Food-Derived Opioid Peptides in Human Health: A Review




Department of Food Science and Biotechnology, College of Agriculture and Life Sciences, Kangwon National University, Chuncheon 200-701, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(22), 8825; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228825
Submission received: 20 October 2020
/
Revised: 19 November 2020
/
Accepted: 19 November 2020
/
Published: 21 November 2020
(This article belongs to the Special Issue Peptides for Health Benefits 2020)
Abstract
:World Health Organization data suggest that stress, depression, and anxiety have a noticeable prevalence and are becoming some of the most common causes of disability in the Western world. Stress-related disorders are considered to be a challenge for the healthcare system with their great economic and social impact. The knowledge on these conditions is not very clear among many people, as a high proportion of patients do not respond to the currently available medications for targeting the monoaminergic system. In addition, the use of clinical drugs is also associated with various side effects such as vomiting, dizziness, sedation, nausea, constipation, and many more, which prevents their effective use. Therefore, opioid peptides derived from food sources are becoming one of the safe and natural alternatives because of their production from natural sources such as animals and plant proteins. The requirement for screening and considering dietary proteins as a source of bioactive peptides is highlighted to understand their potential roles in stress-related disorders as a part of a diet or as a drug complementing therapeutic prescription. In this review, we discussed current knowledge on opioid endogenous and exogenous peptides concentrating on their production, purification, and related studies. To fully understand their potential in stress-related conditions, either as a drug or as a therapeutic part of a diet prescription, the need to screen more dietary proteins as a source of novel opioid peptides is emphasized.
1. Introduction
Unhealthy lifestyle and the consumption of an unhealthy diet have been major causes of non-communicable diseases (NCD) in recent years. According to a World Health Organization report, 350 million people suffer from depression, in which 15–25% are Western-type communities [1]. Along with depression, anxiety, stress, cardiovascular diseases, and high blood pressure are becoming part of the compelling health problems worldwide [2]. For several years, sleep apnea, stress, and anxiety responses are mutual and related to each other. Stress is a response by the body to restore homeostatic balance, and these responses may cause damage or may lead to disease conditions. Several diseases arise as a consequence of stress having co-morbidity with sleep disorders [3], In particular, prolonged or persistent stress contributes to elevated hormones such as cortisol, the “stress hormone”, and decreased serotonin and other neurotransmitters in the brain, including dopamine, which have been correlated with depression. As these chemical systems function properly, they control biological processes such as sleep, appetite, energy, and sex drive, and they enable normal moods and emotions to be expressed. However, if the stress response fails to shut down and reset after a stressful situation has passed, it may lead to depression in susceptible individuals.
Sleep problems lead to issues such as reduction of appetite, a decrease of attention, unstable mood, and fatigue. According to statistics from the Mental Health Services administration (USA), 12 months of major depressive disorder in 2017 was around 13.3% for adolescents and 7.1% for adults [4,5]. It was reported that anxiety and depression disorders are more common in women as compared to men with an approximate 2:1 ratio during women’s reproductive years [4,5]. Furthermore, according to a worldwide survey, 45.7% of individuals with long-term major depressive disorders had a lifetime history of one or more than one anxiety disorder [6]. The importance of understanding stress disorders for health especially considering youth should be overemphasized [7].
Food proteins have long been used as good sources of potent bioactive peptides for preventing, managing, and treating human health [8,9]. Food proteins are biomolecules that are involved in different biological functions for improving human health, and bioactive peptides are encrypted in the sequences of proteins and are released at the time of digestion and play important roles in improving health [10,11]. They may have two or several proteinogenic amino acid groups linked with each other by peptide bonds and are released from their native proteins when fermented or treated with enzymes [12,13,14].
Currently available clinical treatments for stress-related disorders are also associated with various side effects such as vomiting, dizziness, sedation, nausea, and constipation. Therefore, nutraceuticals are becoming a promising target, as they are derived from food naturally and are known as food with medicinal benefits. These foods not only contain basic nutrients but are also rich in probiotics, antioxidants, polyphenols, bioactive compounds, and beneficial fatty acids and are known as functional foods. Their promising therapies and regulatory functions with active compounds may become a powerful tool against synthetic drugs [15], but a lot of studies are still needed to understand their roles. Hence, promoting healthy dietary intake has become a central part of the world health organization’s plan of action to prevent and monitor NCDs [8].
Opioid Peptides
Opioids were first identified in 1975 and consist of small molecules of 5–80 amino acids [16]. They diffuse locally and act on another adjacent neuron in much lower concentration as compared with neurotransmitters and give a more prolonged response [17]. Opioid peptides bind with opioid receptors μ-, κ-, and δ- for activation [18]. They are known for a wide range of functions such as neuromodulation, sleepiness [19], and pain modulation [20,21]. Except for casexins and lactoferroxins, which are opioid antagonists, the majority of peptide ligands are opioid agonists [22,23]. Opioid peptides can be categorized into endogenous or exogenous peptides based on their origin: endogenous opioid peptides that are produced by the body itself and exogenous opioid peptides that are produced after the ingestion of food when food proteins are digested by body enzymes [24,25,26]. Meanwhile, the number of studies on peptides related to stress is also increasing exponentially, which supports our interest in studying bioactive peptides and their roles in human health (Figure 1). Additionally, several attempts in human, as well as translational animal studies, have confirmed the potential of opioids in stress and related condition. The exponential increase in the search for bioactive peptides from food is also due to their low safety concerns and slow clearance from body tissues compared to synthetic drugs [27].
Here, we will summarize the current literature on opioid peptides, their classification as endogenous and exogenous, production methods, mechanism of action, as well as some opioid-like well-known peptides and their potential roles in stress-related conditions (Table 1).
So far, the potential roles of opioid-like peptides explained in Table 1 in stress-related conditions as well as in human health are well known, but their affinities as opioid peptides are still unclear. However, they are reported to exert opioid-like behaviors, but still, the mechanism of action is not well known. Therefore, further studies should be done to investigate their affinities as well as the mechanism of action as opioid peptides.
2. Classification of Opioid Peptides
They are divided into two categories as endogenous opioid peptides that are self-produced by the body and exogenous opioid peptides that are produced by different food sources such as plants and animals [25,26].
2.1. Endogenous Opioid Peptides
The endogenous opioid peptides are naturally produced in the mammalian system, which can operate as hormones (secreted by the gland and delivered to the target tissues) or neuromodulators (secreted by nerve cells and functioning in the central and peripheral nervous systems) [51]. In 1975 [16], the first endogenous ligand for opioid receptors was discovered and named as Enkephalins, and later, other endogenous peptides named Endorphins, Endomorphins, and Dynomorphins [10,52] were introduced. Opioid peptides that contain the conserved Tyr-Gly-Gly-Phe sequence at their N terminus are known as typical opioid peptides [51]. Therefore, for better understanding, Table 2A,B contains the amino acid sequences of endogenous opioid peptides, along with various binding affinities of endomorphin analogues. As it is well known, there are three opioid receptor types, µ-opioid receptor (MOR), δ-opioid receptor (DOR), and κ-opioid receptor and (KOR), which are responsible for the physiological and pharmacological effects of opioid peptides. Site-directed mutagenesis, receptor chimaera experiments, and NMR data have shown that MOR are the main opioid receptors, and μ-selectivity is primarily characterized by the second and third extracellular loops, as well as the intracellular carboxyl termini and extracellular amino termini of the MOR [53]. Apart from this, very significant features of the μ-selective agonists are the inclusion of Tyr1, Pro2, or d-Ala2 lipophilic residues at the third or fourth position and the amidation at the C-terminal [54]. So, with this theoretical basis, it would be possible to determine the conformational changes of the Endomorphins (EMs) belonging to the ligand–receptor binding and, eventually, to conjecture the detailed details of the MOR selectivity mechanism.
Enkephalins opioid peptides are present in the pituitary gland, brain, gastrointestinal tract, and kidney, and they are subdivided into two classes: Met-enkephalins and Leu-enkephalins. Another class of endogenous opioid peptides are endorphins, which are subdivided into four groups: α, β, γ, and σ. All are produced in the hypothalamus, pituitary gland, and in different parts of the nervous system and brain. Among all of them, β-endorphins are the most powerful one and play a crucial role as a neuromodulator [55]; they help to alleviate stress, body pain, and anxiety behaviors [55,56]. Endomorphins consist of types 1 and 2 endomorphins and dynorphins; both are located in the central nervous system and play significant roles in pain and stress-related conditions.
Using amino acid substitution, addition, deletion, cyclization, or the hybridization of two ligands, endogenous peptides have been modified into semisynthetic analogues to incorporate conformational constraints and make them more potent to be used as clinical analgesics [73]. Endomorphines have been transformed into analogues, which have improved protease stability by the addition of unnatural amino acids accompanied by cyclization [73,74,75]. Modifications of leu-enkephaline by the substitution, addition, and deletion of amino acids have resulted in a variety of agonists with improved δ receptor selectivity [51]. Extensive research has been done to synthesize analogues with the desired characteristics, which are not addressed in this review, but this information is available in different articles focusing only on therapeutics [20,73].
2.2. Food Derived Exogenous Opioid Peptides
These peptides are also known as exorphins having morphine-like activity, and they are derived exogenously, outside the body via different food sources. Dietary proteins are known as one of the essential sources of opioids because of their structural similarities to endogenous opioids [76]. Accumulating evidence suggests that opioid receptors may recognize peptides derived from the enzymatic hydrolysis of food proteins as they carry amino acid sequences with conserved tyrosine residue at the N terminus, and therefore, some of these opioid peptides are presented in Table 3 exhibit opioid activity along with physiological activity [77]. However, bovine α-casein peptides lacking tyrosine at the amino terminus Arg-Tyr-Leu-Gly-Tyr-Leu-Glu have also shown opioid activity [22]. To date, opioid peptides originating from animal proteins have been established as binding to μ receptors and those from plant proteins to δ receptors [78], except for soymorphins. Mostly exogenous opioid peptides are generated in the gastrointestinal tract and are absorbed in the bloodstream. They are also known to resist breakdown by intestinal enzymes such as proteases and can cross the blood–brain barrier to interact with opiate receptors [79]. There are various sources of these food-derived exogenous opioid peptides such as casein from human milk [19], β casein (buffalo milk) [25], and β-casomorphin (parmesan cheese [80], cheddar cheese [81]). Milk protein fermentation with lactic acid bacteria (LAB) is a desirable method for the production of functional foods enriched by bioactive peptides given its low cost and a good nutritional picture of fermented generated milk [82]. α-casein and β-casein are known as a good source of exogenous opioid peptides [76]. Milk was indicated to have opioid activity in 1979 [83], and morphine was isolated from milk as a drug at concentrations of 200 to 500 ng/L [84]. The opioid activity was attributed to the presence of β-casein [85] and f90-96 (Arg-Tyr-Leu-Gly-Tyr-Leu-Glu) and f90-95 (Arg-Tyr-Leu-Gly-Tyr-Leu) β-casein [86] peptides corresponding to f60-66 (Tyr-Pro-Phe-Pro-Gly-Pro-Ile, β-casomorphin-7). The opioid activity was also shown by the sequence corresponding to f91-96 (Tyr-Leu-Gly-Tyr-Leu-Glu) and f91-95 (Tyr-Leu-Gly-Tyr-Leu) amino acid residues, and Arg-Tyr-Leu-Gly-Tyr-Leu-Glu was the most potent [86]. β-casomorphin-7 from bovine β-casein was the first identified opioid peptide (Tyr-Pro-Phe-Pro-Gly-Pro-Ile) [87] and is known as the most potent opioid peptide in different βb-casomorphins (6, 5, and 4). As a result, βh-casomorphine-4,-5,-6 and-8 with Tyr-Pro-Phe-amino-termine were tested for opioid activity [57]. Based on the primary structure of human β-casein (βh-casein) and the sequence comparison with βb-casein, 10 residual shifted alignment relationships and 47% identity were established [57,88]. Moreover, β-casomorphins (BCM), which are produced by β-casein [89] (region 57–70), have shown potential effects on brain functions [89], calming, and sleep of infants [90] as well as in the modulation of behaviors such as anxiety [91]. Meanwhile, both βh-casomorphines and βb-casomorphines bind particularly to μ receptors, with the highest affinity for μ receptors and the lowest affinity for κ receptors [88].
Furthermore, there are various other food sources too that are reported as a source of exogenous opioid peptides such as barley (hordein peptide [79]), wheat (Gluten (gluten exorphins), a major wheat protein complex, Gliadin (gliadorphin), Glutenin (gluten morphin) peptides [55]), while gliadorphin-7 (Tyr-Pro-Gln-Pro-Gln-Pro-Phe) derived from α-gliadin has shown opioid activity [92]. In another study, whey protein consisting of β-lactoglobulin, immunoglobulins, α-lactalbumin, lactoperoxidase, lactoferrin, etc. was reported to exert opioid-like activity [57]. In contrast, soymorphins are known as specific ligands of the μ-opioid receptor, and many soymorphins were isolated from different sources showing opioid activities such as soymorphins 5 (Tyr-Pro-Phe-Val-Val), 6 (Tyr-Pro-Phe-Val-Val-Asn), and 7 (Tyr-Pro-Phe-Val-Val-Asn-Ala) [76]. Out of all three, soymorphin 5 (Tyr-Pro-Phe-Val-Val) has shown the highest opioid activity [93]. In another study, by the amidation or esterification, (by methyl group) of the peptides at carboxyl terminals such as valmuceptin (βh-casein 51–54 amide, Tyr-Pro-Phe-Val-NH2), morphiceptin (β-casein amide, Tyr-Pro-Phe-Pro-NH2), α-lactorphin (αh-lactalbumin 50–53 amide, Tyr-Gly-Leu-Phe-NH2), β-casorphin (βh-casein 41–44 amide, Try-Pro-Ser-Phe-NH2), β-casomorphin-4 and β-casomorphin-5 amides [94,95], several opioid peptide analogues have been identified. Lactoferroxin A (Tyr-Gly-Ser-Gly-Tyr-OCH3), B (Arg-Tyr-Tyr-Gly-Tyr-OCH3), and C (Lys-Tyr-Leu-Gly-Pro-Gln-Tyr-OCH3) are opioid antagonists derived from methyl-esterified human lactoferrin peptic digest [96]. The opioid activity of some of these peptides derived from food proteins is shown in Table 4.
