Effects of Excess Manganese on the Xylem Sap Protein Profile of Tomato (Solanum lycopersicum) as Revealed by Shotgun Proteomic Analysis

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Xylem Sap Collection and Mineral Composition
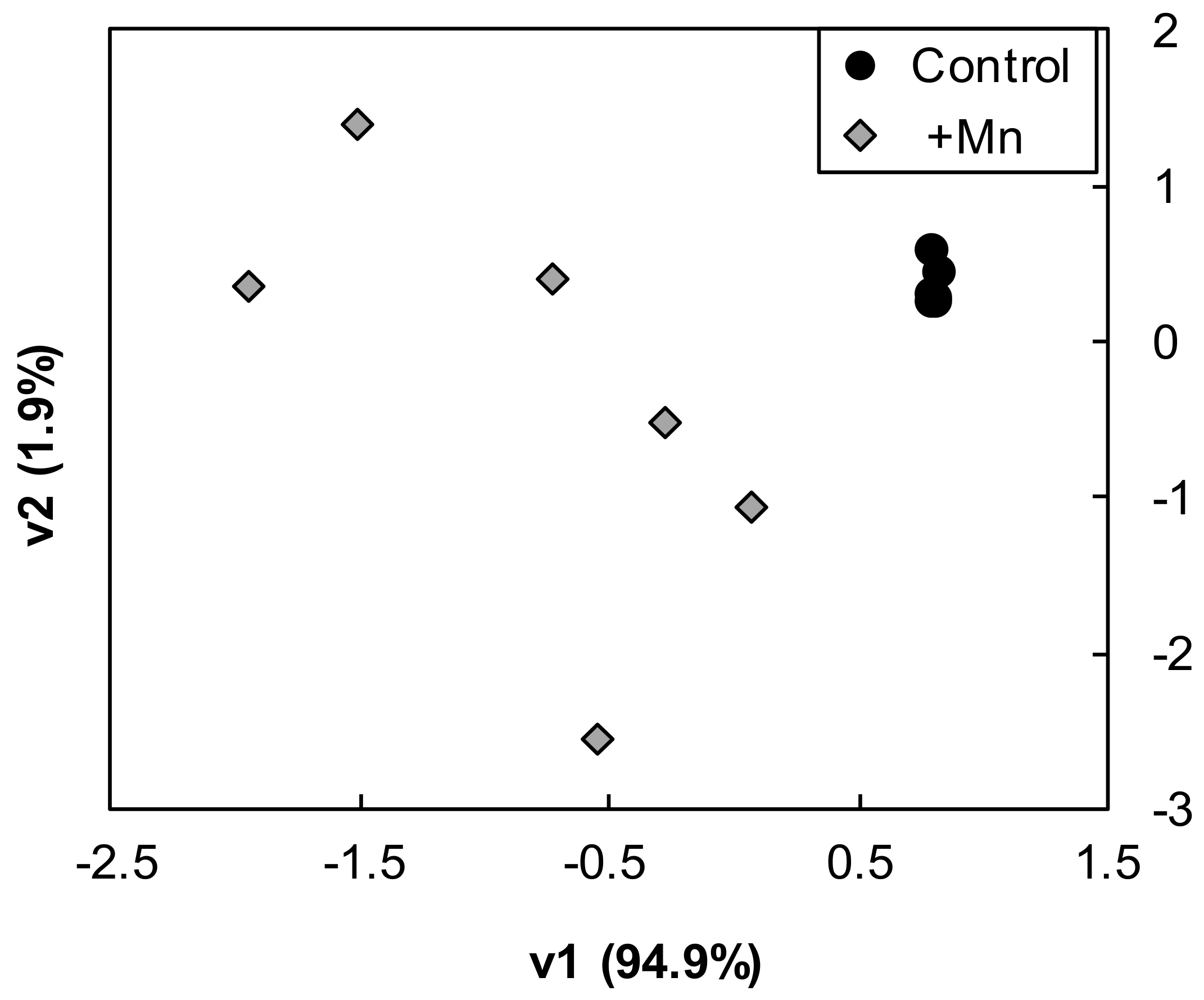
2.2. Proteins Detected, Class Annotation and PCA Analysis
2.3. Effect of Excess Mn on the Xylem Sap Protein Profile
2.4. Protein Species Decreasing in Abundance
2.5. Protein Species Increasing in Abundance
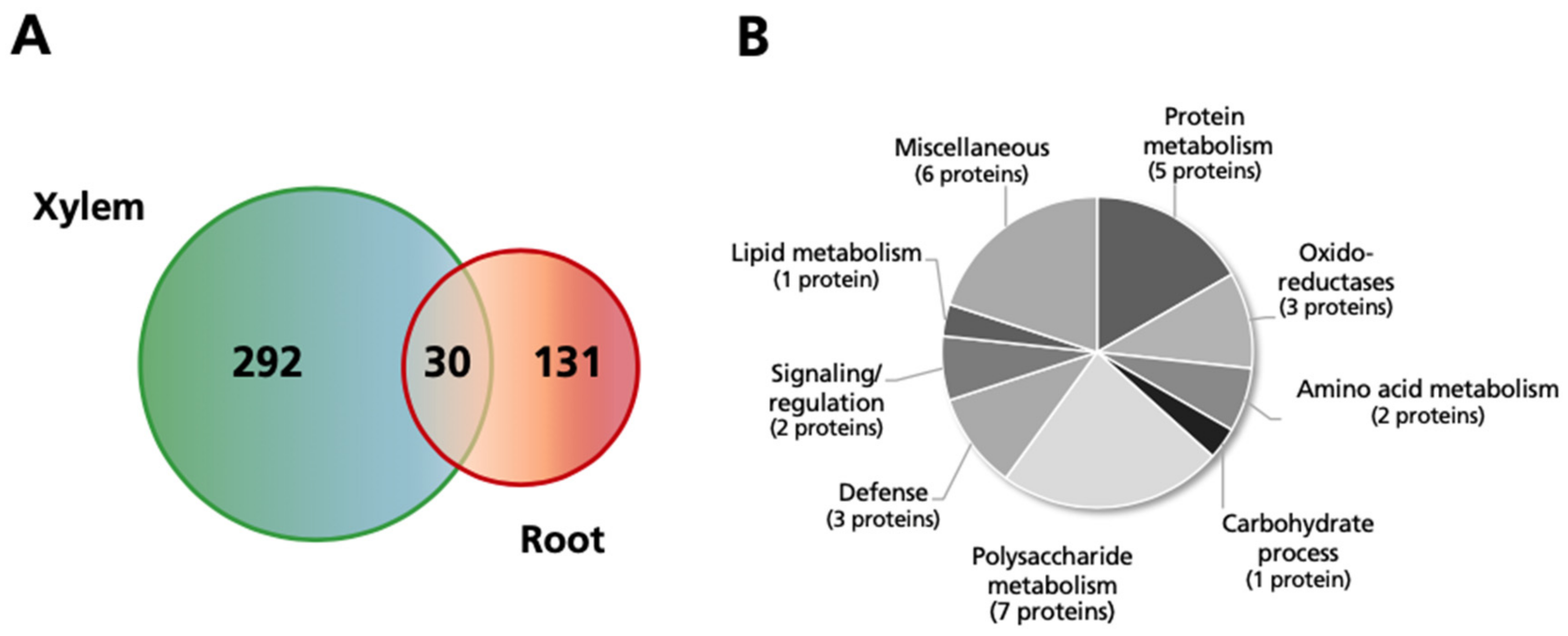
2.6. Comparison of Changes Observed in the Xylem and Root Protein Profiles in Response to Excess Mn
3. Discussion
3.1. Excess Mn Affects the Metabolism of the Cell Wall
3.2. Excess Mn Alters Protein Turnover
3.3. Excess Mn Alters Defense Mechanisms
3.4. Excess Mn Effects on Signaling and Regulation Mechanisms
3.5. Excess Mn Affects General Metabolism
3.6. Comparison of Changes Induced by Excess Mn in the Root and Xylem Sap Proteomes
4. Materials and Methods
4.1. Plant Material and Sampling
4.2. Experimental Design
4.3. Mineral Analysis
4.4. Protein Extraction
4.5. Label Free Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
4.6. Root Protein Profiling and Comparison with the Xylem Sap
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Boston, MA, USA, 2012. [Google Scholar]
- Pittman, J. Managing the manganese: Molecular mechanisms of manganese transport and homeostasis. New Phytol. 2008, 167, 733–742. [Google Scholar] [CrossRef]
- Millaleo, R.; Reyes-Díaz, M.; Ivanov, A.G.; Mora, M.L.; Alberdi, M. Manganese as essential and toxic element for plants: Transport, accumulation and resistance mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Socha, A.L.; Guerinot, M.L. Mn-euvering manganese: The role of transporter gene family members in manganese uptake and mobilization in plants. Front. Plant Sci. 2014, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, M.; Sharma, A. Manganese in cell metabolism of higher plants. Bot. Rev. 1991, 57, 117–149. [Google Scholar] [CrossRef]
- Graham, M.H.; Haynes, R.J.; Meyer, J.H. Changes in soil chemistry and aggregate stability induced by fertilizer applications, burning and trash retention on a long-term sugarcane experiment in South Africa. Eur. J. Soil Sci. 2002, 53, 589–598. [Google Scholar] [CrossRef]
- Driscoll, C.T.; Lawrence, G.B.; Bulger, A.J.; Butler, T.J.; Cronan, C.S.; Eagar, C.; Lambert, K.F.; Likens, G.E.; Stoddard, J.L.; Weathers, K.C. Acidic deposition in the Northeastern United States: Sources and inputs, ecosystem effects, and management strategies. BioScience 2001, 51, 180–198. [Google Scholar] [CrossRef] [Green Version]
- Foy, C.D. Physiological effects of hydrogen, aluminum and manganese toxicities in acid soils. In Soil Acidicity and Liming, 2nd ed.; Adams, J., Ed.; American Society of Agronomy: Madison, WI, USA, 1984; pp. 57–97. [Google Scholar]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, K.W.; Goulding, K.W.; Vitousek, P.M.; Zhang, F.S. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernando, D.R.; Lynch, J.P. Manganese phytotoxicity: New light on an old problem. Ann. Bot. 2010, 116, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Zhu, Z.; Xu, M.; Qian, Q.; Yu, J. Effect of excess manganese on the antioxidant system in Cucumis sativus L. under two light intensities. Environ. Exp. Bot. 2006, 58, 197–205. [Google Scholar] [CrossRef]
- Venkatesan, S.; Hemalatha, K.V.; Jayaganesh, S. Characterization of manganese toxicity and its influence on nutrient uptake, antioxidant enzymes and biochemical parameters in tea. Res. J. Phytochem. 2007, 1, 52–60. [Google Scholar]
- Führs, H.; Behrens, C.; Gallien, S.; Heintz, D.; Van Dorsselaer, A.; Braun, H.P.; Horst, W.J. Physiological and proteomic characterization of manganese sensitivity and tolerance in rice (Oryza sativa) in comparison with barley (Hordeum vulgare). Ann. Bot. 2010, 105, 1129–1140. [Google Scholar]
- Gangwar, S.; Singh, V.P.; Prasad, S.M.; Maurya, J.N. Modulation of manganese toxicity in Pisum sativum L. seedlings by kinetin. Sci. Hortic. 2010, 126, 467–474. [Google Scholar] [CrossRef]
