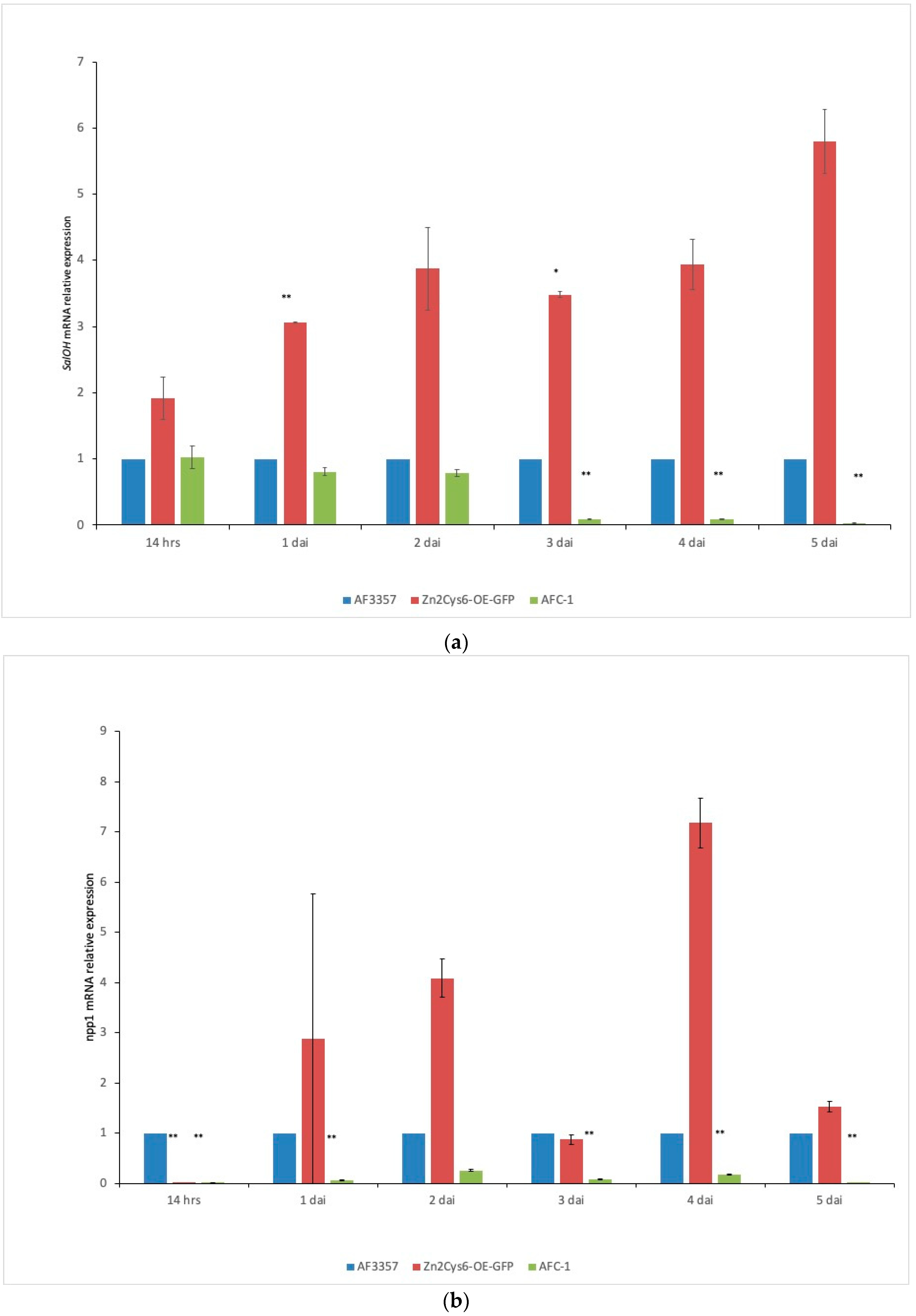
2.3. Expression of Genes Related to the Pathogenesis
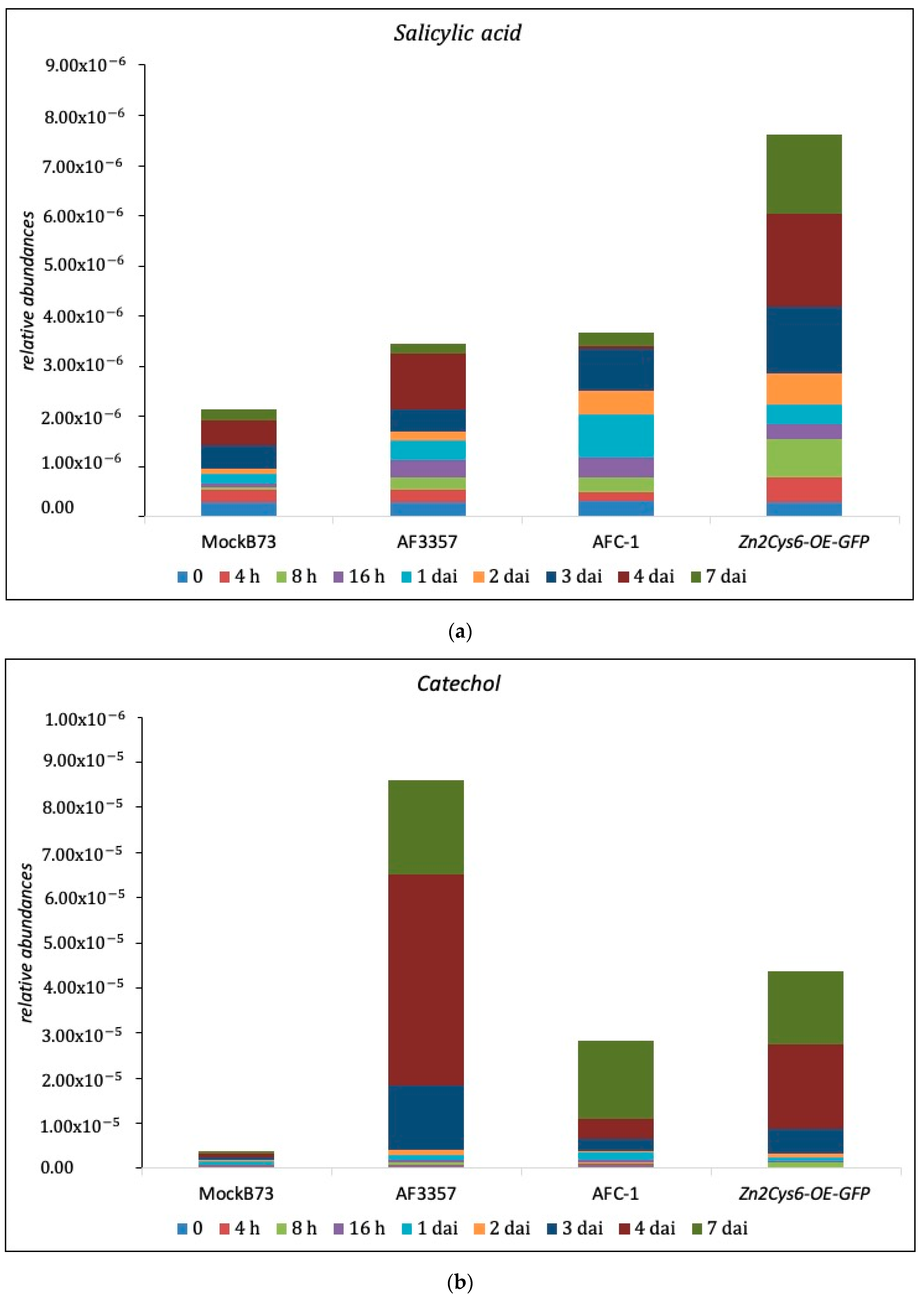
2.4. SA and Cathecol Accumulation in Maize-Infected Kernel
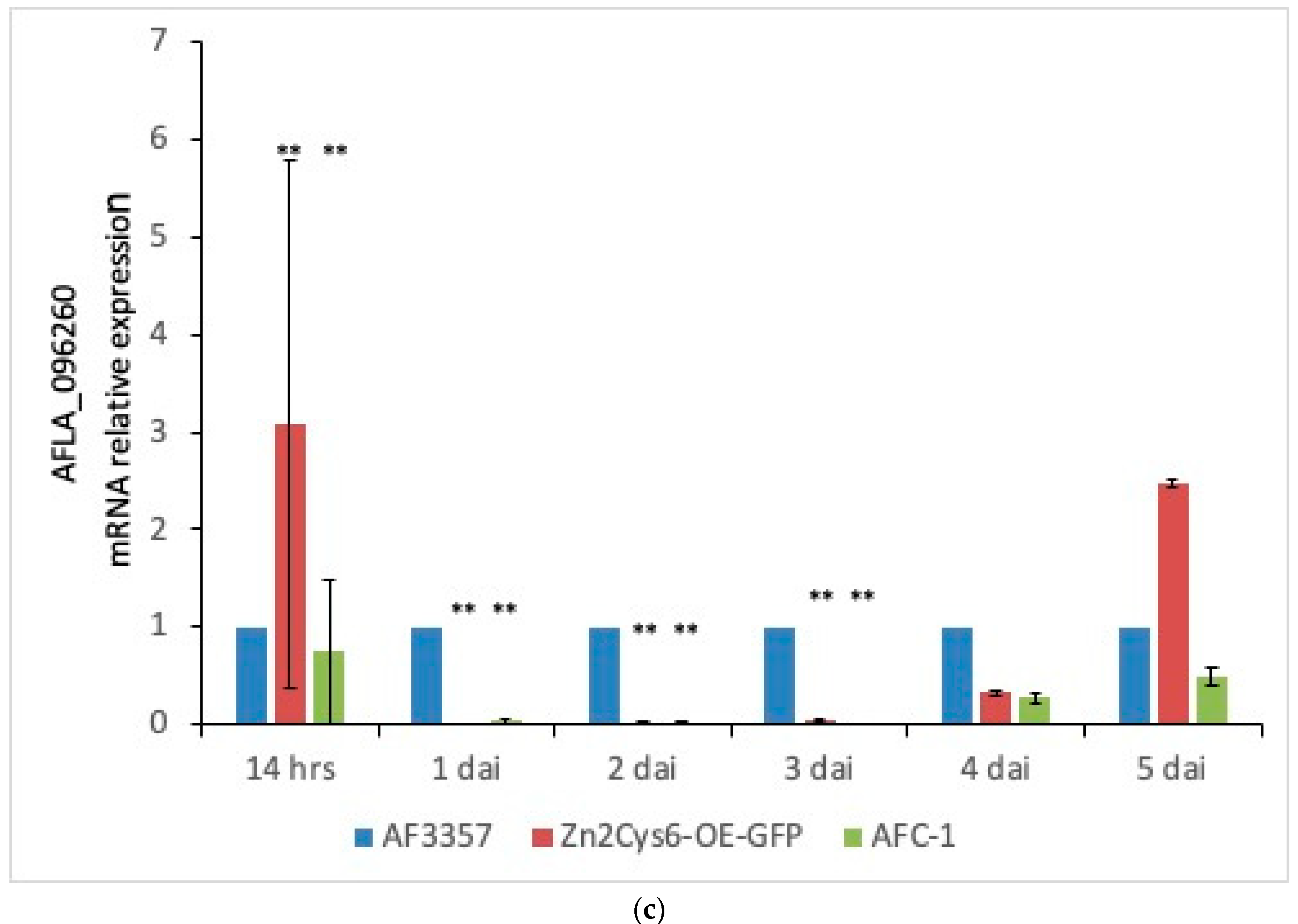
2.5. Npp1 Induces Necrotic Cell Death
3. Materials and methods
3.1. Aspergillus flavus Strains
3.2. Generation of Zn2Cys6∆ and Zn2Cys6-OE-GFP Strains
3.3. Zea Mays Crops
3.4. Gene Expression Analysis
3.5. Rutin Degradation
3.6. Salicylic Acid, Catechol, Rutin, Quercetin and Aflatoxin B1 Analysis by HPLC-MS/MS
3.7. Histological Protocol
3.8. Histochemical Assay
3.9. TUNEL Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | Aflatoxin |
| dai | Day After Inoculation |
| EFSA | European Food Safety Authority |
| GUS | β-Glucuronidase |
| HR | Hypersensitive Response |
| PCD | Programmed Cell Death |
| PPA | protocatechuoyl-phloroglucinolcarboxylic acid |
| ROS | Reactive Oxygen Species |
| SA | Salicylic Acid |
| SM | Secondary Metabolites |
| SMURF | Secondary Metabolite Unknown Regions Finder |
| TUNEL | Terminal Deoxynucleotidyl Transferase dUTP Nick end Labeling |
References
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, B.W. Biodiversity of Aspergillus section Flavi in the United States: A review. Food Addit. Contam. 2007, 24, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
- Dowd, P.F. Involvement of Arthropods in the Establishment of Mycotoxigenic Fungi under Field Conditions; Marcel Dekker: New York, NY, USA, 1998. [Google Scholar]
- Payne, G.A.; Brown, M.P. Genetics and physiology of aflatoxin biosynthesis. Annu. Rev. Phytopathol. 1998, 36, 329–362. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.Z.; Butrón, A.; Li, H.; Widstrom, N.W.; Lynch, R.E. Restriction fragment length polymorphism assessment of the heterogeneous nature of maize population GT-MAS: Gk and field evaluation of resistance to aflatoxin production by Aspergillus flavus. J. Food Prot. 2002, 65, 167–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raruang, Y.; Omolehin, O.; Hu, D.; Wei, Q.; Han, Z.