Gestational Exposure to Bisphenol A Affects Testicular Morphology, Germ Cell Associations, and Functions of Spermatogonial Stem Cells in Male Offspring

 ,
,
Abstract
:1. Introduction
2. Results
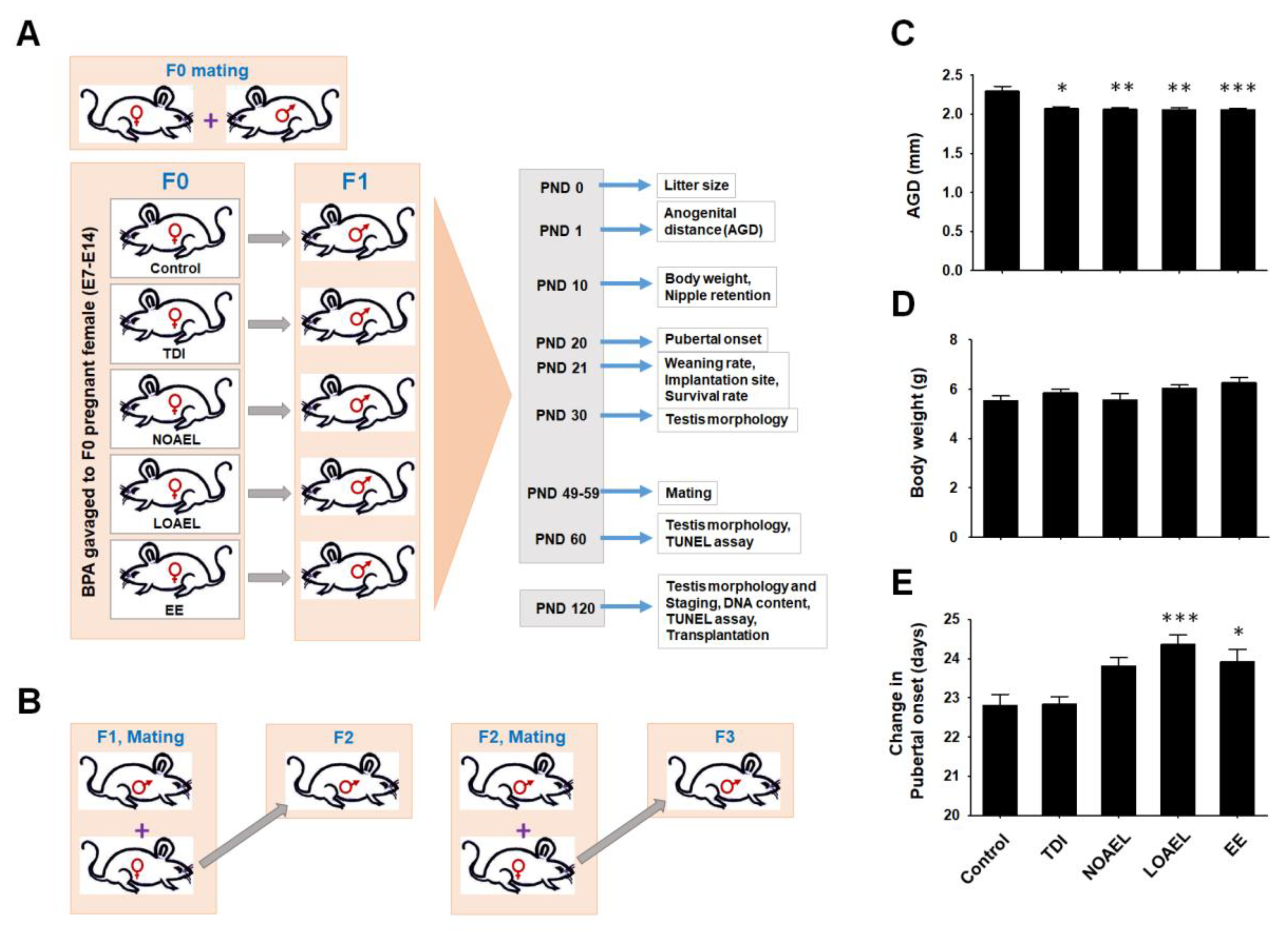
2.1. Effects of BPA on Embryo Survival, Anogenital Distance, and Nipple Retention in CD1 Male Pups
2.2. Effects of BPA on Body Weight, Pubertal Onset, and Testis Weight of F1 Male Pups
2.3. BPA Increases Frequency of Abnormal Seminiferous Tubules in F1 and F2 Offspring
2.4. BPA Affects the Area of the Seminiferous Epithelium and Alters the Stages of Seminiferous Tubule
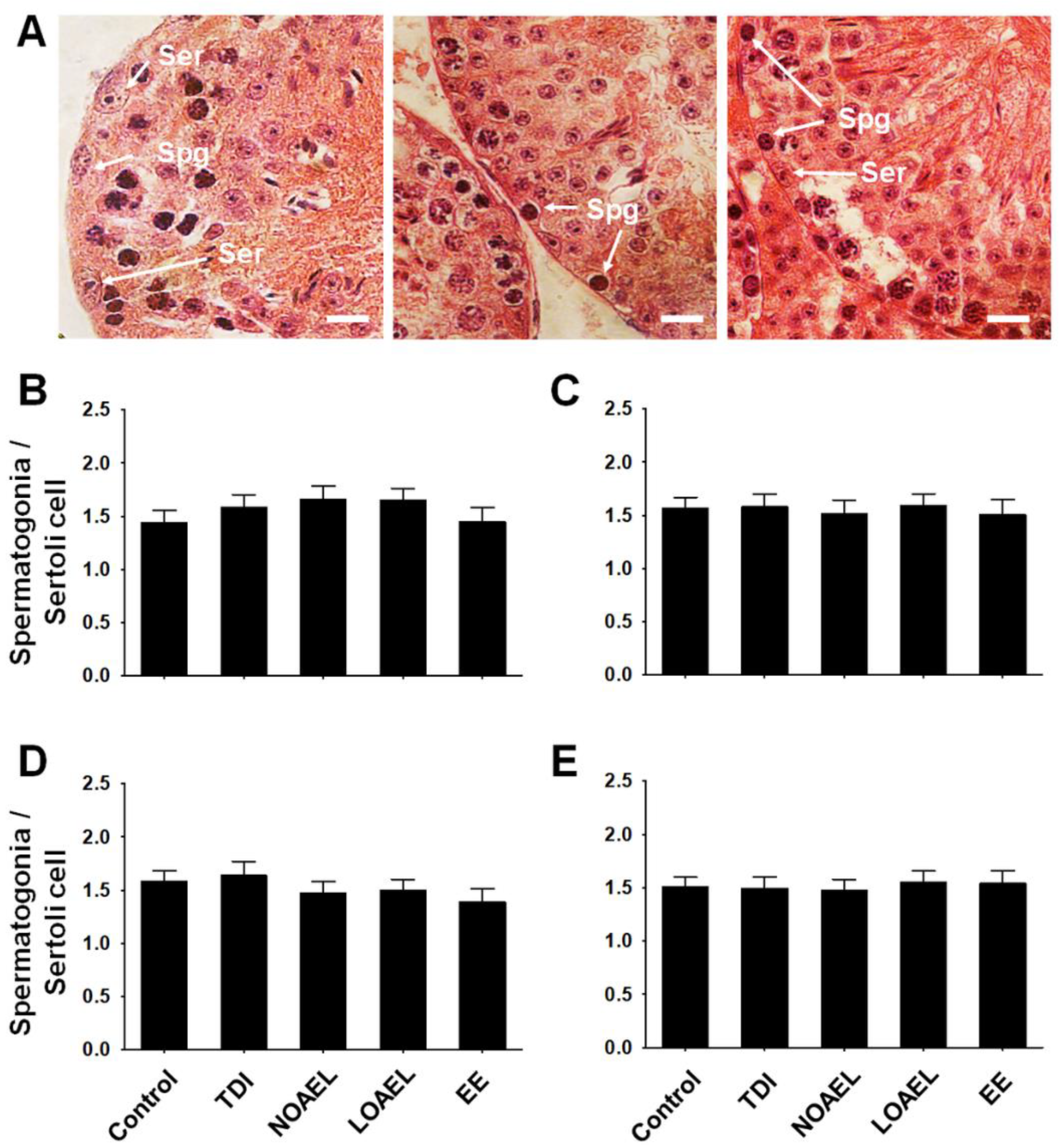
2.5. Effects of BPA on the Number of Spermatogonia and Germ Cells
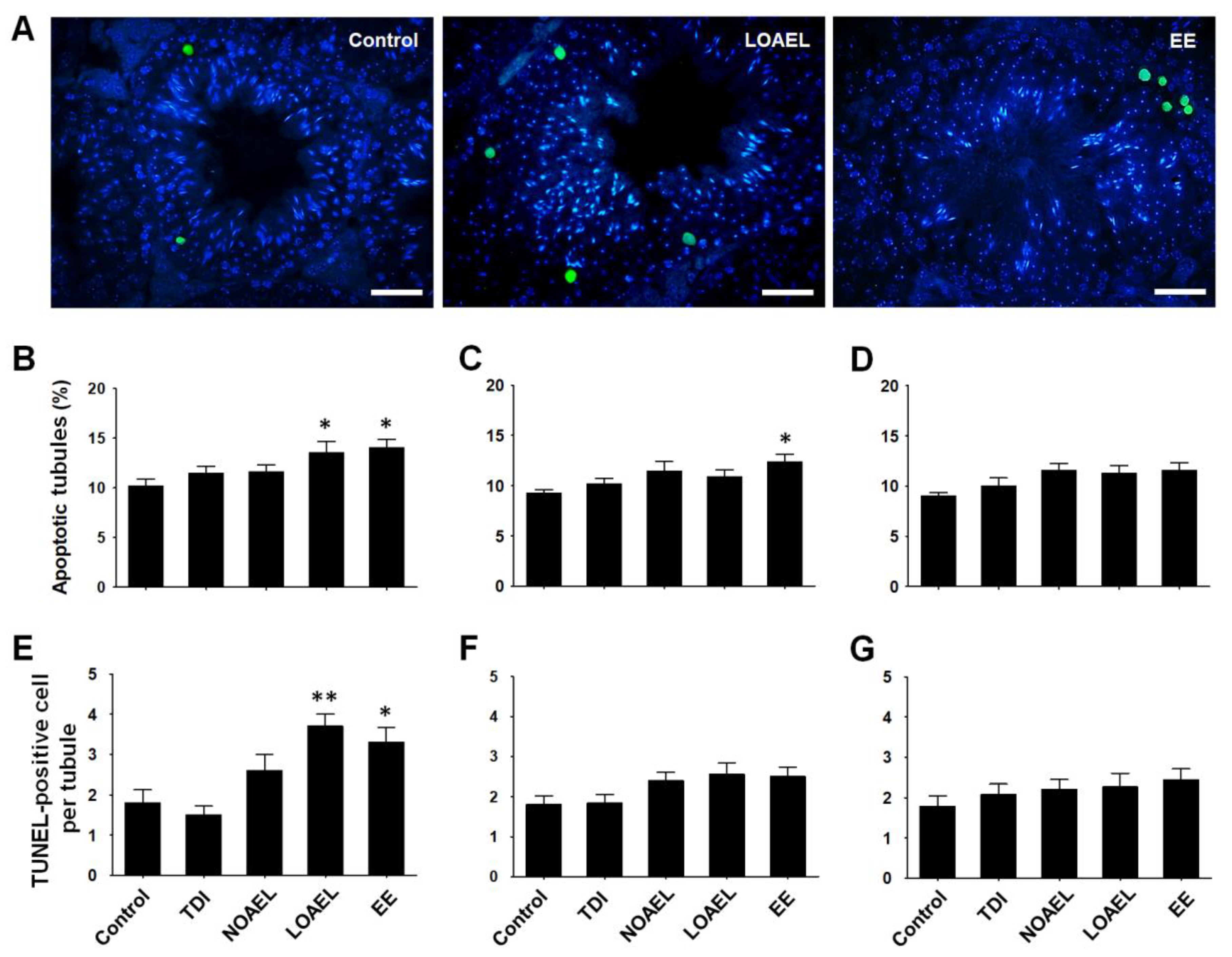
2.6. BPA Exposure Induces Germ Cell Apoptosis
2.7. BPA Exposure Does Not Change Litter Size of F1 and F2 Generations
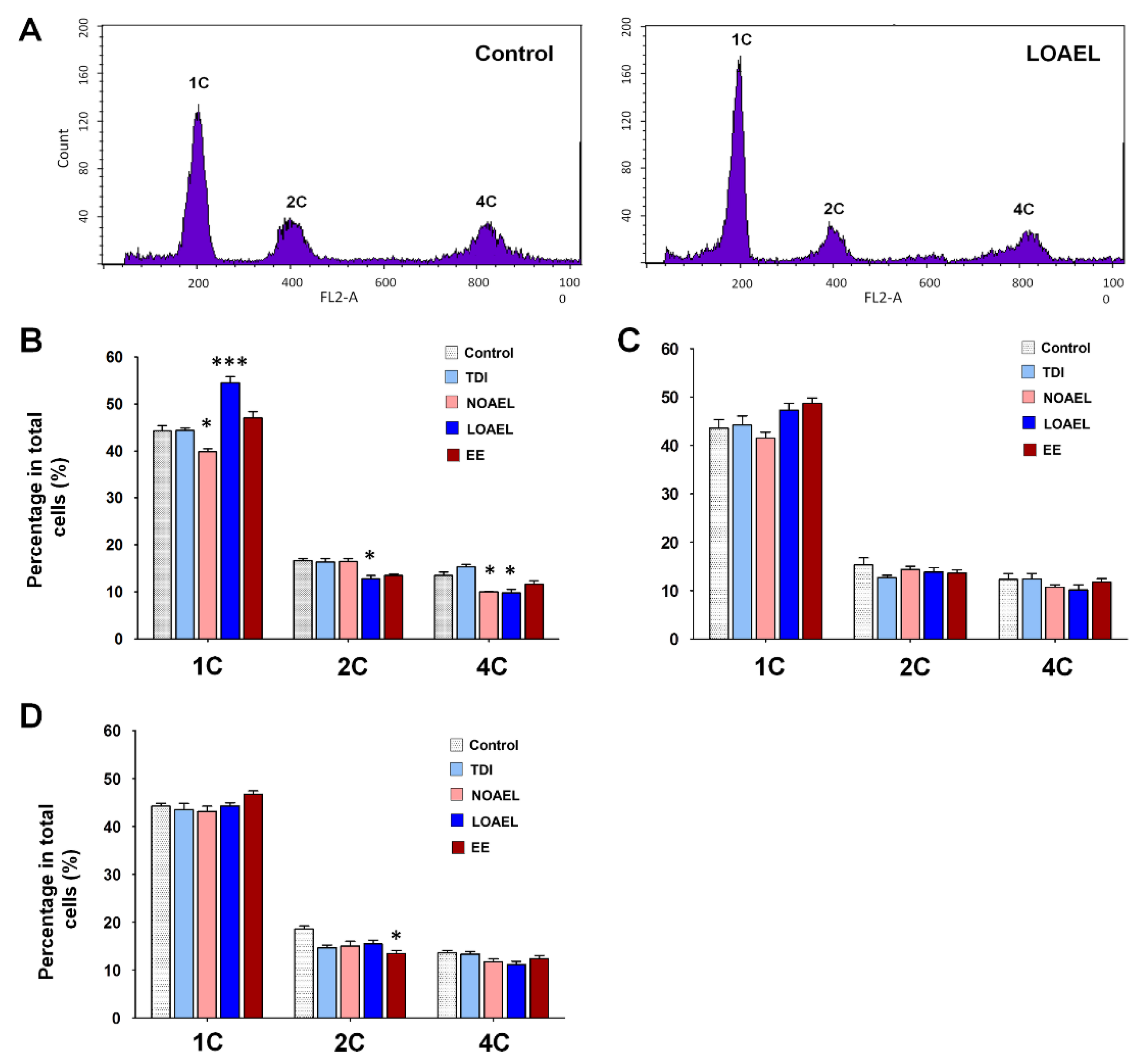
2.8. BPA Affects the Stemness Properties of Spermatogonial Stem Cell in F1 Offspring
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design, BPA Exposure, and Breeding to Generate F1–F3 Offspring
4.3. Embryo Survival, Neonatal and Pubertal Period Analysis, Implantation Sites, and Weaning Rates
4.4. Collection of Testis and Determination of Testicular Abnormalities
4.5. PAS-Hematoxylin Staining, Staging of Seminiferous Epithelium, and Counting of Spermatogonia
4.6. Flow Cytometric Analysis
4.7. Detection of Germ Cell Apoptosis
4.8. Germ Cell Transplantation for Evaluating SSCs Activity after BPA Exposure
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AGD | Anogenital distance measurements |
| ANOVA | Analysis of variance |
| BPA | Bisphenol A |
| BW | Body weight |
| ED | Endocrine disruptor |
| EE | Ethinylestradiol |
| FACS | Fluorescence-activated cell sorting |
| LOAEL | Lowest-observed-adverse-effect-level |
| NOAEL | No-observed-adverse-effect-level |
| PAS | Periodic acid-schiff |
| PND | Postnatal day |
| SE | Seminiferous epithelium |