- Führs, H.; Specht, A.; Erban, A.; Kopka, J.; Horst, W.J. Functional associations between the metabolome and manganese tolerance in Vigna unguiculata. J. Exp. Bot. 2012, 63, 329–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Xu, G.; Mou, D.; Wang, J.; Ma, J. Subcellular Mn compartation, anatomic and biochemical changes of two grape varieties in response to excess manganese. Chemosphere 2012, 89, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Singh, S. Biochemical markers and nutrient constraints diagnosis in Citrus: A perspective. J. Plant Nutr. 2006, 29, 827–855. [Google Scholar] [CrossRef]
- Millaleo, R.; Reyes-Díaz, M.; Alberdi, M.; Ivanov, A.G.; Krol, M.; Huner, N.P. Excess manganese differentially inhibits photosystem I versus II in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.P.; Qi, Y.P.; You, X.; Yang, L.T.; Guo, P.; Ye, X.; Zhou, X.X.; Ke, F.J.; Chen, L.S. Leaf cDNA-AFLP analysis of two citrus species differing in manganese tolerance in response to long-term manganese-toxicity. BMC Genom. 2013, 14, 621. [Google Scholar] [CrossRef] [Green Version]
- You, X.; Yang, L.T.; Lu, Y.B.; Li, H.; Zhang, S.Q.; Chen, L.S. Proteomic changes of citrus roots in response to long-term manganese toxicity. Trees 2014, 28, 1383–1399. [Google Scholar] [CrossRef]
- Chen, Z.; Yan, W.; Sun, L.; Tian, J.; Liao, H. Proteomic analysis reveals growth inhibition of soybean roots by manganese toxicity is associated with alteration of cell wall structure and lignification. J. Proteom. 2016, 143, 151–160. [Google Scholar] [CrossRef]
- Wu, Z.; Liang, F.; Hong, B.; Young, J.C.; Sussman, M.R.; Harper, J.F.; Sze, H. An endoplasmic reticulum-bound Ca2+/Mn2+ pump, ECA1, supports plant growth and confers tolerance to Mn2+ stress. Plant Physiol. 2002, 130, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Delhaize, E.; Kataoka, T.; Hebb, D.M.; White, R.G.; Ryan, P.R. Genes encoding proteins of the cation diffusion facilitator family that confer manganese tolerance. Plant Cell 2003, 15, 1131–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, C.; Fu, X.; Chen, X.; Shi, J.; Chen, Y. Accumulation and interaction of calcium and manganese in Phytolacca. Am. Plant Sci. 2009, 177, 601–606. [Google Scholar] [CrossRef]
- Oparka, K.J.; Cruz, S.S. The great escape: Phloem transport and unloading of macromolecules. Annu. Rev. Plant Biol. 2000, 51, 323–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, W.J.; Groover, A.; Lichtenberger, R.; Furuta, K.; Yadav, S.R.; Helariutta, Y.; He, X.Q.; Fukuda, H.; Kang, J.; Brady, S.M.; et al. The plant vascular system: Evolution, development and functions. J. Integr. Plant Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.B. Long-distance transport. In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; Wiley Blackwell: Rockville, MD, USA, 2000; pp. 729–784. [Google Scholar]
- Dickson, R.E. Xylem translocation of amino-acids from roots to shoots in cottonwood plants. Can. J. For. Res. 1979, 9, 374–378. [Google Scholar] [CrossRef]
- Friedman, R.; Levin, N.; Altman, A. Presence and identification of polyamines in xylem and phloem exudates of plants. Plant Physiol. 1986, 82, 1154–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Millán, A.F.; Morales, F.; Abadía, A.; Abadía, J. Effects of iron deficiency on the composition of the leaf apoplastic fluid and xylem sap in sugar beet. Implications for iron and carbon transport. Plant Physiol. 2000, 124, 873–884. [Google Scholar] [CrossRef] [Green Version]
- Escher, P.; Eiblmeier, M.; Hetzger, I.; Rennenberg, H. Seasonal and spatial variation of carbohydrates in mistletoes (Viscum album) and the xylem sap of its hosts (Populus x euamericana and Abies alba). Physiol. Plant. 2004, 120, 212–219. [Google Scholar] [CrossRef]
