Organic Cation Transporters in the Lung—Current and Emerging (Patho)Physiological and Pharmacological Concepts


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Expression and Subcellular Localisation of OCT/Ns in Lung-Derived Cell Lines, Pulmonary Cell Cultures and Lung Tissues in Health and Disease
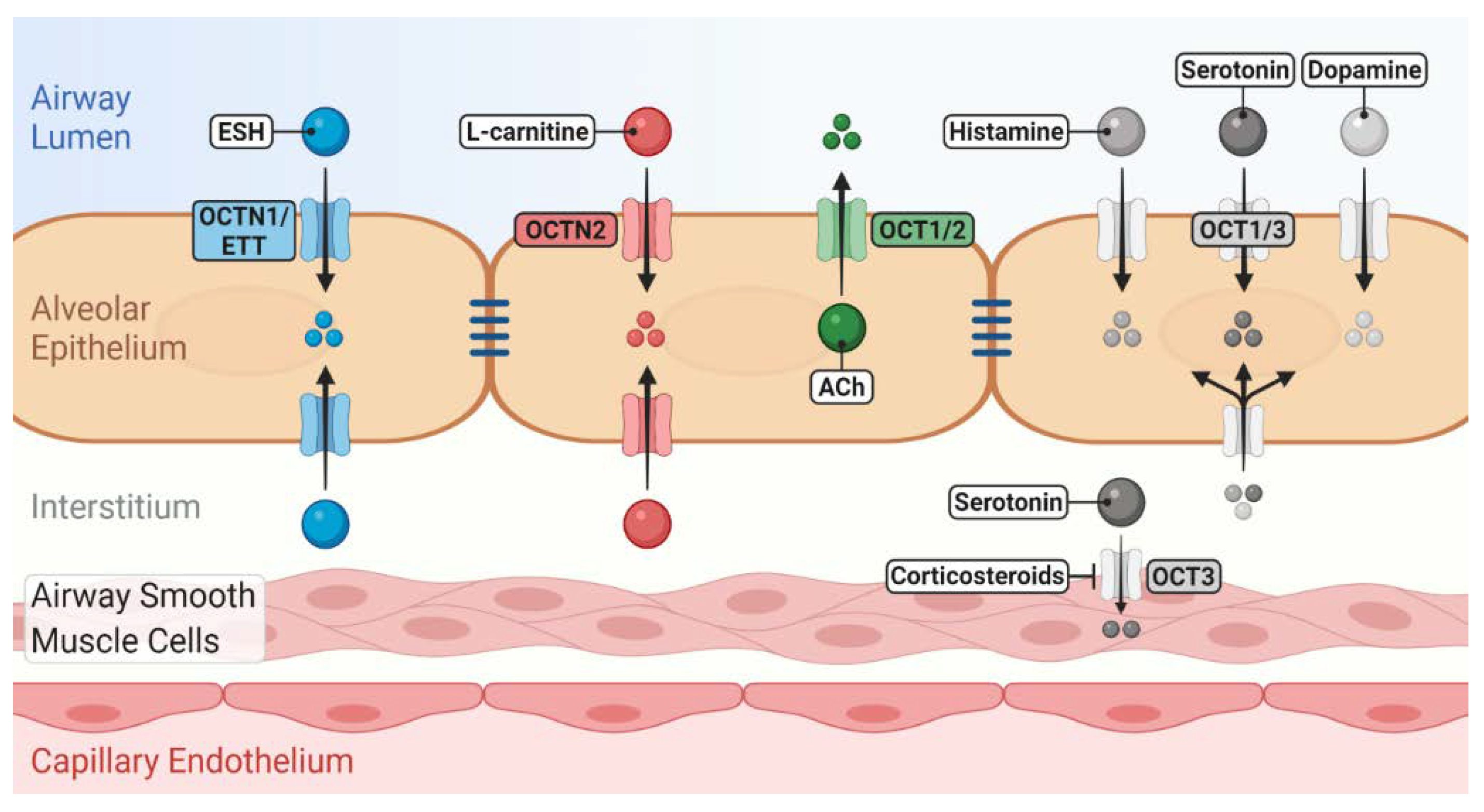
3. OCT/Ns (SLC22A1–A5) Transporter Function in Lung Physiology and Pathophysiology
3.1. OCT1, 2 and 3
3.2. OCTN1
3.3. OCTN2
3.4. Functional Studies Using Exogenous OCT/N Substrates
3.5. Relation of OCT/N Expression, Subcellular Localisation and Function
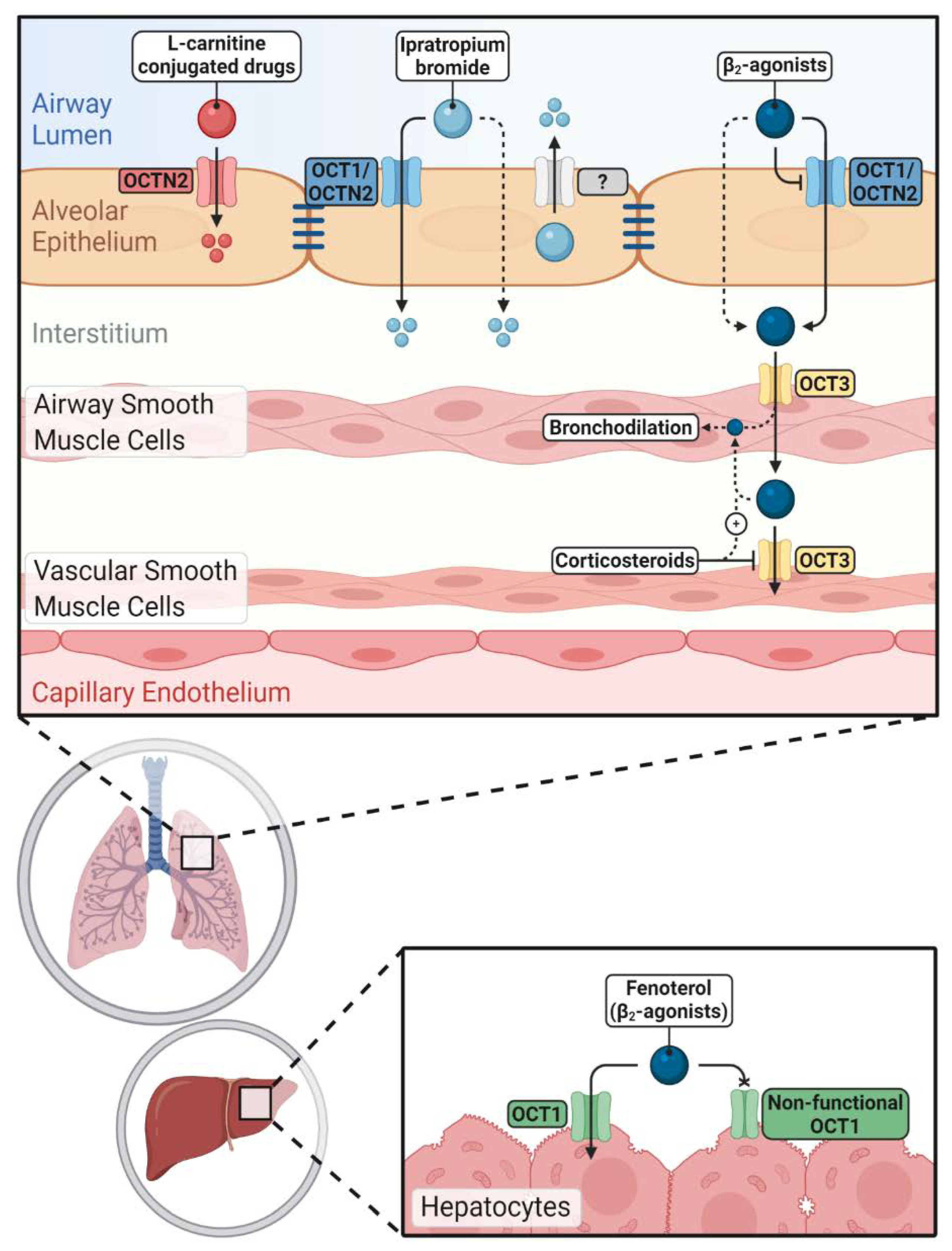
4. Pharmacological Aspects of OCT/N Transporters in the Lung
4.1. Interaction of OCT/N Transporters with Inhaled Drugs
4.1.1. Interaction with β2-Agonists
4.1.2. Interaction with Anticholinergic Drugs
4.2. OCTN2 as a Target to Enhance Pulmonary Drug Delivery
5. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACh | Acetylcholine |
| AIC | Air-interfaced culture |
| ASP+ | 4-[4-(dimethylamino)-styryl]-N-methylpyridinium |
| ATB0,+ | Amino acid transporter B0,+ |
| COPD | Chronic obstructive pulmonary disease |
| CSE | Cigarette smoke extract |
| ESH | Ergothioneine |
| ETT | Ergothioneine transporter |
| HDM | House dust mite extract |
| IHC | Immunohistochemistry |
| LCC | Liquid-covered conditions |
| LC-MS/MS | Liquid chromatography–tandem mass spectrometry |
| LPS | Lipopolysaccharide |
| MATE | Multidrug and toxin extrusion |
| MPP+ | 1-methyl-4-phenylpyridinium |
| MRPs | Multidrug-resistance-associated proteins |
| NF-κB | Nuclear factor kappa B |
| NHBE | Normal human bronchial epithelial |
| OATs | Organic anion transporters |
| OCT | Organic cation transporter |
| OCTN | Novel organic cation transporter |
| PDSC | Prednisolone succinate-l-carnitine |
| RDS | Respiratory distress syndrome |
| SLC | Solute carrier |
| TEA | Tetraethylammonium |
| TNF-α | Tumour necrosis factor α |
References
- Bosquillon, C. Drug transporters in the lung—Do they play a role in the biopharmaceutics of inhaled drugs? J. Pharm. Sci. 2010, 99, 2240–2255. [Google Scholar] [CrossRef] [PubMed]
- Salomon, J.J.; Ehrhardt, C. Organic cation transporters in the blood–air barrier: Expression and implications for pulmonary drug delivery. Ther. Deliv. 2012, 3, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Nickel, S.; Clerkin, C.G.; Selo, M.A.; Ehrhardt, C. Transport mechanisms at the pulmonary mucosa: Implications for drug delivery. Expert Opin. Drug Deliv. 2016, 13, 667–690. [Google Scholar] [CrossRef] [PubMed]
- Engelhart, D.; Granados, J.C.; Shi, D.; Saier, M.H.; Baker, M.E.; Abagyan, R.; Nigam, S.K.; Saier, M.H. Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs. Int. J. Mol. Sci. 2020, 21, 1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selo, M.A.; Al-Alak, H.H.; Ehrhardt, C. Lung transporters and absorption mechanisms in the lungs. In Inhalation Aerosols: Physical and Biological Basis for Therapy, 3rd ed.; Hickey, A.J., Mansour, H.M., Eds.; CRC Press: New York, NY, USA, 2019; Volume 1, pp. 57–70. [Google Scholar]
- Koepsell, H.; Lips, K.; Volk, C. Polyspecific Organic Cation Transporters: Structure, Function, Physiological Roles, and Biopharmaceutical Implications. Pharm. Res. 2007, 24, 1227–1251. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef]
- The Human Protein Atlas 19 November 2020. Available online: https://www.proteinatlas.org/ (accessed on 23 November 2020).
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- The International Transporter Consortium; International Transporter Consortium; Giacomini, K.M.; Huang, S.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.R.; Chu, X.; Dahlin, A.; Evers, R.; et al. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [CrossRef]
- Chen, L.; Yee, S.W.; Giacomini, K.M. OCT1 in hepatic steatosis and thiamine disposition. Cell Cycle 2015, 14, 283–284. [Google Scholar] [CrossRef]
- Chen, L.; Shu, Y.; Liang, X.; Chen, E.C.; Yee, S.W.; Zur, A.A.; Li, S.; Xu, L.; Keshari, K.R.; Lin, M.J.; et al. OCT1 is a high-capacity thiamine transporter that regulates hepatic steatosis and is a target of metformin. Proc. Natl. Acad. Sci. USA 2014, 111, 9983–9988. [Google Scholar] [CrossRef] [Green Version]
- Bacq, A.; Balasse, L.; Biala, G.; Guiard, B.P.; Gardier, A.; Schinkel, A.; Louis, F.; Vialou, V.; Martres, M.-P.; Chevarin, C.; et al. Organic cation transporter 2 controls brain norepinephrine and serotonin clearance and antidepressant response. Mol. Psychiatry 2012, 17, 926–939. [Google Scholar] [CrossRef] [Green Version]
- Couroussé, T.; Gautron, S. Role of organic cation transporters (OCTs) in the brain. Pharmacol. Ther. 2015, 146, 94–103. [Google Scholar] [CrossRef]
- Vialou, V.V.; Amphoux, A.; Zwart, R.; Giros, B.; Gautron, S. Organic Cation Transporter 3 (Slc22a3) Is Implicated in Salt-Intake Regulation. J. Neurosci. 2004, 24, 2846–2851. [Google Scholar] [CrossRef]
- Samodelov, S.L.; Kullak-Ublick, G.A.; Gai, Z.; Visentin, M. Organic Cation Transporters in Human Physiology, Pharmacology, and Toxicology. Int. J. Mol. Sci. 2020, 21, 7890. [Google Scholar] [CrossRef]