Hydrolysate opioid activity can be assessed using one of many available assays. The naloxone-reversible inhibition of adenylate cyclase activity [97], naloxone-reversible inhibition of electrically induced contraction of isolated organ preparation, either mouse vas deferens and guinea-pig ileum [98], receptor binding assay or radio-receptor assay [99], are widely used assays for food opioid research. The main focus of the standard opioid activity determination tests in vitro was on µ and δ receptor interactions. These experiments are based on the inhibition of electrically evoked contractions of the mouse vas deferens (MVD) and the guinea pig ileum (GPI). The opioid effect in GPI preparations is primarily mediated by µ receptors, whereas the predominant MVD receptors are of the δ type [51]. Saturation and competition studies include receptor binding assays on tissue homogenates. The affinity of various compounds to opioid receptors is defined in saturation binding studies. Competition analyses can be performed subsequently or separately to validate these findings [51].
Exogenous opioid peptides have demonstrated promising effects in various investigations and the effects of administration of these opioid peptides at various doses in different animal models are presented in Table 5. Doses and results are difficult to compare, since various animal models and routes of administration have been used by different researchers. For example, Rubiscolin–6 improves memory consolidation [100], exerts orexigenic (oral administration) [101,102] and anxiolytic effects [103], and suppresses high-fat consumption [104]. β-casomorphine induces the release of somatostatin and insulin [105] and has been shown to prolong the gastrointestinal transit time [106] as well as modulate intestinal mucus secretion [107]. Apart from the central and peripheral nervous system effects by opioid receptors, β-casomorphin-7 also improved plasma insulin and superoxide dismutase and catalase activity in diabetic rats, thereby shielding them from hyperglycemia and free radical-mediated oxidative stress [108]. Gluten exorphin B5 enhanced the secretion of prolactin [109] and gluten exorphin C enhanced exploratory activity, improved learning, and decreased anxiety [110]; thus, studies show the positive effect of opioid peptides in human health.
The IC50 value is the concentration that would inhibit the electrically-evoked maximal contractions of the organ by 50%. As explained earlier, the opioid effect in GPI is primarily mediated by µ receptors, whereas in MVD receptors are of the δ type. So, the values in Table 4 represent the minimum concentration required for the activity with their selective receptors. For example, in soymorphins 5, 6, and 7, the IC50 value is lower in GPI assay as compared to values obtained by MVD assay, which shows there selectivity toward µ opioid receptors for activity.
Some current knowledge on the biological effects observed upon the intracerebroventricular, intraperitoneal, and oral administration of exogenous opioid peptides in the animal models are highlighted in Table 5. The fact that food-derived peptides can cross the small intestine and be present in blood and tissues has been appreciated for a long time [121,122,123]. In contrast, some pieces of evidence also suggests that [124] the degree of peptide absorption decreases with increasing chain length, so peptide length is also a point of consideration.
Although these studies (Table 5) provide useful information on the possible roles of the exogenous opioid peptides, still there are shortcomings in their research: for example, the exogenous peptides applied (similar to their endogenous counterparts) are unstable and are hydrolyzed into shorter forms, which typically have distinct activities after hydrolyzation from the parent peptide administered, leading to difficulties in the prediction of outcomes. So, technologies such as probe/radiolabeling will be effective here for the administration of parent peptides before hydrolysis inside the animal or human body. However, peptides can be secured against enzymatic cleavage by inserting a structure that induces a tail probe [125], by lactam bridge [126], by stacking or clipping peptide sequences [127], or by cycling [128].
For better understanding, deeper research with an explored signal cascade mechanism at the cellular and molecular level is needed to explore food-derived opioid peptides as therapeutic mediators, functional foods, or nutraceuticals for human health promotion.
3. Production of Opioid Peptides from Food Proteins
There are various emerging improvements in the methodologies for the analysis and the development of food-derived peptides. The main methods for their discovery can be classified as a traditional, in silico approach, or integrated approaches from the present literature.
3.1. The Traditional Approach
The traditional approach focuses on enzyme selection and production of the peptide through protein hydrolysis, purification of peptides, and peptide identification.
This is an extensively used method for recovering bioactive peptides from different food sources and comprises a collection of certain protein sources of interest. The proteins were usually digested with the help of food-grade enzymes (proteases, proteinases, or peptidases) to hydrolyze proteins into peptide fragments [129,130,131,132]. In addition to enzymes, various microorganisms may be used to ferment the proteins to enhance peptide breakdown [133,134,135]. The use of fermentation is possible because of the many proteolytic enzymes they possess for degrading proteins and to satisfy their nitrogen demands. Subsequently, the fermented peptides are extracted and purified according to their structural chemistry. The fractions undergo in vitro testing to evaluate their potential health effects. In an earlier study, β-casomorphin-7 was detected for the first time in the fermented sample [136]. In another study, gluten and exorphins were identified in a hydrolysate of pepsin and thermolysin [112] as well as in hydrolysate of pepsin–trypsin–chymotrypsin [113]. Gluten exorphins A5 with a weight around 0.747–2.192 mg/kg and C of 3.201–6.689 mg/kg have recently been found in bread and pasta following the simulation of in vitro GI digestion (using pepsin–trypsin–chymotrypsin) [137]. Similarly, casomorphin releases were analyzed in milk and its products after simulated GI digestion by different enzymes [138].
The hydrolysates with beneficial health effects are purified, and the active peptides were identified. Methods such as high-pressure liquid chromatography joined with mass spectrometry, reverse-phase liquid chromatography joined with mass spectrometry, and liquid chromatography-electrospray ionization along with quantitative time-of-flight tandem mass spectrometry have been used in studies to identify the bioactive peptides present in the sample [139,140,141]. This classical approach is effective in discovering new bioactive peptides from various protein substrates.
3.2. The In Silico Approach
The in silico methods comprise the use of information gathered from databases to find out the occurrence frequency of encrypted bioactive peptides in the primary food protein structure. The protein sequences can be accessible from databases for the analysis of different bioactive peptides (Table 6). As the existence of these peptides does not naturally mean the liberation of encrypted peptides, some bioinformatics software has been created that mimic proteolytic enzyme specificities to produce silico peptide profiles. Then, these proteases can be utilized for the hydrolysis of food protein, and their potential health effects were tested to build their efficacy. This method helps differentiate the identified peptides from unknown sources of protein (Figure 2). Most in silico platforms can predict the bioactive potential of the identified peptides, and the bioactivity can be confirmed through experiments in vitro and in vivo. As the in silico approach is rapid, cost-effective, and a greater number of options are available, various researchers are using this approach [142,143,144] to identify potent bioactive peptides. Proteolysis tools such as ExPASy Peptide Cutter (http:/web.expasy.org/peptide cutter), BIOPEP, and PoPS (http://pops.csse.monash.edu.au) are used to classify the specificities of different enzymes for releasing target peptide from food. By using this approach, sequences of cereal proteins (wheat, oat, barley, and rice) display high concentrations of peptides with dipeptidyl peptidase-inhibitory, anti-thrombotic, angiotensin-converting enzyme-inhibitory, antioxidant, hypotensive, and opioid activity [145]. In recent years, these computer-based databases have been used to predict the existence of bioactive peptides in food proteins [24,146,147,148]. Yet, more research is required to better understand this approach for the prediction of various food-based bioactive peptides.
3.3. Chemical Synthesis Approach
The key chemical approaches for peptide synthesis are solution-phase synthesis (SPS) and solid-phase peptide synthesis (SPPS). SPS is generally performed by coupling single amino acids in solution. Long peptide synthesis is feasible by synthesizing short fragments of the target peptides first and compressing them to produce long peptides [149]. This SPS approach is called the fragments condensation process. In the SPS method, it is possible to deprotect and purify intermediate products to achieve high purity of the target peptide [150,151]. SPS is economical and efficiently extensible, but the long reaction time remains a drawback. Meanwhile, the SPPS approach requires peptide synthesis using resin as a support for a growing peptide chain. An amino acid’s reactive side chain and α-amino group are first covered (mostly using the fluorenylmethoxycarbonyl protecting group (Fmoc) or tert-Butoxycarbonyl (Boc)) and the amino acids C-terminus is bound to the resin [152]. The N-terminal protecting group is normally removed (or cleaved) by using trifluoroacetic acid (Boc) or by 20% piperidine in N-N-dimethylformamide (Fmoc); then, the resin is washed before the introduction of subsequent amino acids. The peptide is separated off the resin after the required sequence is completed [149]. Presently, SPPS is widely used for therapeutic peptide synthesis because of its lower manufacturing costs and advancements in chromatographic equipment [149]. Long peptide or protein chains can also be synthesized by the chemical ligation approaches. The Native Chemical Ligation (NCL) is an efficient process for ligating peptides. A non-protected peptide segment containing an N-terminal cysteine is reacted to ligate peptide fragments with another unprotected peptide to form a thioester-linked intermediate, which is later reconstructed into a peptide bond. This process enables the formation of peptides of high molecular weight such as multivalent peptide-based non-symmetric dendrimer [153] and collagen-like polymers [154]. The advantages of this process are the high strength of the starting materials in NCL, the well-established chemical methods to manufacture peptide thioesters, and the high chemo-selective nature of the peptides. In addition, various researchers have used this approach such as Hartman et al. [155] for the treatment of pathological conditions of oxidation, Meisel et al. [156,157] for the oral administration of chemically synthesized peptides to mice in inflammation and atherosclerosis, González-García et al. [158] to transform bioactive peptide from waste to valuable product, Agyei et al. [159] to produce bioactive peptides to get large-scale recovery in pharmaceuticals, and Kim et al. [160] for the purification of bioactive peptides in food industries.
3.4. The Integrated Approach
An integrated bioinformatics approach may be utilized in the detection of bioactive peptides because of limitations associated with the previous approaches. The strengths of both classical and bioinformatics approaches (Figure 3) can be combined to advance the analysis and need for peptides in health benefits and functional foods. The bioactive peptides determined in food proteins via the in silico method could be chemically synthesized, and this approach will lead to the detection of new peptides from new sources [161]. However, this is possible only if the whole sequence of a protein and the functional activities of the peptides are already known. Nonetheless, various important bioactive peptides of low concentration can further be missing if have not already been identified in the database. Therefore, alternative techniques such as peptide display methodologies were being used for the search of bioactive peptides at present.
3.5. Screening for Bioactive Peptides
After fermentation or enzymatic hydrolysis, it is usually very hard to identify particular bioactive peptides in a sample. Chemically synthesizing all of the peptides that could be found in a peptide digested for the screening purposes is costly and very laborious (if not impossible). Hence, many researchers use the bioassay fractionation approach, in which liquid chromatography separates protein digests into fractions. The fractions are analyzed, and potent fractions are further fractionated before mass spectrometry [162]. The peptides encrypted in the fractions are eventually recognized. While the approach has been successful and efficient, it typically omits the activity of lower concentrated fragments.
Using in silico platforms alone to predict bioactivity may be easy, but it may not be very accurate, as not all of the peptides predicted may be bioactive. Consequently, other researchers correlate their data collected from HPLC-MS along with many house databases that enhance the detection of less concentrated peptides [14]. However, several essential new low-concentration bioactive peptides can still be lost. In such a situation, the recombinant peptide libraries related to the coding sequence (peptide display) of these peptides can be used to identify dynamic bioactive peptides as an effective tool. Peptide display methodologies have been used as an effective research tool to track protein interactions at high throughput [163,164]. Phage display has been widely applied among most available molecular display techniques such as covalent antibody display, yeast and bacterial display, mRNA display, ribosome display, and CIS display. Phage display is a peptide selection approach that comprises the fusion of a peptide or protein with a protein coat of bacteriophage displayed on the virus surface [165]. The random peptide libraries displayed in a phage provide a functional approach to biopeptides, distinguishing peptides binding from those which are nonbinding peptides via affinity purification. The phage displayed random peptide library identification is an effective way of detecting peptides that can bind and control target molecular behaviors. This method has been practiced in identifying receptor-bound bioactive peptides [166], disease-specific antigen mimics [166,167], cell-specific peptides, non-protein-bound peptides [168], or organ-specific peptides [169], as well as in the designing of peptide-mediated drug delivery systems [165]. Consequently, analysis for bioactive peptides applying phage display technology is an advantageous approach that can be used in basic research. This way, the bioactive peptides achieved can be cloned and overexpressed to increase their quantity.
4. Mechanism of Action
A receptor is a protein that binds with a chemical messenger and brings out an intracellular response. In the case of the opioid peptide, they either bind to a receptor or are converted into smaller peptides or amino acids (Figure 4). The receptors present are G-proteins coupled receptors (GPCRs); they are known as the largest gene family among all receptors, consisting of around 1000 different genes in human and other mammals [170].
When an opioid peptide binds to a receptor molecule, several intracellular changes of molecules occur along with other changes such as enzyme activation, the opening of ion channels, and the transcription of genes [170]. G-protein coupled receptors are also known as heptahelical receptors, as they consist of seven-transmembrane spanning receptors, and their signaling is conducted via G-proteins. Opioid peptides when functioning through GPCRs bind to the external surface of the receptor, which changes the conformation of the receptor proteins, leading to intracellular changes of the G proteins with the receptors [170]. The G-protein consists of 3 subunits α, β, and γ; when G-proteins are in an inactive state, the α-subunit binds with GDP (Guanosine diphosphate) along with other two subunits, β and γ, forming a G-protein complex. After receptor activation, the G-proteins attached GDP molecule is released and converted to a GTP (Guanosine triphosphate) molecule, and the GTP-α-subunit complex is dissociated from βγ subunits, which remain attached. The Gα-subunit is now able to modulate the activity of the effector molecules such as phospholipase or adenylyl cyclase [170]. Whereas it was also seen that in some cases, βγ subunits are also able to modulate some effector activities [170]. As soon as GTP bound with the α-subunit is hydrolyzed by cellular proteins, GTP hydrolyzed to GDP [170]. Thereafter, the free α-subunit again reassociates to form a heterotrimeric complex with βγ subunits, and this complex couples with the receptor for the next cycle of G-proteins upon activation (Figure 5). There are different subfamilies of G-proteins based on α-subunit Gi, Gs, Gq, and Go [76]. Everyone has multiple members who work via different pathways. The Gαs subunits activate adenylyl cyclase, whereas Gαo and Gαi subunits inhibit the adenylyl cyclase enzyme [76]. These activations lead to a series of reactions as adenylyl cyclase bring about cAMP(Cyclic adenosine monophosphate) formation from ATP, which activates protein kinase A. Earlier protein kinase A phosphorylates various intracellular substrates that result in biological modulations [76]. Moreover, Gαq subunits activate phospholipases that produce diacylglycerol and inositol 1,4,5-triphosphate [171]. Diacylglycerol activates protein kinase C for the phosphorylation of various molecules, whereas inositol 1,4,5-triphosphate activates receptors present on the endoplasmic reticulum for the opening of the Ca2+ channel [171]. Gα12/13 leads to the activation of guanine-nucleotide factors such as RhoGTPases for the exchange of GDP to proteins leading to biological changes [170].