| SSCs | Spermatogonial stem cells |
| ST | Seminiferous tubules |
| TDI | Tolerable daily intake |
References
- Lang, I.A.; Galloway, T.S.; Scarlett, A.; Henley, W.E.; Depledge, M.; Wallace, R.B.; Melzer, D. Association of urinary bisphenol A concentration with medical disorders and laboratory abnormalities in adults. JAMA 2008, 300, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.R.; Schoenfelder, G. Urinary, circulating, and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Environ. Health Perspect. 2010, 118, 1055–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, H.H.; Carlson, E.M.; Chua, J.P.; Belcher, S.M. Bisphenol A is released from polycarbonate drinking bottles and mimics the neurotoxic actions of estrogen in developing cerebellar neurons. Toxicol. Lett. 2008, 176, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maragou, N.C.; Makri, A.; Lampi, E.N.; Thomaidis, N.S.; Koupparis, M.A. Migration of bisphenol A from polycarbonate baby bottles under real use conditions. Food Addit. Contam. 2008, 25, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Richter, C.A.; Birnbaum, L.S.; Farabollini, F.; Newbold, R.R.; Rubin, B.S.; Talsness, C.E.; Vandenbergh, J.G.; Walser-Kuntz, D.R.; vom Saal, F.S. In vivo effects of bisphenol A in laboratory rodent studies. Reprod. Toxicol. 2007, 24, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Cummings, A.M.; Laws, S.C. Assessment of estrogenicity by using the delayed implanting rat model and examples. Reprod. Toxicol. 2000, 14, 111–117. [Google Scholar] [CrossRef]
- Krishnan, A.V.; Stathis, P.; Permuth, S.F.; Tokes, L.; Feldman, D. Bisphenol-A: An Estrogenic Substance Is Released from Polycarbonate Flasks during Autoclaving. Endocrinology 1993, 132, 2279–2286. [Google Scholar] [CrossRef]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol. 2011, 127, 27–34. [Google Scholar] [CrossRef]
- Morrisey, R.E.; George, J.D.; Price, C.J.; Tyl, R.W.; Marr, M.C.; Kimmel, C.A. The Developmental Toxicity of Bisphenol-a in Rats and Mice. Fundam. Appl. Toxicol. 1987, 8, 571–582. [Google Scholar] [CrossRef]
