Biosynthesis of N-Docosahexanoylethanolamine from Unesterified Docosahexaenoic Acid and Docosahexaenoyl-Lysophosphatidylcholine in Neuronal Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
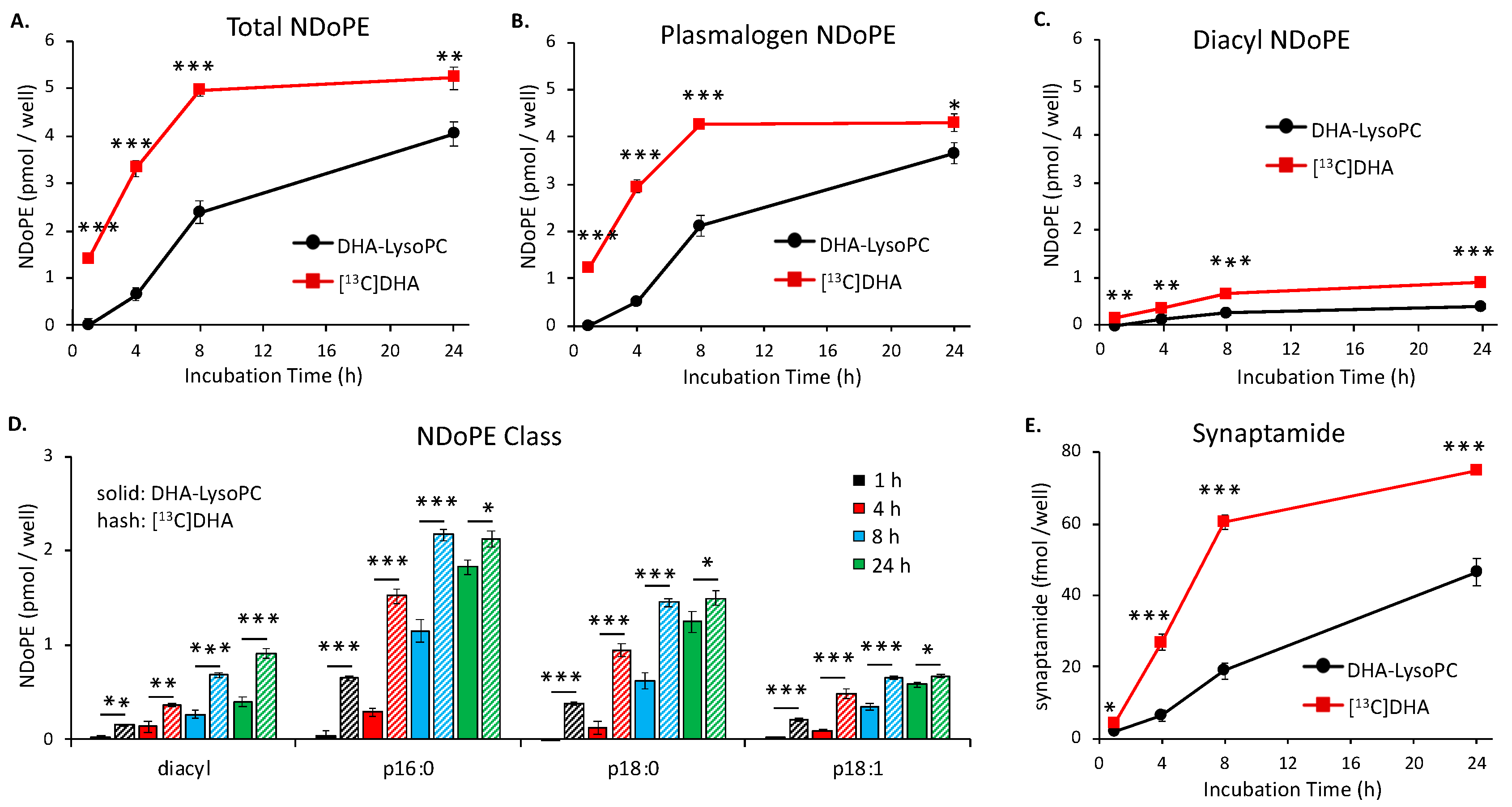
2.1. Time-Dependent NDoPE and Synaptamide Synthesis
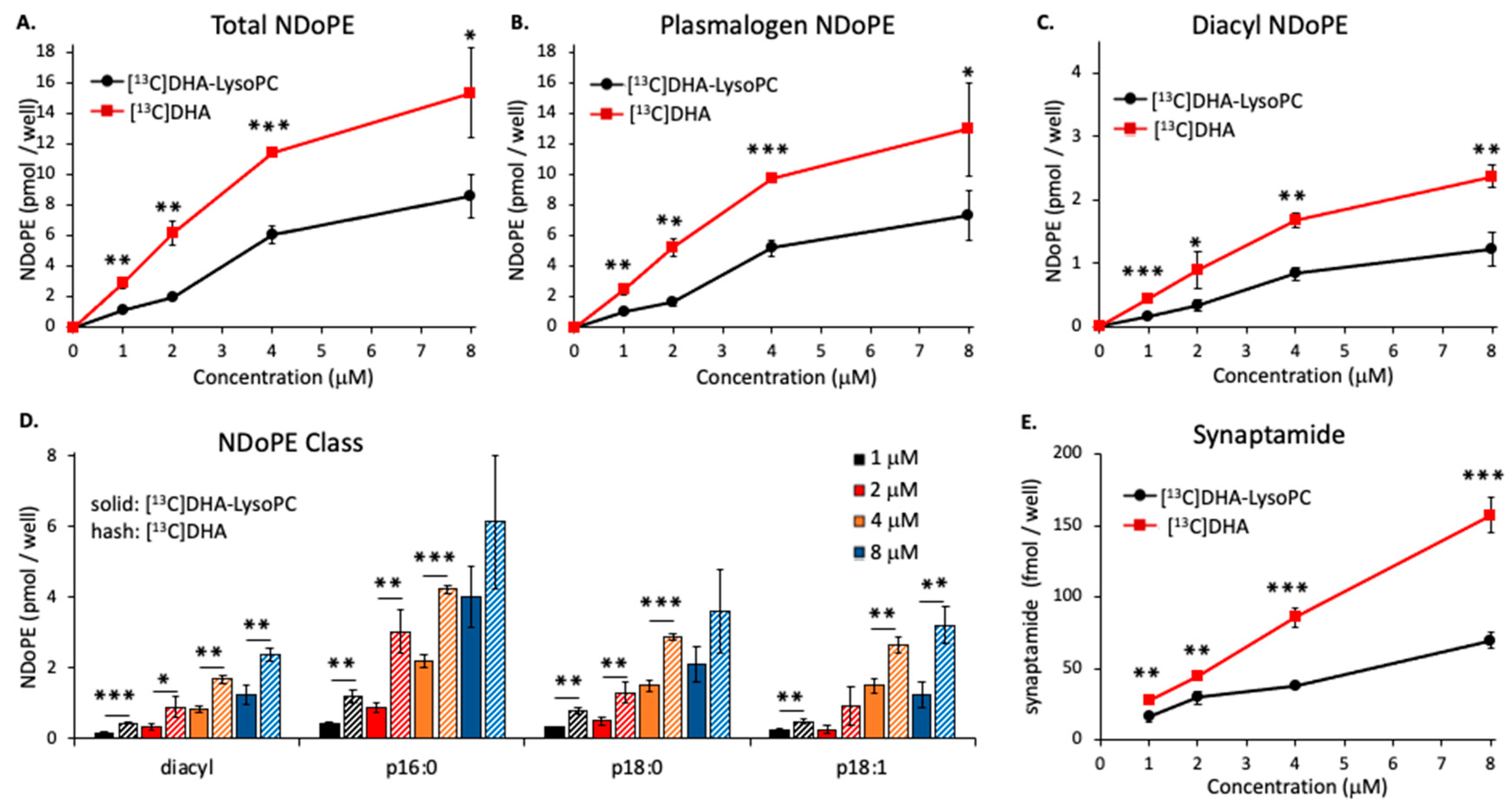
2.2. Concentration-Dependent NDoPE and Synaptamide Synthesis
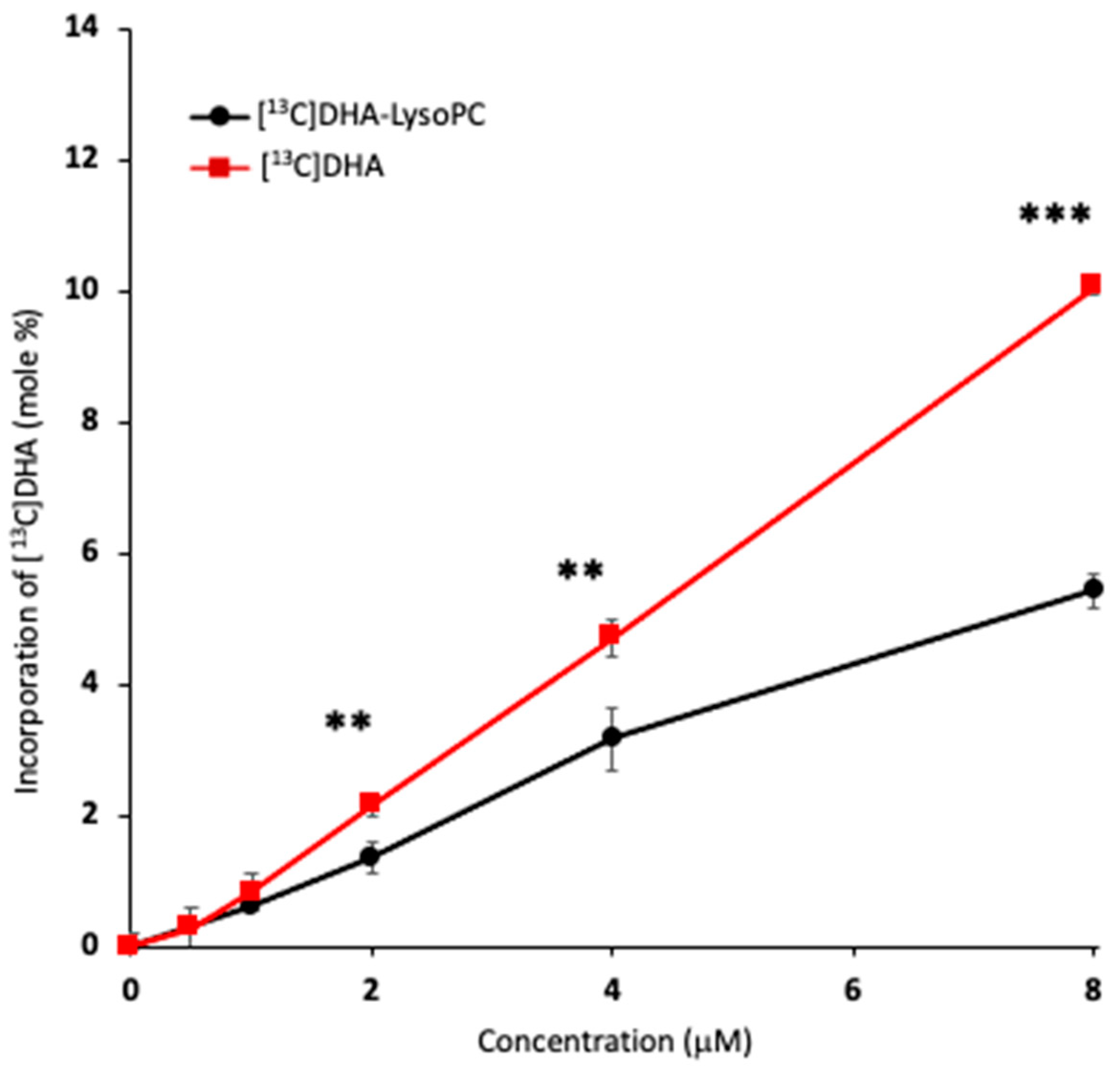
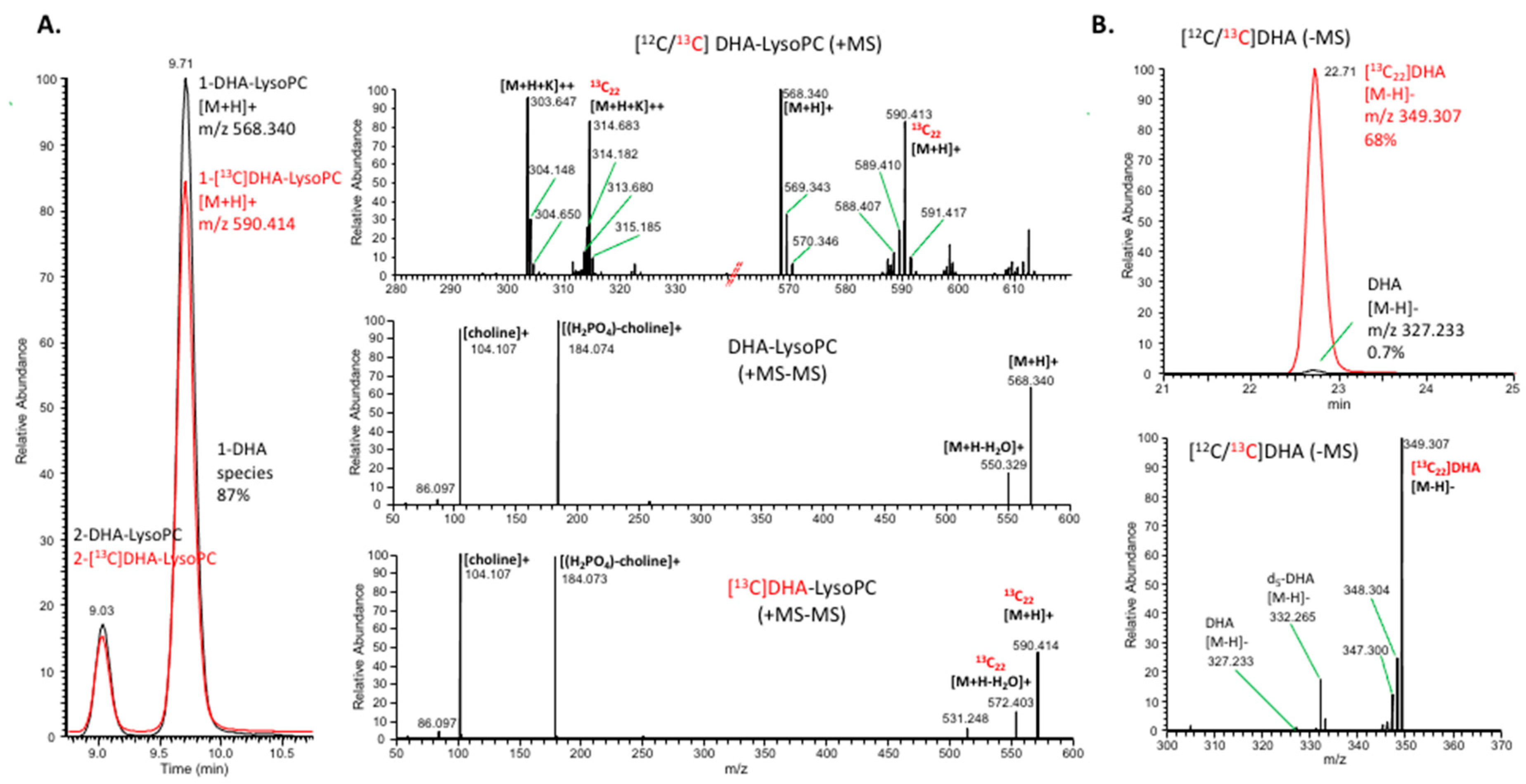
2.3. Uptake of Unesterified DHA and DHA-lysoPC
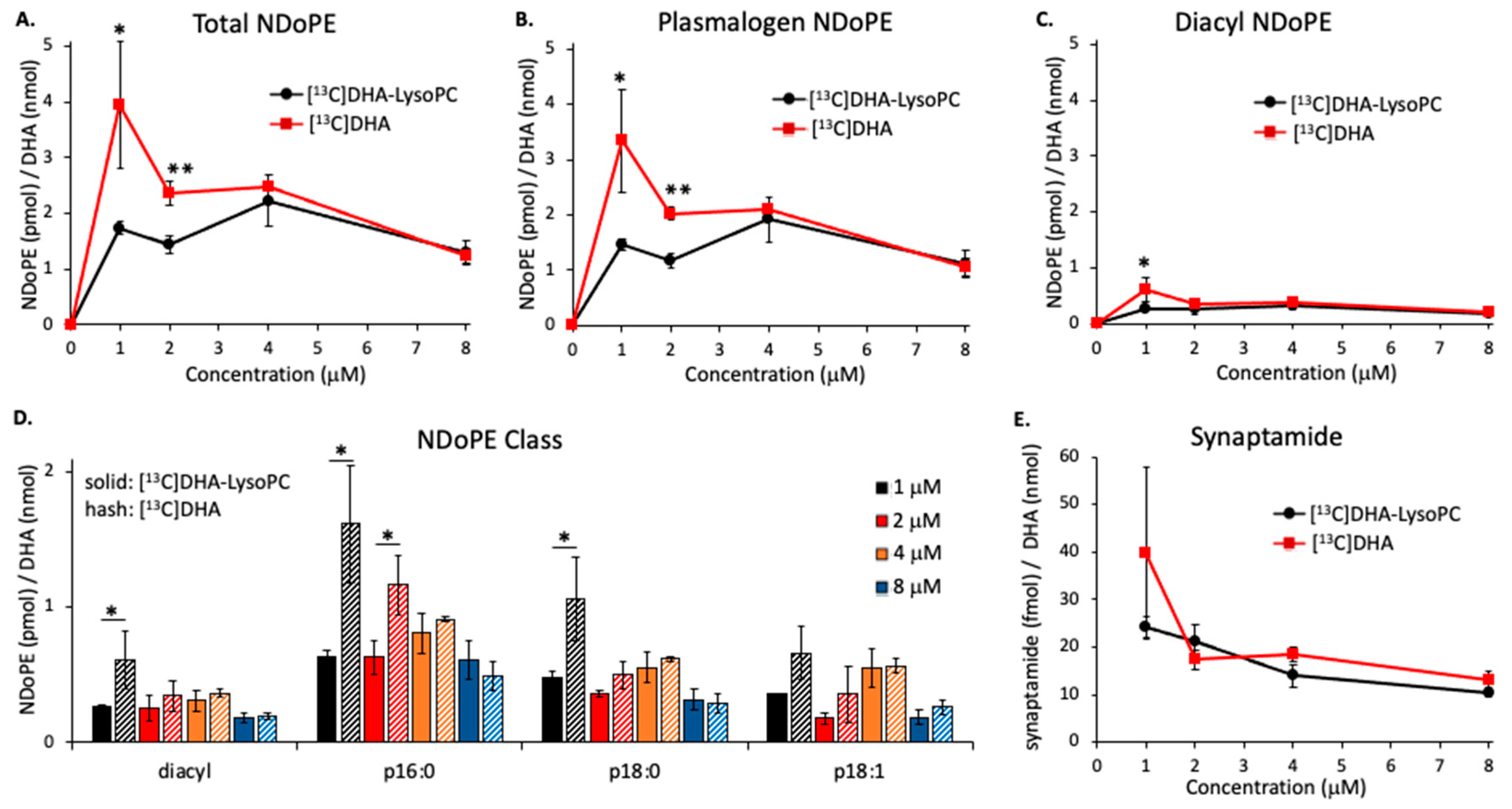
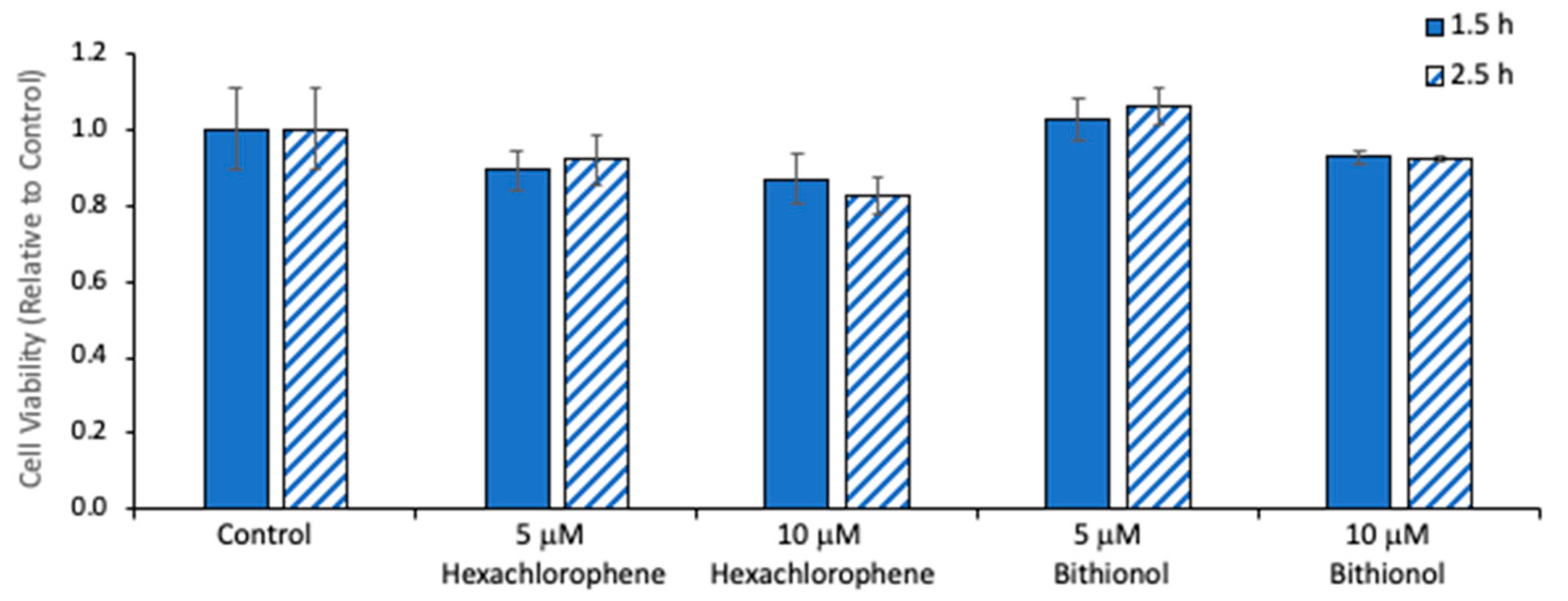
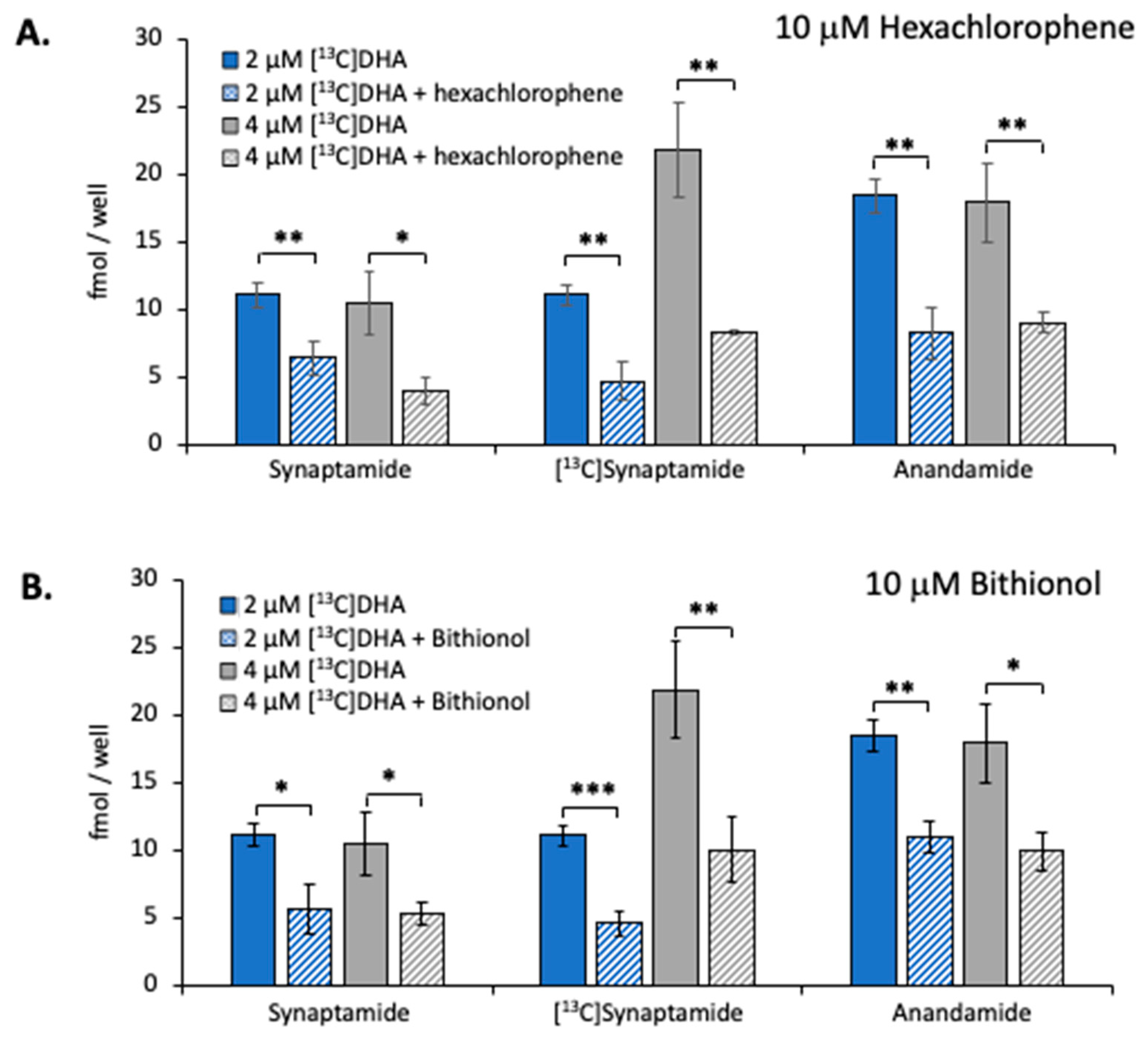
2.4. Effect of NAPE-PLD Inhibition on Synaptamide Production
3. Discussion
4. Materials and Methods
4.1. Substrate Lipids and Internal Standards
4.2. Cell Culture and Incubation with Lipid Substrates
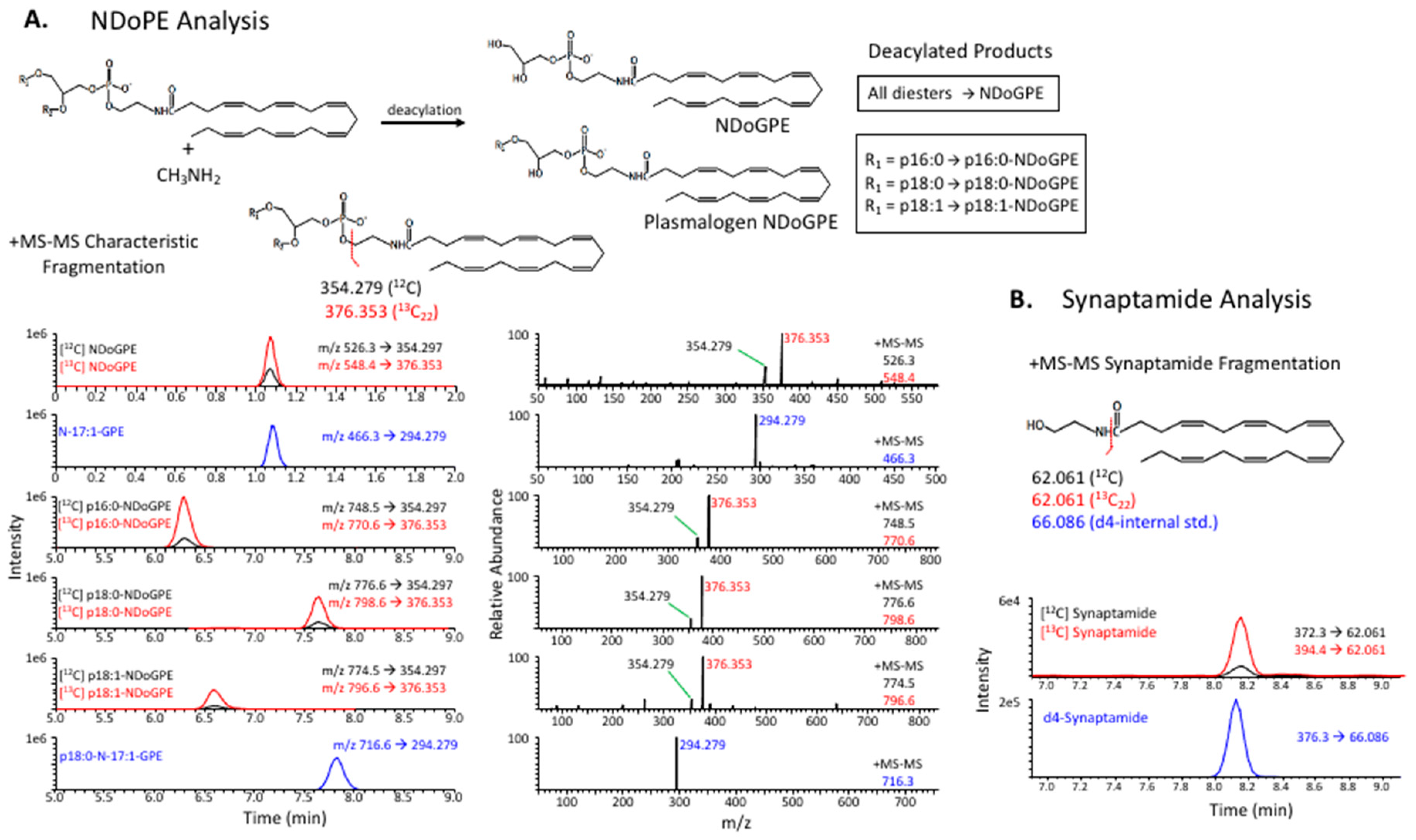
4.3. Deacylated NDoPE Analysis
4.4. Synaptamide Analysis
4.5. Total Lipid Analysis
4.6. NAPE-PLD Inhibition
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BHT | t-Butyl hydroxytoluene |
| DCM | Dichloromethane |
| DHA | Docosahexaenoic acid |
| DHA-lysoPC | Docosahexaenoyl lysophosphatidylcholine |
| DMEM | Dulbecco’s minimum essential medium |
| DMSO | Dimethyl sulfoxide |
| FAAH | Fatty acid amide hydrolase |
| FBS | Fetal bovine serum |
| GC | Gas–liquid chromatography |
| GPC | Glycerophosphorylcholine |
| GPE | Glycerophosphorylethanolamine |
| LC | Liquid chromatography |
| LC–MS | Liquid chromatography/mass spectrometry |
| lysoPC | Lysophosphatidylcholine |
| NAE | N-Acylethanolamine |
| NAPE | N-Acylphosphatidylethanolamine |
| NAPE-PLD | N-Acylphosphatidylethanolamine-phospholipase D |
| NDoGPE | N-Docosahexaenoylglycerolphosphorylethanolamine |
| NDoPE | N-Docosahexaenoylphosphatidylethanolamine |