- Endter, S.; Francombe, D.; Gumbleton, M.; Ehrhardt, C. RT-PCR analysis of ABC, SLC and SLCO drug transporters in human lung epithelial cell models. J. Pharm. Pharmacol. 2009, 61, 583–591. [Google Scholar] [CrossRef]
- Ingoglia, F.; Visigalli, R.; Rotoli, B.M.; Barilli, A.; Riccardi, B.; Puccini, P.; Dall’Asta, V. Functional characterization of the organic cation transporters (OCTs) in human airway pulmonary epithelial cells. Biochim. Biophys. Acta 2015, 1848, 1563–1572. [Google Scholar] [CrossRef] [Green Version]
- Horvath, G.; Schmid, N.; Fragoso, M.A.; Schmid, A.; Conner, G.E.; Salathe, M.; Wanner, A. Epithelial Organic Cation Transporters Ensure pH-Dependent Drug Absorption in the Airway. Am. J. Respir. Cell Mol. Biol. 2007, 36, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, A.; Matsumaru, T.; Yamamura, N.; Suzuki, S.; Uchida, Y.; Tachikawa, M.; Terasaki, T. Drug Transporter Protein Quantification of Immortalized Human Lung Cell Lines Derived from Tracheobronchial Epithelial Cells (Calu-3 and BEAS2-B), Bronchiolar–Alveolar Cells (NCI-H292 and NCI-H441), and Alveolar Type II-like Cells (A549) by Liquid Chromatography–Tandem Mass Spectrometry. J. Pharm. Sci. 2015, 104, 3029–3038. [Google Scholar] [CrossRef]
- Sakamoto, A.; Matsumaru, T.; Yamamura, N.; Uchida, Y.; Tachikawa, M.; Ohtsuki, S.; Terasaki, T. Quantitative expression of human drug transporter proteins in lung tissues: Analysis of regional, gender, and interindividual differences by liquid chromatography–tandem mass spectrometry. J. Pharm. Sci. 2013, 102, 3395–3406. [Google Scholar] [CrossRef]
- Salomon, J.J.; Gausterer, J.C.; Selo, M.A.; Hosoya, K.-I.; Huwer, H.; Schneider-Daum, N.; Lehr, C.-M.; Ehrhardt, C. OCTN2-Mediated Acetyl-l-Carnitine Transport in Human Pulmonary Epithelial Cells In Vitro. J. Pharm. Sci. 2019, 11, 396. [Google Scholar] [CrossRef] [Green Version]
- Courcot, E.; Leclerc, J.; Lafitte, J.-J.; Mensier, E.; Jaillard, S.; Gosset, P.; Shirali, P.; Pottier, N.; Broly, F.; Lo-Guidice, J.-M. Xenobiotic Metabolism and Disposition in Human Lung Cell Models: Comparison with In Vivo Expression Profiles. Drug Metab. Dispos. 2012, 40, 1953–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barilli, A.; Visigalli, R.; Ferrari, F.; Di Lascia, M.; Riccardi, B.; Puccini, P.; Dall’Asta, V.; Rotoli, B.M. Organic Cation Transporters (OCTs) in EpiAirway™, a Cellular Model of Normal Human Bronchial Epithelium. Biomedicines 2020, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Pritchard, D.; Bosquillon, C. Evaluation of air-interfaced Calu-3 cell layers for investigation of inhaled drug interactions with organic cation transporters in vitro. Int. J. Pharm. 2012, 426, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Latif, M.; Pritchard, D.; Bosquillon, C. In-cell Western™ detection of organic cation transporters in bronchial epithelial cell layers cultured at an air–liquid interface on Transwell ® inserts. J. Pharmacol. Toxicol. Methods 2013, 68, 184–189. [Google Scholar] [CrossRef]
- Salomon, J.J.; Muchitsch, V.E.; Gausterer, J.C.; Schwagerus, E.; Huwer, H.; Daum, N.; Lehr, C.-M.; Ehrhardt, C. The Cell Line NCl-H441 Is a Usefulin VitroModel for Transport Studies of Human Distal Lung Epithelial Barrier. Mol. Pharm. 2014, 11, 995–1006. [Google Scholar] [CrossRef]
- Lips, K.S.; Volk, C.; Schmitt, B.M.; Pfeil, U.; Arndt, P.; Miska, D.; Ermert, L.; Kummer, W.; Koepsell, H. Polyspecific Cation Transporters Mediate Luminal Release of Acetylcholine from Bronchial Epithelium. Am. J. Respir. Cell Mol. Biol. 2005, 33, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Fallon, J.K.; Houvig, N.; Booth-Genthe, C.L.; Smith, P.C. Quantification of membrane transporter proteins in human lung and immortalized cell lines using targeted quantitative proteomic analysis by isotope dilution nanoLC–MS/MS. J. Pharm. Biomed. Anal. 2018, 154, 150–157. [Google Scholar] [CrossRef]
- Horvath, G.; Mendes, E.S.; Schmid, N.; Schmid, A.; Conner, G.E.; Salathe, M.; Wanner, A. The effect of corticosteroids on the disposal of long-acting β2-agonists by airway smooth muscle cells. J. Allergy Clin. Immunol. 2007, 120, 1103–1109. [Google Scholar] [CrossRef]
- Berg, T.; Hegelund-Myrbäck, T.; Öckinger, J.; Zhou, X.; Brännström, M.; Hagemann-Jensen, M.; Werkstrom, V.; Seidegård, J.; Grunewald, J.; Nord, M.; et al. Expression of MATE1, P-gp, OCTN1 and OCTN2, in epithelial and immune cells in the lung of COPD and healthy individuals. Respir. Res. 2018, 19, 68. [Google Scholar] [CrossRef] [Green Version]
- Berg, T.; Myrbäck, T.H.; Olsson, M.; Seidegård, J.; Werkström, V.; Zhou, X.; Grunewald, J.; Gustavsson, L.; Nord, M. Gene expression analysis of membrane transporters and drug-metabolizing enzymes in the lung of healthy and COPD subjects. Pharmacol. Res. Perspect. 2014, 2, e00054. [Google Scholar] [CrossRef]
