CDK5RAP3, a Novel Nucleoplasmic Shuttle, Deeply Regulates HSF1-Mediated Heat Stress Response and Protects Mammary Epithelial Cells from Heat Injury




Abstract
:1. Introduction
2. Results
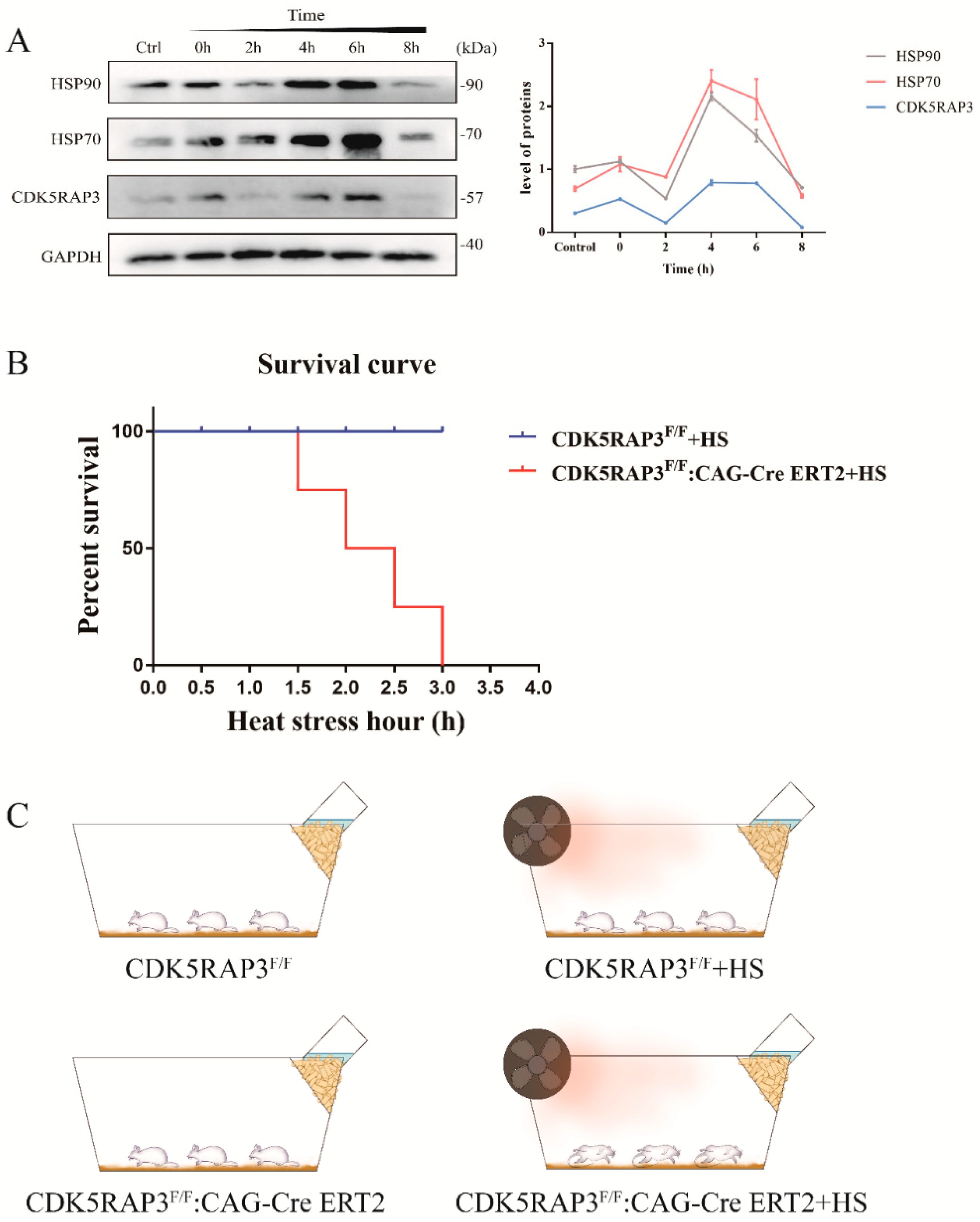
2.1. CDK5RAP3 and HSPs Showed Similar Expression Trend in BMECs after Exposure to Heat Stress
2.2. The Deletion of the CDK5RAP3 Gene Accelerated the Death of Mice
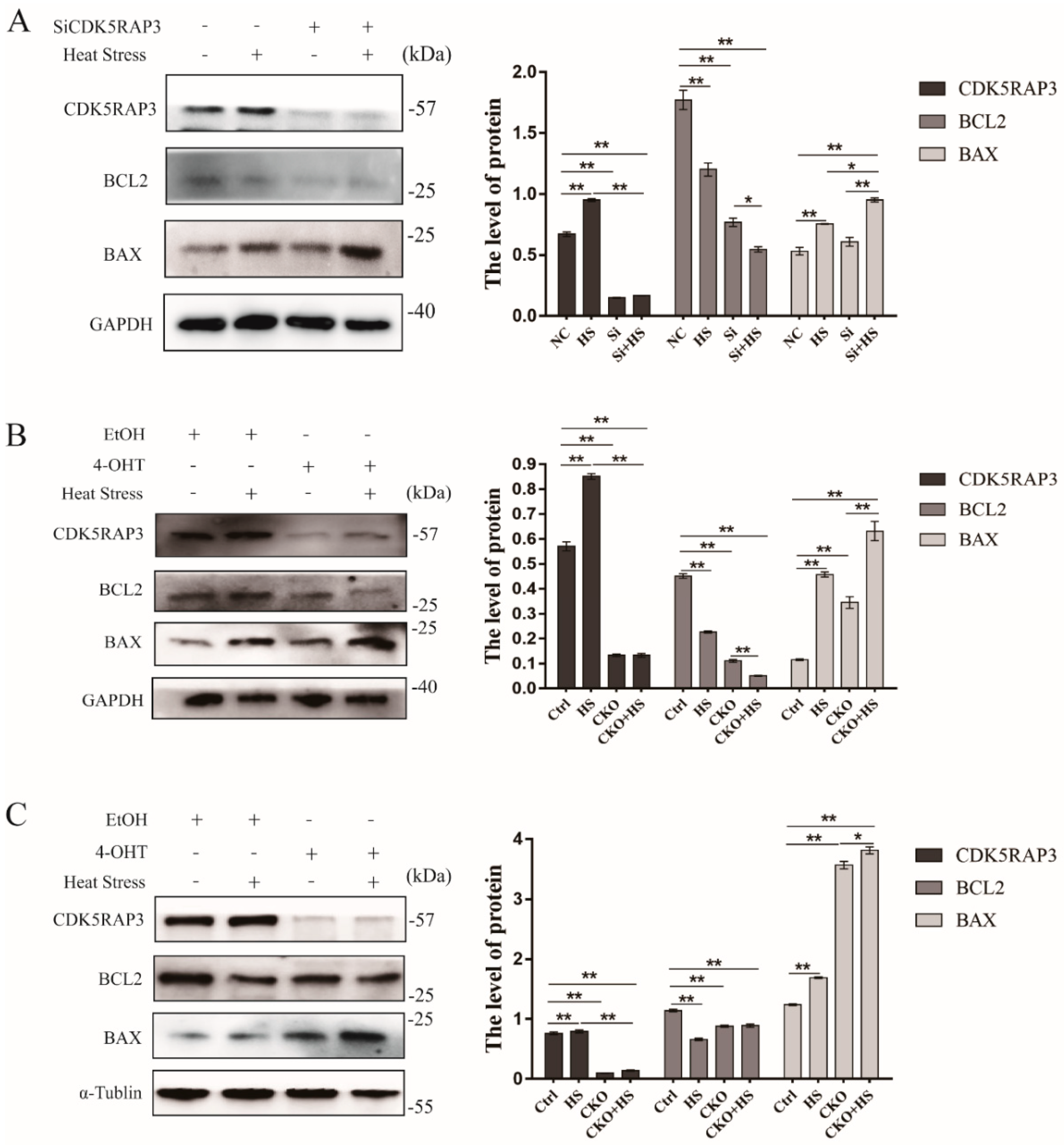
2.3. CDK5RAP3 Induces Apoptosis Due to the Heat Stress
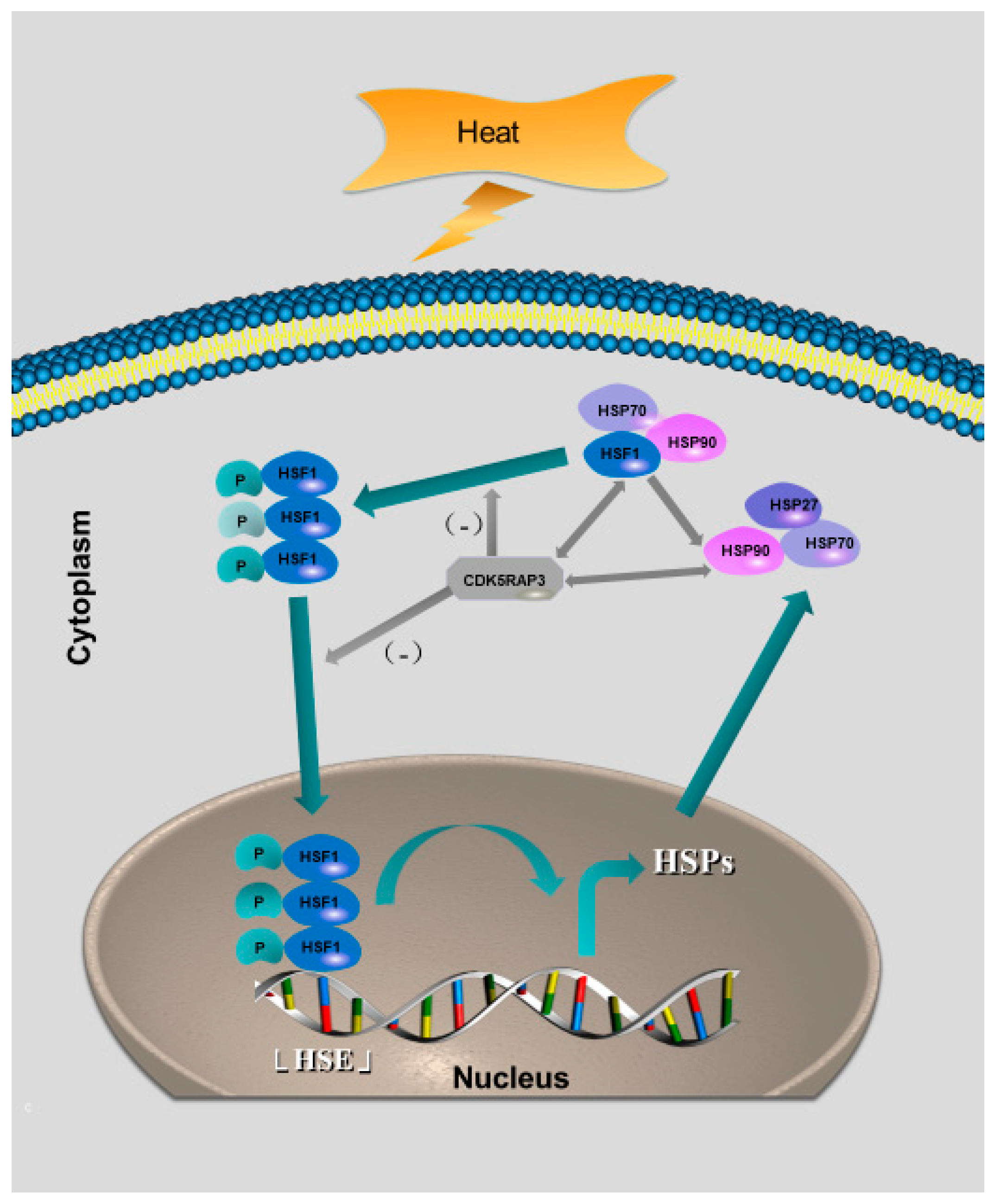
2.4. Regulation of the Heat Shock Factor 1-Mediated Stress Response Pathway by CDK5RAP3
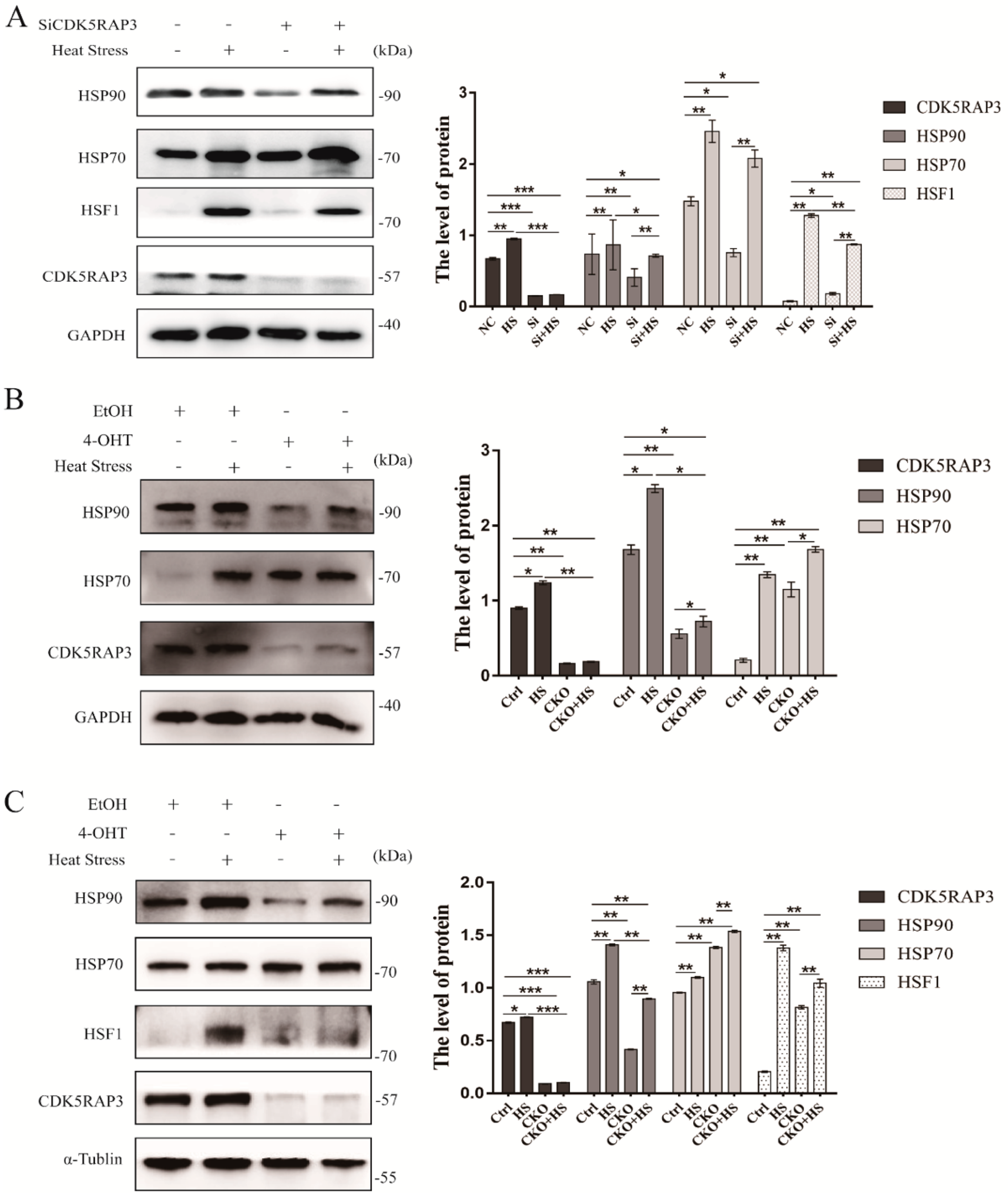
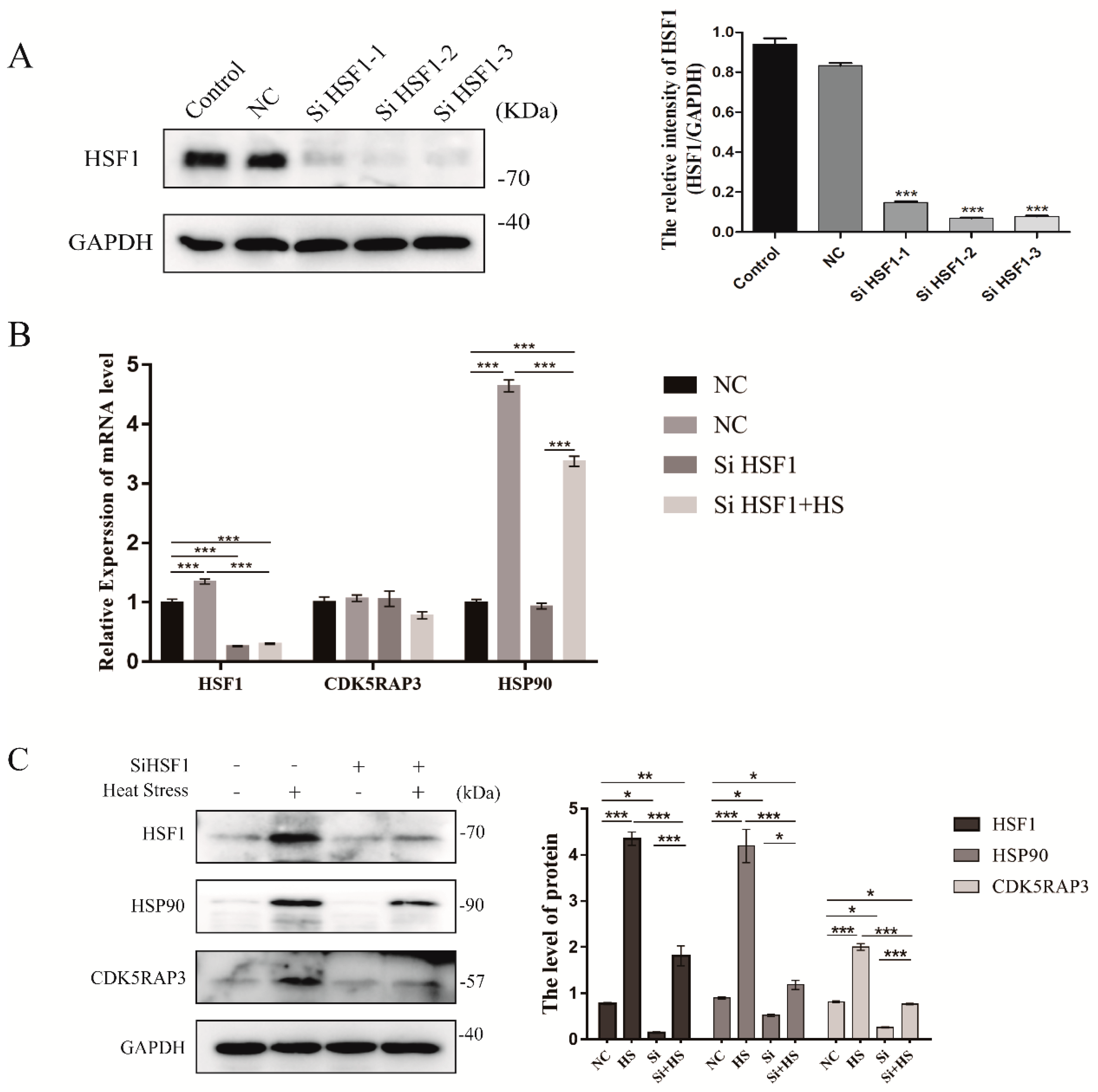
2.4.1. CDK5RAP3 Depletion Inhibited the Expression of HSP90, While HSP70 Showed No Significant Change
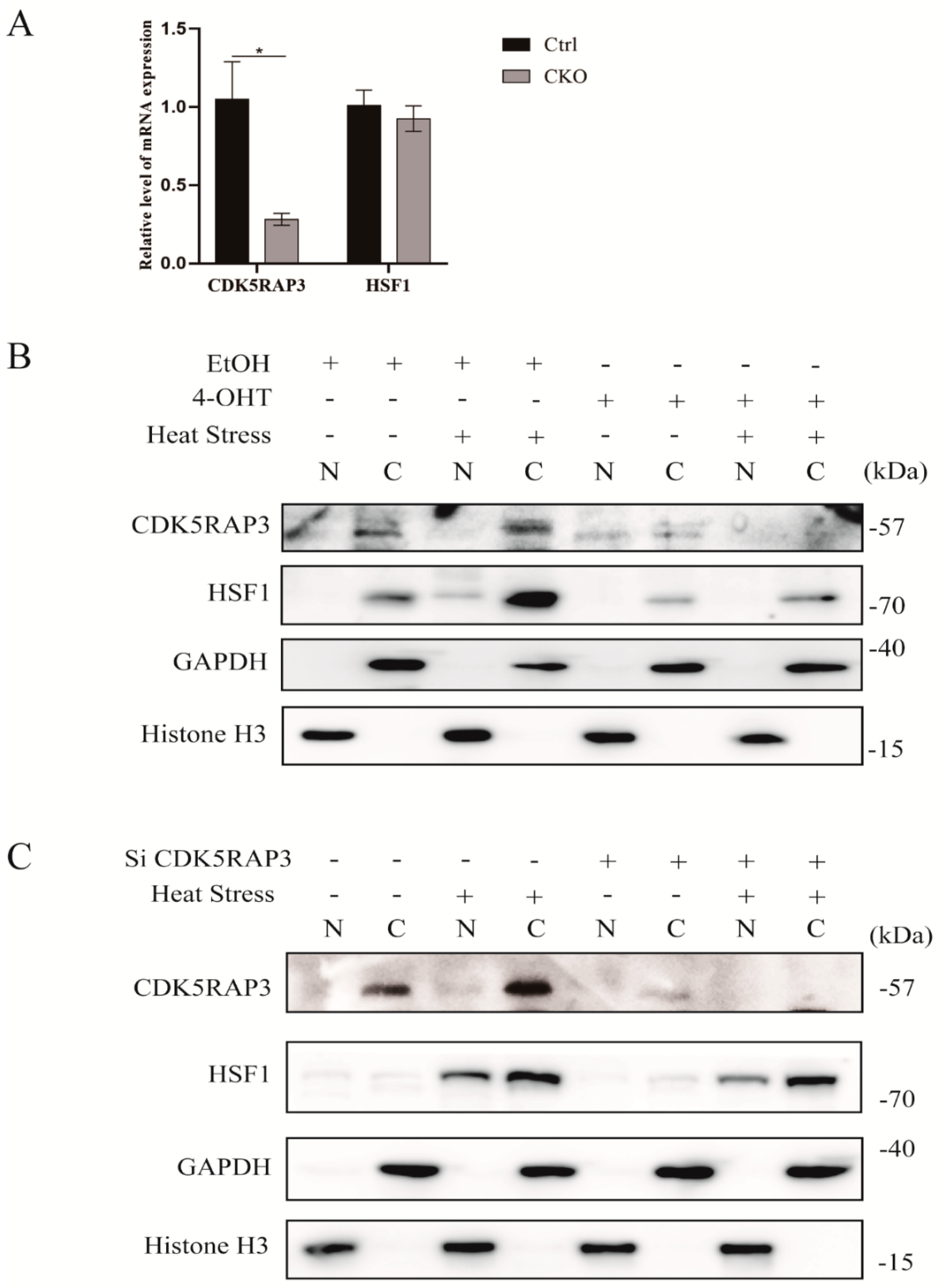
2.4.2. The Knockdown of CDK5RAP3 Did Not Affect the mRNA Expression Level of HSF1, But the Expression of HSF1 Protein
2.4.3. The Knockout of CDK5RAP3 Gene Affected the Nuclear and Cytoplasmic Distribution of HSF1 and Inhibited the Trimer Formation of HSF1
2.4.4. CDK5RAP3 Interacts with HSF1 to Influence the Expression of HSP90
2.4.5. The Knockdown of HSF1 Affect the mRNA and Protein Expression Level of CDK5RAP3 in BMECs
2.4.6. Immunofluorescence Results Show that CDK5RAP3 Binds to HSF1
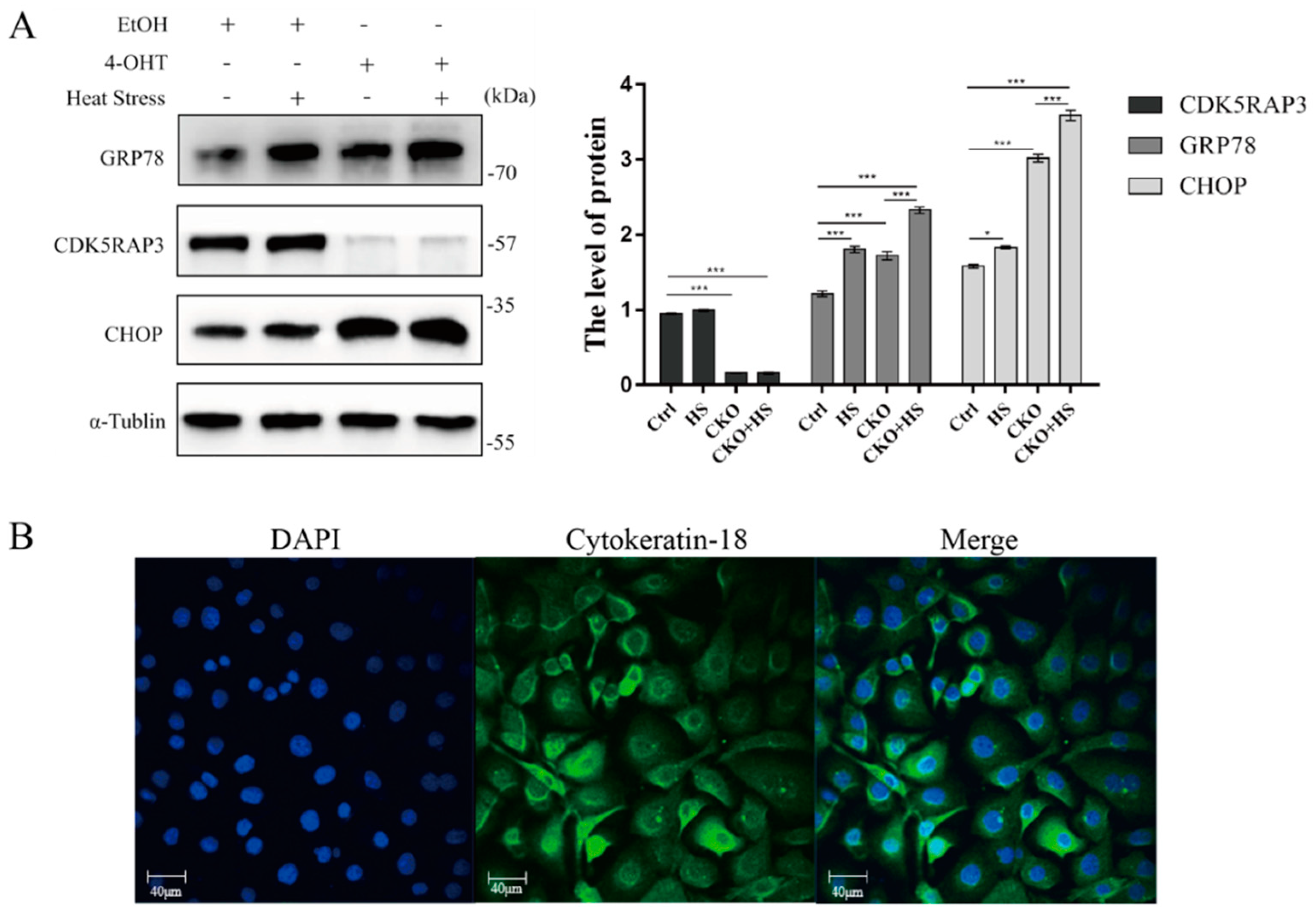
2.5. Alteration in Expression of GRP78 and CHOP Due to Deficiency of CDK5RAP3 and Heat Stress
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemical Reagents
4.3. Cell Culture and Heat Treatment
4.4. Identification of Bovine Mammary Epithelial Cells by Immunofluorescence
4.5. Total RNA Isolation and qRT-PCR Analysis
4.6. The Extraction of Cellular Nuclear and Cytoplasmic Protein
4.7. The Detection of Tissue and Cellular Protein Concentration
4.8. Western Blot Analysis
4.9. Trimerization Detection Experiment
4.10. Co-Immunoprecipitation
4.11. Immunofluorescent Staining
4.12. Statistical Method
