Abnormal Expression of Mitochondrial Ribosomal Proteins and Their Encoding Genes with Cell Apoptosis and Diseases

Abstract
:1. Introduction
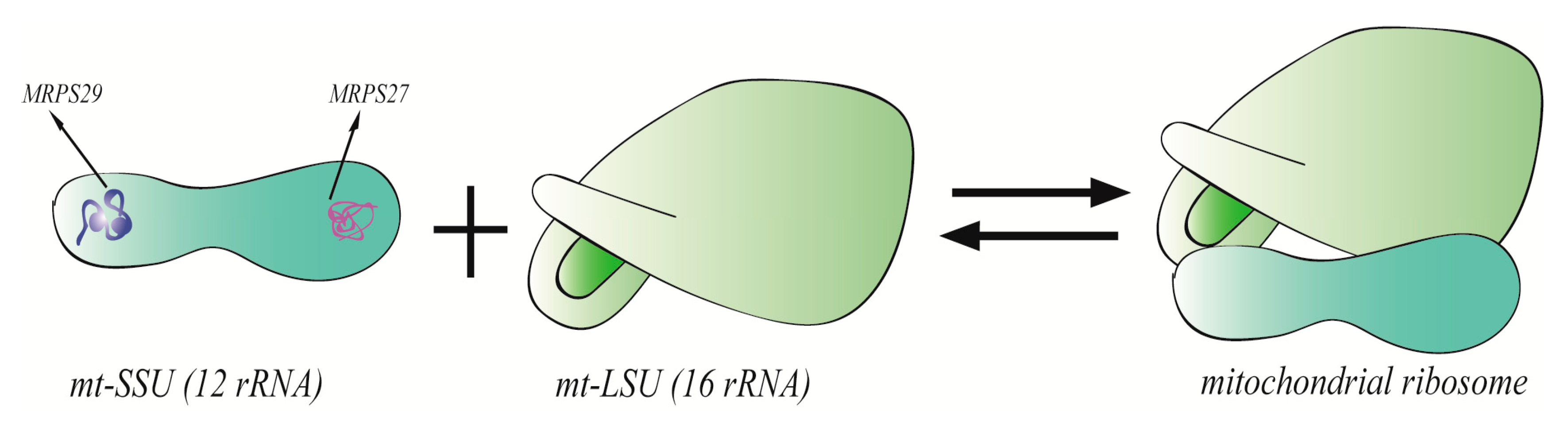
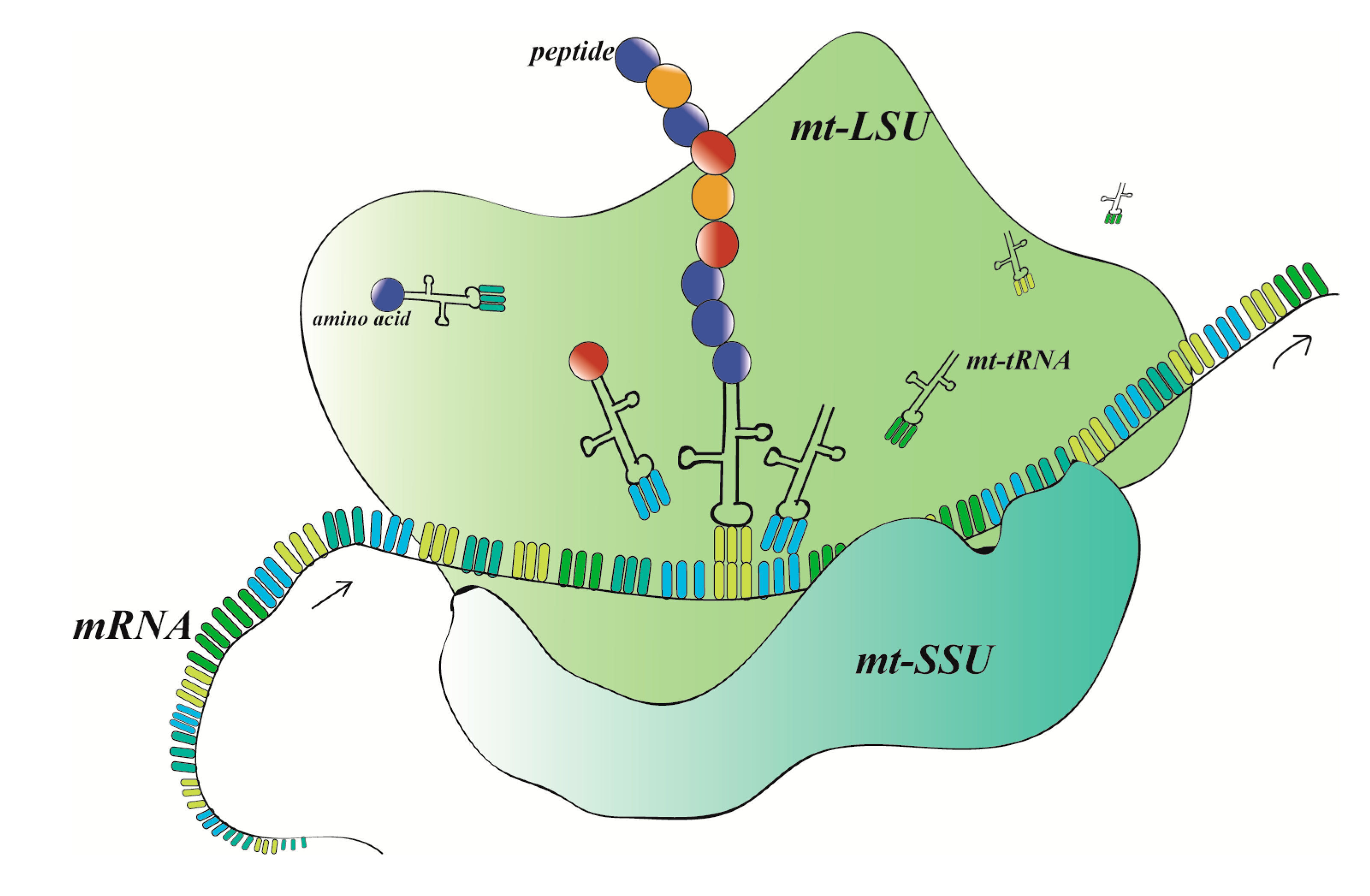
2. MRPs on Structure and Function of Mitochondrial Ribosomes
3. The Relationship between MRPs and Cell Apoptosis
3.1. MRPS29 (DAP3)
3.2. MRPL41 (BMRP)
3.3. MRPL65 (PDCD9, MRPS30)
4. MRPs Associated with Cancers
4.1. MRPs and Breast Cancer
4.2. MRPs and Digestive Tract Cancers
4.3. MRPs and Other Cancers
5. MRPs Associated with Diseases Except Cancers
5.1. MRPs Associated with Mitochondrial Diseases
5.2. Clinical Manifestations of MRPs Related Mitochondrial Disease
5.3. MRPs and Heart Diseases
5.4. MRPs Associated with Age and Other Related Mitochondrial Diseases
6. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ban, N.; Beckmann, R.; Cate, J.H.; Dinman, J.D.; Dragon, F.; Ellis, S.R.; Lafontaine, D.L.; Lindahl, L.; Liljas, A.; Lipton, J.M.; et al. A new system for naming ribosomal proteins. Curr. Opin. Struct. Biol. 2014, 24, 165–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki-Nevala, S.; Sarhadi, V.K.; Knuuttila, A.; Scheinin, I.; Ellonen, P.; Lagstrom, S.; Ronty, M.; Kettunen, E.; Husgafvel-Pursiainen, K.; Wolff, H.; et al. Driver Gene and Novel Mutations in Asbestos-Exposed Lung Adenocarcinoma and Malignant Mesothelioma Detected by Exome Sequencing. Lung 2016, 194, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Cahill, L.S.; Cameron, J.M.; Winterburn, J.; Macos, P.; Hoggarth, J.; Dzamba, M.; Brudno, M.; Nutter, L.M.J.; Sproule, T.J.; Burgess, R.W.; et al. Structural Variant in Mitochondrial-Associated Gene (MRPL3) Induces Adult-Onset Neurodegeneration with Memory Impairment in the Mouse. J. Neurosci. 2020, 40, 4576–4585. [Google Scholar] [CrossRef] [PubMed]
- Bursle, C.; Narendra, A.; Chuk, R.; Cardinal, J.; Justo, R.; Lewis, B.; Coman, D. COXPD9 an Evolving Multisystem Disease; Congenital Lactic Acidosis, Sensorineural Hearing Loss, Hypertrophic Cardiomyopathy, Cirrhosis and Interstitial Nephritis. JIMD Rep. 2017, 34, 105–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galmiche, L.; Serre, V.; Beinat, M.; Assouline, Z.; Lebre, A.S.; Chretien, D.; Nietschke, P.; Benes, V.; Boddaert, N.; Sidi, D.; et al. Exome sequencing identifies MRPL3 mutation in mitochondrial cardiomyopathy. Hum. Mutat. 2011, 32, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; He, Y.; Liu, Y.; Yang, X. Gene signature associated with neuro-endocrine activity predicting prognosis of pancreatic carcinoma. Mol. Genet. Genom. Med. 2019, 7, e00729. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; He, J.; Tang, J.; Chen, K.; Wang, Z.; Xia, Q.; Li, H. Investigation of the gene co-expression network and hub genes associated with acute mountain sickness. Hereditas 2020, 157. [Google Scholar] [CrossRef]
- Morais-Rodrigues, F.; Silύerio-Machado, R.; Kato, R.B.; Rodrigues, D.L.N.; Valdez-Baez, J.; Fonseca, V.; San, E.J.; Gomes, L.G.R.; dos Santos, R.G.; Vinicius Canário Viana, M.; et al. Analysis of the microarray gene expression for breast cancer progression after the application modified logistic regression. Gene 2020, 726. [Google Scholar] [CrossRef]
- Klein, R.; Li, X.; Kuo, J.Z.; Klein, B.E.; Cotch, M.F.; Wong, T.Y.; Taylor, K.D.; Rotter, J.I. Associations of candidate genes to age-related macular degeneration among racial/ethnic groups in the multi-ethnic study of atherosclerosis. Am. J. Ophthalmol. 2013, 156, 1010–1020.e1. [Google Scholar] [CrossRef] [Green Version]
- Besse, A.; Brezavar, D.; Hanson, J.; Larson, A.; Bonnen, P.E. LONP1 de novo dominant mutation causes mitochondrial encephalopathy with loss of LONP1 chaperone activity and excessive LONP1 proteolytic activity. Mitochondrion 2020, 51, 68–78. [Google Scholar] [CrossRef]
- Zhang, Q.; Liang, Z.; Gao, Y.; Teng, M.; Niu, L. Differentially expressed mitochondrial genes in breast cancer cells: Potential new targets for anti-cancer therapies. Gene 2017, 596, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Guo, M.; Wei, Y.; Yu, S.; Li, H.; Wang, Y.; Xu, X.; Cui, Y.; Tian, J.; Liang, L.; et al. FoxO3a confers cetuximab resistance in RAS wild-type metastatic colorectal cancer through c-Myc. Oncotarget 2016, 7, 80888–80900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-K.; Lim, J.J.; Jeoun, U.-w.; Min, S.; Lee, E.-b.; Kwon, S.M.; Lee, C.; Yoon, G. Lactate-mediated mitoribosomal defects impair mitochondrial oxidative phosphorylation and promote hepatoma cell invasiveness. J. Biol. Chem. 2017, 292, 20208–20217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Li, L.; Fu, L.; Yuan, Y.; Dai, H.; Zhu, T.; Zhou, Y.; Yuan, F. Integrated Bioinformatics Analysis the Function of RNA Binding Proteins (RBPs) and Their Prognostic Value in Breast Cancer. Front. Pharmacol. 2019, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Deng, J.L.; Wang, L.P.; Zhang, H.B.; Tang, L.; Huang, Y.; Tang, J.; Wang, S.M.; Wang, G. Identification of Candidate Genes Associated with Breast Cancer Prognosis. DNA Cell Biol. 2020, 39, 1205–1227. [Google Scholar] [CrossRef]