- Shrine, N.; Portelli, M.A.; John, C.; Artigas, M.S.; Bennett, N.; Hall, R.; Lewis, J.; Henry, A.P.; Billington, C.K.; Ahmad, A.; et al. Moderate-to-severe asthma in individuals of European ancestry: A genome-wide association study. Lancet Respir. Med. 2019, 7, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, M.F.; Gut, I.G.; Demenais, F.; Strachan, D.P.; Bouzigon, E.; Heath, S.; Von Mutius, E.; Farrall, M.; Lathrop, M.; Cookson, W.O. A Large-Scale, Consortium-Based Genomewide Association Study of Asthma. N. Engl. J. Med. 2010, 363, 1211–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, K.; Shikanai, T.; Nakamura, Y.; Kobayashi, H.; Ogasawara, M.; Maeyama, K. Roles of Histamine in the Pathogenesis of Bronchial Asthma and Reevaluation of the Clinical Usefulness of Antihistamines. Yakugaku Zasshi 2011, 131, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, K.; Ogasawara, M. The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. Int. J. Mol. Sci. 2019, 20, 1733. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, M.; Cingolani, E.; Pritchard, D.; Bosquillon, C. Enhanced expression of Organic Cation Transporters in bronchial epithelial cell layers following insults associated with asthma – Impact on salbutamol transport. Eur. J. Pharm. Sci. 2017, 106, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Rotoli, B.M.; Visigalli, R.; Barilli, A.; Ferrari, F.; Bianchi, M.G.; Di Lascia, M.; Riccardi, B.; Puccini, P.; Dall’Asta, V. Functional analysis of OCTN2 and ATB0,+ in normal human airway epithelial cells. PLoS ONE 2020, 15, e0228568. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Qi, C.; Zhou, J.; Wen, Z.; Zhu, X.; Xia, H.; Song, J. LPS-induced inflammation delays the transportation of ASP+ due to down-regulation of OCTN1/2 in alveolar epithelial cells. J. Drug Target. 2019, 28, 437–447. [Google Scholar] [CrossRef]
- Qi, C.; Zhou, J.; Wang, Z.; Fang, X.; Li, D.; Jin, Y.; Song, J. Cigarette smoke extract combined with lipopolysaccharide reduces OCTN1/2 expression in human alveolar epithelial cells in vitro and rat lung in vivo under inflammatory conditions. Int. Immunopharmacol. 2020, 87, 106812. [Google Scholar] [CrossRef]
- Schulz, C.; Farkas, L.; Wolf, K.; Krätzel, K.; Eissner, G.; Pfeifer, M. Differences in LPS-Induced Activation of Bronchial Epithelial Cells (BEAS-2B) and Type II-Like Pneumocytes (A-549). Scand. J. Immunol. 2002, 56, 294–302. [Google Scholar] [CrossRef]
- Al-Majdoub, Z.M.; Al Feteisi, H.; Achour, B.; Warwood, S.; Neuhoff, S.; Rostami-Hodjegan, A.; Barber, J. Proteomic Quantification of Human Blood–Brain Barrier SLC and ABC Transporters in Healthy Individuals and Dementia Patients. Mol. Pharm. 2019, 16, 1220–1233. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Ligong, C.; Chen, L. The physiological role of drug transporters. Protein Cell 2015, 6, 334–350. [Google Scholar] [CrossRef] [Green Version]
- Majd, H.; King, M.S.; Palmer, S.M.; Smith, A.C.; Elbourne, L.D.; Paulsen, I.T.; Sharples, D.; Henderson, P.J.; Kunji, E.R. Screening of candidate substrates and coupling ions of transporters by thermostability shift assays. eLife 2018, 7. [Google Scholar] [CrossRef]
- Tschirka, J.; Bach, M.; Kisis, I.; Lemmen, J.; Gnoth, M.J.; Gründemann, D. Transporter tandems—Precise tools for normalizing active transporter in the plasma membrane. Biochem. J. 2020, 477, 4191–4206. [Google Scholar] [CrossRef] [PubMed]
- Boxberger, K.H.; Hagenbuch, B.; Lampe, J.N. Common drugs inhibit human organic cation transporter 1 (OCT1)-mediated neurotransmitter uptake. Drug Metab. Dispos. 2014, 42, 990–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, T.; Yanai, K. Histamine Clearance Through Polyspecific Transporters in the Brain. Muscarinic Recept. 2016, 241, 173–187. [Google Scholar] [CrossRef]
- Kummer, W.; Wiegand, S.; Akinci, S.; Wessler, I.; Schinkel, A.H.; Wess, J.; Koepsell, H.; Haberberger, R.V.; Lips, K. Role of acetylcholine and polyspecific cation transporters in serotonin-induced bronchoconstriction in the mouse. Respir. Res. 2006, 7, 65. [Google Scholar] [CrossRef] [Green Version]
- Kummer, W.; Krasteva, G. Non-neuronal cholinergic airway epithelium biology. Curr. Opin. Pharmacol. 2014, 16, 43–49. [Google Scholar] [CrossRef]
- Wessler, I.; Kirkpatrick, C.J.; Racké, K. Non-neuronal acetylcholine, a locally acting molecule, widely distributed in biological systems: Expression and function in humans. Pharmacol. Ther. 1998, 77, 59–79. [Google Scholar] [CrossRef]
- Kolahian, S.; Gosens, R. Cholinergic Regulation of Airway Inflammation and Remodelling. J. Allergy 2012, 2012, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Belmonte, K.E. Cholinergic Pathways in the Lungs and Anticholinergic Therapy for Chronic Obstructive Pulmonary Disease. Proc. Am. Thorac. Soc. 2005, 2, 297–304. [Google Scholar] [CrossRef]
- Reinheimer, T.; Baumgärtner, D.; Höhle, K.-D.; Racké, K.; Wessler, I. Acetylcholine via Muscarinic Receptors Inhibits Histamine Release from Human Isolated Bronchi. Am. J. Respir. Crit. Care Med. 1997, 156, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Gosens, R.; Zaagsma, J.; Meurs, H.; Halayko, A.J. Muscarinic receptor signaling in the pathophysiology of asthma and COPD. Respir. Res. 2006, 7, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, J.R.; Richbart, S.D.; Merritt, J.C.; Brown, K.C.; Nolan, N.A.; Akers, A.T.; Lau, J.K.; Robateau, Z.R.; Miles, S.L.; Dasgupta, P. Acetylcholine signaling system in progression of lung cancers. Pharmacol. Ther. 2019, 194, 222–254. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cohen, J.A.; Wallrapp, A.; Trieu, K.G.; Barrios, J.; Shao, F.; Krishnamoorthy, N.; Kuchroo, V.K.; Jones, M.R.; Fine, A.; et al. Age-Related Dopaminergic Innervation Augments T Helper 2-Type Allergic Inflammation in the Postnatal Lung. Immunity 2019, 51, 1102–1118.e7. [Google Scholar] [CrossRef] [PubMed]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [Green Version]