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sanh, M.V.; Wiktorsson, H.; Ly, L.V. Effect of feeding level on milk production, body weight change, feed conversion and postpartum oestrus of crossbred lactating cows in tropical conditions. J. Shanxi Coll. Tradit. Chin. Med. 2007, 77, 331–338. [Google Scholar] [CrossRef]
- Wang, J.; Xue, X.; Liu, Q.; Zhang, S.; Peng, M.; Zhou, J.; Chen, L.; Fang, F. Effects of duration of thermal stress on growth performance, serum oxidative stress indices, the expression and localization of ABCG2 and mitochondria ROS production of skeletal muscle, small intestine and immune organs in broilers. J. Biol. 2019, 85, 102420. [Google Scholar] [CrossRef]
- Bhanuprakash, V.; Singh, U.; Sengar, G.; Sajjanar, B.; Bhusan, B.; Raja, T.V.; Alex, R.; Kumar, S.; Singh, R.; Ashish, K.; et al. Differential effect of thermal stress on HSP70 expression, nitric oxide production and cell proliferation among native and crossbred dairy cattle. J. Biol. 2016, 59, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Badri, T.M.; Chen, K.L.; Alsiddig, M.A.; Li, L.; Cai, Y.; Wang, G.L. Genetic polymorphism in Hsp90AA1 gene is associated with the thermotolerance in Chinese Holstein cows. Cell Stress Chaperones 2018, 23, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Trifkovic, J.; Jovanovic, L.; Duric, M.; Stevanovic-Dordevic, S.; Milanovic, S.; Lazarevic, M.; Sladojevic, Z.; Kirovski, D. Influence of different seasons during late gestation on Holstein cows’ colostrum and postnatal adaptive capability of their calves. Int. J. Biometeorol. 2018, 62, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Polsky, L.; von Keyserlingk, M.A.G. Invited review: Effects of heat stress on dairy cattle welfare. J. Dairy Sci. 2017, 100, 8645–8657. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Yang, X.; Ren, X.; Maliskova, L.; Li, B.; Jones, I.R.; Wang, C.; Jacob, F.; Wu, K.; Traglia, M.; et al. Mapping cis-regulatory chromatin contacts in neural cells links neuropsychiatric disorder risk variants to target genes. Nat. Genet. 2019, 51, 1252–1262. [Google Scholar] [CrossRef]