- Sotgia, F.; Fiorillo, M.; Lisanti, M.P. Mitochondrial markers predict recurrence, metastasis and tamoxifen-resistance in breast cancer patients: Early detection of treatment failure with companion diagnostics. Oncotarget 2017, 8, 68730–68745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.; Li, J.; Li, R.; Xu, X.; Liu, J.; Qin, L.; Huang, T.; Wu, J.; Jiao, M.; Wei, M.; et al. Potentially Critical Roles of NDUFB5, TIMMDC1, and VDAC3 in the Progression of Septic Cardiomyopathy Through Integrated Bioinformatics Analysis. DNA Cell Biol. 2020, 39, 105–117. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, Y.; Ling, J.; Xiang, Y.; Zhuo, W. Screening of key genes and prediction of therapeutic agents in Arsenic-induced lung carcinoma. Cancer Biomark. 2019, 25, 351–360. [Google Scholar] [CrossRef]
- Ayakannu, T.; Taylor, A.H.; Konje, J.C. Selection of Endogenous Control Reference Genes for Studies on Type 1 or Type 2 Endometrial Cancer. Sci. Rep. 2020, 10, 8468. [Google Scholar] [CrossRef]
- Ayakannu, T.; Taylor, A.H.; Willets, J.M.; Brown, L.; Lambert, D.G.; McDonald, J.; Davies, Q.; Moss, E.L.; Konje, J.C. Validation of endogenous control reference genes for normalizing gene expression studies in endometrial carcinoma. Mol. Hum. Reprod. 2015, 21, 723–735. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Zhao, J.; Ma, H.; Wang, M.C. Integrating transcriptome-wide association study and copy number variation study identifies candidate genes and pathways for diffuse non-Hodgkin‘s lymphoma. Cancer Genet 2020, 243, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.N.; Schmidt, H.; Steiniche, T.; Madsen, M. Identification of robust reference genes for studies of gene expression in FFPE melanoma samples and melanoma cell lines. Melanoma Res. 2020, 30, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Aasebø, E.; Berven, F.S.; Hovland, R.; Døskeland, S.O.; Bruserud, Ø.; Selheim, F.; Hernandez-Valladares, M. The Progression of Acute Myeloid Leukemia from First Diagnosis to Chemoresistant Relapse: A Comparison of Proteomic and Phosphoproteomic Profiles. Cancers 2020, 12, 1466. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Yu, Y.; Zhang, B.; Miao, L.; Wang, L.; Zhao, K.; Ji, Y.; Wang, R.; Ma, H.; Chen, N.; et al. Genetic variants in lncRNA H19 are associated with the risk of oral squamous cell carcinoma in a Chinese population. Oncotarget 2018, 9, 23915–23922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, N.Y.; Chueh, F.S.; Yu, C.C.; Liao, C.L.; Lin, J.J.; Hsia, T.C.; Wu, K.C.; Liu, H.C.; Lu, K.W.; Chung, J.G. Benzyl isothiocyanate alters the gene expression with cell cycle regulation and cell death in human brain glioblastoma GBM 8401 cells. Oncol. Rep. 2016, 35, 2089–2096. [Google Scholar] [CrossRef]
- Di Nottia, M.; Marchese, M.; Verrigni, D.; Mutti, C.D.; Torraco, A.; Oliva, R.; Fernandez-Vizarra, E.; Morani, F.; Trani, G.; Rizza, T.; et al. A homozygous MRPL24 mutation causes a complex movement disorder and affects the mitoribosome assembly. Neurobiol. Dis. 2020, 141. [Google Scholar] [CrossRef]
- Sotgia, F.; Lisanti, M.P. Mitochondrial biomarkers predict tumor progression and poor overall survival in gastric cancers: Companion diagnostics for personalized medicine. Oncotarget 2017, 8, 67117–67128. [Google Scholar] [CrossRef] [Green Version]
- Qiu, R.; Shi, H.; Wang, S.; Leng, S.; Liu, R.; Zheng, Y.; Huang, W.; Zeng, Y.; Gao, J.; Zhang, K.; et al. BRMS1 coordinates with LSD1 and suppresses breast cancer cell metastasis. Am. J. Cancer Res. 2018, 8, 2030–2045. [Google Scholar]
- Liu, L.; Luo, C.; Luo, Y.; Chen, L.; Liu, Y.; Wang, Y.; Han, J.; Zhang, Y.; Wei, N.; Xie, Z.; et al. MRPL33 and its splicing regulator hnRNPK are required for mitochondria function and implicated in tumor progression. Oncogene 2018, 37, 86–94. [Google Scholar] [CrossRef]
- Li, J.; Feng, D.; Gao, C.; Zhang, Y.; Xu, J.; Wu, M.; Zhan, X. Isoforms S and L of MRPL33 from alternative splicing have isoform-specific roles in the chemoresponse to epirubicin in gastric cancer cells via the PI3K/AKT signaling pathway. Int. J. Oncol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Lebedev, T.D.; Vagapova, E.R.; Popenko, V.I.; Leonova, O.G.; Spirin, P.V.; Prassolov, V.S. Two Receptors, Two Isoforms, Two Cancers: Comprehensive Analysis of KIT and TrkA Expression in Neuroblastoma and Acute Myeloid Leukemia. Front. Oncol. 2019, 9, 1046. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Zambo, K.D.A.; Zamuner, F.T.; Ou, T.; Hopkins, C.; Kelley, D.Z.; Wulf, H.A.; Winkler, E.; Erbe, R.; Danilova, L.; et al. Chromatin structure regulates cancer-specific alternative splicing events in primary HPV-related oropharyngeal squamous cell carcinoma. Epigenetics 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Chen, C.; Cao, H.; Wang, J.; Shen, E. Whole transcriptome sequencing reveals biologically significant RNA markers and related regulating biological pathways in cardiomyocyte hypertrophy induced by high glucose. J. Cell. Biochem. 2019, 120, 1018–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshabi, A.M.; Vastrad, B.; Shaikh, I.A.; Vastrad, C. Identification of Crucial Candidate Genes and Pathways in Glioblastoma Multiform by Bioinformatics Analysis. Biomolecules 2019, 9, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Lu, P.; Yan, L.; Yang, L.; Wang, Y.; Chen, J.; Dai, J.; Li, Y.; Kang, Z.; Bai, T.; et al. MRPL35 Is Up-Regulated in Colorectal Cancer and Regulates Colorectal Cancer Cell Growth and Apoptosis. Am. J. Pathol. 2019, 189, 1105–1120. [Google Scholar] [CrossRef] [Green Version]