-Q.; Rajasekaran, K.; Cary, J.W.; Wang, K.; Chen, Z.Y. Host induced gene silencing targeting Aspergillus flavus aflM reduced aflatoxin contamination in transgenic maize under field conditions. Front. Microbiol. 2020, 11, 754. [Google Scholar] [CrossRef]
- Gorman, D.P.; Kang, M.S. Preharvest Aflatoxin Contamination in Maize: Resistance and Genetics. Plant Breed. 1991, 107, 1–10. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.C.; et al. Risk assessment of aflatoxins in food. EFSA J. 2020, 18, e06040. [Google Scholar] [CrossRef]
- Ehrlich, K.C. Non-aflatoxigenic Aspergillus flavus to prevent aflatoxin contamination in crops: Advantages and limitations. Front. Microbiol. 2014, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, H.J.; Moretti, A.; Leggieri, M.C.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B 1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, A.M.; Cary, J.W. Association of fungal secondary metabolism and sclerotial biology. Front. Microbiol. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, P.; Gangurde, S.S.; Ortega-Beltran, A.; Kumar, R.; Parmar, S.; Sudini, H.K.; Lei, Y.; Ni, X.; Huai, D.; Fountain, J.C. Functional Biology and Molecular Mechanisms of Host-Pathogen Interactions for Aflatoxin Contamination in Groundnut (Arachis hypogaea L.) and Maize (Zea mays L.). Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Khaldi, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.H.; Fedorova, N.D. SMURF: Genomic mapping of fungal secondary metabolite clusters. Fungal Genet. Biol. 2010, 47, 736–741. [Google Scholar] [CrossRef] [Green Version]