- Almeida, R.A.; Kerro-Dego, O.; Rius, A.G. Effect of heat stress on the interaction of Streptococcus uberis with bovine mammary epithelial cells. J. Dairy Res. 2018, 85, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Takii, R.; Fujimoto, M.; Matsumoto, M.; Srivastava, P.; Katiyar, A.; Nakayama, K.I.; Nakai, A. The pericentromeric protein shugoshin 2 cooperates with HSF1 in heat shock response and RNA Pol II recruitment. EMBO J. 2019, 38, e102566. [Google Scholar] [CrossRef]
- Pincus, D. Regulation of Hsf1 and the Heat Shock Response. Adv. Exp. Med. Biol. 2020, 1243, 41–50. [Google Scholar]
- Yin, C.F.; Kao, S.C.; Hsu, C.L.; Chang, Y.W.; Cheung, C.H.Y.; Huang, H.C.; Juan, H.F. Phosphoproteome Analysis Reveals Dynamic Heat Shock Protein 27 Phosphorylation in Tanshinone IIA-Induced Cell Death. J. Proteome Res. 2020, 19, 1620–1634. [Google Scholar] [CrossRef] [PubMed]
- Workman, P. Reflections and Outlook on Targeting HSP90, HSP70 and HSF1 in Cancer: A Personal Perspective. Adv. Exp. Med. Biol. 2020, 1243, 163–179. [Google Scholar]
- Bhatti, M.; Dinn, S.; Miskiewicz, E.I.; MacPhee, D.J. Expression of heat shock factor 1, heat shock protein 90 and associated signaling proteins in pregnant rat myometrium: Implications for myometrial proliferation. Reprod. Biol. 2019, 19, 374–385. [Google Scholar] [CrossRef]
- Bickel, D.; Gohlke, H. C-terminal modulators of heat shock protein of 90kDa (HSP90): State of development and modes of action. Bioorg. Med. Chem. 2019, 27, 115080. [Google Scholar] [CrossRef]
- Gaur, D.; Singh, P.; Guleria, J.; Gupta, A.; Kaur, S.; Sharma, D. The Yeast Hsp70 Co-chaperone Ydj1 Regulates Functional Distinction of Ssa Hsp70s in the Hsp90 Chaperoning Pathway. Genetics 2020, 215, 683–698. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Pastor, R.; Burchfiel, E.T.; Thiele, D.J. Regulation of heat shock transcription factors and their roles in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, A.; Fujimoto, M.; Tan, K.; Kurashima, A.; Srivastava, P.; Okada, M.; Takii, R.; Nakai, A. HSF1 is required for induction of mitochondrial