- Bindhumol, V.; Chitra, K.C.; Mathur, P.P. Bisphenol A induces reactive oxygen species generation in the liver of male rats. Toxicology 2003, 188, 117–124. [Google Scholar] [CrossRef]
- Ranciere, F.; Lyons, J.G.; Loh, V.H.Y.; Botton, J.; Galloway, T.; Wang, T.G.; Shaw, J.E.; Magliano, D.J. Bisphenol A and the risk of cardiometabolic disorders: A systematic review with meta-analysis of the epidemiological evidence. Environ. Heal. 2015, 14, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustieles, V.; Perez-Lobato, R.; Olea, N.; Fernandez, M.F. Bisphenol A: Human exposure and neurobehavior. Neurotoxicology 2015, 49, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Urriola-Munoz, P.; Lagos-Cabre, R.; Moreno, R.D. A Mechanism of Male Germ Cell Apoptosis Induced by Bisphenol-A and Nonylphenol Involving ADAM17 and p38 MAPK Activation. PLoS ONE 2014, 9, e113793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oatley, J.M.; Brinster, R.L. Regulation of Spermatogonial Stem Cell Self-Renewal in Mammals. Annu. Rev. Cell Dev. Biol. 2008, 24, 263–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinster, R.L. Male germline stem cells: From mice to men. Science 2007, 316, 404–405. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C.; Rubin, B.S.; Soto, A.M. Bisphenol-A and the Great Divide: A Review of Controversies in the Field of Endocrine Disruption. Endocr. Rev. 2009, 30, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Kwon, W.S.; Yoon, S.J.; Park, Y.J.; Ryu, B.Y.; Pang, M.G. A novel approach to assessing bisphenol-A hazards using an in vitro model system. BMC Genom. 2016, 17, 577. [Google Scholar] [CrossRef]
- Li, J.; Mao, R.; Zhou, Q.; Ding, L.; Tao, J.; Ran, M.M.; Gao, E.S.; Yuan, W.; Wang, J.T.; Hou, L.F. Exposure to bisphenol A (BPA) in Wistar rats reduces sperm quality with disruption of ERK signal pathway. Toxicol. Mech. Methods 2016, 26, 180–188. [Google Scholar] [CrossRef]