- Gelfand, E.W. Role of histamine in the pathophysiology of asthma: Immunomodulatory and anti-inflammatory activities of H1-receptor antagonists. Am. J. Med. 2002, 113, 2–7. [Google Scholar] [CrossRef]
- Cazzola, M.; Matera, M.G. 5-HT modifiers as a potential treatment of asthma. Trends Pharmacol. Sci. 2000, 21, 13–16. [Google Scholar] [CrossRef]
- Pirina, P.; Zinellu, E.; Paliogiannis, P.; Fois, A.G.; Marras, V.; Sotgia, S.; Carru, C.; Zinellu, A. Circulating serotonin levels in COPD patients: A pilot study. BMC Pulm. Med. 2018, 18, 167. [Google Scholar] [CrossRef]
- Wultsch, T.; Grimberg, G.; Schmitt, A.; Painsipp, E.; Wetzstein, H.; Breitenkamp, A.F.S.; Gründemann, D.; Schömig, E.; Lesch, K.P.; Gerlach, M.; et al. Decreased anxiety in mice lacking the organic cation transporter 3. J. Neural Transm. 2009, 116, 689–697. [Google Scholar] [CrossRef]
- Yee, S.W.; Lin, L.; Merski, M.; Keiser, L.G.M.J.; Gupta, A.; Zhang, Y.; Chien, H.-C.; Shoichet, B.K.; Giacomini, K.M. Prediction and validation of enzyme and transporter off-targets for metformin. J. Pharmacokinet. Pharmacodyn. 2015, 42, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Sekizawa, K.; Nakazawa, H.; Morikawa, M.; Yamauchi, K.; Maeyama, K.; Watanabe, T.; Sasaki, H. Histamine N-methyltransferase inhibitor potentiates histamine- and antigen-induced airway microvascular leakage in guinea pigs. J. Allergy Clin. Immunol. 1995, 96, 910–916. [Google Scholar] [CrossRef]
- Gründemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schömig, E. Discovery of the ergothioneine transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschirka, J.; Kreisor, M.; Betz, J.; Gründemann, D. Substrate Selectivity Check of the Ergothioneine Transporter. Drug Metab. Dispos. 2018, 46, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Grigat, S.; Harlfinger, S.; Pal, S.; Striebinger, R.; Golz, S.; Geerts, A.; Lazar, A.; Schömig, E.; Gründemann, D. Probing the substrate specificity of the ergothioneine transporter with methimazole, hercynine, and organic cations. Biochem. Pharmacol. 2007, 74, 309–316. [Google Scholar] [CrossRef]
- Cheah, I.K.; Tang, R.M.; Yew, T.S.; Lim, K.H.; Halliwell, B. Administration of Pure Ergothioneine to Healthy Human Subjects: Uptake, Metabolism, and Effects on Biomarkers of Oxidative Damage and Inflammation. Antioxid. Redox Signal. 2017, 26, 193–206. [Google Scholar] [CrossRef]
- Halliwell, B.; Cheah, I.K.; Tang, R.M.Y. Ergothioneine—A diet-derived antioxidant with therapeutic potential. FEBS Lett. 2018, 592, 3357–3366. [Google Scholar] [CrossRef] [Green Version]
- Cheah, I.; Halliwell, B. Could Ergothioneine Aid in the Treatment of Coronavirus Patients? Antioxidants 2020, 9, 595. [Google Scholar] [CrossRef]
- Asahi, T.; Wu, X.; Shimoda, H.; Hisaka, S.; Harada, E.; Kanno, T.; Nakamura, Y.; Kato, Y.; Osawa, T. A mushroom-derived amino acid, ergothioneine, is a potential inhibitor of inflammation-related DNA halogenation. Biosci. Biotechnol. Biochem. 2015, 80, 313–317. [Google Scholar] [CrossRef]
- Waller, S.; Tremelling, M.; Bredin, F.; Godfrey, L.; Howson, J.; Parkes, M. Evidence for association of OCTN genes and IBD5 with ulcerative colitis. Gut 2006, 55, 809–814. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Nelson, L.; Franke, A.; Poritz, L.; Li, T.-Y.; Wu, R.; Wang, Y.; MacNeill, C.; Thomas, N.J.; Schreiber, S.; et al. OCTN1 variant L503F is associated with familial and sporadic inflammatory bowel disease. J. Crohns Colitis 2010, 4, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Peltekova, V.D.; Wintle, R.F.; Rubin, L.A.; Amos, C.I.; Huang, Q.; Gu, X.; Newman, B.; Van Oene, M.; Cescon, D.; Greenberg, G.; et al. Functional variants of OCTN cation transporter genes are associated with Crohn disease. Nat. Genet. 2004, 36, 471–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meisel, P.; Pagels, S.; Grube, M.; Jedlitschky, G.; Völzke, H.; Kocher, T. Tooth loss and adiposity: Possible role of carnitine transporter (OCTN1/2) polymorphisms in women but not in men. Clin. Oral Investig. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Gilmour, P.S.; Jimenez, L.A.; Biswas, S.K.; Antonicelli, F.; Aruoma, O.I. Ergothioneine inhibits oxidative stress- and TNF-α-induced NF-κB activation and interleukin-8 release in alveolar epithelial cells. Biochem. Biophys. Res. Commun. 2003, 302, 860–864. [Google Scholar] [CrossRef]
- Biswas, S.; Hwang, J.W.; Kirkham, P.A.; Rahman, I. Pharmacological and Dietary Antioxidant Therapies for Chronic Obstructive Pulmonary Disease. Curr. Med. Chem. 2013, 20, 1496–1530. [Google Scholar] [CrossRef]
- Rahman, I.; Kilty, I. Antioxidant Therapeutic Targets in COPD. Curr. Drug Targets 2006, 7, 707–720. [Google Scholar] [CrossRef]