- Georgianna, D.R.; Fedorova, N.D.; Burroughs, J.L.; Dolezal, A.L.; Bok, J.W.; Horowitz-Brown, S.; Woloshuk, C.P.; Yu, J.; Keller, N.P.; Payne, G.A. Beyond aflatoxin: Four distinct expression patterns and functional roles associated with Aspergillus flavus secondary metabolism gene clusters. Mol. Plant Pathol. 2010, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Punelli, M.; Scala, V.; Scarpari, M.; Uva, P.; Mentzen, W.I.; Dolezal, A.L.; Woloshuk, C.; Pinzari, F.; Fabbri, A.A. Genotypic and phenotypic versatility of Aspergillus flavus during maize exploitation. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Valdes, J.J.; Cameron, J.E.; Cole, R.J. Aflatrem: A tremorgenic mycotoxin with acute neurotoxic effects. Environ. Health Perspect. 1985, 62, 459–463. [Google Scholar] [CrossRef]
- Ehrlich, K.C.; Mack, B.M. Comparison of expression of secondary metabolite biosynthesis cluster genes in Aspergillus flavus, A. parasiticus, and A. oryzae. Toxins (Basel) 2014, 6, 1916–1928. [Google Scholar] [CrossRef] [PubMed]
- Keates, S.E.; Kostman, T.A.; Anderson, J.D.; Bailey, B.A. Altered gene expression in three plant species in response to treatment with Nep1, a fungal protein that causes necrosis. Plant Physiol. 2003, 132, 1610–1622. [Google Scholar] [CrossRef] [Green Version]
- Staats, M.; van Baarlen, P.; Schouten, A.; van Kan, J.A.L. Functional analysis of NLP genes from Botrytis elliptica. Mol. Plant Pathol. 2007, 8, 209–214. [Google Scholar] [CrossRef]
- Laluk, K.; Mengiste, T. Necrotroph attacks on plants: Wanton destruction or covert extortion? Arab. Book/Am. Soc. Plant Biol. 2010, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra, B.A.; Lohmar, J.M.; Satterlee, T.; McDonald, T.; Cary, J.W.; Calvo, A.M. The 14-3-3 protein homolog ArtA regulates development and secondary metabolism in the opportunistic plant pathogen Aspergillus flavus. Appl. Environ. Microbiol. 2018, 84, e02241-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.-K.; Ehrlich, K.C. Genome-wide analysis of the Zn (II) 2 Cys 6 zinc cluster-encoding gene family in Aspergillus flavus. Appl. Microbiol. Biotechnol. 2013, 97, 4289–4300. [Google Scholar] [CrossRef]