- Tiwari, D.; Vanage, G. Mutagenic effect of Bisphenol A on adult rat male germ cells and their fertility. Reprod. Toxicol. 2013, 40, 60–68. [Google Scholar] [CrossRef]
- Rahman, M.S.; Kwon, W.S.; Lee, J.S.; Yoon, S.J.; Ryu, B.Y.; Pang, M.G. Bisphenol-A Affects Male Fertility via Fertility-related Proteins in Spermatozoa. Sci. Rep. 2015, 5, 9196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Duan, W.; Li, R.; Xu, S.; Zhang, L.; Chen, C.; He, M.; Lu, Y.; Wu, H.; Pi, H.; et al. Exposure to bisphenol A disrupts meiotic progression during spermatogenesis in adult rats through estrogen-like activity. Cell Death Dis. 2013, 4, e676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrooman, L.A.; Oatley, J.M.; Griswold, J.E.; Hassold, T.J.; Hunt, P.A. Estrogenic Exposure Alters the Spermatogonial Stem Cells in the Developing Testis, Permanently Reducing Crossover Levels in the Adult. PLoS Genet. 2015, 11, e1004949. [Google Scholar] [CrossRef] [Green Version]
- Hulak, M.; Gazo, I.; Shaliutina, A.; Linhartova, P. In vitro effects of bisphenol A on the quality parameters, oxidative stress, DNA integrity and adenosine triphosphate content in sterlet (Acipenser ruthenus) spermatozoa. Comp. Biochem. Physiol. C-Toxicol. Pharmacol. 2013, 158, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Doshi, T.; D’souza, C.; Vanage, G. Aberrant DNA methylation at Igf2-H19 imprinting control region in spermatozoa upon neonatal exposure to bisphenol A and its association with post implantation loss. Mol. Biol. Rep. 2013, 40, 4747–4757. [Google Scholar] [CrossRef] [PubMed]
- Bolt, H.M.; Stewart, J.D. Highlight report: The bisphenol A controversy. Arch. Toxicol. 2011, 85, 1491–1492. [Google Scholar] [CrossRef] [Green Version]
- Tyl, R.W. Basic Exploratory Research versus Guideline-Compliant Studies Used for Hazard Evaluation and Risk Assessment: Bisphenol A as a Case Study. Environ. Health Perspect. 2009, 117, 1644–1651. [Google Scholar] [CrossRef]