These opioids peptide signaling systems are very complex as compared with some conventional neurotransmitters where no multiple receptor binding occurs and only a single ligand works with different receptors such as for acetylcholine [170]. Whereas most of them bind with multiple receptors but as an addition, they are also able to bind with receptors subtypes, creating more complexity in the signaling pathways. Yet, with the complexity of receptors and mechanism of action in these bioactive peptides, they still provide a promising and successful role in various clinical trials [170].
Transport of Opioid Peptides in the Body
During digestion, digestive enzymes hydrolyze the food proteins to peptides and amino acids. Several factors influence the GI tract’s transport and absorption of peptides, including pKa, peptide size, and pH microclimate. Gastric emptying and the intestinal transit affect the location where the peptide is present along the GI tract and thus influence absorption. Peptides larger than di-tripeptides are found not easily absorbed in healthy people except in the conditions such as stress or disease when intestinal permeability is increased [172]. It has been seen that there is no absolute barrier to the intestinal mucosa and different peptides, including gluten exorphins A5 and A4, can cross the intestinal epithelium [57,137], whereas the mechanism of transfer is still not clear. In another study, it was observed that in a mammalian system, four different peptide transport systems PTS-1, PTS-2, PTS-3, and PTS-4 were involved that can transfer peptides, including food-derived peptides, from the peripheral circulation to the central nervous system through the blood–brain barrier [173]. In contrast, all four PTS-1 systems carry opioid peptides, including Tyr-MIF-1 met-enkephalin and leu-enkephalin [173]. Food-derived opioids absorbed in the gastrointestinal tract initially interact with receptors that are present on the enteric nervous system (ENS) and thus influence GI functions. The ENS is a network of nerve cells located in the GI tract wall, which control motility and secretion and regulate digestion, absorption, and immunomodulation [174]. In contrast, the glycosylation of peptides has also shown promising results in the transportation of peptides via the GLUT1 glucose carrier, along with the glycosylated analogues of dermorphin and met-encephalin [175]. However, due to peptidase activity, the half-life of opioid peptides in the blood is low. Endogenous opioids leu-enkephalin and dynorphin-A (1–13) have 6.7 min and 1 min half-lives [176,177]. In contrast, dermorphin exhibits a longer half-life than Enkephalins [178], and the half-life can be extended by binding these peptides to carrier proteins such as transferrin [179] or albumin [180]; some of the half-life stability data of opioid peptides are shown in Table 7. Therefore, in vivo half-life investigations of exogenous opioid peptides in blood need investigation for future research benefits.
5. Clinical and Animal Studies Related to Exogenous Opioid Peptides in Stress-Related Conditions
There are various preclinical and clinical studies available to support the role of several bioactive peptides in animals as well as in humans. Interestingly, as of now, opioid peptides are known as a potential target for the development of various new therapies related to stress disorders [184,185]. Here, we highlighted some pieces of evidence to support the role of food-derived peptides in stress-related conditions.
Stress, Anxiety, and Depression
After the discovery of endogenous opioid peptides, experiments were performed to investigate exogenous peptides and their role in animal models. Studies related to exogenous opioid peptides have shown positive impacts on human health.
For instance, Lister et al. [91] had used various nociceptive models, and intracerebral (i.c.) or intracerebroventricular (i.c.v) routes were tested in which it was observed that various β-casomorphins (-3, -4, -5, -6, and -7) had shown anxiolytic effects. Limit et al. have found the effect of peptides isolated from soybean on brain functions [186]. Similarly, Bernet et al. explained the role of fish hydrolysates (Gabolysat PC60) on the levels of GABA and their sympathoadrenal activity leading anxiolytic effects in rats [187].
Additionally, rubiscolin-6 isolated from spinach RuBisCo was administered orally in mice and found to have an anxiolytic effect arbitrated by dopamine receptors [103] as well as a reduction of nociception in mice [188].
It was also seen that certain milk-based β-casomorphins interact with the receptors of opiates to affect the absorption of food in different conditions such as stress and anxiety [189]. Yin et al. has reported the protective role of β-casomorphins against oxidative stress [108]. In another, Chesnokova et al. [120] and Kaneko et al. [119] have shown the potential role of soymorphin-5 in the management of anxiety in mice.
On the other hand, soymorphin-5 demonstrated anxiolytic effects in mice, showing an improvement in the time mice remained in the open arms during an elevated plus-maze test [190]. In a human study, α-lactalbumin from full whey protein induced anxiolytic effects [191]. So, by going through different studies, we found that food-derived opioid peptides play a promising role in stress-related conditions as well as in human health.
6. Conclusions
In this review, we provided a detailed discussion on types of opioid peptides along with their sources, structures generated by enzyme hydrolysis of the food proteins, and their clinical pieces of evidence related to stress, anxiety, and depression. The opioid peptides have been discovered in the 1970s, and to our understanding, they hold great promise as valuable functional ingredients in healthy diets. In the modern generation, stress, anxiety, and depressive disorders are becoming a major issue, and food-derived opioid peptides showing anxiolytic and anti-stress effects can be a beneficial food substrate in maintaining a healthy population. The fermentation and enzymatic hydrolysis of food proteins are widely used in varying quantities and bioactivity to release these peptides. The amino acid profiles of various plants and animal food proteins suggest that these can become a great potential source for the production of bioactive peptides. Their effects are also dependent on their stability in blood, binding affinities, and half-lives as well as the capability of crossing the blood–brain barrier. The relative activity of these opioids depends on their affinity to the receptors, μ, δ, and κ. To date, opioid peptides from animal proteins tend to attach primarily to μ receptors, whereas those from plant proteins bind with δ receptors, except soymorphine. Most food opioid investigations were focused on widely used tissue preparations, mouse vas deferens, and guinea-pig ileum, which are unique to μ and δ receptors, respectively. It is not known whether the receptor of food-derived opioid peptides binds to the κ receptor. There is no proof in the literature of the use of rabbit vas deferens, which is known to be rich in κ opioid receptor, to confirm whether food opioids bind to this receptor [59].
On the other hand, various studies such as meta-analysis, animal model, and clinical examinations have shown the impact of these peptides on the nervous system (analgesia, antinociception, and improved memory), GI functions (increased intestinal transit time, increased appetite, and suppression of high fat intake), and increased β-oxidation and energy consumption, indicating the possibility of their use as nutraceuticals for pain relief, stress reduction, blood sugar, and obesity.
Still, to validate their roles, future consideration is needed to understand the stability of these peptides during digestion in animal and humans by in in vitro and in vivo studies as well as the health-related effects they generate across the gut–brain axis. Therefore, further research is required to understand and develop methods for the development of opioid peptides from food proteins in substantial quantities for pharmaceutical, beverage, and food use and system strategies to ensure their targeted distribution.
Author Contributions
Conceptualization: A.T., E.B.-M.D. and D.-H.O.; Writing—Original Draft Preparation: A.T.; Data curation: A.T., Writing—Review & Editing, A.T., E.B.-M.D. and D.-H.O.; Software: A.T., F.K.O., S.-J.Y.; and D.-H.O., Supervision, D.-H.O. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Brain Korea (BK) 21 Plus Project (Grant No.22A20153713433) funded by the Korean Government, South Korea.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- World Health Organization. World Mental Health Day, 10 October 2012. Available online: http://www.who.int/mental_health/management (accessed on 30 September 2020).
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Szelenberger, W.; Soldatos, C. Sleep disorders in psychiatric practice. World Psychiatry 2005, 4, 186. [Google Scholar] [PubMed]
- Substance Abuse and Mental Health Services Administration. Mental Health Services Administration. Key Substance Use and Mental Health Indicators in the United States: Results from the 2016 National Survey on Drug Use and Health (HHS Publication No. SMA 17-5044, NSDUH Series H-52); Center for Behavioral Health Statistics and Quality: Rockville, MD, USA, 2017.
- Ned, H.; Kalin, M.D. The Critical Relationship Between Anxiety and Depression. Am. J. Psychiatry 2020, 177, 365–367. [Google Scholar] [CrossRef]
- Kessler, R.C.; Sampson, N.A.; Berglund, P.; Gruber, M.; Al-Hamzawi, A.; Andrade, L.; Bunting, B.; Demyttenaere, K.; Florescu, S.; De Girolamo, G. Anxious and non-anxious major depressive disorder in the World Health Organization World Mental Health Surveys. Epidemiol. Psychiatr. Sci. 2015, 24, 210–226. [Google Scholar] [CrossRef] [Green Version]
- Gee, D.G.; Kribakaran, S. Developmental Differences in Neural Responding to Threat and Safety: Implications for Treating Youths with Anxiety. Am. Psychiatr. Assoc. 2020. [Google Scholar] [CrossRef] [PubMed]
- Poppitt, S.D. Milk proteins and human health. In Milk Proteins; Elsevier: Amsterdam, The Netherlands, 2020; pp. 651–669. [Google Scholar]
- González, S. Dietary Bioactive Compounds and Human Health and Disease. Nutrients 2020, 12, 348. [Google Scholar] [CrossRef] [Green Version]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. Nutr. 2018, 58, 2273–2284. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Oh, D.H.; Lee, B.H. Bioactive peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Park, M.H.; Kim, J.-H.; Oh, D.-H. Novel angiotensin I-converting enzyme inhibitory peptides from soybean protein isolates fermented by Pediococcus pentosaceus SDL1409. LWT 2018, 93, 88–93. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Ofosu, F.K.; Chelliah, R.; Park, M.H.; Kim, J.-H.; Oh, D.-H. Development of a soy protein hydrolysate with an antihypertensive effect. Int. J. Mol. Sci. 2019, 20, 1496. [Google Scholar] [CrossRef] [Green Version]
- Daliri, E.B.-M.; Lee, B.H.; Park, B.-J.; Kim, S.-H.; Oh, D.-H. Antihypertensive peptides from whey proteins fermented by lactic acid bacteria. Food Sci. Biotechnol. 2018, 27, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Villaluenga, C.; Hernández-Ledesma, B. Peptides for Health Benefits 2019. Int. J. Mol. Sci. 2020, 21, 2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, J.; Smith, T.; Kosterlitz, H.; Fothergill, L.A.; Morgan, B.; Morris, H. Identification of two related pentapeptides from the brain with potent opiate agonist activity. Nature 1975, 258, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Gulpinar, M.A.; Yegen, B.C. The Physiology of Learning and Memory: Role of Peptides and Stress. Curr. Protein Pept. Sci. 2004, 5, 457–473. [Google Scholar] [CrossRef]
- Wang, Y.; Van Bockstaele, E.J.; Liu-Chen, L.-Y. In vivo trafficking of endogenous opioid receptors. Life Sci. 2008, 83, 693–699. [Google Scholar] [CrossRef] [Green Version]
- Rutherfurd-Markwick, K.J. Food proteins as a source of bioactive peptides with diverse functions. Br. J. Nutr. 2012, 108, S149–S157. [Google Scholar] [CrossRef] [Green Version]
- Bodnar, R.J. Endogenous opiates and behavior: 2012. Peptides 2013, 50, 55–95. [Google Scholar] [CrossRef]
- Bagley, E.E.; Ingram, S.L. Endogenous opioid peptides in the descending pain modulatory circuit. Neuropharmacology 2020, 173, 108131. [Google Scholar] [CrossRef]
- Teschemacher, H.; Koch, G.; Brantl, V. Milk protein-derived opioid receptor ligands. Pept. Sci. 1997, 43, 99–117. [Google Scholar] [CrossRef]