- Corrêa, T.; Feltes, B.C.; Riegel, M. Integrated analysis of the critical region 5p15.3–p15.2 associated with cri-du-chat syndrome. Genet. Mol. Biol. 2019, 42 (Suppl. 1), 186–196. [Google Scholar] [CrossRef]
- Sundquist, K.; Ahmad, A.; Svensson, P.J.; Zoller, B.; Sundquist, J.; Memon, A.A. Polymorphisms in PARK2 and MRPL37 are associated with higher risk of recurrent venous thromboembolism in a sex-specific manner. J. Thromb. Thrombolysis 2018, 46, 154–165. [Google Scholar] [CrossRef] [Green Version]
- Sultana, N.; Rahman, M.; Myti, S.; Islam, J.; Mustafa, M.G.; Nag, K. A novel knowledge-derived data potentizing method revealed unique liver cancer-associated genetic variants. Hum. Genom. 2019, 13. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.J.; Zhao, N.; Shen, H.; Wang, H. Long Noncoding RNA MRPL39 Inhibits Gastric Cancer Proliferation and Progression by Directly Targeting miR-130. Genet. Test. Mol. Biomark. 2018, 22, 656–663. [Google Scholar] [CrossRef]
- Napoli, E.; Tassone, F.; Wong, S.; Angkustsiri, K.; Simon, T.J.; Song, G.; Giulivi, C. Mitochondrial Citrate Transporter-dependent Metabolic Signature in the 22q11.2 Deletion Syndrome. J. Biol. Chem. 2015, 290, 23240–23253. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ryan, S.K.; Deboer, E.; Cook, K.; Fitzgerald, S.; Lachman, H.M.; Wallace, D.C.; Goldberg, E.M.; Anderson, S.A. Mitochondrial deficits in human iPSC-derived neurons from patients with 22q11.2 deletion syndrome and schizophrenia. Transl. Psychiatry 2019, 9, 302. [Google Scholar] [CrossRef] [Green Version]
- Devaraju, P.; Yu, J.; Eddins, D.; Mellado-Lagarde, M.M.; Earls, L.R.; Westmoreland, J.J.; Quarato, G.; Green, D.R.; Zakharenko, S.S. Haploinsufficiency of the 22q11.2 microdeletion gene Mrpl40 disrupts short-term synaptic plasticity and working memory through dysregulation of mitochondrial calcium. Mol. Psychiatry 2016, 22, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.; Duan, H.; Li, H.; Wang, H.; Liu, Y.; Fan, Y.; Zhang, C. Knockdown of MRPL42 suppresses glioma cell proliferation by inducing cell cycle arrest and apoptosis. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Yuan, W.; Shen, Y.; Lu, X.; Li, Y.; Tian, T.; Jiang, L.; Zhuang, X.; Wu, J.; Chu, M. The miR-608 rs4919510 polymorphism may modify cancer susceptibility based on type. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilescu, C.; Ojala, T.H.; Brilhante, V.; Ojanen, S.; Hinterding, H.M.; Palin, E.; Alastalo, T.P.; Koskenvuo, J.; Hiippala, A.; Jokinen, E.; et al. Genetic Basis of Severe Childhood-Onset Cardiomyopathies. J. Am. Coll. Cardiol. 2018, 72, 2324–2338. [Google Scholar] [CrossRef] [PubMed]
- Yeo, J.H.; Skinner, J.P.; Bird, M.J.; Formosa, L.E.; Zhang, J.G.; Kluck, R.M.; Belz, G.T.; Chong, M.M. A Role for the Mitochondrial Protein Mrpl44 in Maintaining OXPHOS Capacity. PLoS ONE 2015, 10, e0134326. [Google Scholar] [CrossRef]
- Distelmaier, F.; Haack, T.B.; Catarino, C.B.; Gallenmüller, C.; Rodenburg, R.J.; Strom, T.M.; Baertling, F.; Meitinger, T.; Mayatepek, E.; Prokisch, H.; et al. MRPL44 mutations cause a slowly progressive multisystem disease with childhood-onset hypertrophic cardiomyopathy. Neurogenetics 2015, 16, 319–323. [Google Scholar] [CrossRef]
- Morin, A.; Madore, A.M.; Kwan, T.; Ban, M.; Partanen, J.; Rönnblom, L.; Syvänen, A.C.; Sawcer, S.; Stunnenberg, H.; Lathrop, M.; et al. Exploring rare and low-frequency variants in the Saguenay-Lac-Saint-Jean population identified genes associated with asthma and allergy traits. Eur. J. Hum. Genet. EJHG 2019, 27, 90–101. [Google Scholar] [CrossRef]
- Antony, F.; Deantonio, C.; Cotella, D.; Soluri, M.F.; Tarasiuk, O.; Raspagliesi, F.; Adorni, F.; Piazza, S.; Ciani, Y.; Santoro, C.; et al. High-throughput assessment of the antibody profile in ovarian cancer ascitic fluids. OncoImmunology 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Abaji, R.; Ceppi, F.; Patel, S.; Gagné, V.; Xu, C.J.; Spinella, J.F.; Colombini, A.; Parasole, R.; Buldini, B.; Basso, G.; et al. Genetic risk factors for VIPN in childhood acute lymphoblastic leukemia patients identified using whole-exome sequencing. Pharmacogenomics 2018, 19, 1181–1193. [Google Scholar] [CrossRef]
- Maiuthed, A.; Prakhongcheep, O.; Chanvorachote, P. Microarray-based Analysis of Genes, Transcription Factors, and Epigenetic Modifications in Lung Cancer Exposed to Nitric Oxide. Cancer Genom. Proteom. 2020, 17, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Abdul Aziz, N.A.; Mokhtar, N.M.; Harun, R.; Mollah, M.M.; Mohamed Rose, I.; Sagap, I.; Mohd Tamil, A.; Wan Ngah, W.Z.; Jamal, R. A 19-Gene expression signature as a predictor of survival in colorectal cancer. BMC Med. Genom. 2016, 9, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Cortés, A.; Cabrera-Andrade, A.; Vázquez-Naya, J.M.; Pazos, A.; Gonzáles-Díaz, H.; Paz, Y.M.C.; Guerrero, S.; Pérez-Castillo, Y.; Tejera, E.; Munteanu, C.R. Prediction of breast cancer proteins involved in immunotherapy, metastasis, and RNA-binding using molecular descriptors and artificial neural networks. Sci. Rep. 2020, 10, 8515. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, R.; Lu, D.; Zhuo, J.; Zhang, X.; Wang, K.; Wei, X.; Wei, Q.; Wang, W.; Xie, H.; Zhou, L.; et al. CR6-interacting factor 1 inhibits invasiveness by suppressing TGF-β-mediated epithelial-mesenchymal transition in hepatocellular carcinoma. Oncotarget 2017, 8, 94759–94768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahedi, S.; Chueh, F.Y.; Chandran, B.; Yu, C.L. Lymphocyte-specific protein tyrosine kinase (Lck) interacts with CR6-interacting factor 1 (CRIF1) in mitochondria to repress oxidative phosphorylation. BMC Cancer 2015, 15, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Ran, Q.; Xiang, Y.; Xiang, L.; Chen, L.; Li, F.; Wu, J.; Wu, C.; Li, Z. Co-Activation of PKC-δ by CRIF1 Modulates Oxidative Stress in Bone Marrow Multipotent Mesenchymal Stromal Cells after Irradiation by Phosphorylating NRF2 Ser40. Theranostics 2017, 7, 2634–2648. [Google Scholar] [CrossRef] [PubMed]
- Ramchandran, R.; Piao, S.; Lee, J.W.; Nagar, H.; Jung, S.-b.; Choi, S.; Kim, S.; Lee, I.; Kim, S.-m.; Shin, N.; et al. CR6 interacting factor 1 deficiency promotes endothelial inflammation by SIRT1 downregulation. PLoS ONE 2018, 13, e192693. [Google Scholar] [CrossRef] [Green Version]
- Nagar, H.; Jung, S.B.; Ryu, M.J.; Choi, S.J.; Piao, S.; Song, H.J.; Kang, S.K.; Shin, N.; Kim, D.W.; Jin, S.A.; et al. CR6-Interacting Factor 1 Deficiency Impairs Vascular Function by Inhibiting the Sirt1-Endothelial Nitric Oxide Synthase Pathway. Antioxid. Redox Signal. 2017, 27, 234–249. [Google Scholar] [CrossRef]
- Park, J.S.; Choi, S.Y.; Hwang, S.H.; Kim, S.M.; Choi, J.; Jung, K.A.; Kwon, J.Y.; Kong, Y.Y.; Cho, M.L.; Park, S.H. CR6-interacting factor 1 controls autoimmune arthritis by regulation of signal transducer and activator of transcription 3 pathway and T helper type 17 cells. Immunology 2019, 156, 413–421. [Google Scholar] [CrossRef]
- Zhou, C.; Chen, Z.; Peng, C.; Chen, C.; Li, H. Long Noncoding RNA TRIM52-AS1 Sponges miR-514a-5p to Facilitate Hepatocellular Carcinoma Progression Through Increasing MRPS18A. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef]
- Tian, A.; Pu, K.; Li, B.; Li, M.; Liu, X.; Gao, L.; Mao, X. Weighted gene coexpression network analysis reveals hub genes involved in cholangiocarcinoma progression and prognosis. Hepatol. Res. 2019, 49, 1195–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbergenov, R.; Duscha, S.; Fritz, A.K.; Juskeviciene, R.; Oishi, N.; Schmitt, K.; Shcherbakov, D.; Teo, Y.; Boukari, H.; Freihofer, P.; et al. Mutant MRPS5 affects mitoribosomal accuracy and confers stress-related behavioral alterations. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.; Xie, X.; Wang, Z.; Lei, Q. Identification of potential crucial genes and key pathways in osteosarcoma. Hereditas 2020, 157. [Google Scholar] [CrossRef] [PubMed]
- Menezes, M.J.; Guo, Y.; Zhang, J.; Riley, L.G.; Cooper, S.T.; Thorburn, D.R.; Li, J.; Dong, D.; Li, Z.; Glessner, J.; et al. Mutation in mitochondrial ribosomal protein S7 (MRPS7) causes congenital sensorineural deafness, progressive hepatic and renal failure and lactic acidemia. Hum. Mol. Genet. 2015, 24, 2297–2307. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.D.; Zhao, X.; Li, W.T. Identification of differentially expressed metastatic genes and their signatures to predict the overall survival of uveal melanoma patients by bioinformatics analysis. Int. J. Ophthalmol. 2020, 13, 1046–1053. [Google Scholar] [CrossRef]
- Jackson, C.B.; Huemer, M.; Bolognini, R.; Martin, F.; Szinnai, G.; Donner, B.C.; Richter, U.; Battersby, B.J.; Nuoffer, J.M.; Suomalainen, A.; et al. A variant in MRPS14 (uS14m) causes perinatal hypertrophic cardiomyopathy with neonatal lactic acidosis, growth retardation, dysmorphic features and neurological involvement. Hum. Mol. Genet. 2019, 28, 639–649. [Google Scholar] [CrossRef]
- Mushtaq, M.; Jensen, L.; Davidsson, S.; Grygoruk, O.V.; Andrén, O.; Kashuba, V.; Kashuba, E. The MRPS18-2 protein levels correlate with prostate tumor progression and it induces CXCR4-dependent migration of cancer cells. Sci. Rep. 2018, 8, 2268. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, M.; Ali, R.H.; Kashuba, V.; Klein, G.; Kashuba, E. S18 family of mitochondrial ribosomal proteins: Evolutionary history and Gly132 polymorphism in colon carcinoma. Oncotarget 2016, 7, 55649–55662. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Hsiao, C.C.; Chen, T.W.; Wu, C.C.; Chao, T.Y.; Leung, S.Y.; Eng, H.L.; Lee, C.P.; Wang, T.Y.; Lin, M.C. Whole Genome DNA Methylation Analysis of Active Pulmonary Tuberculosis Disease Identifies Novel Epigenotypes: PARP9/miR-505/RASGRP4/GNG12 Gene Methylation and Clinical Phenotypes. Int. J. Mol. Sci. 2020, 21, 3180. [Google Scholar] [CrossRef]