- Doyle, T.J.; Bowman, J.L.; Windell, V.L.; McLean, D.J.; Kim, K.H. Transgenerational Effects of Di-(2-ethylhexyl) Phthalate on Testicular Germ Cell Associations and Spermatogonial Stem Cells in Mice. Biol. Reprod. 2013, 88, 1–15. [Google Scholar] [CrossRef]
- Xie, M.N.; Bu, P.L.; Li, F.J.; Lan, S.J.; Wu, H.J.; Yuan, L.; Wang, Y. Neonatal bisphenol A exposure induces meiotic arrest and apoptosis of spermatogenic cells. Oncotarget 2016, 7, 10606–10615. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, P.C.; Kang, H.G.; Kim, Y.H.; Jung, S.E.; Rahman, M.S.; Lee, H.S.; Kim, Y.H.; Pang, M.G.; Ryu, B.Y. Bisphenol an Affects on the Functional Properties and Proteome of Testicular Germ Cells and Spermatogonial Stem Cells in vitro Culture Model. Sci. Rep. 2017, 7, 11858. [Google Scholar] [CrossRef]
- Ge, L.C.; Chen, Z.J.; Liu, H.; Zhang, K.S.; Su, Q.; Ma, X.Y.; Huang, H.B.; Zhao, Z.D.; Wang, Y.Y.; Giesy, J.P.; et al. Signaling related with biphasic effects of bisphenol A (BPA) on Sertoli cell proliferation: A comparative proteomic analysis. Biochim. Biophys. Acta-General. Subj. 2014, 1840, 2663–2673. [Google Scholar] [CrossRef] [PubMed]
- Okada, A.; Kai, O. Effects of estradiol-17beta and bisphenol A administered chronically to mice throughout pregnancy and lactation on the male pups’ reproductive system. Asian J. Androl. 2008, 10, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Salian, S.; Doshi, T.; Vanage, G. Perinatal exposure of rats to Bisphenol A affects the fertility of male offspring. Life Sci. 2009, 85, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Peretz, J.; Vrooman, L.; Ricke, W.A.; Hunt, P.A.; Ehrlich, S.; Hauser, R.; Padmanabhan, V.; Taylor, H.S.; Swan, S.H.; VandeVoort, C.A.; et al. Bisphenol a and reproductive health: Update of experimental and human evidence, 2007–2013. Environ. Health Perspect. 2014, 122, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Dobrzynska, M.M.; Radzikowska, J. Genotoxicity and reproductive toxicity of bisphenol A and X-ray/bisphenol A combination in male mice. Drug Chem. Toxicol. 2013, 36, 19–26. [Google Scholar] [CrossRef]
