Glycosylation in Indolent, Significant and Aggressive Prostate Cancer by Automated High-Throughput N-Glycan Profiling

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Patient Cohort
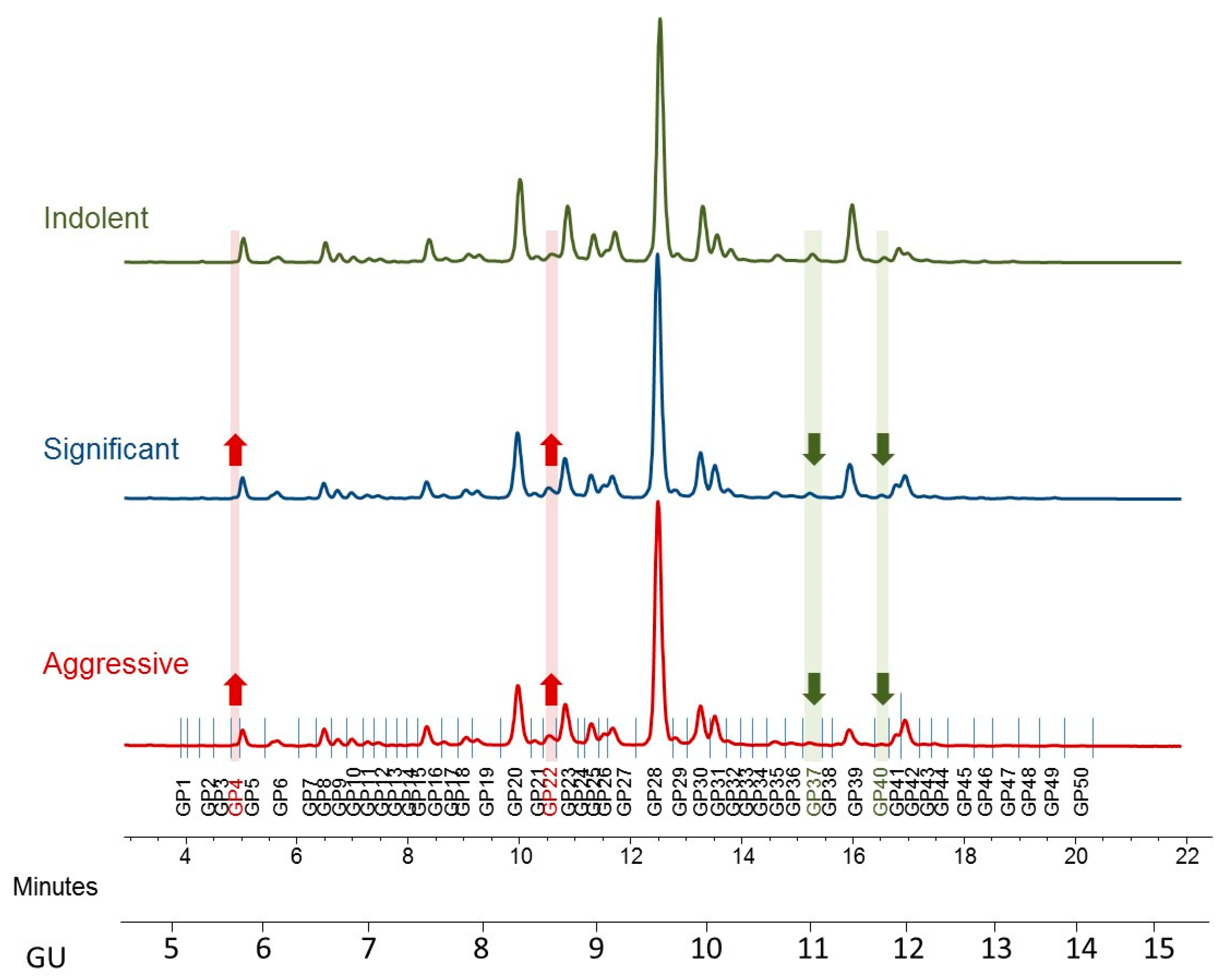
2.2. HILIC-UPLC Profiling of PCa Patients’ Serum
2.3. Significant Changes in Whole Serum N-Glycome in Prostate Cancer
3. Discussion
3.1. Increases in Oligomannose Glycans
3.2. Alterations in Branching and Bisecting Glycans
4. Materials and Methods
4.1. Serum Samples
4.2. N-Glycan Release and Clean up
4.3. Hydrophilic Interaction Chromatography-Ultra-Performance Liquid Chromatography (UPLC)
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2AB | 2-aminobenzamide |
| AIC | Akaike information criterion |
| ANOVA | Analysis of variance |
| AUC | Area under the curve |
| BEH | Ethylene Bridged Hybrid |
| BPH | Benign prostate hyperplasia |
| CE | Capillary electrophoresis |
| DRE | Digital rectal examination |
| EGFR | Epidermal growth factor receptor |
| GS | Gleason score |
| GU | Glucose unit |
| HILIC | Hydrophilic interaction liquid chromatography |
| HPAEC | High performance anion-exchange chromatography |
| HPLC | High performance liquid chromatography |
| HSD | Honestly significant difference |
| MALDI | Matrix-assisted laser desorption/ionization |
| NIBRT | National Institute of Bioprocessing Research and Training |
| PCa | Prostate cancer |
| PCRC | Prostate Cancer Research Consortium |
| PNI | Perineural invasion |
| PSA | Prostate specific antigen |
| ROC | Receiver operating characteristic |
| RP–LC | Reverse phase liquid chromatography |
| SPE | Solid phase extraction |
| TRIPOD | Transparent Reporting of a multivariable prediction model for Individual Prognosis Or Diagnosis |
| UPLC | Ultra-performance liquid chromatography |
| WAX | Weak anion exchange |
References
- Rawla, P. Epidemiology of Prostate Cancer, 2019. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilgunn, S.; Conroy, P.J.; Saldova, R.; Rudd, P.M.; O’Kennedy, R.J. Aberrant PSA glycosylation—A sweet predictor of prostate cancer. Nat. Rev. Urol. 2013, 10, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Dimakakos, A.; Armakolas, A.; Koutsilieris, M. Novel tools for prostate cancer prognosis, diagnosis, and follow-up. Biomed. Res. Int. 2014, 2014, 890697. [Google Scholar] [CrossRef] [PubMed]
- Dube, D.H.; Bertozzi, C.R. Glycans in cancer and inflammation—Potential for therapeutics and diagnostics. Nat. Rev. Drug Discov. 2005, 4, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Stockmann, H.; Adamczyk, B.; Hayes, J.; Rudd, P.M. Automated, high-throughput IgG-antibody glycoprofiling platform. Anal. Chem. 2013, 85, 8841–8849. [Google Scholar] [CrossRef] [PubMed]