chaperones during the mitochondrial unfolded protein response. FEBS Open Bio. 2020, 10, 1135–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Wang, H.; Kang, B.; Chen, B.; Shi, Y.; Yang, S.; Sun, L.; Liu, Y.; Xiao, W.; Zhang, T.; et al. CDK5RAP3, a UFL1 substrate adaptor, is crucial for liver development. Development 2019, 146, dev169235. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Luo, S.; Li, H. Cdk5 activator-binding protein C53 regulates apoptosis induced by genotoxic stress via modulating the G2/M DNA damage checkpoint. J. Biol. Chem. 2005, 280, 20651–20659. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ching, Y.P.; Lam, W.H.; Qi, Z.; Zhang, M.; Wang, J.H. Identification of a common protein association region in the neuronal Cdk5 activator. J. Biol. Chem. 2000, 275, 31763–31769. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.Y.; Liu, L.C.; Wang, J.B.; Xie, J.W.; Lin, J.X.; Lu, J.; Cao, L.L.; Lin, M.; Tu, R.H.; Huang, C.M.; et al. CDK5RAP3 Inhibits the Translocation of MCM6 to Influence the Prognosis in Gastric Cancer. J. Cancer 2019, 10, 4488–4498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.B.; Wang, Z.W.; Li, Y.; Huang, C.Q.; Zheng, C.H.; Li, P.; Xie, J.W.; Lin, J.X.; Lu, J.; Chen, Q.Y.; et al. CDK5RAP3 acts as a tumor suppressor in gastric cancer through inhibition of beta-catenin signaling. Cancer Lett. 2017, 385, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shi, Y.; Li, Z.; Yu, H.; Han, Y.; Wang, X.; Sun, K.; Yang, T.; Lou, K.; Song, Y.; et al. A functional variant of IC53 correlates with the late onset of colorectal cancer. Mol. Med. 2011, 17, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Mak, G.W.; Chan, M.M.; Leong, V.Y.; Lee, J.M.; Yau, T.O.; Ng, I.O.; Ching, Y.P. Overexpression of a novel activator of PAK4, the CDK5 kinase-associated protein CDK5RAP3, promotes hepatocellular carcinoma metastasis. Cancer Res. 2011, 71, 2949–2958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stav, D.; Bar, I.; Sandbank, J. Usefulness of CDK5RAP3, CCNB2, and RAGE genes for the diagnosis of lung adenocarcinoma. Int. J. Biol. Markers 2007, 22, 108–113. [Google Scholar] [CrossRef]