- Repine, J.E.; Elkins, N.D. Effect of ergothioneine on acute lung injury and inflammation in cytokine insufflated rats. Prev. Med. 2012, 54, S79–S82. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, R.; Tamai, I.; Yabuuchi, H.; Nezu, J.I.; Oku, A.; Sai, Y.; Shimane, M.; Tsuji, A. Na(+)-dependent carnitine transport by organic cation transporter (OCTN2): Its pharmacological and toxicological relevance. J. Pharmacol. Exp. Ther. 1999, 291, 778–784. [Google Scholar]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine transport and fatty acid oxidation. Biochim. Biophys. Acta (BBA) Bioenerg. 2016, 1863, 2422–2435. [Google Scholar] [CrossRef]
- Tamai, I.; Ohashi, R.; Nezu, J.-I.; Yabuuchi, H.; Oku, A.; Shimane, M.; Sai, Y.; Tsuji, A. Molecular and Functional Identification of Sodium Ion-dependent, High Affinity Human Carnitine Transporter OCTN2. J. Biol. Chem. 1998, 273, 20378–20382. [Google Scholar] [CrossRef] [Green Version]
- Kremser, K.; Stangl, H.; Pahan, K.; Singh, I. Nitric Oxide Regulates Peroxisomal Enzyme Activities. Clin. Chem. Lab. Med. 1995, 33, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, G.C.; McKenna, M.C. l-Carnitine and Acetyl-l-carnitine Roles and Neuroprotection in Developing Brain. Neurochem. Res. 2017, 42, 1661–1675. [Google Scholar] [CrossRef]
- Gülçin, I. Antioxidant and antiradical activities of l-carnitine. Life Sci. 2006, 78, 803–811. [Google Scholar] [CrossRef]
- Solarska, K.; Lewinska, A.; Karowicz-Bilińska, A.; Bartosz, G. The antioxidant properties of carnitine in vitro. Cell. Mol. Biol. Lett. 2010, 15, 90–97. [Google Scholar] [CrossRef]
- Şener, G.; Paskaloğlu, K.; Şatiroglu, H.; Alican, I.; Kaçmaz, A.; Sakarcan, A. L-Carnitine Ameliorates Oxidative Damage due to Chronic Renal Failure in Rats. J. Cardiovasc. Pharmacol. 2004, 43, 698–705. [Google Scholar] [CrossRef]
- Le Borgne, F.; Ravaut, G.; Bernard, A.; Demarquoy, J. L-carnitine protects C2C12 cells against mitochondrial superoxide overproduction and cell death. World J. Biol. Chem. 2017, 8, 86–94. [Google Scholar] [CrossRef]
- Cao, Y.; Qu, H.-J.; Li, P.; Wang, C.-B.; Wang, L.-X.; Han, Z.-W. Single dose administration of L-carnitine improves antioxidant activities in healthy subjects. Tohoku J. Exp. Med. 2011, 224, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-J.; Lin, J.-S.; Lin, Y.-C.; Lin, P.-T. Effects of L-carnitine supplementation on oxidative stress and antioxidant enzymes activities in patients with coronary artery disease: A randomized, placebo-controlled trial. Nutr. J. 2014, 13, 79. [Google Scholar] [CrossRef] [Green Version]
- Al-Biltagi, M.; Isa, M.; Bediwy, A.S.; Helaly, N.; El Lebedy, D.D. L-Carnitine Improves the Asthma Control in Children with Moderate Persistent Asthma. J. Allergy 2012, 2012, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Asilsoy, S.; Soylu, O.B.; Karaman, O.; Uzuner, N.; Kavukçu, S. Serum total and free carnitine levels in children with asthma. World J. Pediatr. 2009, 5, 60–62. [Google Scholar] [CrossRef]
- Otsubo, C.; Bharathi, S.; Uppala, R.; Ilkayeva, O.R.; Wang, N.; McHugh, K.; Zou, Y.; Wang, J.; Alcorn, J.F.; Zuo, Y.Y.; et al. Long-chain Acylcarnitines Reduce Lung Function by Inhibiting Pulmonary Surfactant. J. Biol. Chem. 2015, 290, 23897–23904. [Google Scholar] [CrossRef] [Green Version]
- Öztürk, M.A.; Kardas, Z.; Kardas, F.; Güneş, T.; Kurtoglu, S. Effects of L-carnitine supplementation on respiratory distress syndrome development and prognosis in premature infants: A single blind randomized controlled trial. Exp. Ther. Med. 2016, 11, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Gustavsson, L.; Bosquillon, C.; Gumbleton, M.; Hegelund-Myrbäck, T.; Nakanishi, T.; Price, D.; Tamai, I.; Zhou, X.-H. Drug Transporters in the Lung: Expression and Potential Impact on Pulmonary Drug Disposition; Royal Society of Chemistry (RSC): Cambridge, UK, 2016; Chapter 6; Volume 1, pp. 184–228. [Google Scholar]
- Al-Jayyoussi, G.; Price, D.F.; Kreitmeyr, K.; Keogh, J.P.; Smith, M.W.; Gumbleton, M.; Morris, C.J. Absorption of ipratropium and l -carnitine into the pulmonary circulation of the ex-vivo rat lung is driven by passive processes rather than active uptake by OCT/OCTN transporters. Int. J. Pharm. 2015, 496, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, R.; Tamai, I.; Nezu, J.-I.; Nikaido, H.; Hashimoto, N.; Oku, A.; Sai, Y.; Shimane, M.; Tsuji, A. Molecular and Physiological Evidence for Multifunctionality of Carnitine/Organic Cation Transporter OCTN2. Mol. Pharmacol. 2001, 59, 358–366. [Google Scholar] [CrossRef]
- Nakanishi, T.; Hasegawa, Y.; Haruta, T.; Wakayama, T.; Tamai, I. In Vivo Evidence of Organic Cation Transporter-Mediated Tracheal Accumulation of the Anticholinergic Agent Ipratropium in Mice. J. Pharm. Sci. 2013, 102, 3373–3381. [Google Scholar] [CrossRef]
- Salomon, J.J.; Endter, S.; Tachon, G.; Falson, F.; Buckley, S.T.; Ehrhardt, C. Transport of the fluorescent organic cation 4-(4-(dimethylamino)styryl)-N-methylpyridinium iodide (ASP+) in human respiratory epithelial cells. Eur. J. Pharm. Biopharm. 2012, 81, 351–359. [Google Scholar] [CrossRef]