| PBS | Phosphate-buffered saline |
| PE | Phosphatidylethanolamine |
| pNDoPE | Plasmalogen-NDoPE |
| SPE | Solid-phase extraction |
| Synaptamide | N-Docosahexanoylethanolamine |
References
- Kim, H.-Y.; Spector, A.A. N-docosahexanoylethanolamine: A neurotrophic and neuroprotective metabolite of docosahexaenoic acid. Mol. Asp. Med. 2018, 64, 34–44. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Moon, H.S.; Cao, D.; Lee, J.; Kevala, K.; Jun, S.; Lovinger, D.; Akbar, M.; Huang, B.X. N-docosahexaenoylethanolamide promotes development of hippocampal neurons. Biochem. J. 2011, 435, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, M.A.; Katakura, M.; Kharebava, G.; Kevala, K.; Kim, H.-Y. N-docosahexaenoylethanolamine is a potent neurogenic factor for neural stem cell differentiation. J. Neurochem. 2013, 125, 869–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, T.; Chen, H.; Kevala, K.; Lee, J.; Kim, H.-Y. N-docosahexaenoylethanolamine ameliorates LPS-induced neuroinflammation via cAMP/PKA-dependent signaling. J. Neuroinflamm. 2016, 13, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Huang, B.X.; Kwon, H.S.; Rashid, M.A.; Kharebava, G.; Desai, A.; Patnaik, S.; Marugan, J.; Kim, H.-Y. Orphan GPR110 (ADGRF1) targeted by N-docosahexaenoylethanolamine in development of neurons and cognitive function. Nat. Commun. 2016, 7, 13123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devane, W.A.; Hanuš, L.; Breuer, A.; Pertwee, P.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Ettinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Felder, C.C.; Briley, E.M.; Axelrod, J.; Simpson, J.T.; Mackie, K.; Devane, W.A. Arachidonoyl ethanolamide, an endogenous cannabinoid eicosanoid, binds to the cloned human cannabinoid receptor and stimulates receptor-mediated signal transduction. Proc. Natl. Acad. Sci. USA 1993, 90, 7656–7660. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, D.G.; Chin, S.A. Enzymatic synthesis and degradation of anandamide, a cannabinoid receptor agonist. Biochem. Pharmacol. 1993, 46, 791–796. [Google Scholar] [CrossRef]
- Devane, W.A.; Axelrod, J. Enzymatic synthesis of anandamide, an endogenous ligand for the cannabinoid receptor, by brain membranes. Proc. Natl. Acad. Sci. USA 1994, 91, 6698–6701. [Google Scholar] [CrossRef] [Green Version]
- Kruszka, K.A.; Gross, R.W. The ATP- and CoA-independent synthesis of arachidonoylethanolamide. J. Biol. Chem. 1994, 269, 14345–14348. [Google Scholar]
- Schmid, H.H.O.; Schmid, P.C.; Natarajan, V. N-acylation-phosphodiesterase pathway and cell signaling. Chem. Phys. Lipids 1996, 80, 133–142. [Google Scholar] [CrossRef]
- Schmid, H.H.O.; Schmid, P.C.; Natarjan, V. N-acylated glycerophospholipids and their derivatives. Prog. Lipid Res. 1990, 29, 1–43. [Google Scholar] [CrossRef]
- DiMarzo, V.; Fontana, A.; Cadas, H.; Schnelli, S.; Cimino, G.; Schwartz, J.C.; Piomelli, D. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 1994, 372, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadas, H.; Gaillet, S.; Beltramo, M.; Venance, L.; Piomelli, D. Biosynthesis of an endogenous cannabinoid precursor in neurons and its control by calcium and cAMP. J. Neurosci. 1996, 16, 3934–3942. [Google Scholar] [CrossRef] [Green Version]
- Cadas, H.; Tomaso, E.D.; Piomelli, D. Occurrence and biosynthesis of endogenous cannabinoid precursor, N-arachidonoyl phosphatidylethanolamine, in rat brain. J. Neurosci. 1997, 17, 1226–1242. [Google Scholar] [CrossRef] [Green Version]