- Nobili, C.; D’Angeli, S.; Altamura, M.M.; Scala, V.; Fabbri, A.A.; Reverberi, M.; Fanelli, C. ROS and 9-oxylipins are correlated with deoxynivalenol accumulation in the germinating caryopses of Triticum aestivum after Fusarium graminearum infection. Eur. J. Plant Pathol. 2014, 139, 429–444. [Google Scholar] [CrossRef]
- Nayak, S.N.; Agarwal, G.; Pandey, M.K.; Sudini, H.K.; Jayale, A.S.; Purohit, S.; Desai, A.; Wan, L.; Guo, B.; Liao, B. Aspergillus flavus infection triggered immune responses and host-pathogen cross-talks in groundnut during in-vitro seed colonization. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Qi, P.-F.; Zhang, Y.-Z.; Liu, C.-H.; Chen, Q.; Guo, Z.-R.; Wang, Y.; Xu, B.-J.; Jiang, Y.-F.; Zheng, T.; Gong, X. Functional analysis of FgNahG clarifies the contribution of salicylic acid to wheat (Triticum aestivum) resistance against Fusarium head blight. Toxins (Basel) 2019, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Hao, G.; Naumann, T.A.; Vaughan, M.M.; McCormick, S.; Usgaard, T.; Kelly, A.; Ward, T.J. Characterization of a Fusarium graminearum Salicylate Hydroxylase. Front. Microbiol. 2019, 9, 3219. [Google Scholar] [CrossRef]
- Zhou, B.-J.; Jia, P.-S.; Gao, F.; Guo, H.-S. Molecular characterization and functional analysis of a necrosis-and ethylene-inducing, protein-encoding gene family from Verticillium dahliae. Mol. Plant-Microbe Interact. 2012, 25, 964–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, B.-Z.; Zhu, X.-P.; Fu, L.; Lv, R.-F.; Storey, D.; Tooley, P.; Zhang, X.-G. Characterization of necrosis-inducing NLP proteins in Phytophthora capsici. BMC Plant Biol. 2014, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Cobos, R.; Calvo-Peña, C.; Álvarez-Pérez, J.M.; Ibáñez, A.; Diez-Galán, A.; González-García, S.; García-Angulo, P.; Acebes, J.L.; Coque, J.J.R. Necrotic and cytolytic activity on grapevine leaves produced by Nep1-like proteins of Diplodia seriata. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Adv. Res. 2006, 661, 23–67. [Google Scholar]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines (Basel Switz.) 2018, 5, 93. [Google Scholar] [CrossRef]