- Meisel, H.; Fitzgerald, R.J. Opioid peptides encrypted in intact milk protein sequences. Br. J. Nutr. 2000, 84, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Gutiérrez-López, G.F.; Dávila-Ortiz, G. Use of proteomics and peptidomics methods in food bioactive peptide science and engineering. Food Eng. Rev. 2012, 4, 224–243. [Google Scholar] [CrossRef]
- Trivedi, M.S.; Shah, J.S.; Al-Mughairy, S.; Hodgson, N.W.; Simms, B.; Trooskens, G.A.; Van Criekinge, W.; Deth, R.C. Food-derived opioid peptides inhibit cysteine uptake with redox and epigenetic consequences. J. Nutr. Biochem. 2014, 25, 1011–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, Z.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, M.; Hattori, S.; Amano, Y.; Watanabe, M.; Nakamura, K. Blood pressure-lowering peptides from neo-fermented buckwheat sprouts: A new approach to estimating ACE-inhibitory activity. PLoS ONE 2014, 9, e105802. [Google Scholar] [CrossRef]
- Popik, P.; Bobula, B.; Janusz, M.; Lisowski, J.; Vetulani, J. Colostrinin, a polypeptide isolated from early milk, facilitates learning and memory in rats. Pharmacol. Biochem. Behav. 1999, 64, 183–189. [Google Scholar] [CrossRef]
- Gülpınar, M.A.; Özbeyli, D.; Arbak, S.; Yeğen, B.Ç. Anti-inflammatory effect of acute stress on experimental colitis is mediated by cholecystokinin-B receptors. Life Sci. 2004, 75, 77–91. [Google Scholar] [CrossRef]
- Bracci-Laudiero, L.; Aloe, L.; Lundeberg, T.; Theodorsson, E.; Stenfors, C. Altered levels of neuropeptides characterize the brain of lupus prone mice. Neurosci. Lett. 1999, 275, 57–60. [Google Scholar] [CrossRef]
- Sprick, U.; Hasenöhrl, R.; Krauth, J.; Klapdor, K.; Huston, J. Effects of chronic substance P treatment and intracranial fetal grafts on learning after hippocampal kainic acid lesions. Peptides 1996, 17, 275–285. [Google Scholar] [CrossRef]
- Sandin, J.; Georgieva, J.; Schött, P.A.; Ögren, S.O.; Terenius, L. Nociceptin/orphanin FQ microinjected into hippocampus impairs spatial learning in rats. Eur. J. Neurosci. 1997, 9, 194–197. [Google Scholar] [CrossRef]
- Manabe, T.; Noda, Y.; Mamiya, T.; Katagiri, H.; Houtani, T.; Nishi, M.; Noda, T.; Takahashi, T.; Sugimoto, T.; Nabeshima, T. Facilitation of long-term potentiation and memory in mice lacking nociceptin receptors. Nature 1998, 394, 577–581. [Google Scholar] [CrossRef]
- Hiramatsu, M.; Inoue, K. Improvement by low doses of nociceptin on scopolamine-induced impairment of learning and/or memory. Eur. J. Pharmacol. 2000, 395, 149–156. [Google Scholar] [CrossRef]
- Lynch, G.; Larson, J.; Staubli, U.; Granger, R. Variants of Synaptic Potentiation and Different Types of Memory Operations in Hippocampus and Related Structures; Oxford University Press: Washington, DC, USA, 1991. [Google Scholar]
- Chen, Q.; Patel, R.; Sales, A.; Oji, G.; Kim, J.; Monreal, A.; Brinton, R. Vasopressin-induced neurotrophism in cultured neurons of the cerebral cortex: Dependency on calcium signaling and protein kinase C activity. Neuroscience 2000, 101, 19–26. [Google Scholar] [CrossRef]
- Crawley, J.N.; Corwin, R.L. Biological actions of cholecystokinin. Peptides 1994, 15, 731–755. [Google Scholar] [CrossRef]
- Dauge, V.; Pophillat, M.; Crete, D.; Melik-Parsadaniantz, S.; Roques, B. Involvement of brain endogenous cholecystokinin in stress-induced impairment of spatial recognition memory. Neuroscience 2003, 118, 19–23. [Google Scholar] [CrossRef]
- Telegdy, G.; Adamik, A.; Glover, V. The action of isatin (2, 3-dioxoindole) an endogenous indole on brain natriuretic and C-type natriuretic peptide-induced facilitation of memory consolidation in passive-avoidance learning in rats. Brain Res. Bull. 2000, 53, 367–370. [Google Scholar] [CrossRef]
- Telegdy, G.; Kokavszky, K.; Nyerges, A. Action of C-type natriuretic peptide (CNP) on passive avoidance learning in rats: Involvement of transmitters. Eur. J. Neurosci. 1999, 11, 3302–3306. [Google Scholar] [CrossRef]
- Telegdy, G.; Kokavszky, K. The action of pituitary adenylate cyclase activating polypeptide (PACAP) on passive avoidance learning. The role of transmitters. Brain Res. 2000, 874, 194–199. [Google Scholar] [CrossRef]
- McDONALD, M.P.; Gleason, T.C.; Robinson, J.K.; Crawley, J.N. Galanin inhibits performance on rodent memory tasks. Ann. N. Y. Acad. Sci. 1998, 863, 305–322. [Google Scholar] [CrossRef]
- Counts, S.E.; Perez, S.E.; Kahl, U.; Bartfai, T.; Bowser, R.P.; Deecher, D.C.; Mash, D.C.; Crawley, J.N.; Mufson, E.J. Galanin: Neurobiologic mechanisms and therapeutic potential for Alzheimer’s disease. CNS Drug Rev. 2001, 7, 445–470. [Google Scholar] [CrossRef]
- Wrenn, C.; Marriott, L.; Kinney, J.; Holmes, A.; Wenk, G.; Crawley, J. Galanin peptide levels in hippocampus and cortex of galanin-overexpressing transgenic mice evaluated for cognitive performance. Neuropeptides 2002, 36, 413–426. [Google Scholar] [CrossRef]
- Santo-Yamada, Y.; Yamada, K.; Wada, E.; Goto, Y.-I.; Wada, K. Blockade of bombesin-like peptide receptors impairs inhibitory avoidance learning in mice. Neurosci. Lett. 2003, 340, 65–68. [Google Scholar] [CrossRef]
- Yamada, K.; Santo-Yamada, Y.; Wada, K. Stress-induced impairment of inhibitory avoidance learning in female neuromedin B receptor-deficient mice. Physiol. Behav. 2003, 78, 303–309. [Google Scholar] [CrossRef]
- Ojika, K.; Mitake, S.; Tohdoh, N.; Appel, S.H.; Otsuka, Y.; Katada, E.; Matsukawa, N. Hippocampal cholinergic neurostimulating peptides (HCNP). Prog. Neurobiol. 2000, 60, 37–83. [Google Scholar] [CrossRef]
- Gasparini, L.; Netzer, W.J.; Greengard, P.; Xu, H. Does insulin dysfunction play a role in Alzheimer’s disease? Trends Pharmacol. Sci. 2002, 23, 288–293. [Google Scholar] [CrossRef]
- Moriguchi, T.; Sakurai, T.; Nambu, T.; Yanagisawa, M.; Goto, K. Neurons containing orexin in the lateral hypothalamic area of the adult rat brain are activated by insulin-induced acute hypoglycemia. Neurosci. Lett. 1999, 264, 101–104. [Google Scholar] [CrossRef]
- Aou, S.; Li, X.-L.; Li, A.-J.; Oomura, Y.; Shiraishi, T.; Sasaki, K.; Imamura, T.; Wayner, M. Orexin-A (hypocretin-1) impairs Morris water maze performance and CA1-Schaffer collateral long-term potentiation in rats. Neuroscience 2003, 119, 1221–1228. [Google Scholar] [CrossRef]
- Janecka, A.; Fichna, J.; Janecki, T. Opioid receptors and their ligands. Curr. Top. Med. Chem. 2004, 4, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Koneru, A.; Satyanarayana, S.; Rizwan, S. Endogenous opioids: Their physiological role and receptors. Glob. J. Pharm. 2009, 3, 149–153. [Google Scholar]
- Law, P.-Y.; Wong, Y.H.; Loh, H.H. Mutational analysis of the structure and function of opioid receptors. Pept. Sci. 1999, 51, 440–455. [Google Scholar] [CrossRef]
- Yang, Y.-R.; Chiu, T.-H.; Chen, C.-L. Structure–activity relationships of naturally occurring and synthetic opioid tetrapeptides acting on locus coeruleus neurons. Eur. J. Pharmacol. 1999, 372, 229–236. [Google Scholar] [CrossRef]
- Kaur, J.; Kumar, V.; Sharma, K.; Kaur, S.; Gat, Y.; Goyal, A.; Tanwar, B. Opioid Peptides: An Overview of Functional Significance. Int. J. Pept. Res. Ther. 2020, 26, 33–41. [Google Scholar] [CrossRef]
- Froehlich, J.C. Opioid peptides. Alcohol Health Res. World 1997, 21, 132. [Google Scholar] [PubMed]
- Garg, S.; Nurgali, K.; Kumar Mishra, V. Food proteins as source of opioid peptides-a review. Curr. Med. Chem. 2016, 23, 893–910. [Google Scholar] [CrossRef] [PubMed]
- Hackler, L.; Zadina, J.E.; Ge, L.-J.; Kastin, A.J. Isolation of relatively large amounts of endomorphin-1 and endomorphin-2 from human brain cortex. Peptides 1997, 18, 1635–1639. [Google Scholar] [CrossRef]
- Aldrich, J.V.; Kulkarni, S.S.; Senadheera, S.N.; Ross, N.C.; Reilley, K.J.; Eans, S.O.; Ganno, M.L.; Murray, T.F.; McLaughlin, J.P. Unexpected Opioid Activity Profiles of Analogues of the Novel Peptide Kappa Opioid Receptor Ligand CJ-15,208. ChemMedChem 2011, 6, 1739–1745. [Google Scholar] [CrossRef]
- Goldstein, A. Opioid peptides (endorphins) in pituitary and brain. Science 1976, 193, 1081–1086. [Google Scholar] [CrossRef]
- Li, C.H.; Chung, D. Isolation and structure of an untriakontapeptide with opiate activity from camel pituitary glands. Proc. Natl. Acad. Sci. USA 1976, 73, 1145–1148. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, A.; Fischli, W.; Lowney, L.I.; Hunkapiller, M.; Hood, L. Porcine pituitary dynorphin: Complete amino acid sequence of the biologically active heptadecapeptide. Proc. Natl. Acad. Sci. USA 1981, 78, 7219–7223. [Google Scholar] [CrossRef] [Green Version]
- Chavkin, C.; James, I.F.; Goldstein, A. Dynorphin is a specific endogenous ligand of the kappa opioid receptor. Science 1982, 215, 413–415. [Google Scholar] [CrossRef]
- Okada, Y.; Fukumizu, A.; Takahashi, M.; Shimizu, Y.; Tsuda, Y.; Yokoi, T.; Bryant, S.D.; Lazarus, L.H. Synthesis of stereoisomeric analogues of endomorphin-2, H-Tyr-Pro-Phe-Phe-NH2, and examination of their opioid receptor binding activities and solution conformation. Biochem. Biophys. Res. Commun. 2000, 276, 7–11. [Google Scholar] [CrossRef]
- Li, T.; Fujita, Y.; Tsuda, Y.; Miyazaki, A.; Ambo, A.; Sasaki, Y.; Jinsmaa, Y.; Bryant, S.D.; Lazarus, L.H.; Okada, Y. Development of Potent μ-Opioid Receptor Ligands Using Unique Tyrosine Analogues of Endomorphin-2. J. Med. Chem. 2005, 48, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Torino, D.; Mollica, A.; Pinnen, F.; Lucente, G.; Feliciani, F.; Davis, P.; Lai, J.; Ma, S.-W.; Porreca, F.; Hruby, V.J. Synthesis and evaluation of new endomorphin analogues modified at the Pro2 residue. Bioorgan. Med. Chem. Lett. 2009, 19, 4115–4118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.; Murray, T.F.; Aldrich, J.V. Synthesis and evaluation of potential affinity labels derived from endomorphin-2. J. Pept. Res. 2003, 61, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Tömböly, C.; Kövér, K.E.; Péter, A.; Tourwé, D.; Biyashev, D.; Benyhe, S.; Borsodi, A.; Al-Khrasani, M.; Rónai, A.Z.; Tóth, G. Structure−Activity Study on the Phe Side Chain Arrangement of Endomorphins Using Conformationally Constrained Analogues. J. Med. Chem. 2004, 47, 735–743. [Google Scholar] [CrossRef]
- Honda, T.; Shirasu, N.; Isozaki, K.; Kawano, M.; Shigehiro, D.; Chuman, Y.; Fujita, T.; Nose, T.; Shimohigashi, Y. Differential receptor binding characteristics of consecutive phenylalanines in μ-opioid specific peptide ligand endomorphin-2. Bioorgan. Med. Chem. 2007, 15, 3883–3888. [Google Scholar] [CrossRef]
- Sasaki, Y.; Sasaki, A.; Niizuma, H.; Goto, H.; Ambo, A. Endomorphin 2 analogues containing Dmp residue as an aromatic amino acid surrogate with high μ-opioid receptor affinity and selectivity. Bioorgan. Med. Chem. 2003, 11, 675–678. [Google Scholar] [CrossRef]
- Vázquez, M.E.; Blanco, J.B.; Salvadori, S.; Trapella, C.; Argazzi, R.; Bryant, S.D.; Jinsmaa, Y.; Lazarus, L.H.; Negri, L.; Giannini, E. 6-N, N-Dimethylamino-2, 3-naphthalimide: A new environment-sensitive fluorescent probe in δ-and μ-selective opioid peptides. J. Med. Chem. 2006, 49, 3653–3658. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Shao, X.; Cui, Y.; Liu, H.M.; Wang, C.L.; Fan, Y.Z.; Liu, J.; Dong, S.L.; Cui, Y.X.; Wang, R. Structure–activity study on the spatial arrangement of the third aromatic ring of endomorphins 1 and 2 using an atypical constrained C terminus. ChemMedChem 2007, 2, 309–317. [Google Scholar] [CrossRef]
- Liu, W.X.; Wang, R. Endomorphins: Potential roles and therapeutic indications in the development of opioid peptide analgesic drugs. Med. Res. Rev. 2012, 32, 536–580. [Google Scholar] [CrossRef]
- Cardillo, G.; Gentilucci, L.; Tolomelli, A. Unusual amino acids: Synthesis and introduction into naturally occurring peptides and biologically active analogues. Mini Rev. Med. Chem. 2006, 6, 293–304. [Google Scholar] [CrossRef]
- Gentilucci, L. SNew Trends in the Development of Opioid Peptide Analogues as Advanced Remedies for Pain Relief. Curr. Top. Med. Chem. 2004, 4, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Udenigwe, C.C. Role of food-derived opioid peptides in the central nervous and gastrointestinal systems. J. Food Biochem. 2019, 43, e12629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastin, A. Handbook of Biologically Active Peptides; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Yoshikawa, M.; Takahashi, M.; Yang, S. Delta opioid peptides derived from plant proteins. Curr. Pharm. Des. 2003, 9, 1325–1330. [Google Scholar] [CrossRef] [PubMed]
- Zioudrou, C.; Streaty, R.A.; Klee, W.A. Opioid peptides derived from food proteins. The exorphins. J. Biol. Chem. 1979, 254, 2446–2449. [Google Scholar]
- Bell, S.J.; Grochoski, G.T.; Clarke, A.J. Health implications of milk containing β-casein with the A2 genetic variant. Crit. Rev. Food Sci. Nutr. 2006, 46, 93–100. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Johnson, S.K.; Busetti, F.; Solah, V.A. Formation and degradation of beta-casomorphins in dairy processing. Crit. Rev. Food Sci. Nutr. 2015, 55, 1955–1967. [Google Scholar] [CrossRef]
