Letrozole Suppresses the Fusion of Osteoclast Precursors through Inhibition of p38-Mediated DC-STAMP Pathway



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
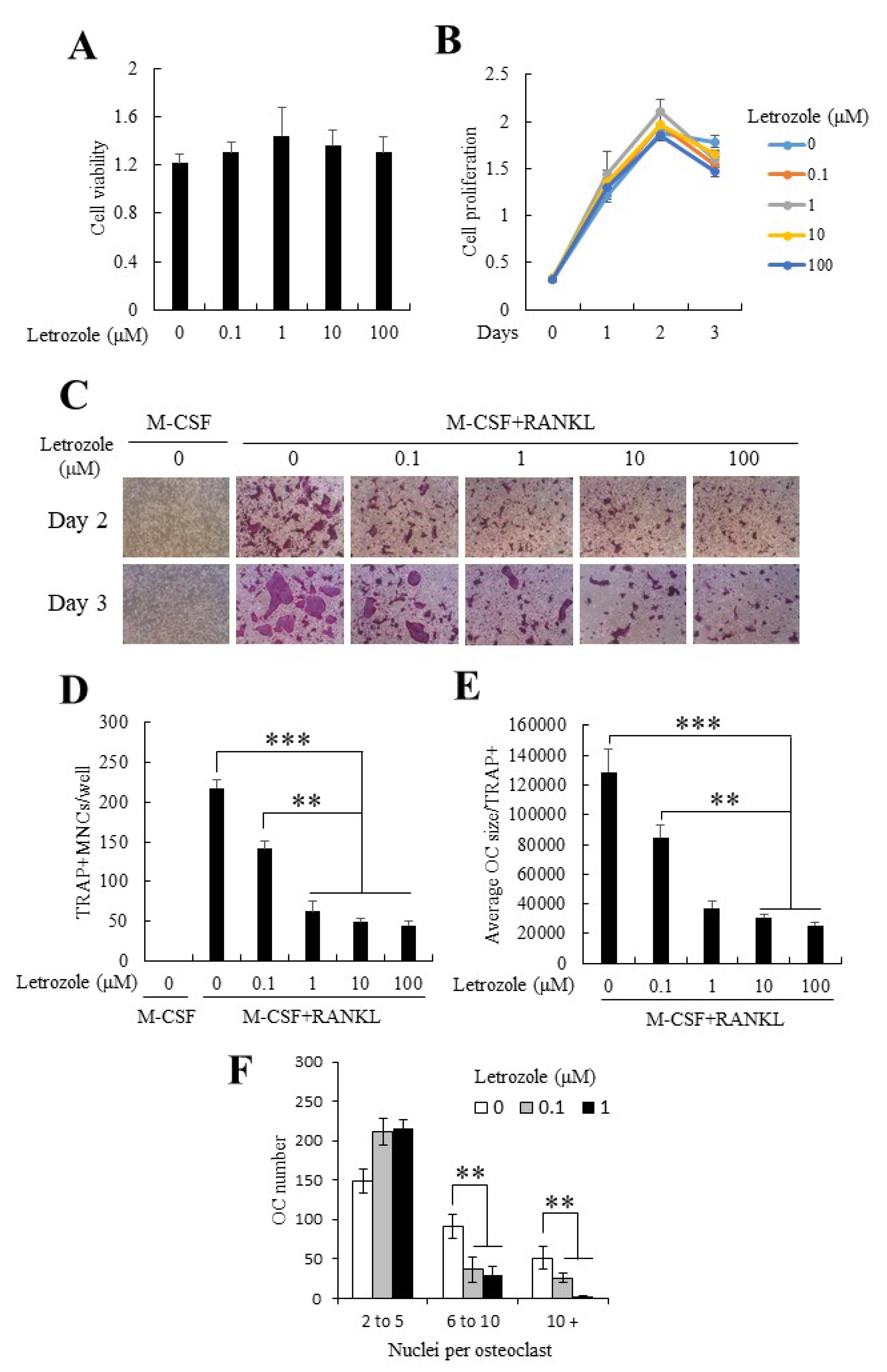
2.1. Letrozole Demonstrated No Cytotoxicity on Mouse Bone Marrow-Derived Macrophages (BMMs) and Did Not Alter BMM Proliferation
2.2. Letrozole Inhibited RANKL-Induced Osteoclastogenesis of BMMs
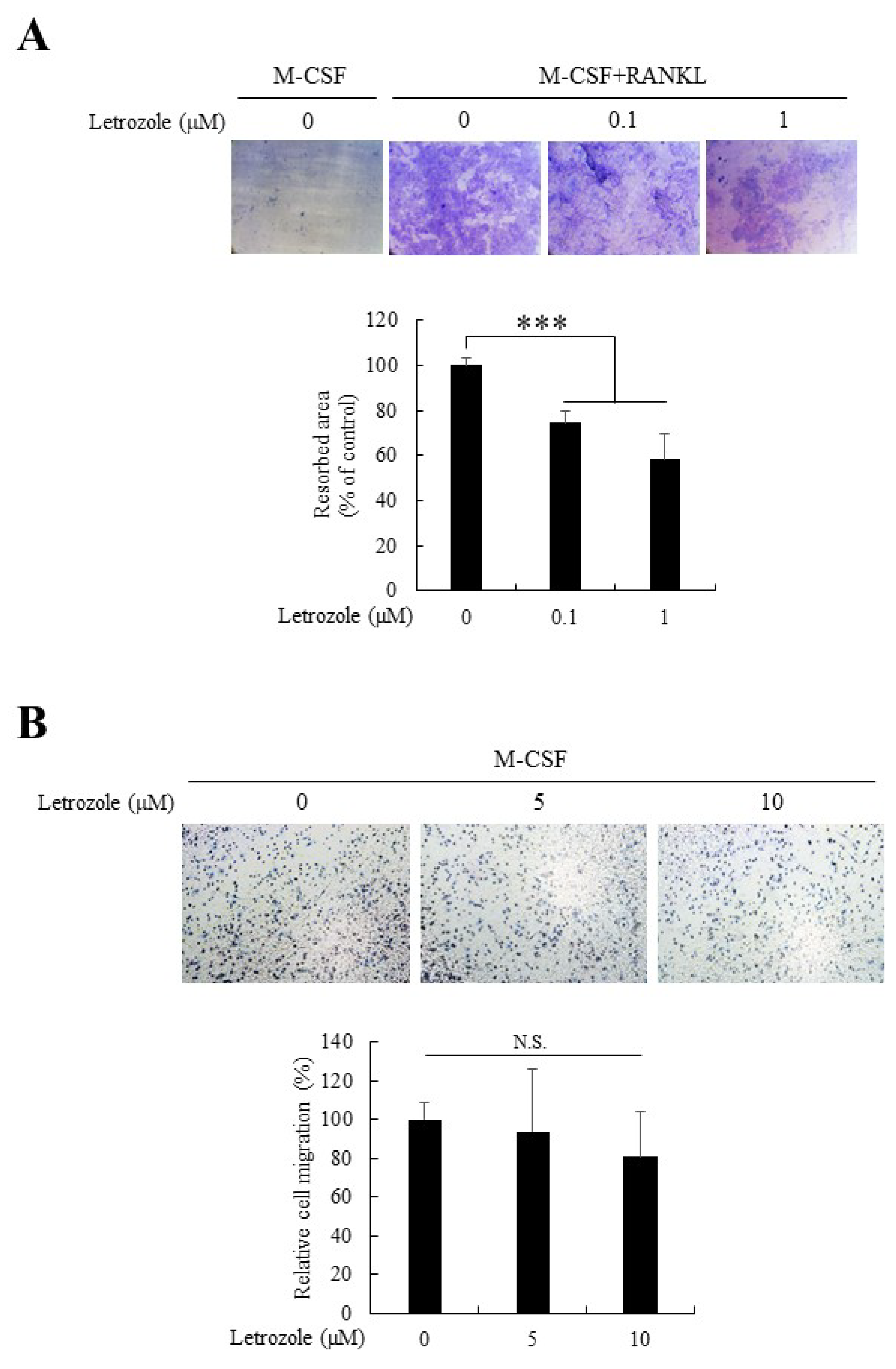
2.3. Letrozole Decreased Osteoclastic Bone Resorption But Did Not Affect Cell Migration
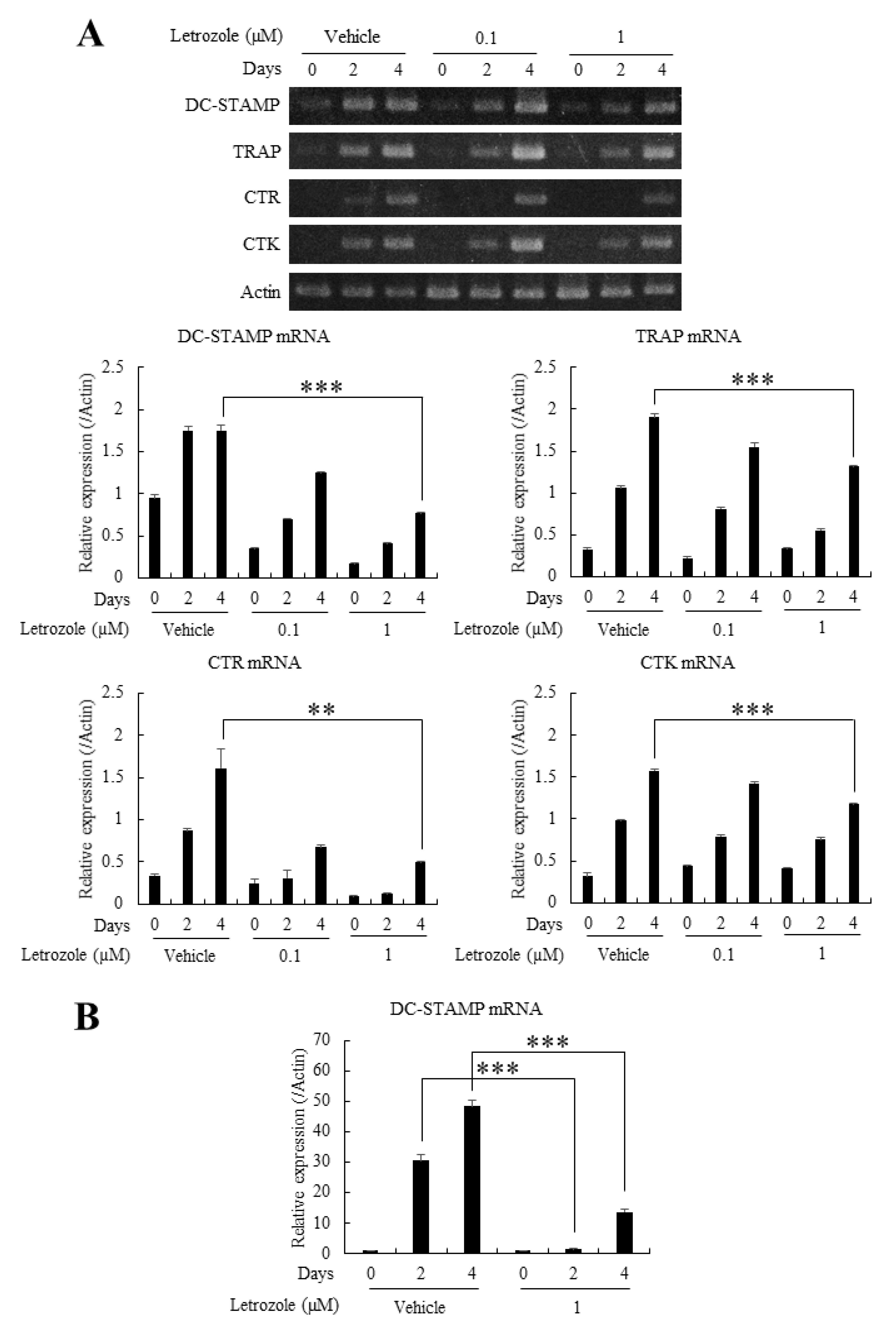
2.4. Letrozole Reduced the mRNA Expression of TRAP, CTR, CTK, and DC-STAMP
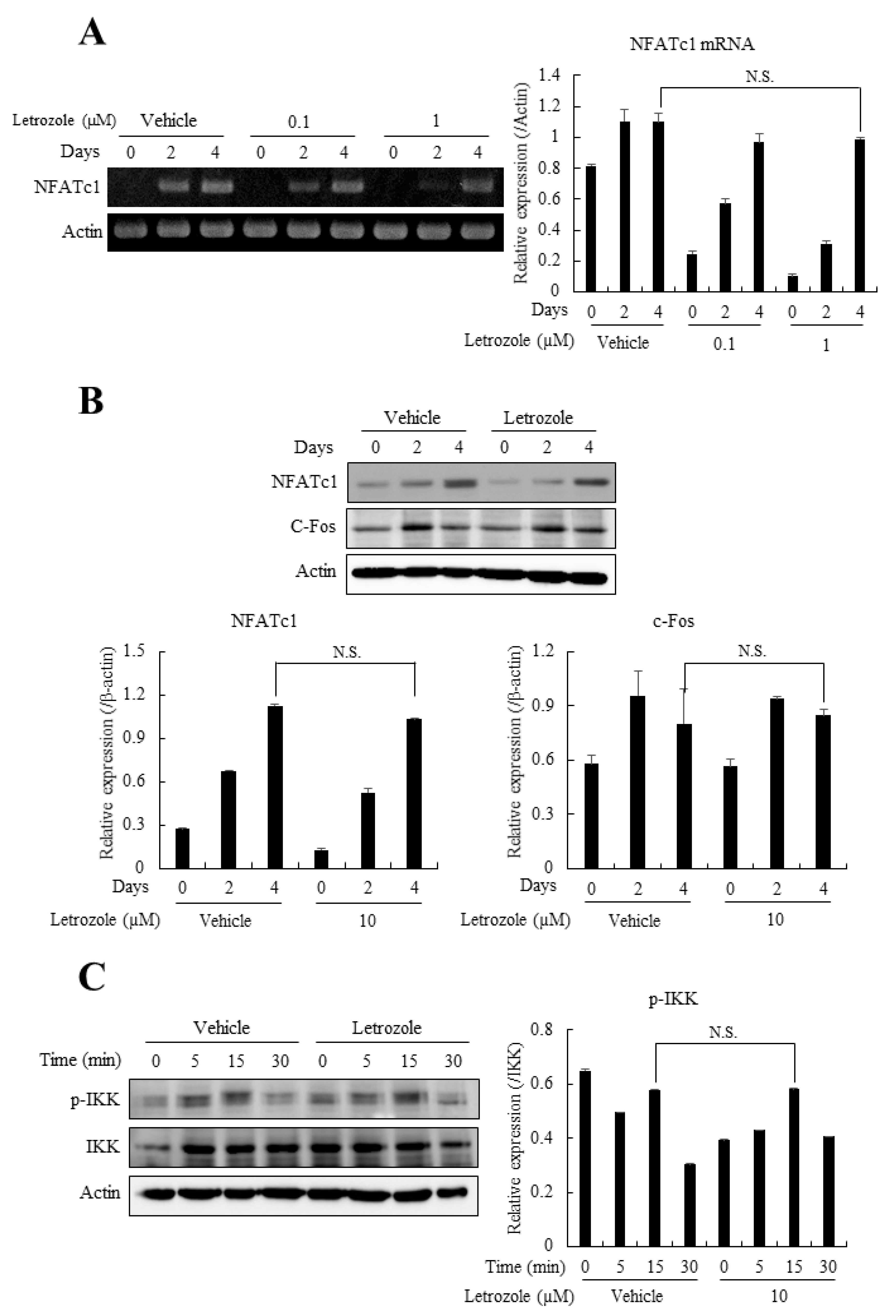
2.5. Letrozole Did Not Affect the Expression of Osteoclastogenic Transcription Factors, NFATc1, c-Fos, or the Activation of NF-κB
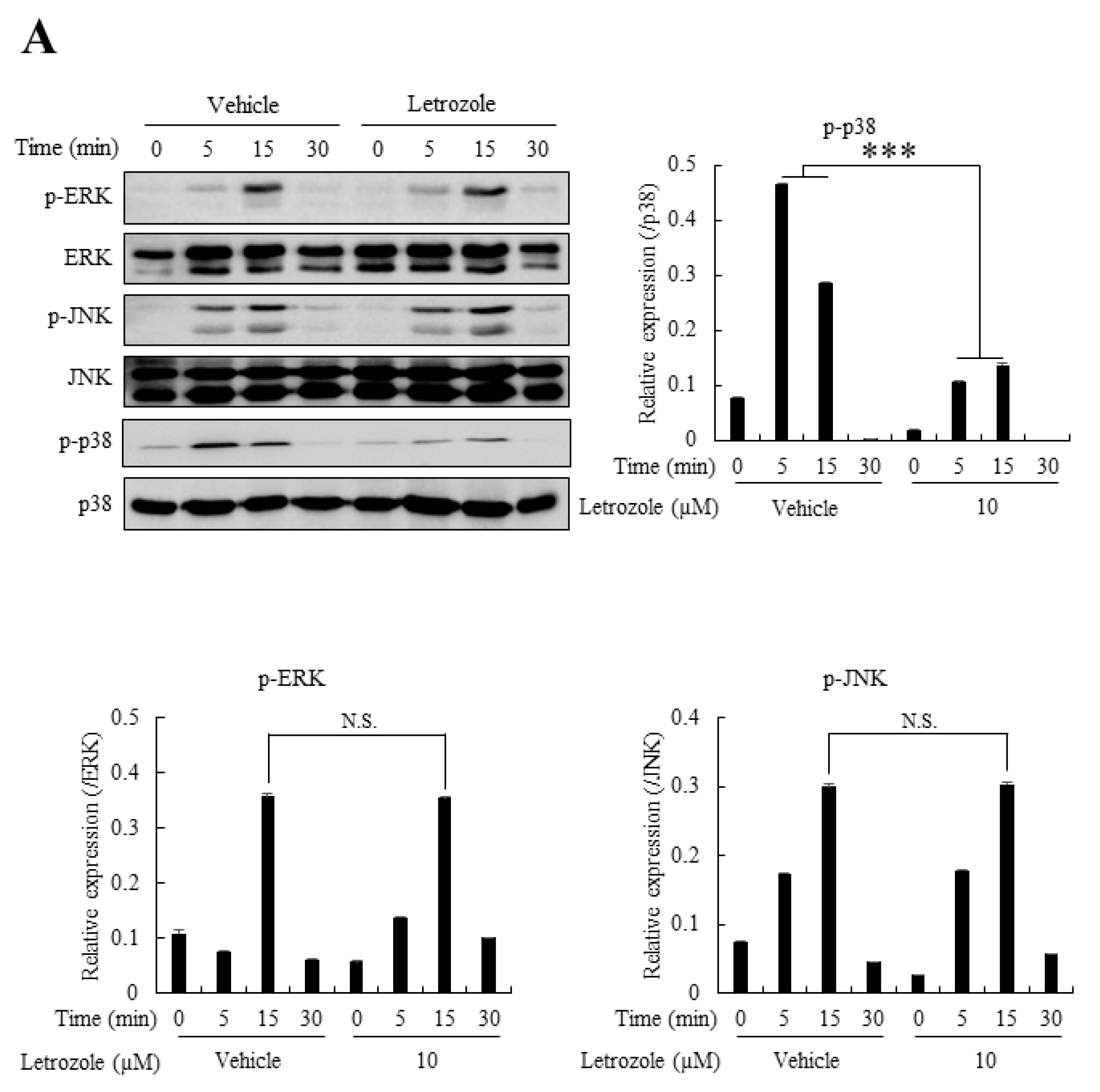
2.6. Letrozole Diminished p38 Activation in RANKL-Stimulated BMMs
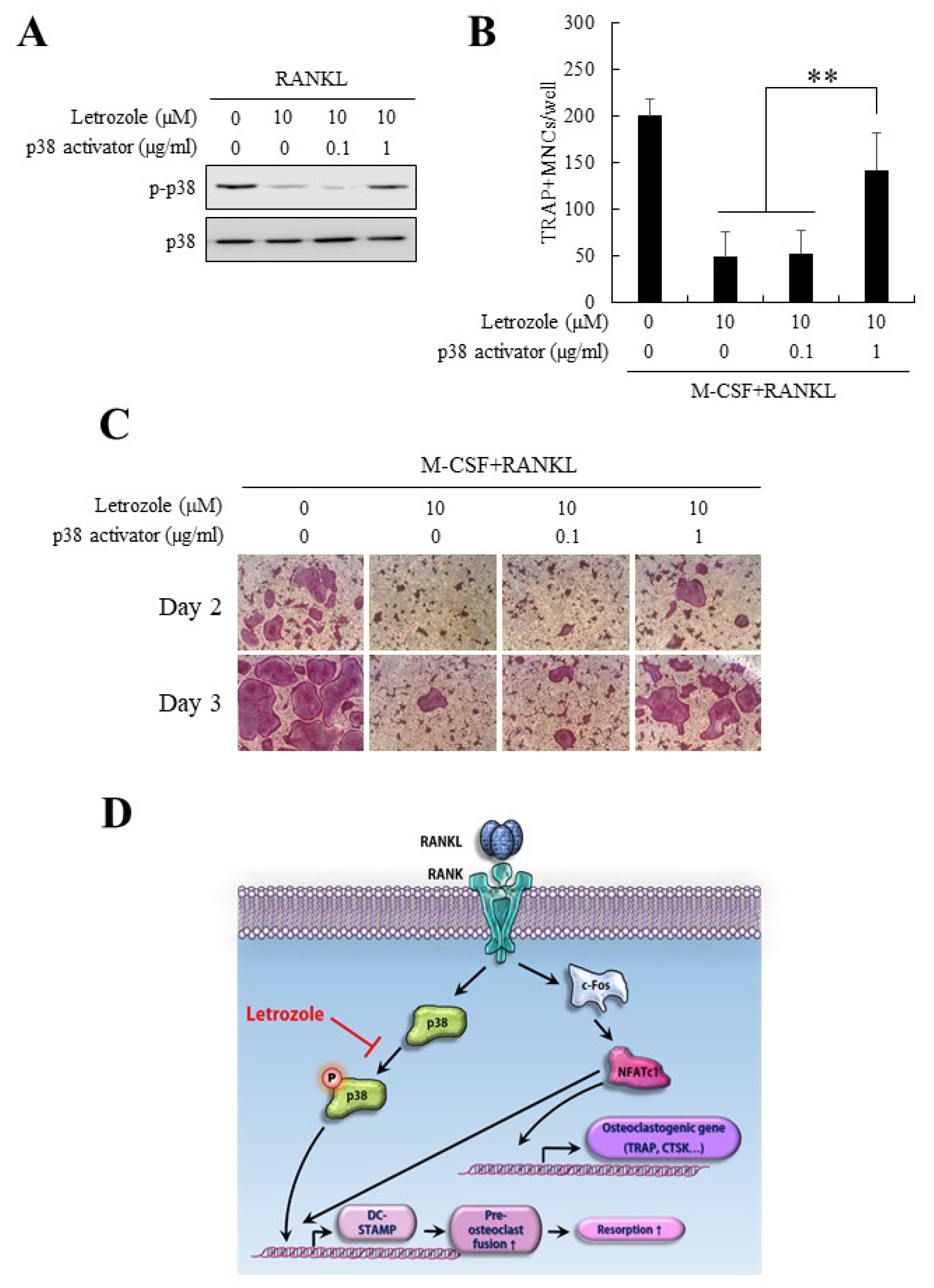
2.7. Anti-Osteoclastogenic Effect of Letrozole Was Abolished by p38 Activation