- Rahman, M.S.; Kwon, W.S.; Karmakar, P.C.; Yoon, S.J.; Ryu, B.Y.; Pang, M.G. Gestational Exposure to Bisphenol A Affects the Function and Proteome Profile of F1 Spermatozoa in Adult Mice. Environ. Health Perspect. 2017, 125, 238–245. [Google Scholar] [CrossRef]
- Durcova-Hills, G.; Capel, B. Development of germ cells in the mouse. Curr. Top. Dev. Biol. 2008, 83, 185–212. [Google Scholar] [CrossRef]
- Jasarevic, E.; Sieli, P.T.; Twellman, E.E.; Welsh, T.H., Jr.; Schachtman, T.R.; Roberts, R.M.; Geary, D.C.; Rosenfeld, C.S. Disruption of adult expression of sexually selected traits by developmental exposure to bisphenol A. Proc. Natl. Acad. Sci. USA 2011, 108, 11715–11720. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhou, Z.; Qing, D.; He, Y.; Wu, T.; Miao, M.; Wang, J.; Weng, X.; Ferber, J.R.; Herrinton, L.J.; et al. Occupational exposure to bisphenol-A (BPA) and the risk of self-reported male sexual dysfunction. Hum. Reprod. 2010, 25, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Erler, C.; Novak, J. Bisphenol an exposure: Human risk and health policy. J. Pediatr. Nurs. 2010, 25, 400–407. [Google Scholar] [CrossRef]
- Brisken, C. Endocrine disruptors and breast cancer. CHIMIA Int. J. Chem. 2008, 62, 406–409. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.T.; Zheng, J.C.; Xiao, X.M.; Zheng, S.; Dong, K.R.; Liu, J.B.; Wang, Y. Environmental endocrine disruptors promote invasion and metastasis of SK-N-SH human neuroblastoma cells. Oncol. Rep. 2010, 23, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoeller, R.T. Environmental chemicals impacting the thyroid: Targets and consequences. Thyroid 2007, 17, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Lubick, N. Cardiovascular Health Exploring a Potential Link between BPA and Heart Disease. Environ. Health Perspect. 2010, 118, A116. [Google Scholar] [CrossRef]