- Wamsley, J.J.; Gary, C.; Biktasova, A.; Hajek, M.; Bellinger, G.; Virk, R.; Issaeva, N.; Yarbrough, W.G. Loss of LZAP inactivates p53 and regulates sensitivity of cells to DNA damage in a p53-dependent manner. Oncogenesis 2017, 6, e314. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Wu, J.; He, C.; Yang, W.; Li, H. Tumor suppressor protein C53 antagonizes checkpoint kinases to promote cyclin-dependent kinase 1 activation. Cell Res. 2009, 19, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Pi, W.; Sivaprakasam, S.; Zhu, X.; Zhang, M.; Chen, J.; Makala, L.; Lu, C.; Wu, J.; Teng, Y.; et al. UFBP1, a Key Component of the Ufm1 Conjugation System, Is Essential for Ufmylation-Mediated Regulation of Erythroid Development. PLoS Genet. 2015, 11, e1005643. [Google Scholar] [CrossRef]
- Lin, J.X.; Xie, X.S.; Weng, X.F.; Zheng, C.H.; Xie, J.W.; Wang, J.B.; Lu, J.; Chen, Q.Y.; Cao, L.L.; Lin, M.; et al. Low expression of CDK5RAP3 and DDRGK1 indicates a poor prognosis in patients with gastric cancer. World J. Gastroenterol. 2018, 24, 3898–3907. [Google Scholar] [CrossRef]
- Zheng, C.H.; Wang, J.B.; Lin, M.Q.; Zhang, P.Y.; Liu, L.C.; Lin, J.X.; Lu, J.; Chen, Q.Y.; Cao, L.L.; Lin, M.; et al. CDK5RAP3 suppresses Wnt/beta-catenin signaling by inhibiting AKT phosphorylation in gastric cancer. J. Exp. Clin. Cancer Res. 2018, 37, 59. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.X.; Weng, X.F.; Xie, X.S.; Lian, N.Z.; Qiu, S.L.; Wang, J.B.; Lu, J.; Chen, Q.Y.; Cao, L.L.; Lin, M.; et al. CDK5RAP3 inhibits angiogenesis in gastric neuroendocrine carcinoma by modulating AKT/HIF-1alpha/VEGFA signaling. Cancer Cell Int. 2019, 19, 282. [Google Scholar] [CrossRef] [Green Version]
- Egusquiaguirre, S.P.; Liu, S.; Tosic, I.; Jiang, K.; Walker, S.R.; Nicolais, M.; Saw, T.Y.; Xiang, M.; Bartel, K.; Nelson, E.A.; et al. CDK5RAP3 is a co-factor for the oncogenic transcription factor STAT3. Neoplasia 2020, 22, 47–59. [Google Scholar] [CrossRef]