- Macdonald, C.; Shao, D.; Oli, A.; Agu, R.U. Characterization of Calu-3 cell monolayers as a model of bronchial epithelial transport: Organic cation interaction studies. J. Drug Target. 2013, 21, 97–106. [Google Scholar] [CrossRef]
- Solis, E.; Zdravkovic, I.; Tomlinson, I.D.; Noskov, S.Y.; Rosenthal, S.J.; DeFelice, L.J. 4-(4-(Dimethylamino)phenyl)-1-methylpyridinium (APP+) Is a Fluorescent Substrate for the Human Serotonin Transporter. J. Biol. Chem. 2012, 287, 8852–8863. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, J.W.; Blakely, R.D.; DeFelice, L.J. Binding and Transport in Norepinephrine Transporters. J. Biol. Chem. 2003, 278, 9768–9777. [Google Scholar] [CrossRef] [Green Version]
- Salomon, J.J.; Gausterer, J.C.; Yahara, T.; Hosoya, K.-I.; Huwer, H.; Hittinger, M.; Schneider-Daum, N.; Lehr, C.-M.; Ehrhardt, C. Organic cation transporter function in different in vitro models of human lung epithelium. Eur. J. Pharm. Sci. 2015, 80, 82–88. [Google Scholar] [CrossRef]
- Koepsell, H. Multiple binding sites in organic cation transporters require sophisticated procedures to identify interactions of novel drugs. Biol. Chem. 2019, 400, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Koepsell, H. Role of organic cation transporters in drug–drug interaction. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1619–1633. [Google Scholar] [CrossRef] [PubMed]
- Salomon, J.J.; Hagos, Y.; Petzke, S.; Kühne, A.; Gausterer, J.C.; Hosoya, K.-I.; Ehrhardt, C. Beta-2 Adrenergic Agonists Are Substrates and Inhibitors of Human Organic Cation Transporter 1. Mol. Pharm. 2015, 12, 2633–2641. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Monteiro, R.; Pestana, D.; Martel, F.; De Freitas, V.; Mateus, N.; Calhau, C. Impact of culture media glucose levels on the intestinal uptake of organic cations. Cytotechnology 2010, 62, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreft, M.E.; Jerman, U.D.; Lasič, E.; Hevir-Kene, N.; Rižner, T.L.; Peternel, L.; Kristan, K. The characterization of the human cell line Calu-3 under different culture conditions and its use as an optimized in vitro model to investigate bronchial epithelial function. Eur. J. Pharm. Sci. 2015, 69, 1–9. [Google Scholar] [CrossRef]
- Kreft, M.E.; Tratnjek, L.; Lasič, E.; Hevir, N.; Rižner, T.L.; Kristan, K. Different Culture Conditions Affect Drug Transporter Gene Expression, Ultrastructure, and Permeability of Primary Human Nasal Epithelial Cells. Pharm. Res. 2020, 37, 170. [Google Scholar] [CrossRef]
- Forbes, B.; Ehrhardt, C. Human respiratory epithelial cell culture for drug delivery applications. Eur. J. Pharm. Biopharm. 2005, 60, 193–205. [Google Scholar] [CrossRef]
- Sporty, J.L.; Horálková, L.; Ehrhardt, C. In vitrocell culture models for the assessment of pulmonary drug disposition. Expert Opin. Drug Metab. Toxicol. 2008, 4, 333–345. [Google Scholar] [CrossRef]
- Sakagami, M. In vitro, ex vivo and in vivo methods of lung absorption for inhaled drugs. Adv. Drug Deliv. Rev. 2020. [Google Scholar] [CrossRef]
- Bosquillon, C.; Madlova, M.; Patel, N.; Clear, N.; Forbes, B. A Comparison of Drug Transport in Pulmonary Absorption Models: Isolated Perfused rat Lungs, Respiratory Epithelial Cell Lines and Primary Cell Culture. Pharm. Res. 2017, 34, 2532–2540. [Google Scholar] [CrossRef]
- Bleasby, K.; Castle, J.C.; Roberts, C.J.; Cheng, C.; Bailey, W.J.; Sina, J.F.; Kulkarni, A.V.; Hafey, M.J.; Evers, R.; Johnson, J.M.; et al. Expression profiles of 50 xenobiotic transporter genes in humans and pre-clinical species: A resource for investigations into drug disposition. Xenobiotica 2006, 36, 963–988. [Google Scholar] [CrossRef]
- Leclerc, J.; Ngangue, E.C.-N.; Cauffiez, C.; Allorge, D.; Pottier, N.; Lafitte, J.-J.; Debaert, M.; Jaillard, S.; Broly, F.; Lo-Guidice, J.-M. Xenobiotic metabolism and disposition in human lung: Transcript profiling in non-tumoral and tumoral tissues. Biochimie 2011, 93, 1012–1027. [Google Scholar] [CrossRef] [PubMed]
- Selo, M.A.; Delmas, A.-S.; Springer, L.; Zoufal, V.; Sake, J.A.; Clerkin, C.G.; Huwer, H.; Schneider-Daum, N.; Lehr, C.-M.; Nickel, S.; et al. Tobacco Smoke and Inhaled Drugs Alter Expression and Activity of Multidrug Resistance-Associated Protein-1 (MRP1) in Human Distal Lung Epithelial Cells in vitro. Front. Bioeng. Biotechnol. 2020, 8, 1030. [Google Scholar] [CrossRef] [PubMed]
- Kolli, A.R.; Kuczaj, A.K.; Martin, F.; Hayes, A.W.; Peitsch, M.C.; Hoeng, J. Bridging inhaled aerosol dosimetry to physiologically based pharmacokinetic modeling for toxicological assessment: Nicotine delivery systems and beyond. Crit. Rev. Toxicol. 2019, 49, 725–741. [Google Scholar] [CrossRef]
- Bäckman, P.; Arora, S.; Couet, W.; Forbes, B.; De Kruijf, W.; Paudel, A. Advances in experimental and mechanistic computational models to understand pulmonary exposure to inhaled drugs. Eur. J. Pharm. Sci. 2018, 113, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, C.; Kneuer, C.; Bies, C.; Lehr, C.-M.; Kim, K.-J.; Bakowsky, U. Salbutamol is actively absorbed across human bronchial epithelial cell layers. Pulm. Pharmacol. Ther. 2005, 18, 165–170. [Google Scholar] [CrossRef]
- Unwalla, H.J.; Horvath, G.; Roth, F.D.; Conner, G.E.; Salathe, M. Albuterol Modulates Its Own Transepithelial Flux via Changes in Paracellular Permeability. Am. J. Respir. Cell Mol. Biol. 2012, 46, 551–558. [Google Scholar] [CrossRef]