- Midoro-Horiuti, T.; Tiwari, R.; Watson, C.S.; Goldblum, R.M. Maternal Bisphenol an Exposure Promotes the Development of Experimental Asthma in Mouse Pups. Environ. Health Perspect. 2010, 118, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Vom Saal, F.S.; Nagel, S.C.; Coe, B.L.; Angle, B.M.; Taylor, J.A. The estrogenic endocrine disrupting chemical bisphenol A (BPA) and obesity. Mol. Cell. Endocrinol. 2012, 354, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Plastics Derived Endocrine Disruptors (BPA, DEHP and DBP) Induce Epigenetic Transgenerational Inheritance of Obesity, Reproductive Disease and Sperm Epimutations. PLoS ONE 2013, 8, e55387. [Google Scholar] [CrossRef]
- Manikkam, M.; Guerrero-Bosagna, C.; Tracey, R.; Haque, M.M.; Skinner, M.K. Transgenerational actions of environmental compounds on reproductive disease and identification of epigenetic biomarkers of ancestral exposures. PLoS ONE 2012, 7, e31901. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.; Rashid, C.; Xin, F.; Li, C.; Polyak, E.; Duemler, A.; van der Meer, T.; Stefaniak, M.; Wajid, S.; Doliba, N.; et al. Sex- and dose-specific effects of maternal bisphenol A exposure on pancreatic islets of first- and second-generation adult mice offspring. Environ. Health Perspect. 2017, 125, 097022. [Google Scholar] [CrossRef]
- Wing, T.Y.; Christensen, A.K. Morphometric Studies on Rat Seminiferous Tubules. Am. J. Anat. 1982, 165, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, E.A.; de Rooij, D.G. Staging of mouse seminiferous tubule cross-sections. Methods Mol. Biol. 2009, 558, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Timms, B.G.; Howdeshell, K.L.; Barton, L.; Bradley, S.; Richter, C.A.; vom Saal, F.S. Estrogenic chemicals in plastic and oral contraceptives disrupt development of the fetal mouse prostate and urethra. Proc. Natl. Acad. Sci. USA 2005, 102, 7014–7019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oatley, J.M.; Brinster, R.L. Spermatogonial stem cells. Methods Enzym. 