- Carella, P.; Wilson, D.C.; Kempthorne, C.J.; Cameron, R.K. Vascular sap proteomics: Providing insight into long-distance signaling during stress. Front. Plant Sci. 2016, 7, 651. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Celma, J.; Ceballos-Laita, L.; Grusak, M.A.; Abadía, J.; López-Millán, A.F. Plant fluid proteomics: Delving into the xylem sap, phloem sap and apoplastic fluid proteomes. BBA-Proteins Proteom. 2016, 1864, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, H. Xylogenesis: Initiation, progression, and cell death. Annu. Rev. Plant Biol. 1996, 47, 199–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, S.; Goodger, J.Q.D.; Marsh, E.L.; Chen, S.; Asirvatham, V.S.; Schachtman, D.P. Characterization of the maize xylem sap proteome. J. Proteome Res. 2006, 5, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Ligat, L.; Lauber, E.; Albenne, C.; Clemente, H.S.; Valot, B.; Zivy, M.; Pont-Lezica, R.; Arlat, M.; Jamet, E. Analysis of the xylem sap proteome of Brassica oleracea reveals a high content in secreted proteins. Proteomics 2011, 11, 1798–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Chung, K.; Lin, W.; Jiang, L. Protein secretion in plants: Conventional and unconventional pathways and new techniques. J. Exp. Bot. 2018, 69, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Bendtsen, J.D.; Kiemer, L.; Fausbøll, A.; Brunak, S. Non-classical protein secretion in bacteria. BMC Microbiol. 2005, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Lonsdale, A.; Davis, M.J.; Doblin, M.S.; Bacic, A. Better than nothing? Limitations of the prediction tool SecretomeP in the search for leaderless secretory proteins (LSPs) in plants. Front. Plant Sci. 2016, 7, 1451. [Google Scholar] [CrossRef] [Green Version]
- Lonsdale, A. Computational biology methods for identifying leaderless secretory proteins in Arabidopsis thaliana and other plant species. Ph.D. Thesis, University of Melbourne, Parkville, VIC, Australia, 2019. [Google Scholar]
- Ceballos-Laita, L.; Gutierrez-Carbonell, E.; Takahashi, D.; Abadía, A.; Uemura, M.; Abadía, J.; López-Millán, A.F. Effects of Fe and Mn deficiencies on the protein profiles of tomato (Solanum lycopersicum) xylem sap as revealed by shotgun analyses. J. Proteom. 2018, 170, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Ceballos-Laita, L.; Gutierrez-Carbonell, E.; Imai, H.; Abadía, A.; Uemura, M.; Abadía, J.; López-Millán, A.F. Effects of manganese toxicity on the protein profile of tomato (Solanum lycopersicum) roots as revealed by two complementary proteomic approaches, two-dimensional electrophoresis and shotgun analysis. J. Proteom. 2018, 185, 51–63. [Google Scholar] [CrossRef]
- González, A.; Steffen, K.L.; Lynch, J.P. Light and excess manganese. Implications for oxidative stress in common bean. Plant Physiol. 1998, 118, 493–504. [Google Scholar]
- Fecht-Christoffers, M.M.; Braun, H.P.; Lemaitre-Guillier, C.; Van Dorsselaer, A.; Horst, W.J. Effect of manganese toxicity on the proteome of the leaf apoplast in cowpea. Plant Physiol. 2003, 133, 1935–1946. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Sun, L.; Liu, P.; Liu, G.; Tian, J.; Liao, H. Malate synthesis and secretion mediated by a manganese-enhanced malate dehydrogenase confers superior manganese tolerance in Stylosanthes guianensis. Plant Physiol. 2014, 167, 176–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Führs, H.; Hartwig, M.; Molina, L.E.; Heintz, D.; Van Dorsselaer, A.; Braun, H.; Horst, W.J. Early manganese-toxicity response in Vigna unguiculata L.—A proteomic and transcriptomic study. Proteomics 2008, 8, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Kaida, R.; Serada, S.; Norioka, N.; Norioka, S.; Neumetzler, L.; Pauly, M.; Sampedro, J.; Zarra, I.; Hayashi, T.; Kaneko, T.S. Potential role for purple acid phosphatase in the dephosphorylation of wall proteins in tobacco cells. Plant Physiol. 2010, 153, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Vecchio, H.A.; Ying, S.; Park, J.; Knowles, V.L.; Kanno, S.; Tanoi, K.; She, Y.-M.; Plaxton, W.C. The cell wall-targeted purple acid phosphatase AtPAP25 is critical for acclimation of Arabidopsis thaliana to nutritional phosphorus deprivation. Plant J. 2014, 80, 569–581. [Google Scholar] [CrossRef]
- Zhang, Z.; Xin, W.; Wang, S.; Zhang, X.; Dai, H.; Sun, R.; Frazier, T.; Zhang, B.; Wang, Q. Xylem sap in cotton contains proteins that contribute to environmental stress response and cell wall development. Funct. Integr. Genom. 2015, 15, 17–26. [Google Scholar] [CrossRef]
- MEROPS the Peptidase Database. Available online: https://www.ebi.ac.uk/merops/ (accessed on 7 October 2020).