- Suguira, T.; Kondo, S.; Sukagawa, A.; Tonegawa, T.; Nakane, S.; Yamashita, A.; Ishima, Y.; Waku, K. Transacylase-mediated and phosphodiesterase-mediated synthesis of N-arachidonoylethanolamine, an endogenous cannabinoid-receptor ligand, in rat brain microsomes. Comparison with synthesis from free arachidonic acid and ethanolamine. Eur. J. Biochem. 1996, 240, 53–62. [Google Scholar] [CrossRef]
- Hillard, C.J.; Campbell, W.B. Biochemistry and pharmacology of arachidonoylethanolamide, a putative endogenous endocannabinoid. J. Lipid Res. 1997, 38, 2383–2398. [Google Scholar]
- Natarajan, V.; Reddy, P.V.; Schmid, P.C.; Schmid, H.H.O. On the biosynthesis and metabolism of N-acylethanolamine phospholipids in infarcted dog heart. Biochim. Biophys. Acta 1981, 664, 445–448. [Google Scholar] [CrossRef]
- Reddy, P.V.; Natarajan, V.; Schmid, P.C.; Schmid, H.H.O. N-acylation of dog heart ethanolamine phospholipids by transacylase activity. Biochim. Biophys. Acta 1983, 750, 472–480. [Google Scholar] [CrossRef]
- Natarajan, V.; Schmid, P.C.; Schmid, H.H.O. N-acylethanolamine phospholipid metabolism in normal and ischemic rat brain. Biochim. Biophys. Acta 1986, 878, 32–41. [Google Scholar] [CrossRef]
- Uyama, T.; Ikematsu, N.; Inoue, M.; Shinohara, N.; Jin, X.H.; Tsuboi, K.; Tonai, T.; Tokumura, A.; Ueda, N. Generation of N-acylphosphatidylethanolamine by members of the phospholipase A/acyltransferase (PLA/AT) family. J. Biol. Chem. 2012, 287, 31905–31919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisogno, T.; Delton-Vanderbroucke, I.; Milone, A.; Lagarde, M.; DiMarzo, V. Biosynthesis and inactivation of N-arachidonoyl ethanolamine (anandamide) and N-docosahexaenoyl ethanolamine in bovine retina. Arch. Biochem. Biophys. 