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. In Vitro Osteoclastogenesis
4.3. MTT Assay
4.4. Resorption Pit Formation Assay
4.5. Trans-Well Migration Assay
4.6. RT-PCR and Quantitative Real-Time PCR Analyses
4.7. Western Blotting
4.8. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Teitelbaum, S.L.; Ross, F.P. Genetic regulation of osteoclast development and function. Nat. Rev. Genet. 2003, 4, 638–649. [Google Scholar] [CrossRef]
- Karsenty, G.; Kronenberg, H.M.; Settembre, C. Genetic control of bone formation. Annu. Rev. Cell Dev. Biol. 2009, 25, 629–648. [Google Scholar] [CrossRef] [PubMed]
- Telebaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Darnay, B.G.; Moore, J.; Aggarwal, B.B. Activation of NF-kappaB by RANK requires tumor necrosis factor receptor-associate factor (TRAF) 6 and NF-kappaB-inducing kinase. Identification of a novel TRAF6 interaction motif. J. Biol. Chem. 1999, 274, 7724–7731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Z.H.; Kim, H.H. Signal transduction by receptor activator of nuclear factor kappa B in osteoclasts. Biochem. Biophys. Res. Commun. 2003, 305, 211–214. [Google Scholar] [CrossRef]
- Takako, N.K.; Hiroshi, T. Ca2+ -NFATc1 signaling is an essential axis of osteoclast differentiation. Annu. Rev. Immunol. 2009, 231, 241–256. [Google Scholar]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Dou, C.E.; Xu, J.; Dong, S. DC-STAMP, the key fusion-mediating molecule in osteoclastogenesis. J. Cell Physiol. 2014, 229, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.H.; Ritchlin, C.T. DC-STAMP: A key regulator in osteoclast differentiation. J. Cell Physiol. 2016, 231, 2402–2407. [Google Scholar] [CrossRef] [Green Version]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, L.C.; Khosla, S.; Dunstan, C.R.; Lacey, D.L.; Boyle, W.J.; Riggs, B.L. The roles of osteoprotegerin and osteoprotegerin ligand in the paracrine regulation of bone resorption. J. Bone Miner. Res. 2000, 15, 2–12. [Google Scholar] [CrossRef]
- Shevde, N.K.; Bendixen, A.C.; Dienger, K.M.; Pike, J.W. Estrogens suppress RANK ligand-induced osteoclast differentiation via a stromal cell independent mechanism involving c-Jun repression. Proc. Natl. Acad. Sci. USA 2000, 97, 7829–7834. [Google Scholar] [CrossRef] [Green Version]