- Vose, P.B.; Randall, P.J. Resistance to aluminum and manganese toxicities in plants related to variety and cation-exchange capacity. Nature 1962, 196, 85–86. [Google Scholar] [CrossRef]
- Minic, Z. Physiological roles of plant glycoside hydrolases. Planta 2008, 227, 723–740. [Google Scholar] [CrossRef]
- CAZy. Carbohydrate-Active enZYmes. Available online: http://www.cazy.org/ (accessed on 7 October 2020).
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, 233–238. [Google Scholar] [CrossRef]
- Lavres, J.; Malavolta, E.; Nogueira, N.L.; Moraes, M.F.; Reis, A.R.; Rossi, M.L.; Cabral, C.P. Changes in anatomy and root cell ultrastructure of soybean genotypes under manganese stress. Rev. Bras. Cienc. Solo 2009, 33, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Freudenberg, K. Biosynthesis and constitution of lignin. Nature 1959, 183, 1152–1155. [Google Scholar] [CrossRef]
- Castillo, F.J.; Greppin, H. Balance between anionic and cationic extracellular peroxidase activities in Sedum album leaves after ozone exposure. Analysis by high-performance liquid chromatography. Physiol. Plant. 1986, 68, 201–208. [Google Scholar] [CrossRef]
- Fernández-Pérez, F.; Pomar, F.; Pedreño, M.A.; Novo-Uzal, E. The suppression of AtPrx52 affects fibers but not xylem lignification in Arabidopsis by altering the proportion of syringyl units. Physiol. Plant. 2015, 154, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pérez, F.; Pomar, F.; Pedreño, M.A.; Novo-Uzal, E. Suppression of Arabidopsis peroxidase 72 alters cell wall and phenylpropanoid metabolism. Plant Sci. 2015, 239, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pérez, F.; Vivar, T.; Pomar, F.; Pedreño, M.A.; Novo-Uzal, E. Peroxidase 4 is involved in syringyl lignin formation in Arabidopsis thaliana. J. Plant Physiol. 2015, 175, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. New Phytol. 2016, 209, 1395–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreps, J.A.; Wu, Y.; Chang, H.S.; Zhu, T.; Wang, X.; Harper, J.F. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol. 2002, 130, 2129–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokunaga, N.; Kaneta, T.; Sato, S.; Sato, Y. Analysis of expression profiles of three peroxidase genes associated with lignification in Arabidopsis thaliana. Physiol. Plant. 2009, 136, 237–249. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, B.H.; Lim, C.J.; Lim, C.O.; Nam, K.H. Constitutive activation of stress-inducible genes in a brassinosteroid-insensitive 1 (bri1) mutant results in higher tolerance to cold. Physiol. Plant. 2010, 138, 191–204. [Google Scholar] [CrossRef]
- Santandrea, G.; Pandolfini, T.; Bennici, A. A physiological characterization of Mn-tolerant tobacco plants selected by in vitro culture. Plant Sci. 2000, 150, 163–170. [Google Scholar] [CrossRef]
- Fecht-Christoffers, M.M.; Führs, H.; Braun, H.P.; Horst, W.J. The role of hydrogen peroxide-producing and hydrogen peroxide-consuming peroxidases in the leaf apoplast of cowpea in manganese tolerance. Plant Physiol. 2006, 140, 1451–1463. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.F.; Eriksson, K.E.L. Laccase and the deposition of lignin in vascular plants. Holzforschung 1994, 48, 21–33. [Google Scholar] [CrossRef]
- Ezaki, B.; Sasaki, K.; Matsumoto, H.; Nakashima, S. Functions of two genes in aluminium (Al) stress resistance: Repression of oxidative damage by the AtBCB gene and promotion of efflux of Al ions by the NtGDI1gene. J. Exp. Bot. 2005, 56, 2661–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Zhang, Z.; Yang, G.; Mao, J.; Xu, F. Ultrastructural topochemistry of cell wall polymers in Populus nigra by transmission electron microscopy and Raman imaging. BioResources 2011, 6, 3944–3959. [Google Scholar]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Buhner-Zaharieva, T.; Abadía, J. Iron deficiency enhances the levels of ascorbate, glutathione, and related enzymes in sugar beet roots. Protoplasma 2003, 221, 269–275. [Google Scholar] [CrossRef]
- Lan, P.; Li, W.; Wen, T.N.; Shiau, J.Y.; Wu, Y.C.; Lin, W.; Schmidt, W. iTRAQ protein profile analysis of Arabidopsis roots reveals new aspects critical for iron homeostasis. Plant Physiol. 2011, 155, 821–834. [Google Scholar] [CrossRef] [Green Version]
- Schaller, A. A cut above the rest: The regulatory function of plant proteases. Planta 2004, 220, 183–197. [Google Scholar] [CrossRef]
- Xia, Y.; Suzuki, H.; Borevitz, J.; Blount, J.; Guo, Z.; Patel, K.; Dixon, R.A.; Lamb, C. An extracellular aspartic protease functions in Arabidopsis disease resistance signaling. EMBO J. 2004, 23, 980–988. [Google Scholar] [CrossRef]
- Meng, L.; Wong, J.H.; Feldman, L.J.; Lemaux, P.G.; Buchanan, B.B. A membrane-associated thioredoxin required for plant growth moves from cell to cell, suggestive of a role in intercellular communication. Proc. Natl. Acad. Sci. USA 2010, 107, 3900–3905. [Google Scholar] [CrossRef] [Green Version]
- Kawano, T. Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep. 2003, 21, 829–837. [Google Scholar] [CrossRef]
- Edreva, A.M. Induction of ‘pathogenesis-related’ proteins in tobacco leaves by physiological (non-pathogenic) disorders. J. Exp. Bot. 1990, 41, 701–703. [Google Scholar] [CrossRef]
- Milne, T.J.; Abbenante, G.; Tyndall, J.D.A.; Halliday, J.; Lewis, R.J. Isolation and characterization of a cone snail protease with homology to CRISP proteins of the pathogenesis-related protein superfamily. J. Biol. Chem. 2003, 278, 31105–31110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, G.M.; Scanlon, M.J.; Swarbrick, J.; Curtis, S.; Gallant, E.; Dulhunty, A.F.; O’Bryan, M.K. The cysteine-rich secretory protein domain of Tpx-1 is related to ion channel toxins and regulates ryanodine receptor Ca2+ signaling. J. Biol. Chem. 2006, 281, 4156–4163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacMillan, C.P.; Mansfield, S.D.; Stachurski, Z.H.; Evans, R.; Southerton, S.G. Fasciclin-like arabinogalactan proteins: Specialization for stem biomechanics and cell wall architecture in Arabidopsis and Eucalyptus. Plant J. 2010, 62, 689–703. [Google Scholar] [CrossRef]