- Chitarrini, G.; Nobili, C.; Pinzari, F.; Antonini, A.; De Rossi, P.; Del Fiore, A.; Procacci, S.; Tolaini, V.; Scala, V.; Scarpari, M. Buckwheat achenes antioxidant profile modulates Aspergillus flavus growth and aflatoxin production. Int. J. Food Microbiol. 2014, 189, 1–10. [Google Scholar] [CrossRef]
- Asters, M.C.; Williams, W.P.; Perkins, A.D.; Mylroie, J.E.; Windham, G.L.; Shan, X. Relating significance and relations of differentially expressed genes in response to Aspergillus flavus infection in maize. Sci. Rep. 2014, 4, 4815. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P.; Adams, T.H. Analysis of a mycotoxin gene cluster in Aspergillus nidulans. SAAS Bull. Biochem. Biotechnol. 1995, 8, 14–21. [Google Scholar]
- Keller, N.P.; Butchko, R.A.E.; Sarr, B.; Phillips, T.D. A visual pattern of mycotoxin production in maize kernels by Aspergillus spp. Phytopathology 1994, 84, 483–488. [Google Scholar] [CrossRef]
- Bhatnagar, D.; Ehrlich, K.C.; Cleveland, T.E. Molecular genetic analysis and regulation of aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2003, 61, 83–93. [Google Scholar] [CrossRef]
- Payne, G.A.; Yu, J.; Nierman, W.C.; Machida, M.; Bhatnagar, D.; Cleveland, T.E.; Dean, R.A. A First Glance into the Genome Sequence of Aspergillus fl avus. In The Aspergilli; CRC Press: Boca Raton, FL, USA, 2007; pp. 35–44. [Google Scholar]
- Du, W.; Huang, Z.; Flaherty, J.E.; Wells, K.; Payne, G.A. Green fluorescent protein as a reporter to monitor gene expression and food colonization by Aspergillus flavus. Appl. Environ. Microbiol. 1999, 65, 834–836. [Google Scholar] [CrossRef] [Green Version]