- Hamdi, Y.; Soucy, P.; Adoue, V.; Michailidou, K.; Canisius, S.; Lemaçon, A.; Droit, A.; Andrulis, I.L.; Anton-Culver, H.; Arndt, V.; et al. Association of breast cancer risk with genetic variants showing differential allelic expression: Identification of a novel breast cancer susceptibility locus at 4q21. Oncotarget 2016, 7, 80140–80163. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-García, M.E.; Cotrina-Vinagre, F.J.; Carnicero-Rodríguez, P.; Martínez-Azorín, F. An innovative strategy to clone positive modifier genes of defects caused by mtDNA mutations: MRPS18C as suppressor gene of m.3946G > A mutation in MT-ND1 gene. Hum. Genet. 2017, 136, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Bertini, V.; Valetto, A.; Baldinotti, F.; Azzara, A.; Cambi, F.; Toschi, B.; Giacomina, A.; Gatti, G.L.; Gana, S.; Caligo, M.A.; et al. Blepharophimosis, Ptosis, Epicanthus Inversus Syndrome: New Report with a 197-kb Deletion Upstream of FOXL2 and Review of the Literature. Mol. Syndromol. 2019, 10, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Baertling, F.; Haack, T.B.; Rodenburg, R.J.; Schaper, J.; Seibt, A.; Strom, T.M.; Meitinger, T.; Mayatepek, E.; Hadzik, B.; Selcan, G.; et al. MRPS22 mutation causes fatal neonatal lactic acidosis with brain and heart abnormalities. Neurogenetics 2015, 16, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Tiosano, D.; Guran, T.; Baris, H.N.; Bayram, Y.; Mory, A.; Shapiro-Kulnane, L.; Hodges, C.A.; Akdemir, Z.C.; Turan, S.; et al. Mutations in the mitochondrial ribosomal protein MRPS22 lead to primary ovarian insufficiency. Hum. Mol. Genet. 2018, 27, 1913–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franca, M.M.; Mendonca, B.B. Genetics of Primary Ovarian Insufficiency in the Next-Generation Sequencing Era. J. Endocr. Soc. 2020, 4, bvz037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiosano, D.; Mears, J.A.; Buchner, D.A. Mitochondrial Dysfunction in Primary Ovarian Insufficiency. Endocrinology 2019, 160, 2353–2366. [Google Scholar] [CrossRef]
- Pu, M.; Wang, J.; Huang, Q.; Zhao, G.; Xia, C.; Shang, R.; Zhang, Z.; Bian, Z.; Yang, X.; Tao, K. High MRPS23 expression contributes to hepatocellular carcinoma proliferation and indicates poor survival outcomes. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Borna, N.N.; Kishita, Y.; Kohda, M.; Lim, S.C.; Shimura, M.; Wu, Y.; Mogushi, K.; Yatsuka, Y.; Harashima, H.; Hisatomi, Y.; et al. Mitochondrial ribosomal protein PTCD3 mutations cause oxidative phosphorylation defects with Leigh syndrome. Neurogenetics 2019, 20, 9–25. [Google Scholar] [CrossRef]
- Amunts, A.; Brown, A.; Toots, J.; Scheres, S.H.W.; Ramakrishnan, V. Ribosome. The structure of the human mitochondrial ribosome. Science 2015, 348, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Yusupov, M.M.; Yusupova, G.Z.; Baucom, A.; Lieberman, K.; Earnest, T.N.; Cate, J.H.; Noller, H.F. Crystal structure of the ribosome at 5.5 A resolution. Science 2001, 292, 883–896. [Google Scholar] [CrossRef]
- Gray, M.W.; Burger, G.; Lang, B.F. The origin and early evolution of mitochondria. Genome Biol. 2001, 2, Reviews1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.R.; Koc, E.C.; Datta, P.P.; Booth, T.M.; Spremulli, L.L.; Agrawal, R.K. Structure of the mammalian mitochondrial ribosome reveals an expanded functional role for its component proteins. Cell 2003, 115, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, P.S.; Sharma, M.R.; Booth, T.M.; Haque, E.M.; Tung, C.S.; Sanbonmatsu, K.Y.; Spremulli, L.L.; Agrawal, R.K. Cryo-EM structure of the small subunit of the mammalian mitochondrial ribosome. Proc. Natl. Acad. Sci. USA 2014, 111, 7284–7289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greber, B.J.; Boehringer, D.; Leibundgut, M.; Bieri, P.; Leitner, A.; Schmitz, N.; Aebersold, R.; Ban, N. The complete structure of the large subunit of the mammalian mitochondrial ribosome. Nature 2014, 515, 283–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greber, B.J.; Bieri, P.; Leibundgut, M.; Leitner, A.; Aebersold, R.; Boehringer, D.; Ban, N. Ribosome. The complete structure of the 55S mammalian mitochondrial ribosome. Science 2015, 348, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.; Amunts, A.; Bai, X.C.; Sugimoto, Y.; Edwards, P.C.; Murshudov, G.; Scheres, S.H.W.; Ramakrishnan, V. Structure of the large ribosomal subunit from human mitochondria. Science 2014, 346, 718–722. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.M.; Rackham, O.; Shearwood, A.M.; Hamilton, K.L.; Narsai, R.; Whelan, J.; Filipovska, A. Pentatricopeptide repeat domain protein 3 associates with the mitochondrial small ribosomal subunit and regulates translation. FEBS Lett. 2009, 583, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Rabl, J.; Leibundgut, M.; Ataide, S.F.; Haag, A.; Ban, N. Crystal structure of the eukaryotic 40S ribosomal subunit in complex with initiation factor 1. Science 2011, 331, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Chae, H.J.; Thomas, M.; Miyazaki, T.; Monosov, A.; Monosov, E.; Krajewska, M.; Krajewski, S.; Reed, J.C. Mammalian dap3 is an essential gene required for mitochondrial homeostasis in vivo and contributing to the extrinsic pathway for apoptosis. FASEB J. 2007, 21, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.M.; Anselmi, C.; Wittig, I.; Faraldo-Gomez, J.D.; Kuhlbrandt, W. Structure of the yeast F1Fo-ATP synthase dimer and its role in shaping the mitochondrial cristae. Proc. Natl. Acad. Sci. USA 2012, 109, 13602–13607. [Google Scholar] [CrossRef] [Green Version]