- Seifert, G.J.; Xue, H.; Acet, T. The Arabidopsis thaliana FASCICLIN LIKE ARABINOGALACTAN PROTEIN 4 gene acts synergistically with abscisic acid signalling to control root growth. Ann. Bot. 2014, 114, 1125–1133. [Google Scholar] [CrossRef] [Green Version]
- Lafarguette, F.; Leple, J.C.; Dejardin, A.; Laurans, F.; Costa, G.; Lesage-Descauses, M.C.; Pilate, G. Poplar genes encoding fasciclin-like arabinogalactan proteins are highly expressed in tension wood. New Phytol. 2004, 164, 107–121. [Google Scholar] [CrossRef]
- Brown, D.M.; Zeef, L.A.H.; Ellis, J.; Goodacre, R.; Turner, S.R. Identification of novel genes in Arabidopsis involved in secondary cell wall formation using expression profiling and reverse genetics. Plant Cell 2005, 17, 2281–2295. [Google Scholar] [CrossRef]
- Persson, S.; Wei, H.; Milne, J.; Page, G.P.; Somerville, C.R. Identification of genes required for cellulose synthesis by regression analysis of public microarray data sets. Proc. Natl. Acad. Sci. USA 2008, 102, 8633–8638. [Google Scholar] [CrossRef] [Green Version]
- Eyüboglu, B.; Pfister, K.; Haberer, G.; Chevalier, D.; Fuchs, A.; Mayer, K.F.X.; Schneitz, K. Molecular characterisation of the STRUBBELIG-RECEPTOR FAMILY of genes encoding putative leucine-rich repeat receptor-like kinases in Arabidopsis thaliana. BMC Plant Biol. 2007, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Miyazono, K.-I.; Miyakawa, T.; Sawano, Y.; Kubota, K.; Kang, H.-J.; Asano, A.; Miyauchi, Y.; Takahashi, M.; Zhi, Y.; Fujita, Y.; et al. Structural basis of abscisic acid signalling. Nat. Cell Biol. 2009, 462, 609. [Google Scholar] [CrossRef]
- Hao, Q.; Yin, P.; Li, W.; Wang, L.; Yan, C.; Lin, Z.; Wu, J.Z.; Wang, J.; Yan, S.F.; Yan, N. The molecular basis of ABA-independent inhibition of PP2Cs by a subclass of PYL proteins. Mol. Cell. 2011, 42, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, L.; Wang, G.; Yu, L.; Zhang, Q.; Xin, Q.; Wu, W.; Gong, Z.; Chen, Z. Structural insights into the abscisic acid stereospecificity by the ABA receptors PYR/PYL/RCAR. PLoS ONE 2013, 8, e67477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benning, U.F.; Tamot, B.; Guelette, B.S.; Hoffmann-Benning, S. New aspects of phloem-mediated long-distance lipid signaling in plants. Front. Plant Sci. 2012, 3, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbaglia, A.M.; Tamot, B.; Greve, V.; Hoffmann-Benning, S. Phloem proteomics reveals new lipid-binding proteins with a putative role in lipid-mediated signaling. Front. Plant Sci. 2016, 7, 563. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.D.G.; Putterill, J.J.; Plummer, K.M.; Newcomb, R.D. The carboxylesterase gene family from Arabidopsis thaliana. J. Mol. Evol. 2003, 57, 487–500. [Google Scholar]
- Ko, M.K.; Jeon, W.B.; Kim, K.S.; Lee, H.H.; Seo, H.H.; Kim, Y.S.; Oh, B.J. A Colletotrichum gloeosporioides-induced esterase gene of nonclimacteric pepper (Capsicum annuum) fruit during ripening plays a role in resistance against fungal infection. Plant Mol. Biol. 2008, 58, 529–541. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.O.N.; Miller, G.A.D. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Bidwell, S.D.; Woodrow, I.E.; Batianoff, G.N.; Sommer-Knudsen, J. Hyperaccumulation of manganese in the rainforest tree Austromyrtus bidwillii (Myrtaceae) from Queensland, Australia. Funct. Plant Biol. 2002, 29, 899–905. [Google Scholar] [CrossRef]
- Fernando, D.R.; Mizuno, T.; Woodrow, I.E.; Baker, A.J.M.; Collins, R.N. Characterization of foliar manganese (Mn) in Mn (hyper)accumulators using X-ray absorption spectroscopy. New Phytol. 2010, 188, 1014–1027. [Google Scholar] [CrossRef]
- Tenhaken, R.; Thulke, O. Cloning of an enzyme that synthesizes a key nucleotide-sugar precursor of hemicellulose biosynthesis from soybean. UDP-glucose dehydrogenase. Plant Physiol. 1996, 112, 1127–1134. [Google Scholar] [CrossRef] [Green Version]
- Klinghammer, M.; Tenhaken, R. Genome-wide analysis of the UDP-glucose dehydrogenase gene family in Arabidopsis, a key enzyme for matrix polysaccharides in cell walls. J. Exp. Bot. 2007, 58, 3609–3621. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, J.E.; Wimmer, R.; Larsen, J.N.; Spangfort, M.D.; Otzen, D.E. The major birch allergen, Bet v 1, shows affinity for a broad spectrum of physiological ligands. J. Biol. Chem. 2002, 277, 23684–23692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Takahashi, D.; Kawamura, Y.; Uemura, M. Comparison of plasma membrane proteomic changes of Arabidopsis suspension-cultured cells (T87 Line) after cold and ABA treatment in association with freezing tolerance development. Plant Cell Physiol. 2012, 53, 543–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, D.; Li, B.; Nakayama, T.; Kawamura, Y.; Uemura, M. Shotgun proteomics of plant plasma membrane and microdomain proteins using nano-LC-MS/MS. In Plant Proteomics: Methods and Protocols; Jorrin-Novo, J.V., Komatsu, S., Weckwerth, W., Wienkoop, S., Eds.; Humana Press: Totowa, NJ, USA, 2014; pp. 481–498. [Google Scholar]
- Gutierrez-Carbonell, E.; Takahashi, D.; Lüthje, S.; González-Reyes, J.A.; Mongrand, S.; Contreras-Moreira, B.; Abadía, A.; Uemura, M.; Abadía, J.; López-Millán, A.F. A shotgun proteomic approach reveals that Fe deficiency causes marked changes in the protein profiles of plasma membrane and detergent-resistant microdomain preparations from Beta vulgaris roots. J. Proteome Res. 2014, 15, 2510–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceballos-Laita, L.; Gutierrez-Carbonell, E.; Takahashi, D.; Abadía, A.; Uemura, M.; Abadía, J.; López-Millán, A.F. Data on xylem sap from Mn- and Fe-deficient tomato plants. Data Brief 2018, 17, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Resource. Available online: http://geneontology.org/ (accessed on 24 September 2020).
- SignalP-5.0 Server. Available online: http://www.cbs.dtu.dk/services/SignalP/ (accessed on 15 September 2020).
- TargetP-2.0 Server. Available online: http://www.cbs.dtu.dk/services/TargetP/ (accessed on 15 September 2020).
- SecretomeP 2.0 Server. Available online: http://www.cbs.dtu.dk/services/SecretomeP/ (accessed on 15 September 2020).