- El-Salam, M.A.; El-Shibiny, S. Bioactive peptides of buffalo, camel, goat, sheep, mare, and yak milks and milk products. Food Rev. Int. 2013, 29, 1–23. [Google Scholar] [CrossRef]
- Brantl, V.; Teschemacher, H. A material with opioid activity in bovine milk and milk products. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1979, 306, 301–304. [Google Scholar] [CrossRef]
- Hazum, E.; Sabatka, J.J.; Chang, K.-J.; Brent, D.A.; Findlay, J.; Cuatrecasas, P. Morphine in cow and human milk: Could dietary morphine constitute a ligand for specific morphine (mu) receptors? Science 1981, 213, 1010–1012. [Google Scholar] [CrossRef]
- Henschen, A.; Brantl, V.; Teschemacher, H.; Lottspeich, F. β-Casomorphins–Novel Opioid Peptides Derived from Bovine Casein–Isolation and Structure. In Endogenous and Exogenous Opiate Agonists and Antagonists; Elsevier: Amsterdam, The Netherlands, 1980; pp. 233–236. [Google Scholar]
- Loukas, S.; Varoucha, D.; Zioudrou, C.; Streaty, R.A.; Klee, W.A. Opioid activities and structures of alpha-casein-derived exorphins. Biochemistry 1983, 22, 4567–4573. [Google Scholar] [CrossRef]
- Brantl, V.; Teschemacher, H.; Bläsig, J.; Henschen, A.; Lottspeich, F. Opioid activities of β-casomorphins. Life Sci. 1981, 28, 1903–1909. [Google Scholar] [CrossRef]
- Koch, G.; Wiedemann, K.; Teschemacher, H. Opioid activities of human β-casomorphins. Naunyn-Schmiedeberg Arch. Pharmacol. 1985, 331, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Bouglé, D.; Bouhallab, S. Dietary bioactive peptides: Human studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Calvo, C.F.; Cesselin, F.; Gelman, M.; Glowinski, J. Identification of an opioid peptide secreted by rat embryonic mixed brain cells as a promoter of macrophage migration. Eur. J. Neurosci. 2000, 12, 2676–2684. [Google Scholar] [CrossRef] [PubMed]
- Lister, J.; Fletcher, P.J.; Nobrega, J.N.; Remington, G. Behavioral effects of food-derived opioid-like peptides in rodents: Implications for schizophrenia? Pharmacol. Biochem. Behav. 2015, 134, 70–78. [Google Scholar] [CrossRef]
- Pruimboom, L.; De Punder, K. The opioid effects of gluten exorphins: Asymptomatic celiac disease. J. HealthPopul. Nutr. 2015, 33, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohinata, K.; Agui, S.; Yoshikawa, M. Soymorphins, novel μ opioid peptides derived from soy β-conglycinin β-subunit, have anxiolytic activities. Biosci. Biotechnol. Biochem. 2007, 71, 2618–2621. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Tani, F.; Yoshimura, T.; Chiba, H. Opioid peptides from milk proteins. Agric. Biol. Chem. 1986, 50, 2419–2421. [Google Scholar] [CrossRef]
- Brantl, V.; Pfeiffer, A.; Herz, A.; Henschen, A.; Lottspeich, F. Antinociceptive potencies of β-casomorphin analogs as compared to their affinities towards μ and δ opiate receptor sites in brain and periphery. Peptides 1982, 3, 793–797. [Google Scholar] [CrossRef]
- Tani, F.; Iio, K.; Chiba, H.; Yoshikawa, M. Isolation and characterization of opioid antagonist peptides derived from human lactoferrin. Agric. Biol. Chem. 1990, 54, 1803–1810. [Google Scholar]
- Sharma, S.K.; Klee, W.A.; Nirenberg, M. Dual regulation of adenylate cyclase accounts for narcotic dependence and tolerance. Proc. Natl. Acad. Sci. USA 1975, 72, 3092–3096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, J.; Kosterlitz, H.; Leslie, F.M. Effect of morphine on adrenergic transmission in the mouse vas deferens. Assessment of agonist and antogonist potencies of narcotic analgesics. Br. J. Pharmacol. 1975, 53, 371. [Google Scholar] [CrossRef] [PubMed]
- Pert, C.B.; Pasternak, G.; Snyder, S.H. Opiate agonists and antagonists discriminated by receptor binding in brain. Science 1973, 182, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Kawamura, Y.; Yoshikawa, M. Effect of rubiscolin, a δ opioid peptide derived from Rubisco, on memory consolidation. Peptides 2003, 24, 325–328. [Google Scholar] [CrossRef]
- Kaneko, K.; Lazarus, M.; Miyamoto, C.; Oishi, Y.; Nagata, N.; Yang, S.; Yoshikawa, M.; Aritake, K.; Furuyashiki, T.; Narumiya, S. Orally administered rubiscolin-6, a δ opioid peptide derived from Rubisco, stimulates food intake via leptomeningeal lipocallin-type prostaglandin D synthase in mice. Mol. Nutr. Food Res. 2012, 56, 1315–1323. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Kaneko, K.; Iguchi, S.; Mizushige, T.; Kanamoto, R.; Yoshikawa, M.; Shimizu, T.; Ohinata, K. Orally administered δ opioid agonist peptide rubiscolin-6 stimulates food intake in aged mice with ghrelin resistance. Mol. Nutr. Food Res. 2014, 58, 2046–2052. [Google Scholar] [CrossRef]
- Hirata, H.; Sonoda, S.; Agui, S.; Yoshida, M.; Ohinata, K.; Yoshikawa, M. Rubiscolin-6, a δ opioid peptide derived from spinach Rubisco, has anxiolytic effect via activating σ1 and dopamine D1 receptors. Peptides 2007, 28, 1998–2003. [Google Scholar] [CrossRef]
- Kaneko, K.; Mizushige, T.; Miyazaki, Y.; Lazarus, M.; Urade, Y.; Yoshikawa, M.; Kanamoto, R.; Ohinata, K. δ-Opioid receptor activation stimulates normal diet intake but conversely suppresses high-fat diet intake in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R265–R272. [Google Scholar] [CrossRef]
- Schusdziarra, V.; Schick, A.; de la Fuente, A.; Specht, J.; Klier, M.; Brantl, V.; Pfeiffer, E.-F. Effect of β-casomorphins and analogs on insulin release in dogs. Endocrinology 1983, 112, 885–889. [Google Scholar] [CrossRef]
- Mihatsch, W.; Franz, A.; Kuhnt, B.; Högel, J.; Pohlandt, F. Hydrolysis of casein accelerates gastrointestinal transit via reduction of opioid receptor agonists released from casein in rats. Neonatology 2005, 87, 160–163. [Google Scholar] [CrossRef]
- Zoghbi, S.; Trompette, A.; Claustre, J.; Homsi, M.E.; Garzón, J.; Jourdan, G.; Scoazec, J.-Y.; Plaisancié, P. β-Casomorphin-7 regulates the secretion and expression of gastrointestinal mucins through a μ-opioid pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G1105–G1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Miao, J.; Zhang, Y. Protective effect of β-casomorphin-7 on type 1 diabetes rats induced with streptozotocin. Peptides 2010, 31, 1725–1729. [Google Scholar] [CrossRef]
- Fanciulli, G.; Dettori, A.; Demontis, M.P.; Tomasi, P.A.; Anania, V.; Delitala, G. Gluten exorphin B5 stimulates prolactin secretion through opioid receptors located outside the blood-brain barrier. Life Sci. 2005, 76, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Belyaeva, Y.A.; Dubynin, V.; Stovolosov, I.; Dobryakova, Y.V.; Bespalova, Z.D.; Kamenskii, A. Effects of acute and chronic administration of exorphin C on behavior and learning in white rat pups. Mosc. Univ. Biol. Sci. Bull. 2009, 64, 66–70. [Google Scholar] [CrossRef]
- Jinsmaa, Y.; Yoshikawa, M. Enzymatic release of neocasomorphin and β-casomorphin from bovine β-casein. Peptides 1999, 20, 957–962. [Google Scholar] [CrossRef]
- Fukudome, S.-I.; Yoshikawa, M. Opioid peptides derived from wheat gluten: Their isolation and characterization. Febs Lett. 1992, 296, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Fukudome, S.-I.; Yoshikawa, M. Gluten exorphin C: A novel opioid peptide derived from wheat gluten. FEBS Lett. 1993, 316, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Yunden, J.; Sonoda, S.; Doyama, N.; Lipkowski, A.W.; Kawamura, Y.; Yoshikawa, M. Rubiscolin, a δ selective opioid peptide derived from plant Rubisco. FEBS Lett. 2001, 509, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Matthies, H.; Stark, H.; Hartrodt, B.; Ruethrich, H.-L.; Spieler, H.-T.; Barth, A.; Neubert, K. Derivatives of β-casomorphins with high analgesic potency. Peptides 1984, 5, 463–470. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Koseki, M.; Wakamatsu, M.; Matsumura, E. Effects of systemic administration of β-casomorphin-5 on learning and memory in mice. Eur. J. Pharmacol. 2006, 530, 81–87. [Google Scholar] [CrossRef]
- Grecksch, G.; Schweigert, C.; Matthies, H. Evidence for analgesic activity of β-casomorphin in rats. Neurosci. Lett. 1981, 27, 325–328. [Google Scholar] [CrossRef]
- Lin, L.; Umahara, M.; York, D.; Bray, G. β-Casomorphins stimulate and enterostatin inhibits the intake of dietary fat in rats. Peptides 1998, 19, 325–331. [Google Scholar] [CrossRef]
- Kaneko, K.; Iwasaki, M.; Yoshikawa, M.; Ohinata, K. Orally administered soymorphins, soy-derived opioid peptides, suppress feeding and intestinal transit via gut μ1-receptor coupled to 5-HT1A, D2, and GABAB systems. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G799–G805. [Google Scholar] [CrossRef] [Green Version]
- Chesnokova, E.; Saricheva, N.; Dubynin, V.; Kamenskij, A.; Kalikhevich, V.; Adermasova, Z. Behavioral effect of soymorphin-5-amide in rats. Mosc. Univ. Biol. Sci. Bull. 2014, 69, 103–107. [Google Scholar] [CrossRef]
- Boullin, D.; Crampton, R.; Heading, C.E.; Pelling, D. Intestinal absorption of dipeptides containing glycine, phenylalanine, proline, β-alanine or histidine in the rat. Clin. Sci. Mol. Med. 1973, 45, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D. Intestinal absorption of peptides. Physiol. Rev. 1975, 55, 537–608. [Google Scholar] [CrossRef]
- Gardner, M. Absorption of amino acids and peptides from a complex mixture in the isolated small intestine of the rat. J. Physiol. 1975, 253, 233–256. [Google Scholar] [CrossRef] [Green Version]
- Zaloga, G.P.; Siddiqui, R.A. Biologically active dietary peptides. Mini Rev. Med. Chem. 2004, 4, 815–821. [Google Scholar] [CrossRef]
- Kaspar, A.A.; Reichert, J.M. Future directions for peptide therapeutics development. Drug Discov. Today 2013, 18, 807–817. [Google Scholar] [CrossRef]
- Houston, M.E.; Campbell, A.P.; Lix, B.; Kay, C.M.; Sykes, B.D.; Hodges, R.S. Lactam bridge stabilization of α-helices: The role of hydrophobicity in controlling dimeric versus monomeric α-helices. Biochemistry 1996, 35, 10041–10050. [Google Scholar] [CrossRef]
- Timmerman, P.; Puijk, W.C.; Meloen, R.H. Functional reconstruction and synthetic mimicry of a conformational epitope using CLIPS™ technology. J. Mol. Recognit. Interdiscip. J. 2007, 20, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Kim, Y.; Kim, T.; Lim, S.; Lee, M. Directional assembly of α-helical peptides induced by cyclization. J. Am. Chem. Soc. 2012, 134, 20270–20272. [Google Scholar] [CrossRef]
- Bougatef, A.; Nedjar-Arroume, N.; Ravallec-Plé, R.; Leroy, Y.; Guillochon, D.; Barkia, A.; Nasri, M. Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained by treatment with microbial and visceral fish serine proteases. Food Chem. 2008, 111, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Y.; Zhong, Q.; Wu, Y.; Xia, W. Purification and characterization of a novel angiotensin-I converting enzyme (ACE) inhibitory peptide derived from enzymatic hydrolysate of grass carp protein. Peptides 2012, 33, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, M.P.C.; Silva, M.R.; Silva, V.D.M.; Souza, M.W.S.d.; Junior, L.; de Oliveira, C.; Afonso, W.d.O. Analysis of whey protein hydrolysates: Peptide profile and ACE inhibitory activity. Braz. J. Pharm. Sci. 2012, 48, 747–757. [Google Scholar] [CrossRef] [Green Version]
- Darewicz, M.; Borawska, J.; Vegarud, G.E.; Minkiewicz, P.; Iwaniak, A. Angiotensin I-converting enzyme (ACE) inhibitory activity and ACE inhibitory peptides of salmon (Salmo salar) protein hydrolysates obtained by human and porcine gastrointestinal enzymes. Int. J. Mol. Sci. 2014, 15, 14077–14101. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Naramoto, K.; Koyama, M. Blood-pressure-lowering effect of fermented buckwheat sprouts in spontaneously hypertensive rats. J. Funct. Foods 2013, 5, 406–415. [Google Scholar] [CrossRef]
- García-Tejedor, A.; Saánchez-Rivera, L.; Castelló-Ruiz, M.; Recio, I.; Salom, J.B.; Manzanares, P. Novel antihypertensive lactoferrin-derived peptides produced by Kluyveromyces marxianus: Gastrointestinal stability profile and in vivo angiotensin I-converting enzyme (ACE) inhibition. J. Agric. Food Chem. 2014, 62, 1609–1616. [Google Scholar] [CrossRef]
- Ha, G.E.; Chang, O.K.; Jo, S.-M.; Han, G.-S.; Park, B.-Y.; Ham, J.-S.; Jeong, S.-G. Identification of antihypertensive peptides derived from low molecular weight casein hydrolysates generated during fermentation by Bifidobacterium longum KACC 91563. Korean J. Food Sci. Anim. Resour. 2015, 35, 738. [Google Scholar] [CrossRef]
- Jarmołowska, B.; Kostyra, E.; Krawczuk, S.; Kostyra, H. β-Casomorphin-7 isolated from Brie cheese. J. Sci. Food Agric. 1999, 79, 1788–1792. [Google Scholar] [CrossRef]