- Gopisetty, G.; Thangarajan, R. Mammalian mitochondrial ribosomal small subunit (MRPS) genes: A putative role in human disease. Gene 2016, 589, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Cavdar Koc, E.; Ranasinghe, A.; Burkhart, W.; Blackburn, K.; Koc, H.; Moseley, A.; Spremulli, L.L. A new face on apoptosis: Death-associated protein 3 and PDCD9 are mitochondrial ribosomal proteins. FEBS Lett. 2001, 492, 166–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilkerson, R.W.; Margineantu, D.H.; Capaldi, R.A.; Selker, J.M. Mitochondrial DNA depletion causes morphological changes in the mitochondrial reticulum of cultured human cells. FEBS Lett. 2000, 474, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Shen, M.; Fujikura, D.; Tosa, N.; Kim, H.R.; Kon, S.; Uede, T.; Reed, J.C. Functional role of death-associated protein 3 (DAP3) in anoikis. J. Biol. Chem. 2004, 279, 44667–44672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, O.; Inbal, B.; Kissil, J.L.; Raveh, T.; Berissi, H.; Spivak-Kroizaman, T.; Feinstein, E.; Kimchi, A. DAP-kinase participates in TNF-alpha- and Fas-induced apoptosis and its function requires the death domain. J. Cell Biol. 1999, 146, 141–148. [Google Scholar] [CrossRef]
- Mukamel, Z.; Kimchi, A. Death-associated protein 3 localizes to the mitochondria and is involved in the process of mitochondrial fragmentation during cell death. J. Biol. Chem. 2004, 279, 36732–36738. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Reed, J.C. A GTP-binding adapter protein couples TRAIL receptors to apoptosis-inducing proteins. Nat. Immunol. 2001, 2, 493–500. [Google Scholar] [CrossRef]
- Berger, T.; Kretzler, M. Interaction of DAP3 and FADD only after cellular disruption. Nat. Immunol. 2002, 3, 3–5. [Google Scholar] [CrossRef]
- Wazir, U.; Orakzai, M.M.; Khanzada, Z.S.; Jiang, W.G.; Sharma, A.K.; Kasem, A.; Mokbel, K. The role of death-associated protein 3 in apoptosis, anoikis and human cancer. Cancer Cell Int. 2015, 15, 39. [Google Scholar] [CrossRef] [Green Version]
- Chintharlapalli, S.R.; Jasti, M.; Malladi, S.; Parsa, K.V.; Ballestero, R.P.; González-García, M. BMRP is a Bcl-2 binding protein that induces apoptosis. J. Cell. Biochem. 2005, 94, 611–626. [Google Scholar] [CrossRef]
- Malladi, S.; Parsa, K.V.; Bhupathi, D.; Rodríguez-González, M.A.; Conde, J.A.; Anumula, P.; Romo, H.E.; Claunch, C.J.; Ballestero, R.P.; González-García, M. Deletion mutational analysis of BMRP, a pro-apoptotic protein that binds to Bcl-2. Mol. Cell. Biochem. 2011, 351, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.A.; Kim, M.J.; Park, J.K.; Chung, Y.M.; Lee, J.H.; Chi, S.G.; Kim, J.S.; Yoo, Y.D. Mitochondrial ribosomal protein L41 suppresses cell growth in association with p53 and p27Kip1. Mol. Cell. Biochem. 2005, 25, 6603–6616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Yoo, Y.A.; Kim, H.J.; Kang, S.; Kim, Y.G.; Kim, J.S.; Yoo, Y.D. Mitochondrial ribosomal protein L41 mediates serum starvation-induced cell-cycle arrest through an increase of p21(WAF1/CIP1). Biochem. Biophys. Res. Commun. 2005, 338, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, Y.; Frémont, M.; Schwarz, S.; Siegmann, M.; Matthies, R.; Jost, J.P. A novel 52 kDa protein induces apoptosis and concurrently activates c-Jun N-terminal kinase 1 (JNK1) in mouse C3H10T1/2 fibroblasts. Gene 1998, 208, 157–166. [Google Scholar] [CrossRef]
- Andrawus, M.; Sharvit, L.; Shekhidem, H.A.; Roichman, A.; Cohen, H.Y.; Atzmon, G. The effects of environmental stressors on candidate aging associated genes. Exp. Gerontol. 2020, 137, 110952. [Google Scholar] [CrossRef] [PubMed]
- Ghoussaini, M.; French, J.D.; Michailidou, K.; Nord, S.; Beesley, J.; Canisus, S.; Hillman, K.M.; Kaufmann, S.; Sivakumaran, H.; Moradi Marjaneh, M.; et al. Evidence that the 5p12 Variant rs10941679 Confers Susceptibility to Estrogen-Receptor-Positive Breast Cancer through FGF10 and MRPS30 Regulation. Am. J. Hum. Genet. 2016, 99, 903–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Manjunath, M.; Zhang, S.; Chasman, D.; Roy, S.; Song, J.S. Integrative Genomic Analysis Predicts Causative Cis-Regulatory Mechanisms of the Breast Cancer-Associated Genetic Variant rs4415084. Cancer Res. 2018, 78, 1579–1591. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Lin, W.; Bao, J.; Cai, Q.; Pan, X.; Bai, M.; Yuan, Y.; Shi, J.; Sun, Y.; Han, M.R.; et al. A Comprehensive cis-eQTL Analysis Revealed Target Genes in Breast Cancer Susceptibility Loci Identified in Genome-wide Association Studies. Am. J. Hum. Genet. 2018, 102, 890–903. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Pan, Y.; Liu, G.; Yang, T.; Jin, Y.; Zhou, F.; Wei, Y. MRPS30-DT Knockdown Inhibits Breast Cancer Progression by Targeting Jab1/Cops5. Front. Oncol. 2019, 9, 1170. [Google Scholar] [CrossRef]
- Chen, C.W.; Fu, M.; Du, Z.H.; Zhao, F.; Yang, W.W.; Xu, L.H.; Li, S.L.; Ge, X.Y. Long Noncoding RNA MRPL23-AS1 Promoteoid Cystic Carcinoma Lung Metastasis. Cancer Res. 2020, 80, 2273–2285. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.X.; Zhang, Y.J.; Zhang, Y.; Ren, X.; Shen, Y.F.; Cheng, M.B.; Zhang, Y. CRIF1 enhances p53 activity via the chromatin remodeler SNF5 in the HCT116 colon cancer cell lines. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Li, H.; Wei, J.; Luo, Y.; Liu, H.; Zhang, J.; Luo, X. Risk Scoring System based on lncRNA Expression for Predicting Survival in Hepatocellular Carcinoma with Cirrhosis. Asian Pac. J. Cancer Prev. 2020, 21, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, J.; Long, X.; Jiao, L.; Zhou, M.; Wu, K. MRPS16 facilitates tumor progression via the PI3K/AKT/Snail signaling axis. J. Cancer 2020, 11, 2032–2043. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.Y.; Wang, W.; Luo, X.G.; Jiang, Y.F.; He, X.; Xu, P.; Chen, X.; Li, X.Y. Screening of prognostic risk microRNAs for acute myeloid leukemia. Hematology 2018, 23, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Li, H.B.; Wang, R.X.; Jiang, H.B.; Zhang, E.D.; Tan, J.Q.; Xu, H.Z.; Zhou, R.R.; Xia, X.B. Mitochondrial Ribosomal Protein L10 Associates with Cyclin B1/Cdk1 Activity and Mitochondrial Function. DNA Cell Biol. 2016, 35, 680–690. [Google Scholar] [CrossRef] [Green Version]