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef]
- LSPpred. Leaderless Secretory Protein Predictor for Plants. Available online: http://lsppred.lspdb.org/ (accessed on 7 October 2020).




{kind=link}
{kind=link}
{kind=link}
| Collection Parameters | Control | Excess Mn |
|---|---|---|
| Exudation rate (mL g−1 DW h−1) | 0.56 ± 0.11 | 0.52 ± 0.21 |
| Protein yield (ng protein µL−1 xylem) | 9.0 ± 3.4 | 27.0 ± 12.5 * |
| Leaf transpiration rate (mmol H2O m−2 s−1) | 10.9 ± 0.5 | 5.2 ± 0.1 *** |
| Micronutrient concentrations (µM) | ||
| Mn | 6.5 ± 0.8 | 385.7 ± 16.1 *** |
| Fe | 47.8 ± 12.5 | 15.7 ± 2.5 ** |
| Zn | 9.5 ± 1.3 | 8.0 ± 0.9 * |
| Cu | 3.6 ± 0.8 | 3.1 ± 0.5 |
| Accession | Comp. 1 Score | Comp. 2 Score | Description | Fold +Mn/ Control |
|---|---|---|---|---|
| Polysaccharide metabolism (9 proteins) | ||||
| Solyc01g104950.3.1 | 0.001 | 0.013 | LEXYL2 (GH3) | 0.5 |
| Solyc12g056960.2.1 | −0.002 | −0.061 | glucan endo-1,3-β-d-glucosidase (GH17) | 4.0 |
| Solyc10g079860.2.1 | −0.008 | −0.034 | glucan endo-1,3-β-d-glucosidase (GH17) | 51.6 |
| Solyc01g008620.3.1 | −0.024 | 0.169 | glucan endo-1,3-β-glucosidase (GH17) | 233.6 |
| Solyc01g059965.1.1 | −0.492 | −2.371 | glucan endo-1,3- β-glucosidase (GH17) | 188.7 |
| Solyc02g086700.3.1 | −0.001 | −0.018 | glucan endo-1,3-β-glucosidase-like (GH17) | 48.8 |
| Solyc05g050130.3.1 | −0.001 | 0.002 | acidic endochitinase (GH18) | 18.3 |
| Solyc10g055800.2.1 | −0.004 | −0.041 | endochitinase (GH19) | 51.1 |
| Solyc02g082920.3.1 | −0.001 | 0.005 | chitinase (GH19) | 91.3 |
| Protein metabolism (5 proteins) | ||||
| Solyc04g078110.1.1 | 0.002 | 0.014 | serine protease SBT1 (MEROPS peptidase family S8) | 0.5 |
| Solyc06g074850.3.1 | 0.001 | 0.006 | carboxypeptidase (MEROPS peptidase family S10) | 0.3 |
| Solyc08g067100.2.1 | 0.024 | 0.047 | eukaryotic aspartyl protease family protein (MEROPS peptidase family A1) | 0.5 |
| Solyc08g079870.3.1 | −0.011 | 0.072 | Subtilisin (MEROPS peptidase family S8) | 74.3 |
| Solyc08g079900.3.1 | −0.390 | 3.028 | subtilisin-like protease (MEROPS peptidase family S8) | 57.1 |
| Oxido-reductases (5 proteins) | ||||
| Solyc06g076630.3.1 | 0.007 | 0.093 | peroxidase | 0.3 |
| Solyc12g094620.2.1 | 0.015 | 0.247 | catalase | 0.2 |
| Solyc01g105070.3.1 | −0.012 | −0.205 | LECEVI16G peroxidase precursor | 14.1 |
| Solyc06g005940.3.1 | −0.001 | −0.001 | protein disulfide-isomerase-like | 11.5 |
| Solyc09g009390.3.1 | −0.001 | −0.004 | monodehydroascorbate reductase | 2.2 |
| Defense (5 proteins) | ||||
| Solyc01g097240.3.1 | −0.003 | 0.011 | pathogenesis-related protein PR-4 | 153.6 |
| Solyc00g174340.2.1 | −0.001 | 0.003 | pathogenesis-related protein PR-1 | 27.2 |
| Solyc09g090980.3.1 | 0.003 | −0.024 | major allergen Mal d 1 | 25.7 |
| Solyc12g005720.1.1 | −0.001 | −0.012 | cysteine-rich receptor-like kinase protein | 9.8 |
| Solyc09g091000.3.1 | 0.003 | −0.016 | major allergen d 1 | 54.3 |
| Lipid metabolism (2 proteins) | ||||
| Solyc03g079880.3.1 | 0.003 | 0.043 | protease inhibitor/seed storage/lipid transfer family protein | 0.4 |
| Solyc08g067500.1.1 | 0.002 | 0.030 | non-specific lipid-transfer protein | 0.2 |
| Carbohydrate metabolism (1 protein) | ||||
| Solyc06g073190.3.1 * | 0.003 | 0.66 | fructokinase-2 | 0.7 |
| Miscellaneous (2 proteins) | ||||
| Solyc01g111170.3.1 | 0.002 | 0.032 | diageotropica/Peptidyl-prolyl cis-trans isomerase | 0.2 |
| Solyc08g074682.1.1 | 0.001 | −0.015 | polyphenol oxidase precursor | 22.3 |
| # | Accession ITAG3.2 | Accession UniProt | Change +Mn/Control | Description | Category | Prediction Tool |
|---|---|---|---|---|---|---|
| Polysaccharide metabolism (13 proteins) | ||||||
| 1 | Solyc09g092170.2.1 | A0A3Q7IA20 | 0.1 | β-galactosidase STBG2 (GH35) | CPS | TarP, SecP |
| 2 | Solyc07g062210.3.1 | A0A3Q7HE74 | 0.1 | protein trichome birefringence-like 41 | CPS | SigP, TarP, SecP |
| 3 | Solyc04g016470.3.1 | A0A3Q7GST1 | 111.6 | glucan endo-1,3-β-D-glucosidase (GH17) | CPS | TarP, SecP |
| 4 | Solyc10g079860.2.1 | A0A3Q7IKF2 | 51.6 | glucan endo-1,3-β-D-glucosidase (GH17) | CPS | SigP, TarP, SecP |
| 5 | Solyc01g008620.3.1 | A0A3Q7E938 | 233.6 | glucan endo-1,3-β-glucosidase (GH17) | CPS | SigP, TarP, SecP |
| 6 | Solyc01g059965.1.1 | Q01413 | 188.7 | glucan endo-1,3-β-glucosidase (GH17) | CPS | SigP, TarP, SecP |
| 7 | Solyc02g086700.3.1 | A0A3Q7FVX4 | 48.8 | glucan endo-1,3-β-glucosidase-like (GH17) | CPS | TarP, SecP |
| 8 | Solyc07g005100.3.1 | A0A3Q7H377 | 30.4 | Chitinase/lysozyme (GH18) | CPS | SigP, TarP, SecP |
| 9 | Solyc10g055800.2.1 | A0A3Q7IHS3 | 51.1 | endochitinase (GH19) | CPS | SigP, TarP, SecP |