1999, 370, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Urquhart, P.; Wang, J.; Woodward, D.F.; Nicolaou, A. Identification of prostamides, fatty acyl ethanolamides and their biosynthetic precursors in rabbit cornea. J. Lipid Res. 2015, 56, 1419–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiès, F.; Pillon, C.; Molière, P.; Lagarde, M.; Lecerf, J. Preferential incorporation of sn-2 lysoPC DHA over unesterified DHA in the young rat brain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1994, 36, R1273–R1279. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, S.I.; Chang, M.C.J.; Spector, A.A. Delivery and turnover of plasma-derived and essential PUFAs in mammalian brain. J. Lipid Res. 2001, 42, 678–685. [Google Scholar] [PubMed]
- Nguyen, L.N.; Ma, D.; Shui, G.; Wong, P.; Cazenave-Gassiot, A.; Nguyen, L.N.; Zhang, X.; Wenk, M.; Goh, E.L.K.; Silver, D.L. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature 2014, 509, 503–506. [Google Scholar] [CrossRef]
- Bazinet, R.P.; Bernoud-Hubac, N.; Lagarde, M. How the plasma lysophospholipid and unesterified fatty acid pools supply the brain with docosahexaenoic acid. Prostaglandins Leukot. Essent. Fat. Acids 2019, 142, 1–3. [Google Scholar] [CrossRef]
- Kharebava, G.; Rashid, M.A.; Li, J.W.; Sarkar, S.; Kevala, K.; Kim, H.-Y. N-docosahexaenoylethanolamine regulates Hedgehog signaling and promotes growth of cortical axons. Biol. Open 2015, 4, 1660–1670. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, G.; Zarrow, J.E.; Mashhadi, Z.; Flynn, C.R.; Vinson, P.; Weaver, C.D.; Davies, S.S. Symmetrically substituted dichlorophenes inhibit N-acyl-phosphatidylethanolamine phospholipase D. J. Biol. Chem. 2020, 295, 7289–7300. [Google Scholar] [CrossRef] [Green Version]
- Croset, M.; Brossard, N.; Polette, N.; Lagard, M. Characterization of plasma unsaturated lysophosphatidylcholines in human and rat. Biochem. J. 2000, 345, 61–67. [Google Scholar] [CrossRef]