- Mamlouk, M.; Young, P.M.; Bebawy, M.; Haghi, M.; Mamlouk, S.; Mulay, V.; Traini, D. Salbutamol Sulfate Absorption across Calu-3 Bronchial Epithelia Cell Monolayer is Inhibited in the Presence of Common Anionic NSAIDs. J. Asthma 2013, 50, 334–341. [Google Scholar] [CrossRef]
- Gnadt, M.; Trammer, B.; Freiwald, M.; Kardziev, B.; Bayliss, M.K.; Edwards, C.D.; Schmidt, M.; Friedel, G.; Högger, P. Methacholine delays pulmonary absorption of inhaled β2-agonists due to competition for organic cation/carnitine transporters. Pulm. Pharmacol. Ther. 2012, 25, 124–134. [Google Scholar] [CrossRef]
- Hendrickx, R.; Johansson, J.G.; Lohmann, C.; Jenvert, R.-M.; Blomgren, A.; Börjesson, L.; Gustavsson, L. Identification of Novel Substrates and Structure–Activity Relationship of Cellular Uptake Mediated by Human Organic Cation Transporters 1 and 2. J. Med. Chem. 2013, 56, 7232–7242. [Google Scholar] [CrossRef]
- Tzvetkov, M.V.; Matthaei, J.; Pojar, S.; Faltraco, F.; Vogler, S.; Prukop, T.; Seitz, T.; Brockmöller, J. Increased Systemic Exposure and Stronger Cardiovascular and Metabolic Adverse Reactions to Fenoterol in Individuals with HeritableOCT1Deficiency. Clin. Pharmacol. Ther. 2018, 103, 868–878. [Google Scholar] [CrossRef]
- Horvath, G.; Mendes, E.S.; Schmid, N.; Schmid, A.; Conner, G.E.; Fregien, N.L.; Salathe, M.; Wanner, A. Rapid nongenomic actions of inhaled corticosteroids on long-acting β2-agonist transport in the airway. Pulm. Pharmacol. Ther. 2011, 24, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, T.; Haruta, T.; Shirasaka, Y.; Tamai, I. Organic Cation Transporter-Mediated Renal Secretion of Ipratropium and Tiotropium in Rats and Humans. Drug Metab. Dispos. 2010, 39, 117–122. [Google Scholar] [CrossRef]
- Nakamura, T.; Nakanishi, T.; Haruta, T.; Shirasaka, Y.; Keogh, J.P.; Tamai, I. Transport of Ipratropium, an Anti-Chronic Obstructive Pulmonary Disease Drug, Is Mediated by Organic Cation/Carnitine Transporters in Human Bronchial Epithelial Cells: Implications for Carrier-Mediated Pulmonary Absorption. Mol. Pharm. 2010, 7, 187–195. [Google Scholar] [CrossRef]
- Panduga, V.; Stocks, M.J.; Bosquillon, C. Ipratropium is ‘luminally recycled’ by an inter-play between apical uptake and efflux transporters in Calu-3 bronchial epithelial cell layers. Int. J. Pharm. 2017, 532, 328–336. [Google Scholar] [CrossRef]
- Zhou, S.-F.; Wang, L.-L.; Di, Y.M.; Xue, C.C.; Duan, W.; Li, C.G.; Li, Y. Substrates and inhibitors of human multidrug resistance associated proteins and the implications in drug development. Curr. Med. Chem. 2008, 15, 1981–2039. [Google Scholar] [CrossRef]
- Shitara, Y.; Sato, H.; Sugiyama, Y. EVALUATION OF DRUG-DRUG INTERACTION IN THE HEPATOBILIARY AND RENAL TRANSPORT OF DRUGS. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 689–723. [Google Scholar] [CrossRef]
- Hutter, V.; Chau, D.Y.; Hilgendorf, C.; Brown, A.; Cooper, A.; Zann, V.; Pritchard, D.I.; Bosquillon, C. Digoxin net secretory transport in bronchial epithelial cell layers is not exclusively mediated by P-glycoprotein/MDR1. Eur. J. Pharm. Biopharm. 2014, 86, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Brockmöller, J.; Seitz, T.; König, J.; Tzvetkov, M.V.; Chen, X. Tropane alkaloids as substrates and inhibitors of human organic cation transporters of the SLC22 (OCT) and the SLC47 (MATE) families. Biol. Chem. 2017, 398, 237–249. [Google Scholar] [CrossRef]
- Müller, F.; König, J.; Hoier, E.; Mandery, K.; Fromm, M.F. Role of organic cation transporter OCT2 and multidrug and toxin extrusion proteins MATE1 and MATE2-K for transport and drug interactions of the antiviral lamivudine. Biochem. Pharmacol. 2013, 86, 808–815. [Google Scholar] [CrossRef]
- Ingoglia, F.; Visigalli, R.; Rotoli, B.M.; Barilli, A.; Riccardi, B.; Puccini, P.; Dall’Asta, V. Functional activity of L-carnitine transporters in human airway epithelial cells. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 210–219. [Google Scholar] [CrossRef]
- Bäckström, E.; Lundqvist, A.; Boger, E.; Svanberg, P.; Ewing, P.; Hammarlund-Udenaes, M.; Fridén, M. Development of a Novel Lung Slice Methodology for Profiling of Inhaled Compounds. J. Pharm. Sci. 2016, 105, 838–845. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Wang, Y.; Liu, S.; Wang, Y.; Liu, Q.; Liu, G.; Chen, Q. Emerging transporter-targeted nanoparticulate drug delivery systems. Acta Pharm. Sin. B 2019, 9, 49–58. [Google Scholar] [CrossRef]
- Kou, L.; Bhutia, Y.D.; Yao, Q.; He, Z.; Sun, J.; Ganapathy, V. Transporter-Guided Delivery of Nanoparticles to Improve Drug Permeation across Cellular Barriers and Drug Exposure to Selective Cell Types. Front. Pharmacol. 2018, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Kou, L.; Yao, Q.; Sun, M.; Wu, C.; Wang, J.; Luo, Q.; Wang, G.; Du, Y.; Fu, Q.; He, Z.; et al. Cotransporting Ion is a Trigger for Cellular Endocytosis of Transporter-Targeting Nanoparticles: A Case Study of High-Efficiency SLC22A5 (OCTN2)-Mediated Carnitine-Conjugated Nanoparticles for Oral Delivery of Therapeutic Drugs. Adv. Health Mater. 2017, 6. [Google Scholar] [CrossRef]