| 10 | Solyc10g055810.2.1 | Q05538 | 80.9 | basic 30 kDa endochitinase (GH19) | CPS | SigP, TarP, SecP |
| 11 | Solyc02g082920.3.1 | Q05539 | 91.3 | acidic extracellular 26 kD chitinase (GH19) | CPS | SigP, TarP, SecP |
| 12 | Solyc10g055820.2.1 | A0A3Q7IIQ3 | 44.4 | endochitinase (GH19) | CPS | SigP, TarP, SecP |
| 13 | Solyc11g005480.2.1 | A0A3Q7IP30 | 26.1 | citrate-binding protein-like | CPS | SigP, TarP, SecP |
| Protein metabolism (12 proteins) | ||||||
| 14 | Solyc00g005000.3.1 | A0A494G8A2 | 0.1 | eukaryotic aspartyl protease family protein | CPS | SigP, TarP, SecP |
| 15 | Solyc07g064590.3.1 | A0A3Q7HFD0 | 0.1 | inducible plastid-lipid associated protein | UPS | LSP, SecP |
| 16 | Solyc06g008170.3.1 | K4CUW3 | 0.1 | 50S ribosomal protein L14 | sUPS | SecP |
| 17 | Solyc09g066430.3.1 | A0A3Q7J0J5 | 0.1 | 60S ribosomal protein L14 | sUPS | SecP |
| 18 | Solyc08g079870.3.1 | - | 74.3 | subtilisin (MEROPS S8, clan SB) | CPS | SigP, TarP, SecP |
| 19 | Solyc08g079900.3.1 | - | 57.1 | subtilisin-like protease (MEROPS S8, clan SB) | CPS | SigP, TarP, SecP |
| 20 | Solyc06g008620.1.1 | A0A3Q7GN27 | 31.7 | tolB protein-like protein (MEROPS S9) | UPS | SPL, SecP |
| 21 | Solyc01g087850.2.1 | O82777 | 31.0 | serine protease SBT3 (MEROPS I13, clan IG) | CPS | SigP, TarP, SecP |
| 22 | Solyc03g020010.1.1 | O48625 | 93.3 | Miraculin (MEROPS I13, clan IG) | CPS | SigP, TarP, SecP |
| 23 | Solyc03g019690.1.1 | A0A3Q7FGU5 | 37.8 | kunitz-type protease inhibitor (MEROPS I13, clan IG) | CPS | SigP, TarP, SecP |
| 24 | Solyc03g098740.1.1 | A0A3Q7FNG4 | 208.5 | kunitz trypsin inhibitor (MEROPS I3, clan IC) | CPS | SigP, TarP, SecP |
| 25 | Solyc08g080630.3.1 | A0A3Q7IQ00 | 31.8 | ethylene-responsive proteinase inhibitor 1 (MEROPS I3, clan IC) | CPS | SigP, TarP, SecP |
| Oxido-reductases (6 proteins) | ||||||
| 26 | Solyc06g005160.3.1 | Q3I5C4 | 0.1 | Ascorbate peroxidase | UPS | LSP |
| 27 | Solyc04g074740.3.1 | A0A3Q7G526 | 0.0 | blue copper protein-like | CPS | SigP, TarP, SecP |
| 28 | Solyc08g066740.3.1 | A0A3Q7HR33 | 0.1 | early nodulin-like protein 1-like | CPS | SigP, TarP, SecP |
| 29 | Solyc08g028690.3.1 | A0A3Q7HMQ9 | 0.0 | NAD(P)-binding Rossmann-fold superfamily protein | UPS | LSP, SPL, SecP |
| 30 | Solyc03g006700.3.1 | A0A3Q7FFR5 | 28.7 | peroxidase (AtPrx52) | CPS | SigP, TarP, SecP |
| 31 | Solyc04g071890.3.1 | - | 71.6 | peroxidase | CPS | TarP, SecP |
| Defense (5 proteins) | ||||||
| 32 | Solyc01g097240.3.1 | P32045 | 153.6 | pathogenesis-related protein PR-4 | CPS | SigP, TarP, SecP |
| 33 | Solyc00g174340.2.1 | A0A494GA45 | 27.2 | pathogenesis-related protein 1 | CPS | SigP, TarP, SecP |
| 34 | Solyc01g106620.2.1 | B2LW68 | 61.8 | pathogenesis-related protein 1 | CPS | SigP, TarP, SecP |
| 35 | Solyc07g005380.3.1 | A0A3Q7H2K6 | 37.7 | pathogenesis-related PR-10-related/norcoclaurine synthase-like protein | UPS | SPL, SecP |
| 36 | Solyc09g090980.3.1 | - | 25.7 | major allergen Mal d 1 | UPS | SPL |
| Carbohydrate metabolism (4 proteins) | ||||||
| 37 | Solyc01g094200.3.1 | A0A3Q7EKQ6 | 0.1 | malic enzyme | UPS | LSP, SecP |
| 38 | Solyc01g101040.3.1 | A0A3Q7EPC2 | 0.1 | ATP-citrate synthase | UPS | LSP |
| 39 | Solyc01g058390.3.1 | A0A3Q7EEN4 | 0.1 | galactokinase | sUPS | SecP |
| 40 | Solyc02g088690.3.1 | A0A3Q7F9B8 | 0.1 | UDP-glucose 6-dehydrogenase family protein | UPS | LSP, SecP |
| Signaling/regulation (5 proteins) | ||||||
| 41 | Solyc02g063090.3.1 | A0A3Q7FJZ3 | 0.1 | T-complex protein 1 subunit zeta 1 | UPS | LSP |
| 42 | Solyc05g056310.3.1 | A0A3Q7GLM4 | 0.1 | T-complex protein 1 subunit gamma | UPS | LSP |
| 43 | Solyc01g086920.3.1 | A0A3Q7F2I5 | 0.0 | leucine-rich repeat receptor-like protein kinase family | CPS | SigP, TarP, SecP |
| 44 | Solyc10g050110.1.1 | A0A3Q7IFT3 | 0.1 | leucine-rich repeat receptor-like protein kinase family | CPS | SigP, TarP, SecP |
| 45 | Solyc08g082820.3.1 | A0A3Q7HX02 | 31.3 | tomato BiP (binding protein)/grp78 (HSP70) | CPS | SigP, TarP, SecP |
| Lipid metabolism (1 protein) | ||||||
| 46 | Solyc01g107990.3.1 | A0A3Q7FCU3 | 0.1 | PI-PLC X domain-containing protein | CPS | SigP, TarP, SecP |
| Miscellaneous (4 proteins) | ||||||
| 47 | Solyc03g115630.3.1 | A0A3Q7FR13 | 0.1 | carbamoyl-phosphate synthase | UPS | LSP, SPL, SecP |
| 48 | Solyc06g005360.3.1 | A0A3Q7GNQ9 | 0.1 | actin-depolymerizing factor family protein | sUPS | SecP |
| 49 | Solyc07g064160.3.1 | A0A3Q7HFB9 | 0.1 | thiamine thiazole synthase | sUP | SecP |
| 50 | Solyc10g049970.2.1 | A0A3Q7IFZ6 | 24.9 | kynurenine formamidase | CPS | SigP, TarP, SecP |
| Unknown function (1 protein) | ||||||
| 51 | Solyc06g035920.3.1 | A0A3Q7GU06 | 0.1 | remorin | sUPS | SecP |
| Xylem sap | Root | |||||