- Cenci, S.; Weitzmann, M.N.; Roggia, C.; Namba, N.; Novack, D.; Woodring, J.; Pacifici, R. Estrogen deficiency induces bone loss by enhancing T cell production of TNF-α. J. Clin. Investig. 2000, 106, 1229–1237. [Google Scholar] [CrossRef]
- Yagi, M.; Miyamoto, T.; Sawatani, Y.; Iwamoto, K.; Hosogane, N.; Fujita, N.; Morita, K.; Ninomiya, K.; Suzuki, T.; Miyamoto, K.; et al. DC-STAMP is essential for cell-cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 2005, 202, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.A.; Schwarz, E.; Li, D.; Xu, Y.; Sheu, T.R.; Li, J.; Bently, K.M.; Feng, C.; Wang, B.; Wang, J.C.; et al. Dendritic Cell-Specific Transmembrane Protein (DC-STAMP) regulates osteoclast differentiation via the Ca2+/NFATc1 axis. J. Cell. Physiol. 2016, 232, 2538–2549. [Google Scholar] [CrossRef]
- Choi, J.W.; Choi, S.Y.; Lee, S.Y.; Lee, J.Y.; Kim, H.S.; Lee, S.Y.; Lee, N.K. Caffeine enhances osteoclast differentiation and maturation through p38 MAP kinase/Mitf and DC-STAMP/CtsK and TRAP pathway. Cell Signal 2013, 25, 1222–1227. [Google Scholar] [CrossRef]
- Fujita, K.; Iwasaki, M.; Ochi, H.; Fukuda, T.; Ma, C.; Miyamoto, T.; Takitani, K.; Koga, T.N.; Sunamura, S.; Kodama, T.; et al. Vitamin E decreases bone mass by stimulating osteoclast fusion. Nat. Med. 2012, 18, 589–594. [Google Scholar] [CrossRef]
- Xiong, W.; Kojic, L.Z.; Zhang, L.; Prasad, S.S.; Douglas, R.; Wang, Y.; Cynader, M.S. Anisomycin activates p38 MAP kinase to induce LTD in mouse primary visual cortex. Brain Res. 2006, 1085, 68–76. [Google Scholar] [CrossRef]
- Hadwen, J.; Farooq, F.; Witherspoon, L.; Schock, S.; Mongeon, K.; MacKenzie, A. Anisomycin Activates Utrophin Upregulation Through a p38 Signaling Pathway. Clin. Transl. Sci. 2018, 5, 506–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukita, T.; Wada, N.; Kukita, A.; Kakimoto, T.; Sandra, F.; Toh, K.; Nagata, K.; Iijima, T.; Horiuchi, M.; Matsusaki, H.; et al. RANKL-induced DC-STAMP is essential for osteoclastogenesis. J. Exp. Med. 2004, 200, 941–946. [Google Scholar] [CrossRef] [Green Version]