- Stuknytė, M.; Maggioni, M.; Cattaneo, S.; De Luca, P.; Fiorilli, A.; Ferraretto, A.; De Noni, I. Release of wheat gluten exorphins A5 and C5 during in vitro gastrointestinal digestion of bread and pasta and their absorption through an in vitro model of intestinal epithelium. Food Res. Int. 2015, 72, 208–214. [Google Scholar] [CrossRef]
- De Noni, I.; Cattaneo, S. Occurrence of β-casomorphins 5 and 7 in commercial dairy products and in their digests following in vitro simulated gastro-intestinal digestion. Food Chem. 2010, 119, 560–566. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected lactic acid bacteria synthesize antioxidant peptides during sourdough fermentation of cereal flours. Appl. Environ. Microbiol. 2012, 78, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Zou, Y.; Yu, N.; Gu, Z. Accumulation and identification of angiotensin-converting enzyme inhibitory peptides from wheat germ. J. Agric. Food Chem. 2011, 59, 3598–3605. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.S.; Baron, C.P.; Nielsen, N.S.; Otte, J.; Jacobsen, C. Antioxidant activity of yoghurt peptides: Part 2–characterisation of peptide fractions. Food Chem. 2010, 123, 1090–1097. [Google Scholar] [CrossRef]
- Pripp, A.H.; Isaksson, T.; Stepaniak, L.; Sørhaug, T. Quantitative structure-activity relationship modelling of ACE-inhibitory peptides derived from milk proteins. Eur. Food Res. Technol. 2004, 219, 579–583. [Google Scholar] [CrossRef]
- Wu, C.-H.; Kuo, Y.-H.; Hong, R.-L.; Wu, H.-C. α-Enolase–binding peptide enhances drug delivery efficiency and therapeutic efficacy against colorectal cancer. Sci. Transl. Med. 2015, 7, 290ra291. [Google Scholar] [CrossRef]
- Wu, C.-H.; Liu, I.-J.; Lu, R.-M.; Wu, H.-C. Advancement and applications of peptide phage display technology in biomedical science. J. Biomed. Sci. 2016, 23, 8. [Google Scholar] [CrossRef] [Green Version]
- Cavazos, A.; Gonzalez de Mejia, E. Identification of bioactive peptides from cereal storage proteins and their potential role in prevention of chronic diseases. Compr. Rev. Food Sci. Food Saf. 2013, 12, 364–380. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Gong, M.; Wu, S. In silico analysis of the large and small subunits of cereal RuBisCO as precursors of cryptic bioactive peptides. Process Biochem. 2013, 48, 1794–1799. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Holton, T.A.; Pollastri, G.; Shields, D.C.; Mooney, C. CPPpred: Prediction of cell penetrating peptides. Bioinformatics 2013, 29, 3094–3096. [Google Scholar] [CrossRef] [PubMed]
- Chandrudu, S.; Simerska, P.; Toth, I. Chemical methods for peptide and protein production. Molecules 2013, 18, 4373–4388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiuchi, Y.; Inui, T.; Nishio, H.; BOdi, J.; Kimura, T.; Tsuji, F.I.; Sakakibara, S. Chemical synthesis of the precursor molecule of the Aequorea green fluorescent protein, subsequent folding, and development of fluorescence. Proc. Natl. Acad. Sci. USA 1998, 95, 13549–13554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpino, L.A.; Ghassemi, S.; Ionescu, D.; Ismail, M.; Sadat-Aalaee, D.; Truran, G.A.; Mansour, E.; Siwruk, G.A.; Eynon, J.S.; Morgan, B. Rapid, continuous solution-phase peptide synthesis: Application to peptides of pharmaceutical interest. Org. Process Res. Dev. 2003, 7, 28–37. [Google Scholar] [CrossRef]
- Murata, H.; Carmali, S.; Baker, S.L.; Matyjaszewski, K.; Russell, A.J. Solid-phase synthesis of protein-polymers on reversible immobilization supports. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Dirksen, A.; Meijer, E.; Adriaens, W.; Hackeng, T.M. Strategy for the synthesis of multivalent peptide-based nonsymmetric dendrimers by native chemical ligation. Chem. Commun. 2006, 1667–1669. [Google Scholar] [CrossRef]
- Lovrinovic, M.; Niemeyer, C.M. Microtiter plate-based screening for the optimization of DNA–protein conjugate synthesis by means of expressed protein ligation. ChemBioChem 2007, 8, 61–67. [Google Scholar] [CrossRef]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef]
- Meisel, H.; FitzGerald, R.J. Biofunctional peptides from milk proteins: Mineral binding and cytomodulatory effects. Curr. Pharm. Des. 2003, 9, 1289–1296. [Google Scholar]
- Mendis, E.; Rajapakse, N.; Byun, H.-G.; Kim, S.-K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- González-García, E.; Marina, M.L.; García, M.C. Plum (Prunus domestica L.) by-product as a new and cheap source of bioactive peptides: Extraction method and peptides characterization. J. Funct. Foods 2014, 11, 428–437. [Google Scholar] [CrossRef]
- Agyei, D.; Danquah, M.K. Industrial-scale manufacturing of pharmaceutical-grade bioactive peptides. Biotechnol. Adv. 2011, 29, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, S.-K. Bioactive peptides from marine sources as potential anti-inflammatory therapeutics. Curr. Protein Pept. Sci. 2013, 14, 177–182. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.; Park, S.; Atwal, A.; Gibbs, B.; Lee, B. Angiotensin I-converting enzyme (ACE) inhibitory peptides from whey fermented by Lactobacillus species. J. Food Biochem. 2009, 33, 587–602. [Google Scholar] [CrossRef]
- Hou, P.; Zhao, G.; He, C.; Wang, H.; He, H. Biopanning of polypeptides binding to bovine ephemeral fever virus G 1 protein from phage display peptide library. BMC Vet. Res. 2018, 14, 3. [Google Scholar] [CrossRef]
- Suire, C.N.; Nainar, S.; Fazio, M.; Kreutzer, A.G.; Paymozd-Yazdi, T.; Topper, C.L.; Thompson, C.R.; Leissring, M.A. Peptidic inhibitors of insulin-degrading enzyme with potential for dermatological applications discovered via phage display. PLoS ONE 2018, 13, e0193101. [Google Scholar] [CrossRef]
- Khondee, S.; Piyawattanametha, W. Targeting Peptides Derived from Phage Display for Clinical Imaging. In Bacteriophages-Biology and Applications; IntechOpen: London, UK, 2019. [Google Scholar]
- Shen, Y.; Ruan, L.; Lian, C.; Li, R.; Tu, Z.; Liu, H. Discovery of HB-EGF binding peptides and their functional characterization in ovarian cancer cell lines. Cell Death Discov. 2019, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, A.; Kringelum, J.V.; Hansen, C.S.; Bøgh, K.L.; Sullivan, E.; Patel, J.; Rigby, N.M.; Eiwegger, T.; Szépfalusi, Z.; De Masi, F. High-throughput sequencing enhanced phage display enables the identification of patient-specific epitope motifs in serum. Sci. Rep. 2015, 5, 12913. [Google Scholar] [CrossRef] [Green Version]
- Ramaraju, H.; Miller, S.J.; Kohn, D.H. Dual-functioning peptides discovered by phage display increase the magnitude and specificity of BMSC attachment to mineralized biomaterials. Biomaterials 2017, 134, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, F.; Luo, J.; Pang, J.; Yan, S.; Luo, F.; Liu, J.; Wang, W.; Cui, Y.; Su, X. A new non-muscle-invasive bladder tumor-homing peptide identified by phage display in vivo. Oncol. Rep. 2016, 36, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricker, L.D. Neuropeptides and other bioactive peptides: From discovery to function. In Colloquium Series on Neuropeptides; Morgan & Claypool Publishers: Williston, VT, USA, 2012. [Google Scholar]
- Law, P.-Y.; Wong, Y.H.; Loh, H.H. Molecular mechanisms and regulation of opioid receptor signaling. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 389–430. [Google Scholar] [CrossRef] [PubMed]
- De Noni, I.; FitzGerald, R.J.; Korhonen, H.J.; Le Roux, Y.; Livesey, C.T.; Thorsdottir, I.; Tomé, D.; Witkamp, R. Review of the potential health impact of β-casomorphins and related peptides. EFSA Sci. Rep. 2009, 231, 1–107. [Google Scholar]
- Ganapathy, V.; Miyauchi, S. Transport systems for opioid peptides in mammalian tissues. AAPS J. 2005, 7, E852–E856. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.-J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2014; pp. 39–71. [Google Scholar]
- Brasnjevic, I.; Steinbusch, H.W.; Schmitz, C.; Martinez-Martinez, P.; Initiative, E.N.R. Delivery of peptide and protein drugs over the blood–brain barrier. Prog. Neurobiol. 2009, 87, 212–251. [Google Scholar] [CrossRef]
- Wang, J.; Hogenkamp, D.J.; Tran, M.; Li, W.-Y.; Yoshimura, R.F.; Johnstone, T.B.; Shen, W.-C.; Gee, K.W. Reversible lipidization for the oral delivery of leu-enkephalin. J. Drug Target. 2006, 14, 127–136. [Google Scholar] [CrossRef]
- Brugos, B.; Hochhaus, G. Metabolism of dynorphin A (1–13). Die Pharm.-Int. J. Pharm. Sci. 2004, 59, 339–343. [Google Scholar]
- Negri, L.; Lattanzi, R.; Tabacco, F.; Scolaro, B.; Rocchi, R. Glycodermorphins: Opioid peptides with potent and prolonged analgesic activity and enhanced blood-brain barrier penetration. Br. J. Pharmacol. 1998, 124, 1516–1522. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-J.; Zhou, J.; Martin, B.; Carlson, O.D.; Maudsley, S.; Greig, N.H.; Mattson, M.P.; Ladenheim, E.E.; Wustner, J.; Turner, A. Transferrin fusion technology: A novel approach to prolonging biological half-life of insulinotropic peptides. J. Pharmacol. Exp. Ther. 2010, 334, 682–692. [Google Scholar] [CrossRef] [Green Version]
- Dennis, M.S.; Zhang, M.; Meng, Y.G.; Kadkhodayan, M.; Kirchhofer, D.; Combs, D.; Damico, L.A. Albumin binding as a general strategy for improving the pharmacokinetics of proteins. J. Biol. Chem. 2002, 277, 35035–35043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egleton, R.D.; Mitchell, S.A.; Huber, J.D.; Janders, J.; Stropova, D.; Polt, R.; Yamamura, H.I.; Hruby, V.J.; Davis, T.P. Improved bioavailability to the brain of glycosylated Met-enkephalin analogs. Brain Res. 2000, 881, 37–46. [Google Scholar] [CrossRef]
- Perlikowska, R.; Gach, K.; Fichna, J.; Toth, G.; Walkowiak, B.; do-Rego, J.-C.; Janecka, A. Biological activity of endomorphin and [Dmt1]endomorphin analogs with six-membered proline surrogates in position 2. Bioorganic Med. Chem. 2009, 17, 3789–3794. [Google Scholar] [CrossRef] [PubMed]
- Tóth, G.; Keresztes, A.; Tömböly, C.; Péter, A.; Fülöp, F.; Tourwé, D.; Navratilova, E.; Varga, É.; Roeske, W.R.; Yamamura, H.I.; et al. New endomorphin analogs with mu-agonist and delta-antagonist properties. Pure Appl. Chem. 2004, 76, 951–957. [Google Scholar] [CrossRef] [Green Version]
- Bali, A.; Singh, N.; Singh Jaggi, A. Neuropeptides as therapeutic targets to combat stress-associated behavioral and neuroendocrinological effects. CNS Neurol. Disord. Drug Targets (Former. Curr. Drug Targets-Cns Neurol. Disord.) 2014, 13, 347–368. [Google Scholar] [CrossRef]
- Clynen, E.; Swijsen, A.; Raijmakers, M.; Hoogland, G.; Rigo, J.-M. Neuropeptides as targets for the development of anticonvulsant drugs. Mol. Neurobiol. 2014, 50, 626–646. [Google Scholar] [CrossRef] [Green Version]
- Yimit, D.; Hoxur, P.; Amat, N.; Uchikawa, K.; Yamaguchi, N. Effects of soybean peptide on immune function, brain function, and neurochemistry in healthy volunteers. Nutrition 2012, 28, 154–159. [Google Scholar] [CrossRef]
- Bernet, F.; Montel, V.; Noël, B.; Dupouy, J.P. Diazepam-like effects of a fish protein hydrolysate (Gabolysat PC60) on stress responsiveness of the rat pituitary-adrenal system and sympathoadrenal activity. Psychopharmacology 2000, 149, 34–40. [Google Scholar] [CrossRef]
- Perlikowska, R.; Janecka, A. Rubiscolins-highly potent peptides derived from plant proteins. Mini Rev. Med. Chem. 2018, 18, 104–112. [Google Scholar] [CrossRef]
- Jahan-Mihan, A.; Luhovyy, B.L.; El Khoury, D.; Anderson, G.H. Dietary proteins as determinants of metabolic and physiologic functions of the gastrointestinal tract. Nutrients 2011, 3, 574–603. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, M. Bioactive peptides derived from natural proteins with respect to diversity of their receptors and physiological effects. Peptides 2015, 72, 208–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booij, L.; Merens, W.; Markus, C.R.; Van der Does, A.W. Diet rich in α-lactalbumin improves memory in unmedicated recovered depressed patients and matched controls. J. Psychopharmacol. 2006, 20, 526–535. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The number of publications on stress-related peptides by year available on PubMed Central as of 30 September 2020.
Figure 1.
The number of publications on stress-related peptides by year available on PubMed Central as of 30 September 2020.

Figure 2.
In silico approach to identify peptides from unknown sources of protein. QSAR: quantitative structure–activity relationship, Str: Structure.
Figure 2.
In silico approach to identify peptides from unknown sources of protein. QSAR: quantitative structure–activity relationship, Str: Structure.

Figure 3.
Representation of integrated approach (conventional/classical and bioinformatics) for the identification of food-derived bioactive peptides.
Figure 3.
Representation of integrated approach (conventional/classical and bioinformatics) for the identification of food-derived bioactive peptides.

Figure 4.
An overview of neuropeptide biosynthesis.

Figure 5.
Mechanism of action.