|---|---|---|---|---|---|---|
| Category | Prediction Tool | Fold +Mn/Control | Fold +Mn/Control | |||
| Proteins decreasing in both proteomes (16 proteins) | ||||||
| Polysaccharide metabolism | Solyc01g104950.3.1 | β -D-xylosidase 2 precursor (LEXYL2) | CPS | SigP, TarP, SecP | 0.5 | 0.4 |
| Solyc05g012070.3.1 | UDP-glucose:protein transglucosylase-like protein | UPS | SPL | 0.2 | 0.5 | |
| Protein metabolism | Solyc02g068740.3.1 | glycine cleavage system H family protein (MEROPS peptidase family C1) | CPS | SecP | 0.3 | 0.5 |
| Solyc12g008630.2.1 | mitochondrial-processing peptidase subunit α-like (MEROPS peptidase family M16) | UPS | LSP, SecP | 0.4 | 0.3 | |
| Solyc02g081700.1.1 | proteasome subunit α type (MEROPS peptidase family T1A) | UPS | LSP, SecP | 0.5 | 0.4 | |
| Solyc06g073310.3.1 | 60S ribosomal protein l9 | sUPS | SecP | 0.5 | 0.5 | |
| Oxido-reductases | Solyc06g005160.3.1 | ascorbate peroxidase | UPS | LSP | 0.1 | 0.2 |
| Solyc01g107590.3.1 | cinnamyl alcohol dehydrogenase | UPS | LSP | 0.2 | 0.3 | |
| Amino acid metabolism | Solyc02g080810.3.1 | aminomethyltransferase | UPS | LSP, SecP | 0.4 | 0.5 |
| Solyc01g080280.3.1 * | glutamine synthetase | lnS | LSP | 0.4 | 0.5 | |
| Carbohydrate metabolism | Solyc08g080140.3.1 | 3,5-epimerase/4-reductase | sUPS | SecP | 0.3 | 0.2 |
| Signaling/regulation | Solyc06g065520.3.1 | T-complex protein 1 subunit eta | UPS | LSP | 0.3 | 0.2 |
| Miscellaneous | Solyc02g087300.1.1 | transducin/WD40 repeat-like superfamily protein | UPS | SPL, SecP | 0.4 | 0.5 |
| Solyc01g006280.3.1 | formate-tetrahydrofolate ligase | UPS | LSP, SPL | 0.4 | 0.3 | |
| Solyc01g109660.2.1 | meloidogyne-induced giant cell protein DB275 (glycine-rich RNA-binding protein) | lnS | None | 0.3 | 0.5 | |
| Solyc12g098150.2.1 | Aldo/keto reductase | lnS | None | 0.2 | 0.5 | |
| Proteins increasing in both proteomes (12 proteins) | ||||||
| Polysaccharide metabolism | Solyc01g059965.1.1 | β-1,3-glucanase (GH17) | CPS | SigP, TarP, SecP | 188.7 | 8.3 |
| Solyc07g005100.3.1 | chitinase/lysozyme (GH18) | CPS | SigP, TarP, SecP | 30.4 | 3.4 | |
| Solyc10g055800.2.1 | chitinase (GH19) | CPS | SigP, TarP, SecP | 51.1 | 3.6 | |
| Solyc10g055810.2.1 | chitinase Z15140 (GH19) | CPS | SigP, TarP, SecP | 80.9 | 4.9 | |
| Solyc10g055820.2.1 | chitinase (GH19) | CPS | SigP, TarP, SecP | 44.4 | 9.5 | |
| Protein metabolism | Solyc03g019690.1.1 | Kunitz-type protease inhibitor (MEROPS I3, clan IC) | CPS | SigP, TarP, SecP | 37.8 | 105.3 |
| Oxido-reductases | Solyc03g006700.3.1 | peroxidase | CPS | SigP, TarP, SecP | 28.7 | 8.4 |
| Defense | Solyc01g097240.3.1 | pathogenesis-related protein PR-4 | CPS | SigP, TarP, SecP | 153.6 | 12.2 |
| Solyc01g106620.2.1 | pathogenesis-related protein 1 | CPS | SigP, TarP, SecP | 61.8 | 4.3 | |
| Solyc09g091000.3.1 | Major allergen d 1 | lnS | None | 54.3 | 10.5 | |
| Miscellaneous | Solyc10g049970.2.1 | kynurenine formamidase | CPS | SigP, TarP, SecP | 24.9 | 32.6 |
| Solyc09g090430.3.1 | cyanate hydratase | lnS | None | 3.4 | 2.4 | |
| Proteins with opposite responses in root and xylem sap proteomes (2 proteins) | ||||||
| Lipid metabolism | Solyc12g017460.1.1 | GDSL esterase/lipase At1g28590-like | CPS | SigP, TarP, SecP | 13.5 | 0.5 |
| Signaling/regulation | Solyc01g106210.3.1 | heat shock protein 70 | UPS | LSP | 3.4 | 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceballos-Laita, L.; Gutierrez-Carbonell, E.; Takahashi, D.; Lonsdale, A.; Abadía, A.; Doblin, M.S.; Bacic, A.; Uemura, M.; Abadía, J.; López-Millán, A.F. Effects of Excess Manganese on the Xylem Sap Protein Profile of Tomato (Solanum lycopersicum) as Revealed by Shotgun Proteomic Analysis. Int. J. Mol. Sci. 2020, 21, 8863. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228863
Ceballos-Laita L, Gutierrez-Carbonell E, Takahashi D, Lonsdale A, Abadía A, Doblin MS, Bacic A, Uemura M, Abadía J, López-Millán AF. Effects of Excess Manganese on the Xylem Sap Protein Profile of Tomato (Solanum lycopersicum) as Revealed by Shotgun Proteomic Analysis. International Journal of Molecular Sciences. 2020; 21(22):8863. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228863
Chicago/Turabian StyleCeballos-Laita, Laura, Elain Gutierrez-Carbonell, Daisuke Takahashi, Andrew Lonsdale, Anunciación Abadía, Monika S. Doblin, Antony Bacic, Matsuo Uemura, Javier Abadía, and Ana Flor López-Millán. 2020. "Effects of Excess Manganese on the Xylem Sap Protein Profile of Tomato (Solanum lycopersicum) as Revealed by Shotgun Proteomic Analysis" International Journal of Molecular Sciences 21, no. 22: 8863. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228863