- Nishida, T.; Emura, K.; Kubota, S.; Lyons, K.M.; Takigawa, M. CCN family 2/connective tissue growth factor (CCN2/CTGF) promotes osteoclastogenesis via induction of and interaction with dendritic cell-specific transmembrane protein (DC-STAMP). J. Bone Miner. Res. 2011, 26, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Bae, H.S.; Yoon, W.J.; Woo, K.M.; Baek, J.H.; Kim, H.H.; Uchida, T.; Ryoo, H.M. Pin1 regulates osteoclast fusion through suppression of the master regulator of cell fusion DC-STAMP. J. Cell Physiol. 2014, 229, 2166–2174. [Google Scholar] [CrossRef]
- Pang, M.; Rodríguez-Gonzalez, M.; Hernandez, M.; Recinos, C.C.; Seldeen, K.L.; Troen, B.R. AP-1 and Mitf interact with NFATc1 to stimulate cathepsin K promoter activity in osteoclast precursors. J. Cell. Biochem. 2019, 120, 12382–12392. [Google Scholar] [CrossRef]
- Lu, S.Y.; Li, M.; Lin, Y.L. Mitf regulates osteoclastogenesis by modulating NFATc1 activity. Exp. Cell Res. 2014, 328, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.M.; Bronisz, A.; Hu, R.; Patel, K.; Mansky, K.C.; Sif, S.; Ostrowski, M.C. MITF and PU.1 recruit p38 MAPK and NFATc1 to target genes during osteoclast differentiation. J. Biol. Chem. 2007, 282, 15921–15929. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.; Curran, M.P.; Perry, C.M. Letrozole: A review of its use in postmenopausal women with breast cancer. Drugs 2004, 64, 1213–1230. [Google Scholar] [CrossRef]
- Henriksen, K.; Karsdal, M.A.; Martin, T.J. Osteoclast-derived coupling factors in bone remodeling. Calcif. Tissue Int. 2014, 94, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Irie, N. Osteoclast-osteoblast communication. Arch. Biochem. Biophys. 2008, 473, 201–209. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Yoo, J.H.; Lee, J.H.; Lee, Y.K.; Bae, M.K.; Kim, Y.D.; Kim, H.J. Connective tissue growth factor (CTGF) regulates the fusion of osteoclast precursors by inhibiting Bcl6 in periodontitis. Int. J. Med. Sci. 2020, 17, 647–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Yoo, J.H.; Lee, Y.K.; Bae, M.K.; Kim, H.J. Calcium-Phosphate Crystals Promote RANKL Expression via the Downregulation of DUSP1. Mol. Cells 2019, 42, 183–188. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Seong, H.-S.; Choi, Y.; Heo, S.C.; Kim, Y.-D. Letrozole Suppresses the Fusion of Osteoclast Precursors through Inhibition of p38-Mediated DC-STAMP Pathway. Int. J. Mol. Sci. 2020, 21, 8396. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218396
Kim HJ, Seong H-S, Choi Y, Heo SC, Kim Y-D. Letrozole Suppresses the Fusion of Osteoclast Precursors through Inhibition of p38-Mediated DC-STAMP Pathway. International Journal of Molecular Sciences. 2020; 21(21):8396. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218396
Chicago/Turabian StyleKim, Hyung Joon, Hwa-Sik Seong, YunJeong Choi, Soon Chul Heo, and Yong-Deok Kim. 2020. "Letrozole Suppresses the Fusion of Osteoclast Precursors through Inhibition of p38-Mediated DC-STAMP Pathway" International Journal of Molecular Sciences 21, no. 21: 8396. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218396