- Barsh, G.S.; Kohda, M.; Tokuzawa, Y.; Kishita, Y.; Nyuzuki, H.; Moriyama, Y.; Mizuno, Y.; Hirata, T.; Yatsuka, Y.; Yamashita-Sugahara, Y.; et al. A Comprehensive Genomic Analysis Reveals the Genetic Landscape of Mitochondrial Respiratory Chain Complex Deficiencies. PLoS Genet. 2016, 12. [Google Scholar] [CrossRef]
- Lake, N.J.; Webb, B.D.; Stroud, D.A.; Richman, T.R.; Ruzzenente, B.; Compton, A.G.; Mountford, H.S.; Pulman, J.; Zangarelli, C.; Rio, M.; et al. Biallelic Mutations in MRPS34 Lead to Instability of the Small Mitoribosomal Subunit and Leigh Syndrome. Am. J. Hum. Genet. 2017, 101, 239–254. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.M.; Choi, D.K.; Sohn, K.C.; Kim, J.Y.; Im, M.; Lee, Y.; Seo, Y.J.; Shong, M.; Lee, J.H.; Kim, C.D. Targeted deletion of Crif1 in mouse epidermis impairs skin homeostasis and hair morphogenesis. Sci. Rep. 2017, 7, 44828. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.K.; Ryu, D.; Kim, K.S.; Chang, J.Y.; Kim, Y.K.; Yi, H.S.; Kang, S.G.; Choi, M.J.; Lee, S.E.; Jung, S.B.; et al. Growth differentiation factor 15 is a myomitokine governing systemic energy homeostasis. J. Cell Biol. 2017, 216, 149–165. [Google Scholar] [CrossRef]
- Jung, S.-B.; Choi, M.J.; Ryu, D.; Yi, H.-S.; Lee, S.E.; Chang, J.Y.; Chung, H.K.; Kim, Y.K.; Kang, S.G.; Lee, J.H.; et al. Reduced oxidative capacity in macrophages results in systemic insulin resistance. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.A.; Kartal, F.; Koc, Z.C.; Murphy, T.; Kim, J.H.; Denvir, J.; Koc, E.C. Mitochondrial oxidative phosphorylation is impaired in TALLYHO mice, a new obesity and type 2 diabetes animal model. Int. J. Biochem. Cell Biol. 2019, 116, 105616. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Wei, D.; Dai, B.; Zheng, L.; Zhao, M.; Xin, N.; Chi, Z.; Zhao, Y.; Ma, T.; Jahane, R.; et al. Mitochondrial Genome Encoded Proteins Expression Disorder, the Possible Mechanism of the Heart Disease in Metabolic Syndrome. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 959–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardeitchik, T.; Mohamed, M.; Ruzzenente, B.; Karall, D.; Guerrero-Castillo, S.; Dalloyaux, D.; van den Brand, M.; van Kraaij, S.; van Asbeck, E.; Assouline, Z.; et al. Bi-allelic Mutations in the Mitochondrial Ribosomal Protein MRPS2 Cause Sensorineural Hearing Loss, Hypoglycemia, and Multiple OXPHOS Complex Deficiencies. Am. J. Hum. Genet. 2018, 102, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulman, J.; Ruzzenente, B.; Bianchi, L.; Rio, M.; Boddaert, N.; Munnich, A.; Rötig, A.; Metodiev, M.D. Mutations in the MRPS28 gene encoding the small mitoribosomal subunit protein bS1m in a patient with intrauterine growth retardation, craniofacial dysmorphism and multisystemic involvement. Hum. Mol. Genet. 2019, 28, 1445–1462. [Google Scholar] [CrossRef]
- Hu, Y.; Deng, L.; Zhang, J.; Fang, X.; Mei, P.; Cao, X.; Lin, J.; Wei, Y.; Zhang, X.; Xu, R. A Pooling Genome-Wide Association Study Combining a Pathway Analysis for Typical Sporadic Parkinson’s Disease in the Han Population of Chinese Mainland. Mol. Neurobiol. 2016, 53, 4302–4318. [Google Scholar] [CrossRef]
- Zhao, Q.; Sun, H.; Yin, L.; Wang, L. miR-126a-5p-Dbp and miR-31a-Crot/Mrpl4 interaction pairs crucial for the development of hypertension and stroke. Mol. Med. Rep. 2019, 20, 4151–4167. [Google Scholar] [CrossRef] [Green Version]
- Bugiardini, E.; Mitchell, A.L.; Rosa, I.D.; Horning-Do, H.T.; Pitmann, A.M.; Poole, O.V.; Holton, J.L.; Shah, S.; Woodward, C.; Hargreaves, I.; et al. MRPS25 mutations impair mitochondrial translation and cause encephalomyopathy. Hum. Mol. Genet. 2019, 28, 2711–2719. [Google Scholar] [CrossRef] [Green Version]
- Carroll, C.J.; Isohanni, P.; Poyhonen, R.; Euro, L.; Richter, U.; Brilhante, V.; Gotz, A.; Lahtinen, T.; Paetau, A.; Pihko, H.; et al. Whole-exome sequencing identifies a mutation in the mitochondrial ribosome protein MRPL44 to underlie mitochondrial infantile cardiomyopathy. J. Med Genet. 2013, 50, 151–159. [Google Scholar] [CrossRef]
- Kılıç, M.; Oğuz, K.K.; Kılıç, E.; Yüksel, D.; Demirci, H.; Sağıroğlu, M.; Yücel-Yılmaz, D.; Özgül, R.K. A patient with mitochondrial disorder due to a novel mutation in MRPS22. Metab. Brain Dis. 2017, 32, 1389–1393. [Google Scholar] [CrossRef]
- Yazdani, A.; Yazdani, A.; Méndez Giráldez, R.; Aguilar, D.; Sartore, L. A Multi-Trait Approach Identified Genetic Variants Including a Rare Mutation in RGS3 with Impact on Abnormalities of Cardiac Structure/Function. Sci. Rep. 2019, 9, 5845. [Google Scholar] [CrossRef]