- Bowden, J.A.; Heckert, A.; Ulmer, C.Z.; Jones, C.M.; Koelmel, J.P.; Abdullah, L.; Ahonen, L.; Alnouti, Y.; Armando, A.M.; Asara, J.M.; et al. Harmonizing lipidomics: NIST interlaboratory comparison exercise for lipidomics using SRM 1950–Metabolites in Frozen Human Plasma. J. Lipid Res. 2017, 58, 2276–2288. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.A.; Johnson, R.A.; Corkey, B.; Kamp, F. Fatty acid transport. The diffusion mechanism in model and biological membranes. J. Mol. Neurosci. 2001, 16, 99–108. [Google Scholar] [CrossRef]
- Jay, A.G.; Simard, J.N.; Huang, N.; Hamilton, J.A. SSO and other putative inhibitors of FA transport across membranes by CD36 disrupt intracellular metabolism but do not affect FA translocation. J. Lipid Res. 2020, 61, 790–807. [Google Scholar] [CrossRef] [Green Version]
- Pownall, H.J. Commentary on SSO and other putative inhibitors of FA transport across membranes by CD36 disrupt intracellular metabolism but do not affect FA translocation. J. Lipid Res. 2020, 61, 595–597. [Google Scholar] [CrossRef] [Green Version]
- Glatz, J.F.C.; Luiken, J.J.F.P.; Bonen, A. Involvement of membrane-associated proteins in the acute regulation of cellular fatty acid uptake. J. Mol. Neurosci. 2001, 16, 123–132. [Google Scholar] [CrossRef]
- Veerkamp, J.H.; Zimmerman, A.W. Fatty acid binding proteins of nervous tissue. J. Mol. Neurosci. 2001, 16, 133–142. [Google Scholar] [CrossRef]
- Glatz, J.F.C.; Luiken, J.J.F.P. Time for a détente in the war on the mechanism of cellular fatty acid uptake. J. Lipid Res. 2020, 61, 1300–1303. [Google Scholar] [CrossRef]
- Epps, D.E.; Natarajan, V.; Schmid, P.C.; Schmid, H.H.O. Accumulation of N-acylethanolamine glycerophospholipids in infarcted myocardium. Biochim. Biophys. Acta 1980, 618, 420–430. [Google Scholar] [CrossRef]