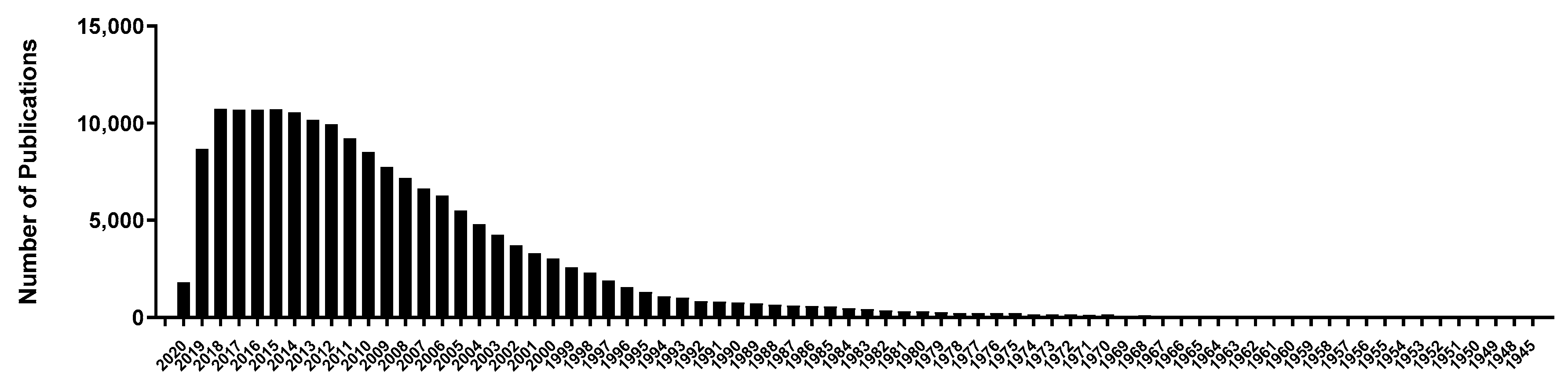{kind=link}
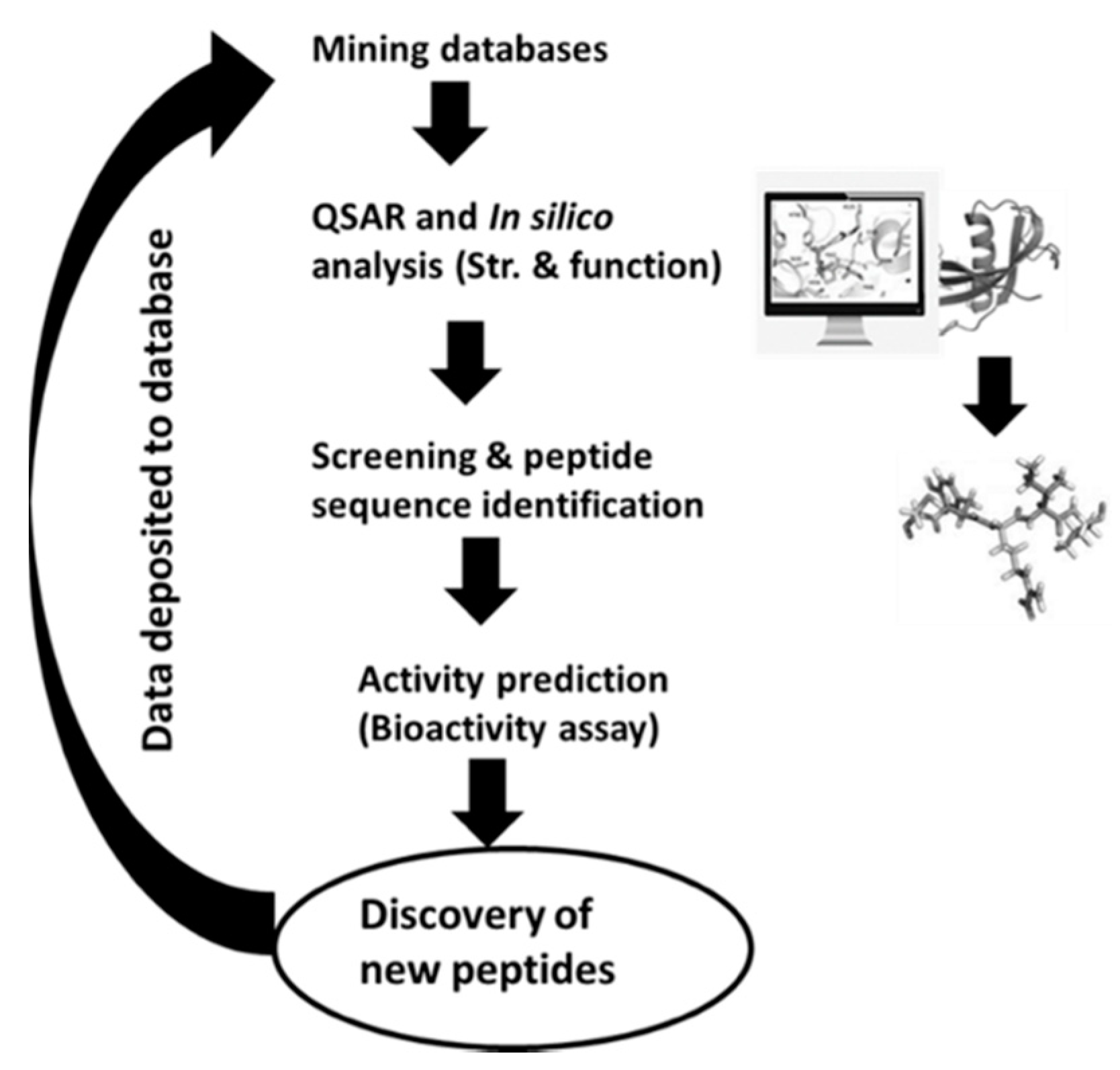{kind=link}
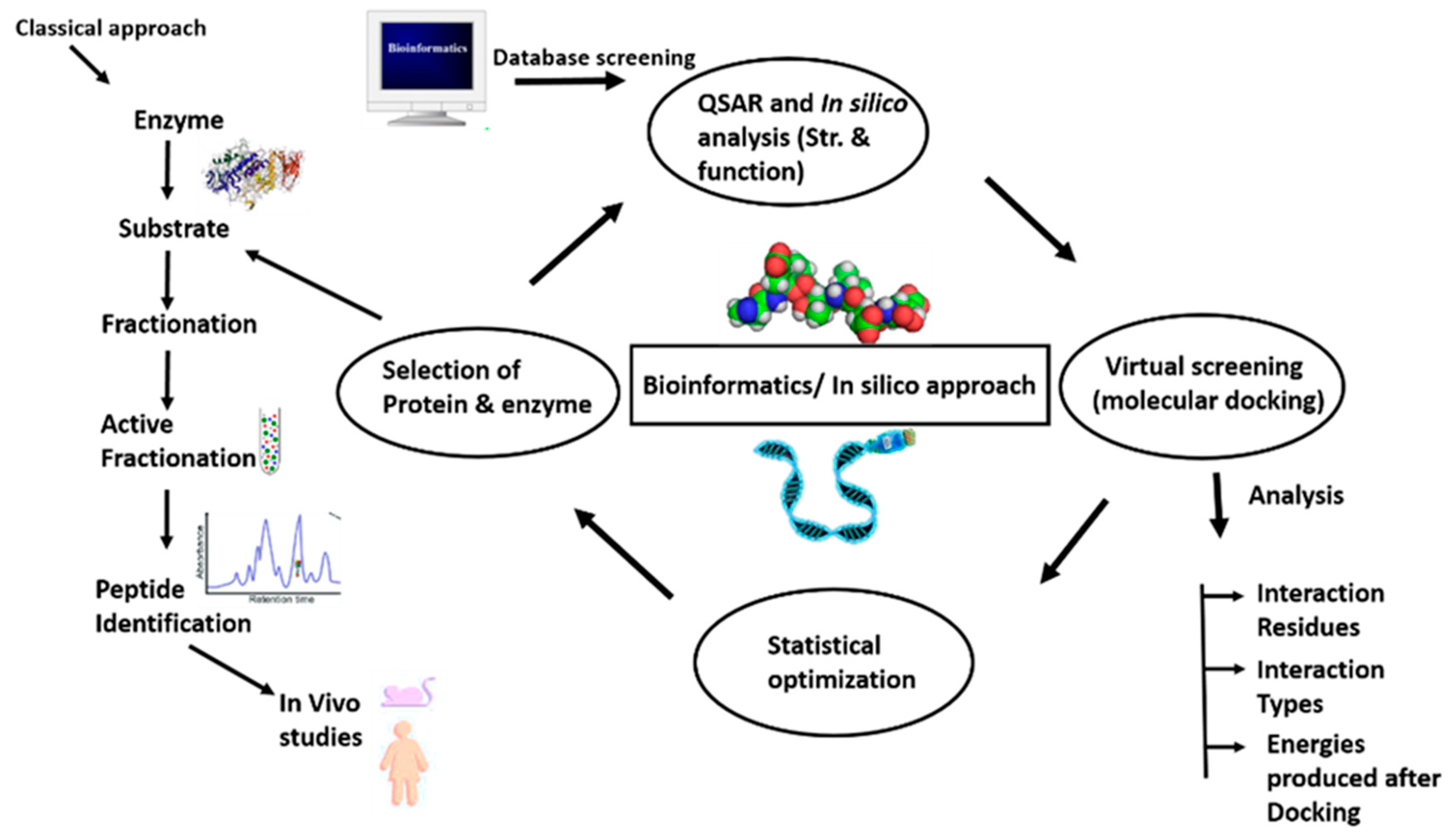{kind=link}
{kind=link}
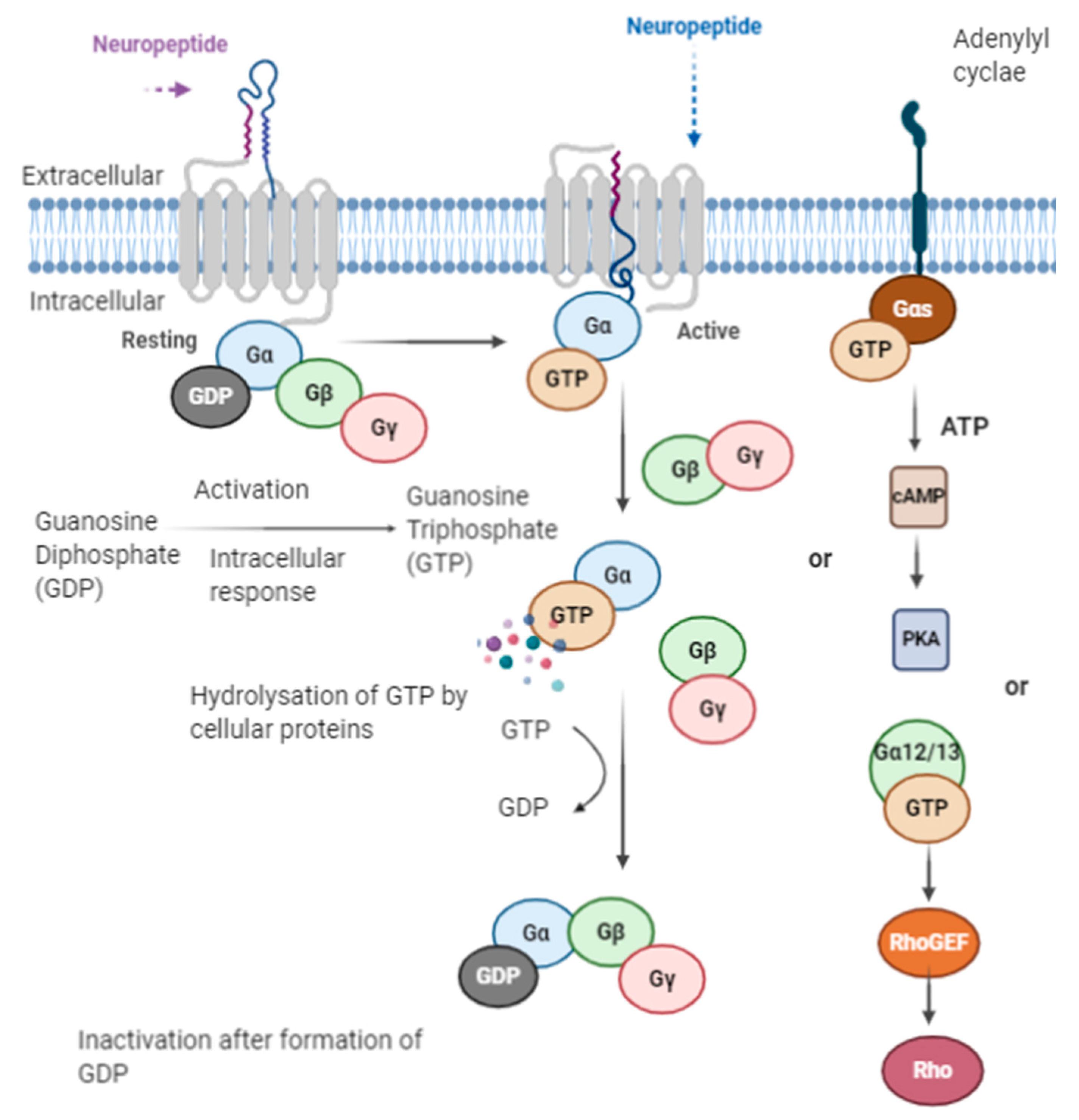{kind=link}
Table 1.
Opioid-like peptides and their roles.
| Peptides | Functions | References |
|---|---|---|
| Colostrinin | Promotes acquisition of spatial learning in aged rats | [28] |
| Corticotropin-releasing factor (CRF) | Improves memory retention Enhance learning performance | [17,29] |
| Neuropeptide Y (NPY) | Neuroprotection as by control of feeding works against neurodegenerative diseases | [17,30] |
| Substance P (SP) | Improves functional recovery and increases the learning ability | [17,31] |
| Nociceptin/orphanin FQ (N/OFQ) | Impairs spatial learning in animal models. Facilitate memory | [32,33,34] |
| Angiotensin-vasopressin (AVP) and Oxytocin (OT) | Promote social memory and learning behaviors. Deficiency of AVP results in memory impairment | [35,36] |
| Cholecystokinin (CCK) | CCK peptides improve learning and memory performance in the patients Help in anxiety states Lack of CCK-A receptors cause impaired learning and memory functions. Play a role in conditioned fear stress and anxiety | [29,37,38] |
| Atrial natriuretic peptide (ANP), Brain-derived natriuretic peptide (BNP), C-type natriuretic peptide (CNP) | Promote action on memory consolidation | [39,40] |
| Pituitary adenylate cyclase-activating polypeptide (PACAP) | Promote learning (consolidation and retrieval) | [41] |
| Galanin | Impairs the learning and memory performances overexpression impairs cognition | [42,43,44] |
| Bombesin/gastrin-releasing peptide (BN/GRP) and Neuromedin (NM) | Improve memory performance | [45,46] |
| Hippocampal cholinergic neurostimulation Peptide (HCNP) | Abnormal accumulation and expression associated with memory and learning disorders | [47] |
| Calcitonin-gene related peptide (CGRP), Substance P(SP) and Neuropeptide Y (NPY) | Enhance memory retention. SP improves functional recovery and increases learning ability. NPY enhances memory | [30] |
| Insulin | Improves short-term memory | [48] |
| Orexin-A | Inhibits long-term potentiation (LTP) and retards spatial learning | [49,50] |
Table 2.
(A) Endogenous opioid peptides. (B) Binding affinities of various Endomorphin analogues.