- Mirzaei, M.; Pushpitha, K.; Deng, L.; Chitranshi, N.; Gupta, V.; Rajput, R.; Mangani, A.B.; Dheer, Y.; Godinez, A.; McKay, M.J.; et al. Upregulation of Proteolytic Pathways and Altered Protein Biosynthesis Underlie Retinal Pathology in a Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 6017–6034. [Google Scholar] [CrossRef] [PubMed]
- Mozhui, K.; Snively, B.M.; Rapp, S.R.; Wallace, R.B.; Williams, R.W.; Johnson, K.C. Genetic Analysis of Mitochondrial Ribosomal Proteins and Cognitive Aging in Postmenopausal Women. Front. Genet. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, Q.; Xiang, Y.; Stephen, P.; Wu, C.; Li, T.; Lin, S.X.; Li, Z. CRIF1-CDK2 Interface Inhibitors: An Unprecedented Strategy for Modulation of Cell Radiosensitivity. J. Am. Chem. Soc. 2019, 141, 1420–1424. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Old Name | New Name | Cancer | Other Diseases |
|---|---|---|---|
| Mitochondrial ribosomal large subunit (mt-LSU) genes and proteins. | |||
| MRPL1 | uL1m | Lung cancer [2] | |
| MRPL3 | uL3m | Neurodegeneration and memory impairment [3], Hypertrophic cardiomyopathy [4,5], Prognosis [6], Acute mountain disease [7] | |
| MRPL9 | bL9m | Breast cancer [8] | |
| MRPL10 | uL10m | Early age-related macular degeneration [9] | |
| MRPL11 | uL11m | Mitochondrial encephalopathy [10] ↓ | |
| MRPL12/L7 | bL12m | Breast cancer [11] ↑, Colorectal cancer [12] | |
| MRPL13 | uL13m | Liver cancer [13] ↓, Breast cancer [13,14,15] ↑ | |
| MRPL15 | uL15m | Breast cancer [16] ↑ | |
| MRPL16 | uL16m | Septic cardiomyopathy [17] ↑ | |
| MRPL17 | bL17m | Lung cancer [18] ↑ | |
| MRPL19 | bL19m | Endometrial cancers [19,20], Diffuse non-Hodgkin lymphoma [21], Melanoma [22] | |
| MRPL21 | bL21m | Acute myeloid leukemia [23] ↑ | |
| MRPL23 | uL23m | Oral squamous cell carcinoma [24] ↓, Glioblastoma multiforme [25] | |
| MRPL24 | uL24m | Cerebellar atrophy, intellectual disability [26] ↓ | |
| MRPL28 | bL28m | Gastric cancer [27] | |
| MRPL33 | bL33m | Breast cancer [28] ↑, Lung cancer, colon cancer [29] ↑, Gastric cancer [30], Acute myeloid leukemia and neuroblastoma [31] ↑, Human papillomavirus associated oropharyngeal squamous cell carcinoma [32] ↑ | |
| MRPL34 | bL34m | Cardiomyocyte hypertrophy [33] ↓ | |
| MRPL35 | bL35m | Glioblastoma multiforme [34] ↑, Colorectal cancer [35] ↑ | |
| MRPL36 | bL36m | Cri-du-chat syndrome [36] | |
| MRPL37 | mL37 | Venous thromboembolism [37] | |
| MRPL38 | mL38 | Liver cancer [38] | |
| MRPL39 | mL39 | Gastric cancer [39] ↓ | |
| MRPL40 | mL40 | Schizophrenia [40,41,42] ↓ | |
| MRPL42 | mL42 | Glioma [43] ↑ | |
| MRPL43 | mL43 | Gastric cancer [44] ↑ | |
| MRPL44 | mL44 | Mitochondrial encephalopathy [10] ↓, Cardiomyopathy [45,46]. Hemiplegia migraine, pigmentary retinopathy, renal insufficiency, Leigh-like lesions on brain MRI [47], Asthma and allergy-related traits [48] | |
| MRPL46 | mL46 | Ovarian cancer [49] | |
| MRPL47 | mL47 | Acute lymphoblastic leukemia [50] | |
| MRPL50 | mL50 | Cardiomyocyte hypertrophy [33] ↑ | |
| MRPL51 | mL51 | Lung cancer [51] ↑ | |
| MRPL52 | mL52 | Colorectal cancer [52] ↓ | |
| MRPL54 | mL54 | Breast cancer [53] | |
| CRIF1 | mL64 | Hepatocellular carcinoma [54] ↓, T-cell leukemia [55] | Acute radiation syndrome [56], Endothelial inflammation [57,58] ↓, Autoimmune arthritis [59] ↑ |
| MRPS18-A | mL66 | Liver cancer [60] ↑, Cholangiocarcinoma [61] ↑ | |
| Mitochondrial ribosomal small subunit (mt-SSU) genes and proteins | |||
| MRPS2 | uS2m | Glioblastoma multiforme [25] | Cardiomyocyte hypertrophy [33] ↑ |
| MRPS5 | uS5m | Noise-induced hearing loss and anxiety related behavior changes [62] ↑ | |
| MRPS7 | uS7m | Osteosarcoma [63] ↑ | Primary hypogonadism, primary adrenal failure [64] ↓ |
| MRPS11 | uS11m | Uveal melanoma [65] ↑ | |
| MRPS12 | uS12m | Glioblastoma multiforme [25] | |
| MRPS14 | uS14m | Perinatal hypertrophic cardiomyopathy [66] ↑ | |
| MRPS18-B | mS40 | Prostate cancer [67] ↑, Colorectal carcinoma [68] | Tuberculosis [69] |
| MRPS18-C | bS18m | Breast cancer [70] ↑ | Epileptic encephalopathy [71] |
| MRPS21 | bS21m | Cardiomyocyte hypertrophy [33] ↑ | |
| MRPS22 | mS22 | Epicanthus inversus syndrome [72], Hypertrophic cardiomyopathy and fallopian tube lesions [73] ↓, Primary ovarian insufficiency [74,75,76] | |
| MRPS23 | mS23 | Hepatocellular carcinoma [77] ↑ | |
| MRPS34 | mS34 | Glioblastoma multiforme [25] | Cardiomyocyte hypertrophy [33] ↓ |
| MRPS37 | mS37 | Acute lymphoblastic leukemia [23] ↑ | |
| MRPS39 | mS39 | Leigh syndrome [78] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, G.; Li, H.; Zhang, H. Abnormal Expression of Mitochondrial Ribosomal Proteins and Their Encoding Genes with Cell Apoptosis and Diseases. Int. J. Mol. Sci. 2020, 21, 8879. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228879
Huang G, Li H, Zhang H. Abnormal Expression of Mitochondrial Ribosomal Proteins and Their Encoding Genes with Cell Apoptosis and Diseases. International Journal of Molecular Sciences. 2020; 21(22):8879. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228879
Chicago/Turabian StyleHuang, Guomin, Hongyan Li, and Hong Zhang. 2020. "Abnormal Expression of Mitochondrial Ribosomal Proteins and Their Encoding Genes with Cell Apoptosis and Diseases" International Journal of Molecular Sciences 21, no. 22: 8879. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228879