- Natarajan, V.; Schmid, P.C.; Reddy, P.V.; Zuzarte-Augustin, M.L.; Schmid, H.H.O. Biosynthesis of N-acylethanolamine phospholipids by dog brain preparations. J. Neurochem. 1983, 41, 1303–1312. [Google Scholar] [CrossRef]
- Natarajan, V.; Schmid, P.C.; Reddy, P.V.; Zuzarte-Augustin, M.L.; Schmid, H.H.O. Occurrence of N-acylethanolamine phospholipids in fish brain and spinal cord. Biochim. Biophys. Acta 1985, 835, 426–433. [Google Scholar] [CrossRef]
- Leishman, E.; Mackie, K.; Luquet, S.; Bradshaw, H.B. Lipidomic profile of a NAPE-PLD KO mouse provides evidence for a broader role of this enzyme in lipid metabolism in the brain. Biochim. Biophys. Acta 2016, 1861, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Simon, G.M.; Cravatt, B.F. Endocannabinoid synthesis proceeding through glycerophospho-N-acylethanolamine and a role for alpha/beta-hydrolase 4 in this pathway. J. Biol. Chem. 2006, 281, 26465–26472. [Google Scholar] [CrossRef] [Green Version]
- Simon, G.M.; Cravatt, B.F. Characterization of mice lacking candidate N-acyl ethanolamine biosynthetic enzymes provides evidence for multiple pathways that contribute to endocannabinoid pathways in vivo. Mol. BioSyst. 2010, 6, 1411–1418. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, L.; Harvey-White, J.; Osei-Hyiaman, D.; Razdan, R.; Gong, Q.; Chan, A.C.; Zhou, Z.; Huang, B.X.; Kim, H.-Y.; et al. A biosynthetic pathway for anandamide. Proc. Natl. Acad. Sci. USA 2006, 103, 13345–13350. [Google Scholar] [CrossRef] [Green Version]
- Hachem, M.; Géloën, A.; Lo Van, A.; Fourmaux, B.; Fenart, L.; Gosselet, F.; Da Silva, P.; Breton, G.; Lagarde, M.; Picq, M.; et al. Efficient docosahexaenoic acid uptake by the brain from a structured phospholipid. Mol. Neurosci. 2016, 53, 3205–3215. [Google Scholar] [CrossRef]