| (A) | ||||
| Opioid Peptide | Amino-Acid Sequence | Protein Precursor | References | |
| endomorphin-1 | Tyr-Pro-Trp-Phe-NH2 | pro-endomorphin | [57,58] | |
| endomorphin-2 | Tyr-Pro-Phe-Phe-NH2 | pro-endomorphin | [57,58] | |
| met-enkephalin | Tyr-Gly-Gly-Phe-Met | pro-enkephalin | [16], | |
| leu-enkephalin | Tyr-Gly-Gly-Phe-Leu | pro-enkephalin | [16,59] | |
| β-endorphin | Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-ThrPro-Leu-Val-Thr-Leu-Phe-Lys-Asn-Ala-Ile-Ile-LysAsn-Ala-Tyr-Lys-Lys-Gly-Glu | pro-opiomelanocortin | [60,61] | |
| dynorphin A | Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Ile-Arg-Pro-Lys-LeuLys-Trp-Asp-Asn-Gln | pro-dynorphin | [62,63] | |
| dynorphin B | Phe-Gly-Gly-Phe-Thr-Gly-Ala-Arg-Lys-Ser-Ala-ArgLys-Leu-Ala-Asn-Gln | pronociceptin | [62,63] | |
(B) | ||||
| Sequence | IC50 (nM) | Ratio of IC50 Ratio δ/µ | References | |
| Affity (µ-Receptor) | Affity (δ-Receptor) | |||
| Endomorphins Modified at First Amino Acid Position | ||||
| d-Tyr1-Pro-Phe-Phe-NH2 | 32.1 ± 1.5 | 4121 ± 1492 | 128 | [64] |
| Dmt1-Pro-Trp-Phe-NH2 | 0.014 ± 0.003 | 12.0 ± 4.05 | 857 | [65] |
| Mmt1-Pro-Phe-Phe-NH2 | 0.132 ± 0.008 | 528.6 ± 47 | 4005 | [65] |
| Emt1-Pro-Phe-Phe NH2 | 0.063 ± 0.006 | 55.7 ± 6.2 | 884 | [65] |
| Dit1-Pro-Phe-Phe-NH2 | 2.29 ± 0.37 | 105 ± 16 | 46 | [65] |
| Det1-Pro-Phe-Phe-NH2 | 0.084 ± 0.006 | 69.7 ± 5.3 | 830 | [65] |
| Tmt1-Pro-Phe-Phe-NH2 | 1.111 ± 0.002 | 593.5 ± 80 | 5347 | [65] |
| Endomorphins Modified at Second Amino Acid Position | ||||
| Tyr-d-Pro2-Phe-Phe-NH2 | 512.4 ± 29 | 30,641 ± 419 | 60 | [64] |
| Tyr-Aze2-Trp-Phe-NH2 | 2.3 ± 0.23 | 3500 ± 360 | 1500 | [66] |
| Tyr-δAla2-Phe-Phe-NH2 | 34 ± 6.3 | 710 ± 130 | 21 | [66] |
| Tyr-3Aze2-Phe-Phe-NH2 | 210 ± 51 | 6900 ± 1200 | 32 | [66] |
| Tyr-Aze2-Phe-Phe-NH2 | 5.6 ± 1.2 | 5100 ± 600 | 920 | [66] |
| Endomorphins Modified at Third Amino Acid Position | ||||
| Tyr-Pro-d-Phe3-Phe-NH2 | 203.2 ± 83 | 4230 ± 344 | 21 | [64] |
| Tyr-Pro-Phe3-(p-NH2)-Phe-NH2 | 185 ± 36 | >10,000 | >1.9 | [67] |
| TyrProPhe3(p-NHCOCH2Br)-PheNH2 | 7210 ± 820 | >10,000 | >1.4 | [67] |
| Tyr-Pro-(2S,3R)-βMePhe3-Phe-NH2 | 106 ± 9 | >10,000 | >10 | [68] |
| Tyr-Pro-(2S,3S)-βMePhe3-Phe-NH2 | 45.3 ± 4.1 | 179 ± 15 | 4 | [68] |
| Tyr-Pro-(2R,3S)-βMePhe3-Phe-NH2 | 4910 ± 328 | >10,000 | >2 | [68] |
| Tyr-Pro-(2R,3R)-βMePhe3-Phe-NH2 | 7090 ± 131 | 6760 ± 865 | 1 | [68] |
| Tyr-Pro-(F5)-Phe3-Phe-NH2 | 11.7 ± 0.503 | 11,700 ± 1010 | 1000 | [69] |
| Endomorphins Modified at C-TERMINAL Position | ||||
| Tyr-Pro-Phe-d-Phe4-NH2 | 45.9 ± 8.6 | 8159 ± 1569 | 177 | [64] |
| Tyr-Pro-Phe-(p-NH2)-Phe4-NH2 | 36.7 ± 2.2 | >10,000 | >270 | [67] |
| TyrProPhePhe4(p-NHCOCH2Br)-NH2 | 158 ± 23 | 1940 ± 310 | 12 | [67] |
| Tyr-Pro-Phe-(p-NCS)-Phe4-NH2 | 345 ± 128 | >10,000 | >29 | [67] |
| Tyr-Pro-Trp-Dmp4-NH2 | 13.2 ± 1.9 | 7624 ± 2571 | 578 | [70] |
| Tyr-Pro-Trp-d-Dmp4-NH2 | 106 ± 20 | 1765 ± 834 | 17 | [70] |
| Tyr-Pro-Phe-Phe4-NH-(CH2)5-CODap(6DMN)-NH2 | 244.5 ± 14 | 5939 ± 1396 | 24 | [71] |
| Tyr-Pro-Phe-d-Val4-NH-Bn | 4.97 ± 1.24 | 3358 ± 414 | 676 | [72] |
| Tyr-Pro-Trp-d-Val4-NH-Bn | 2.32 ± 0.15 | 3287 ± 456 | 1417 | [72] |
2′,6′-dimethyltyrosine (Dmt), 2′-monomethyltyrosine (Mmt), 2′,3′,6′-trimethyltyrosine (Tmt), 2′-ethyl-6′-methyltyrosine (Emt), 2′,6′-diethyltyrosine (Det), 2′6′-dimethylphenylalanine (Dmp), 6-N,N-(dimethylamino)-2,3-naphthalimide (6DMN) and 2′,6′-diisopropyltyrosine (Dit).
Table 3.
Exogenous food-derived opioid peptides.
| Source | Sequences | Peptide Name | References |
|---|---|---|---|
| Bovine milk β-casein | Tyr-Pro-Phe-Pro | βb-casomorphin-4 | [87] |
| Tyr-Pro-Phe-Pro-Gly | βb-casomorphin-5 | ||
| Tyr-Pro-Phe-Pro-Gly-Pro | βb-casomorphin-6 | ||
| Tyr-Pro-Phe-Pro-Gly-Pro-Ile | βb-casomorphin-7 | ||
| Tyr-Pro-Val-Glu-Pro-Phe | Neocasomorphin-6 | [111] | |
| Bovine milk α-lactalbumin | Tyr-Gly-Leu-Phe-NH2 | αb-lactorphin | [22] |
| Human milk β-casein | Tyr-Pro-Phe-Val | βh-casomorphin-4 | [22] |
| Tyr-Pro-Phe-Val-Glu | βh-casomorphin-5 | [57] | |
| Tyr-Pro-Phe-Val-Glu-Pro-Ile | βh-casomorphin-7 | [88] | |
| Tyr-Pro-Phe-Val-Glu-Pro-Ile-pro | βh-casomorphin-8 | [22,88] | |
| Human milk lactalbumin | Tyr-Gly-Leu-Phe-NH2 | αh-lactorphin | [22,94] |
| Bovine/bovine milk lactoferrin | Tyr-Leu-Gly-Ser-Gly-Tyr-OCH3 | lactoferrsoxin A | [96] |
| Arg-Tyr-Tyr-Gly-Tyr-OCH3 | lactoferrsoxin B | ||
| Lys-Tyr-Leu-Gly-Pro-Gln-Tyr-OCH3 | lactoferrsoxin C | ||
| Soy β-conglycinin | Tyr-Pro-Phe-Val-Val | Soymorphin-5 | [93] |
| Tyr-Pro-Phe-Val-Val-Asn | Soymorphin-6 | ||
| Tyr-Pro-Phe-Val-Val-Asn-Ala | Soymorphin-7 | ||
| Wheat HMW glutenin | Gly-Tyr-Tyr-Pro | gluten exorphin A4 | [78,112] |
| Gly-Tyr-Tyr-Pro-Thr | gluten exorphin A5 | ||
| Tyr-Gly-Gly-Trp | gluten exorphin B4 | ||
| Tyr-Gly-Gly-Trp-Leu | gluten exorphin B5 | ||
| Tyr-Pro-Ile-Ser-Leu | gluten exorphin C | [78,113] | |
| Spinach RuBisCo | Tyr-Pro-Leu-Asp-Leu | rubiscolin-5 | [93,114] |
| Tyr-Pro-Leu-Asp-Leu-Phe | rubiscolin-6 |
Table 4.
The opioid activity of exogenous peptides (IC50 in µM).
| Opioid Peptide | Opioid Activity (IC50 in µM) | µ/δ Ratio | Reference | |
|---|---|---|---|---|
| Mouse (vas Deferens) (δ) | Guinea-Pig (ileum) (µ) | |||
| rubiscolin-5 | 51 | 1110 | 21.8 | [114] |
| rubiscolin-6 | 24.4 | 748 | 30.7 | [114] |
| βb-casomorphin-4 | 84 | 22 | 0.26 | [87] |
| βb-casomorphin-5 | 40 | 6.5 | 0.16 | [87] |
| βb-casomorphin-6 | >150 | 27.4 | <0.18 | [87] |
| βb-casomorphin-7 | >200 | 57 | <0.29 | [87] |
| βh-casomorphin-4 | 750 | 19 | 0.025 | [94] |
| βh-casomorphin-5 | ND | 14 | ND | [94] |
| βh-casomorphin-6 | 350 | 25 | 0.071 | [94] |
| βh-casomorphin-8 | 540 | 25 | 0.047 | [94] |
| gluten-exorphin A4 | 70 | >1000 | ND | [112] |
| gluten exorphin A5 | 60 | 1000 | 60.7 | [112] |
| gluten exorphin B4 | 3.4 | 1.5 | 0.44 | [112] |
| gluten exorphin B5 | 0.017 | 0.05 | 2.9 | [112] |
| gluten exorphin C | 30 | 110 | 3.7 | [113] |
| soymorphin-5 | 50 | 6 | 0.12 | [93] |
| soymorphin-6 | 32 | 9.2 | 0.287 | [93] |
| soymorphin-7 | 50 | 13 | 0.26 | [93] |
| Human milk lactalbumin (α-lactorphin) | >1000 | 50 | ND | [94] |
| Bovine milk lactoferrin (lactoferrsoxin) | 4.38 | 5.68 | 0.77 | [96] |
ND—not determined, IC50 is the 50% inhibitory concentration.
Table 5.
Trials of exogenous opioid peptides in animal models.
| Opioid Peptide | Animal Model | Dosage | Administration Route | Effect | Time Duration | Reference |
|---|---|---|---|---|---|---|
| rubiscolin-5 | Mice | 3 nM/mouse | i.c.v | antinociception | Effects observed up to 30 min post-injection | [114] |
| rubiscolin-6 | Mice | 1 nM/mouse | i.c.v | |||
| rubiscolin–6 | Mice | 100 mg/kg 3 nM/mouse | Oral i.c.v | enhancement in memory consolidation | Effects observed up to 24 h post-injection | [100] |
| β-casomorphin- 4,-5,-6,-7 | Rat | 60–2000 nM | i.c.v | analgesic, naloxone reversible | Effects observed up to 30–40 min post-injection | [115] |
| β-casomorphin-5 | Mice Rat | 1mg/kg 166 nM | i.p i.v | improvement in learning and memory, analgestic | Effects observed up to 30-min post-injection Significant analgesia l0-min after injection up to 60 min post-injection | [116,117] |
| β-casomorphin-7 | Rat | 0.1–20 nM | i.c.v | food-intake stimulation | Effects observed up to 6 h post-injection | [118] |
| gluten exorphin C | Mice | 5mg/kg | i.p | improvement in learning and behavior, decreased anxiety | Effects observed up to 15–20 min post-injection | [110] |
| Gluten-exorphin B5 | Rat | 3 mg/kg | i.v | stimulated prolactin secretion | Effects observed after 20 min post-injection | [109] |
| soymorphin-5, 6, and 7 | Mice | 10–30 mg/kg or 3 mg/kg | Oral i.p | anxiolytic effect | Oral—Effects observed up to 20–25 min post-injection i.p—Effects observed <30min of post-injection | [93] |
| soymorphin-5 and 7 | Mice | 30 mg/kg or 48 µ mol/kg | oral | reduced food intake and showed anorexigenic activity | Effects observed after 2 h of oral administration | [119] |
| soymorphin-5 amide | Rat | 5 mg/kg | i.p | decreased anxiety | Effects observed after 30 min of administration | [120] |
i.c.v—intracerebroventricular; i.p—intraperitoneal; i.v—intravenous; nM—nano mol.
Table 6.
In silico databases and online tools for analysis of bioactive peptides.
| Databases Name | Address | Role |
|---|---|---|
| NeuroPIpred | https://webs.iiitd.edu.in/raghava/neuropipred | Neuropeptide database |
| NeuroPP | http://i.uestc.edu.cn/neuropeptide/neuropp/home.html | Neuropeptide database |
| BIOPEP (Bioactivity) (digestion) (Protein) (toxicity) | http://www.uwm.edu.pl/biochemia/index.php/en/biopep | Prediction for precursors of bioactive peptides, |
| ToxinPred (Toxicity) | http://crdd.osdd.net/raghava//toxinpred/ | Prediction of toxicity of peptides |
| I-TASSER (Protein Structure) | https://zhanglab.ccmb.med.umich.edu/I-TASSER/ | Structure and function prediction |
| NCBI (Protein Database) | https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/ | Protein sequences information |
| AlgPred (Toxicity) | http://crdd.osdd.net/raghava//algpred/ | Prediction of toxicity of peptides |
| ProtParam (phytochemical) | http://web.expasy.org/protparam/ | Compute GRAVY (grand average of hydropathicity) |
| UniProtKB (Protein database) | http://www.uniprot.org/ | Structure and sequences information |
| APD (Peptide database) | http://aps.unmc.edu/AP/main.html | Bioactive peptide prediction |
| AntiBP2 (Bioactivity prediction) | http://crdd.osdd.net/raghava//antibp2/ | Antibacterial peptide prediction |
| PEPstrMOD (Peptide database) | http://osddlinux.osdd.net/raghava/pepstrmod | Prediction of tertiary structures |
Table 7.
Stability half-life of opioid peptides.
| Sequence | Half-Life (Mouse Brain) [min] | Reference |
|---|---|---|
| [d-Ala2, p-Cl-Phe4]EM-1 | >300 | [181] |
| [Dmt1,Nip2]EM-1 | 30.9 ± 3.29 | [182] |
| [(2S,3S)β-MePhe4]EM-2 | 35.8 ± 1.8 | [68] |
| [(1S,2R)ACHC2]EM-1 | >12 h | [183] |
| Guanidino-[d-Pro2Gly3, p-Cl-Phe5]EM-1 | 187.3 ± 24 | [181] |
| [(1S,2R)ACPC2]EM-2 | >12 h | [183] |
| [Dmt1,Nip2]EM-2 | 10.7 ± 0.3 | [182] |
| Guanidino-[d-Pro2-Gly3]EM-1 | 111.8 ± 19.2 | [181] |
| Guanidino-[Sar2]EM-1 | 43.9 ± 2.4 | [181] |
EM-1—endomorphin 1, EM-2—endomorphin 2, cis-/trans-2-aminocyclopentanecarboxylic acid (ACPC), cis-/trans-2-aminocyclohexanecarboxylic acid (ACHC), piperidine-3-carboxylic acid (Nip), 2′6′-dimethyltyrosine (Dmt).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tyagi, A.; Daliri, E.B.-M.; Kwami Ofosu, F.; Yeon, S.-J.; Oh, D.-H. Food-Derived Opioid Peptides in Human Health: A Review. Int. J. Mol. Sci. 2020, 21, 8825. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228825
AMA Style
Tyagi A, Daliri EB-M, Kwami Ofosu F, Yeon S-J, Oh D-H. Food-Derived Opioid Peptides in Human Health: A Review. International Journal of Molecular Sciences. 2020; 21(22):8825. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228825
Chicago/Turabian StyleTyagi, Akanksha, Eric Banan-Mwine Daliri, Fred Kwami Ofosu, Su-Jung Yeon, and Deog-Hwan Oh. 2020. "Food-Derived Opioid Peptides in Human Health: A Review" International Journal of Molecular Sciences 21, no. 22: 8825. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228825
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.