2006, 419, 259–282. [Google Scholar] [CrossRef]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis Following Male Germ-Cell Transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Pregnant Mice (n) | Male Pups (n) | Litter Size | Nipple Retention (%) [PND 10] ** | Implantation Sites [PND 21] | Survival Rates (%) [PND 21] | Weaning Rates (%) [PND 21] |
|---|---|---|---|---|---|---|---|
| Control | 12 | 75 | 12.84 ± 0.44 | 4.90 ± 0.88 b | 13.12 ± 0.55 | 97.15 ± 1.31 a | 94.21 ± 2.36 |
| TDI | 12 | 73 | 13.07 ± 0.66 | 7.20 ± 1.50 b | 13.98 ± 1.83 | 94.16 ± 2.71 ab | 89.74 ± 1.81 |
| NOAEL | 13 | 77 | 12.88 ± 1.10 | 14.25 ± 1.60 a | 14.35 ± 1.33 | 92.75 ± 3.01 ab | 93.07 ± 1.93 |
| LOAEL | 14 | 82 | 12.92 ± 0.30 | 13.32 ± 1.88 a | 14.88 ± 0.38 | 89.67 ± 3.16 b | 93.43 ± 2.91 |
| EE control | 11 | 74 | 12.60 ± 0.33 | 12.94 ± 1.66 a | 13.35 ± 0.33 | 94.52 ± 1.78 ab | 92.45 ± 2.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karmakar, P.C.; Ahn, J.S.; Kim, Y.-H.; Jung, S.-E.; Kim, B.-J.; Lee, H.-S.; Ryu, B.-Y. Gestational Exposure to Bisphenol A Affects Testicular Morphology, Germ Cell Associations, and Functions of Spermatogonial Stem Cells in Male Offspring. Int. J. Mol. Sci. 2020, 21, 8644. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228644
Karmakar PC, Ahn JS, Kim Y-H, Jung S-E, Kim B-J, Lee H-S, Ryu B-Y. Gestational Exposure to Bisphenol A Affects Testicular Morphology, Germ Cell Associations, and Functions of Spermatogonial Stem Cells in Male Offspring. International Journal of Molecular Sciences. 2020; 21(22):8644. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228644
Chicago/Turabian StyleKarmakar, Polash Chandra, Jin Seop Ahn, Yong-Hee Kim, Sang-Eun Jung, Bang-Jin Kim, Hee-Seok Lee, and Buom-Yong Ryu. 2020. "Gestational Exposure to Bisphenol A Affects Testicular Morphology, Germ Cell Associations, and Functions of Spermatogonial Stem Cells in Male Offspring" International Journal of Molecular Sciences 21, no. 22: 8644. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228644