- Triebl, A.; Weissengruber, S.; Trötzmüller, M.; Lankmayr, E.; Köfeler, H. Quantitative analysis of N-acylphosphatidylethanolamine molecular species in rat brain using solid-phase extraction combined with reversed-phase chromatography and tandem mass spectrometry. J. Sep. Sci. 2016, 39, 2474–2480. [Google Scholar] [CrossRef]
- Guo, L.; Amamath, V.; Davies, S.S. A liquid chromatography–tandem mass spectrometry method for measurement of N-modified phosphatidylethanolamines. Anal. Biochem. 2010, 405, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Kim, H.-Y. Alterations in hippocampal phospholipid profile by prenatal exposure to ethanol. J. Neurochem. 2004, 89, 1368–1377. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kevala, K.; Lagarde, M.; Spector, A.A.; Kim, H.-Y. Biosynthesis of N-Docosahexanoylethanolamine from Unesterified Docosahexaenoic Acid and Docosahexaenoyl-Lysophosphatidylcholine in Neuronal Cells. Int. J. Mol. Sci. 2020, 21, 8768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228768
Kevala K, Lagarde M, Spector AA, Kim H-Y. Biosynthesis of N-Docosahexanoylethanolamine from Unesterified Docosahexaenoic Acid and Docosahexaenoyl-Lysophosphatidylcholine in Neuronal Cells. International Journal of Molecular Sciences. 2020; 21(22):8768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228768
Chicago/Turabian StyleKevala, Karl, Michel Lagarde, Arthur A. Spector, and Hee-Yong Kim. 2020. "Biosynthesis of N-Docosahexanoylethanolamine from Unesterified Docosahexaenoic Acid and Docosahexaenoyl-Lysophosphatidylcholine in Neuronal Cells" International Journal of Molecular Sciences 21, no. 22: 8768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228768