- He, Z.-M.; Price, M.S.; OBrian, G.R.; Georgianna, D.R.; Payne, G.A. Improved protocols for functional analysis in the pathogenic fungus Aspergillus flavus. BMC Microbiol. 2007, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- Scala, V.; Pietricola, C.; Farina, V.; Beccaccioli, M.; Zjalic, S.; Quaranta, F.; Fornara, M.; Zaccaria, M.; Momeni, B.; Reverberi, M. Tramesan Elicits Durum Wheat Defense against the Septoria Disease Complex. Biomolecules 2020, 10, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, E.M.; Howlett, B.J. Secondary metabolism: Regulation and role in fungal biology. Curr. Opin. Microbiol. 2008, 11, 481–487. [Google Scholar] [CrossRef]
- OBrian, G.R.; Fakhoury, A.M.; Payne, G.A. Identification of genes differentially expressed during aflatoxin biosynthesis in Aspergillus flavus and Aspergillus parasiticus. Fungal Genet. Biol. 2003, 39, 118–127. [Google Scholar] [CrossRef]
- Caceres, I.; Khoury, A.A.; Khoury, R.E.; Lorber, S.; Oswald, I.P.; Khoury, A.E.; Atoui, A.; Puel, O.; Bailly, J.D. Aflatoxin Biosynthesis and Genetic Regulation: A Review. Toxins (Basel) 2020, 12, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Growth Medium | Characteristics | References |
|---|---|---|---|
| Aspergillus flavus 3357 | Czapek Dox Broth or Agar (CD, Difco) | Wild type | Payne et al. (2007) NRRL [39] |
| AFC-1 | Czapek Dox Broth or Agar (CD, Difco) with Uracil 1.122 g/L and 0.26 g/L of Arginine | Mutant (−pyrG, −argD) requires uracil and arginine | Georgianna et al. (2010) [16] |
| Zn2Cys6∆ | Czapek Dox Broth or Agar (CD, Difco) with Uracil 1.122 g/L | auxotroph for uracil in which the transcription factor Zn2Cys6 has been deleted; it derives from AFC-1 strain with arginine auxotrophy restored | This study |
| Zn2Cys6-OE-GFP | Czapek Dox Broth or Agar (CD, Difco) with Uracil 1.122 g/L | auxotroph for uracil in which the transcription factor Zn2Cys6 is overexpressed; it derives from AFC-1 strain with arginine auxotrophy restored | This study |