- Mo, J.; Shi, S.; Zhang, Q.; Gong, T.; Sun, X.; Zhang, Z.-R. Synthesis, Transport and Mechanism of a Type I Prodrug: L-Carnitine Ester of Prednisolone. Mol. Pharm. 2011, 8, 1629–1640. [Google Scholar] [CrossRef]
- Mo, J.; Lim, L.Y.; Zhang, Z.-R. l-Carnitine ester of prednisolone: Pharmacokinetic and pharmacodynamic evaluation of a type I prodrug. Int. J. Pharm. 2014, 475, 123–129. [Google Scholar] [CrossRef]
- Eriksson, J.; Sjögren, E.; Lennernäs, H.; Thörn, H. Drug Absorption Parameters Obtained Using the Isolated Perfused Rat Lung Model Are Predictive of Rat In Vivo Lung Absorption. AAPS J. 2020, 22, 71. [Google Scholar] [CrossRef]
- Eriksson, J.; Thörn, H.; Sjögren, E.; Holmstén, L.; Rubin, K.; Lennernäs, H. Pulmonary Dissolution of Poorly Soluble Compounds Studied in an ex Vivo Rat Lung Model. Mol. Pharm. 2019, 16, 3053–3064. [Google Scholar] [CrossRef]
- Kamusella, P.C.; Wissgott, C.; Grosse-Siestrup, C.; Dittrich, S.; Hegemann, O.; Koios, D.; Von Massenbach, J.; Meissler, M.; Unger, V.; Groneberg, D.A.; et al. A model of isolated, autologously hemoperfused porcine slaughterhouse lungs. ALTEX 2009, 26, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Floerl, S.; Kuehne, A.; Hagos, Y. Functional and Pharmacological Comparison of Human, Mouse, and Rat Organic Cation Transporter 1 toward Drug and Pesticide Interaction. Int. J. Mol. Sci. 2020, 21, 6871. [Google Scholar] [CrossRef]
- Cingolani, E.; Alqahtani, S.; Sadler, R.C.; Prime, D.; Stolnik, S.; Bosquillon, C. In vitro investigation on the impact of airway mucus on drug dissolution and absorption at the air-epithelium interface in the lungs. Eur. J. Pharm. Biopharm. 2019, 141, 210–220. [Google Scholar] [CrossRef]
- Alqahtani, S.; Roberts, C.J.; Stolnik, S.; Bosquillon, C. Development of an In Vitro System to Study the Interactions of Aerosolized Drugs with Pulmonary Mucus. Pharmaceutics 2020, 12, 145. [Google Scholar] [CrossRef] [Green Version]
- Faner, R.; Agustí, À. COPD: Algorithms and clinical management. Eur. Respir. J. 2017, 50, 1701733. [Google Scholar] [CrossRef] [Green Version]
- Aebersold, R.; Mann, M. Mass-spectrometric exploration of proteome structure and function. Nat. Cell Biol. 2016, 537, 347–355. [Google Scholar] [CrossRef]
- Erb-Downward, J.R.; Thompson, D.L.; Han, M.K.; Freeman, C.M.; McCloskey, L.; Schmidt, L.A.; Young, V.B.; Toews, G.B.; Curtis, J.L.; Sundaram, B.; et al. Analysis of the Lung Microbiome in the “Healthy” Smoker and in COPD. PLoS ONE 2011, 6, e16384. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Nuñez, M.; Millares, L.; Pomares, X.; Ferrari, R.; Pérez-Brocal, V.; Gallego, M.; Espasa, M.; Moya, A.; Monsó, E. Severity-Related Changes of Bronchial Microbiome in Chronic Obstructive Pulmonary Disease. J. Clin. Microbiol. 2014, 52, 4217–4223. [Google Scholar] [CrossRef] [Green Version]
- Brewington, J.J.; Filbrandt, E.T.; LaRosa, F.; Ostmann, A.J.; Strecker, L.M.; Szczesniak, R.D.; Clancy, J.P. Detection of CFTR function and modulation in primary human nasal cell spheroids. J. Cyst. Fibros. 2018, 17, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Sachs, N.; Papaspyropoulos, A.; Ommen, D.D.Z.; Heo, I.; Böttinger, L.; Klay, D.; Weeber, F.; Huelsz-Prince, G.; Iakobachvili, N.; Amatngalim, G.D.; et al. Long-term expanding human airway organoids for disease modeling. EMBO J. 2019, 38. [Google Scholar] [CrossRef]
- Yang, L.; Li, Y. Sex Differences in the Expression of Drug-Metabolizing and Transporter Genes in Human Liver. J. Drug Metab. Toxicol. 2012, 3, 1000119. [Google Scholar] [CrossRef]
- Lyn-Cook, B.; Starlard-Davenport, A.; Word, B.; Green, B.; Wise, C.; Ning, B.; Huang, Y. Gender differences in expression of drug transporter genes in human livers: Microarray and real-time PCR analysis. AACR 2008, 14, B34. [Google Scholar]
- Schuetz, E.G.; Furuya, K.N.; Schuetz, J.D. Interindividual variation in expression of P-glycoprotein in normal human liver and secondary hepatic neoplasms. J. Pharmacol. Exp. Ther. 1995, 275, 1011–1018. [Google Scholar]
- Joseph, S.; Nicolson, T.J.; Hammons, G.; Word, B.; Green-Knox, B.; Lyn-Cook, B. Expression of drug transporters in human kidney: Impact of sex, age, and ethnicity. Biol. Sex Differ. 2015, 6, 4. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selo, M.A.; Sake, J.A.; Ehrhardt, C.; Salomon, J.J. Organic Cation Transporters in the Lung—Current and Emerging (Patho)Physiological and Pharmacological Concepts. Int. J. Mol. Sci. 2020, 21, 9168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239168
Selo MA, Sake JA, Ehrhardt C, Salomon JJ. Organic Cation Transporters in the Lung—Current and Emerging (Patho)Physiological and Pharmacological Concepts. International Journal of Molecular Sciences. 2020; 21(23):9168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239168
Chicago/Turabian StyleSelo, Mohammed Ali, Johannes A. Sake, Carsten Ehrhardt, and Johanna J. Salomon. 2020. "Organic Cation Transporters in the Lung—Current and Emerging (Patho)Physiological and Pharmacological Concepts" International Journal of Molecular Sciences 21, no. 23: 9168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239168