- Wang, J.; An, H.; Mayo, M.W.; Baldwin, A.S.; Yarbrough, W.G. LZAP, a putative tumor suppressor, selectively inhibits NF-kappaB. Cancer Cell 2007, 12, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Shao, J.; Li, Y.; Zhao, F.Q.; Liu, J.X.; Liu, H. Protective Effects of Inorganic and Organic Selenium on Heat Stress in Bovine Mammary Epithelial Cells. Oxid. Med. Cell Longev. 2019, 2019, 1503478. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Hu, Y.; Zheng, T.; He, H.; Xiao, W.; Liu, B.; Shi, Y.; Jia, X.; Chen, S.; Wang, J.; et al. MicroRNA-216b inhibits heat stress-induced cell apoptosis by targeting Fas in bovine mammary epithelial cells. Cell Stress Chaperones 2018, 23, 921–931. [Google Scholar] [CrossRef]
- Xiao, Y.; Rungruang, S.; Hall, L.W.; Collier, J.L.; Dunshea, F.R.; Collier, R.J. Effects of niacin and betaine on bovine mammary and uterine cells exposed to thermal shock in vitro. J. Dairy Sci. 2017, 100, 4025–4037. [Google Scholar] [CrossRef]
- Xu, J.; Yin, B.; Huang, B.; Tang, S.; Zhang, X.; Sun, J.; Bao, E. Co-enzyme Q10 protects chicken hearts from in vivo heat stress via inducing HSF1 binding activity and Hsp70 expression. Poult. Sci. 2019, 98, 1002–1011. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Gong, J.; Murshid, A. Extracellular HSPs: The Complicated Roles of Extracellular HSPs in Immunity. Front. Immunol. 2016, 7, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Liu, T.; Rios, Z.; Mei, Q.; Lin, X.; Cao, S. Heat Shock Proteins and Cancer. Trends Pharm. Sci. 2017, 38, 226–256. [Google Scholar] [CrossRef]
- Bepperling, A.; Alte, F.; Kriehuber, T.; Braun, N.; Weinkauf, S.; Groll, M.; Haslbeck, M.; Buchner, J. Alternative bacterial two-component small heat shock protein systems. Proc. Natl. Acad. Sci. USA 2012, 109, 20407–20412. [Google Scholar] [CrossRef] [Green Version]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Erlejman, A.G.; Lagadari, M.; Toneatto, J.; Piwien-Pilipuk, G.; Galigniana, M.D. Regulatory role of the 90-kDa-heat-shock protein (Hsp90) and associated factors on gene expression. Biochim. Biophys. Acta 2014, 1839, 71–87. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Loew, C.; Korner, R.; Pinkert, S.; Theis, M.; Hayer-Hartl, M.; Buchholz, F.; Hartl, F.U. Interplay of acetyltransferase EP300 and the proteasome system in regulating heat shock transcription factor 1. Cell 2014, 156, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Budzynski, M.A.; Puustinen, M.C.; Joutsen, J.; Sistonen, L. Uncoupling Stress-Inducible Phosphorylation of Heat Shock Factor 1 from Its Activation. Mol. Cell Biol. 2015, 35, 2530–2540. [Google Scholar] [CrossRef] [Green Version]
- Millson, S.H.; Piper, P.W. Insights from yeast into whether the inhibition of heat shock transcription factor (Hsf1) by rapamycin can prevent the Hsf1 activation that results from treatment with an Hsp90 inhibitor. Oncotarget 2014, 5, 5054–5064. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Dai, S.; He, Y.; Doty, R.A.; Shultz, L.D.; Sampson, S.B.; Dai, C. MEK guards proteome stability and inhibits tumor-suppressive amyloidogenesis via HSF1. Cell 2015, 160, 729–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Sharma, A.; Sodhi, M.; Verma, P.; Parvesh, K.; Swami, S.K.; Jast, A.; Shandilya, U.K.; Mukesh, M. Characterizing binding sites of heat responsive microRNAs and their expression pattern in heat stressed PBMCs of native cattle, exotic cattle and riverine buffaloes. Mol. Biol. Rep. 2019, 46, 6513–6524. [Google Scholar] [CrossRef]
- Roth, Z. Effect of Heat Stress on Reproduction in Dairy Cows: Insights into the Cellular and Molecular Responses of the Oocyte. Annu. Rev. Anim. Biosci. 2017, 5, 151–170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, X.; Zhang, Y.; Cai, Y.; Chen, J.; Sivaprakasam, S.; Gurav, A.; Pi, W.; Makala, L.; Wu, J.; et al. RCAD/Ufl1, a Ufm1 E3 ligase, is essential for hematopoietic stem cell function and murine hematopoiesis. Cell Death Differ. 2015, 22, 1922–1934. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Y.; Xia, Q.; Song, H.; Wei, R.; Wang, J.; Zou, K. Cadherin 22 participates in the self-renewal of mouse female germ line stem cells via interaction with JAK2 and beta-catenin. Cell Mol. Life Sci. 2018, 75, 1241–1253. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences |
|---|---|
| mβ-ACTIN | Forward: 5′-GACCTCTATGCCAACACAGT-3′ |
| Reverse: 5′-AGTACTTGCGCTCAGGAGGA-3′ | |
| mGAPDH | Forward: 5′-AACTTTGGCATTGTGGAAGG-3′ |
| Reverse: 5′-ACACATTGGGGGTAGGAACA-3′ | |
| mCDK5RAP3 | Forward: 5′-ATGAGATCGACTGGGGTGAC-3′ |
| Reverse: 5′-AGCCTCAGTTCCTGTCTCCA-3′ | |
| mHSF1 | Forward: 5′-TCATCTGCTGGAGCCCGAGTG-3′ |
| Reverse: 5′-ATGTTGAGCTGCCGCACGAAG-3′ | |
| bβ-ACTIN | Forward: 5′-GGGCAGGTCATCACCATCGG-3′ |
| Reverse: 5′-TCATTGTGCTGGGTGCCAGG-3′ | |
| bGAPDH | Forward: 5′-AAGGTCGGAGTGAAC-3′ |
| Reverse: 5′-CGTTCTCTGCCTTGACTGTG-3′ | |
| bCDK5RAP3 | Forward: 5′-TCGACTGGCTGGTAGACAGAAGG-3′ |
| Reverse: 5′-GTCCTGGATGGCGGCATTGATC-3′ | |
| bHSP90 | Forward: 5′-CCAAGTCTGGCACTAAAG-3′ |
| Reverse: 5′-GAAGACTCCCAAGCATAC-3′ | |
| bHSF1 | Forward: 5′-AAAGATCCCCCTGATGCTGAACGAC-3′ |
| Reverse: 5′-CAGTTCGGTGATGTCGGAGATGATG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Zou, Y.; Li, J.; Chen, F.; Li, H.; Cai, Y. CDK5RAP3, a Novel Nucleoplasmic Shuttle, Deeply Regulates HSF1-Mediated Heat Stress Response and Protects Mammary Epithelial Cells from Heat Injury. Int. J. Mol. Sci. 2020, 21, 8400. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218400
Shen Y, Zou Y, Li J, Chen F, Li H, Cai Y. CDK5RAP3, a Novel Nucleoplasmic Shuttle, Deeply Regulates HSF1-Mediated Heat Stress Response and Protects Mammary Epithelial Cells from Heat Injury. International Journal of Molecular Sciences. 2020; 21(21):8400. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218400
Chicago/Turabian StyleShen, Yangyang, Yan Zou, Jun Li, Fanghui Chen, Honglin Li, and Yafei Cai. 2020. "CDK5RAP3, a Novel Nucleoplasmic Shuttle, Deeply Regulates HSF1-Mediated Heat Stress Response and Protects Mammary Epithelial Cells from Heat Injury" International Journal of Molecular Sciences 21, no. 21: 8400. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218400