| nepA-OE-GUS B5-12 | Potato Dextrose Agar (PDA, Difco) | nepA gene over-expressing strain | G.A. Payne (not published) |
| nepA∆-GUS B9-5 | Potato Dextrose Agar (PDA, Difco) | nepA gene knock-out strain | G.A. Payne (not published) |
| Compound | Precursor Ion | Product Ion | Fragmentor (V) | CE (eV) | Polarity |
|---|---|---|---|---|---|
| SA | 137.2 | 92.9, 64.8 | 135 | 20 | [M-H]− |
| Catechol | 109.1 | 53.1 | 135 | 20 | [M-H]− |
| NAA | 245 | 180.8 | 100 | 16 | [M-H]− |
| Quercetin | 303.1 | 152.9 | 100 | 37 | [M-H]+ |
| PPA | 291.3 | 129.1, 112.8 | 90 | 38 | [M-H]+ |
| Chlorogenic acid | 353.31 | 191.2 | 135 | 10 | [M-H]− |
| Rutin | 609.2 | 273.1 | 100 | 56 | [M-H]− |
| Aflatoxin B1 | 313.1 | 241.1 | 135 | 38 | [M-H]+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antiga, L.; La Starza, S.R.; Miccoli, C.; D’Angeli, S.; Scala, V.; Zaccaria, M.; Shu, X.; Obrian, G.; Beccaccioli, M.; Payne, G.A.; et al. Aspergillus flavus Exploits Maize Kernels Using an “Orphan” Secondary Metabolite Cluster. Int. J. Mol. Sci. 2020, 21, 8213. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218213
Antiga L, La Starza SR, Miccoli C, D’Angeli S, Scala V, Zaccaria M, Shu X, Obrian G, Beccaccioli M, Payne GA, et al. Aspergillus flavus Exploits Maize Kernels Using an “Orphan” Secondary Metabolite Cluster. International Journal of Molecular Sciences. 2020; 21(21):8213. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218213
Chicago/Turabian StyleAntiga, Ludovica, Sonia Roberta La Starza, Cecilia Miccoli, Simone D’Angeli, Valeria Scala, Marco Zaccaria, Xiaomei Shu, Gregory Obrian, Marzia Beccaccioli, Gary A. Payne, and et al. 2020. "Aspergillus flavus Exploits Maize Kernels Using an “Orphan” Secondary Metabolite Cluster" International Journal of Molecular Sciences 21, no. 21: 8213. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218213