- Stöckmann, H.; O’Flaherty, R.; Adamczyk, B.; Saldova, R.; Rudd, P.M. Automated, high-throughput serum glycoprofiling platform. Integr. Biol. 2015, 7, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.P.; Royle, L.; Radcliffe, C.M.; Dwek, R.A.; Rudd, P.M. GlycoBase and autoGU: Tools for HPLC-based glycan analysis. Bioinformatics 2008, 24, 1214–1216. [Google Scholar] [CrossRef]
- Royle, L.; Campbell, M.P.; Radcliffe, C.M.; White, D.M.; Harvey, D.J.; Abrahams, J.L.; Kim, Y.G.; Henry, G.W.; Shadick, N.A.; Weinblatt, M.E.; et al. HPLC-based analysis of serum N-glycans on a 96-well plate platform with dedicated database software. Anal. Biochem. 2008, 376, 1–12. [Google Scholar] [CrossRef]
- Royle, L.; Radcliffe, C.M.; Dwek, R.A.; Rudd, P.M. Detailed structural analysis of N-glycans released from glycoproteins in SDS-PAGE gel bands using HPLC combined with exoglycosidase array digestions. Methods Mol. Biol. 2006, 347, 125–143. [Google Scholar]
- Saldova, R.; Asadi Shehni, A.; Haakensen, V.D.; Steinfeld, I.; Hilliard, M.; Kifer, I.; Helland, A.; Yakhini, Z.; Borresen-Dale, A.L.; Rudd, P.M. Association of N-glycosylation with breast carcinoma and systemic features using high-resolution quantitative UPLC. J. Proteome Res. 2014, 13, 2314–2327. [Google Scholar] [CrossRef]
- Mariño, K.; Saldova, R.; Adamczyk, B.; Rudd, P.M. Changes in Serum N-Glycosylation Profiles: Functional Significance and Potential for Diagnostics. In Carbohydrate Chemistry: Chemical and Biological Approaches; Rauter, A.P., Ed.; RSC Publishing: Cambridge, UK, 2012; Volume 37. [Google Scholar]
- Murphy, K.; Murphy, B.T.; Boyce, S.; Flynn, L.; Gilgunn, S.; O’Rourke, C.J.; Rooney, C.; Stockmann, H.; Walsh, A.L.; Finn, S.; et al. Integrating biomarkers across omic platforms: An approach to improve stratification of patients with indolent and aggressive prostate cancer. Mol. Oncol. 2018, 12, 1513–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldova, R.; Fan, Y.; Fitzpatrick, J.M.; Watson, R.W.; Rudd, P.M. Core fucosylation and alpha2-3 sialylation in serum N-glycome is significantly increased in prostate cancer comparing to benign prostate hyperplasia. Glycobiology 2011, 21, 195–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totten, S.M.; Adusumilli, R.; Kullolli, M.; Tanimoto, C.; Brooks, J.D.; Mallick, P.; Pitteri, S.J. Multi-lectin Affinity Chromatography and Quantitative Proteomic Analysis Reveal Differential Glycoform Levels between Prostate Cancer and Benign Prostatic Hyperplasia Sera. Sci. Rep. 2018, 8, 6509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneyama, T.; Ohyama, C.; Hatakeyama, S.; Narita, S.; Habuchi, T.; Koie, T.; Mori, K.; Hidari, K.I.; Yamaguchi, M.; Suzuki, T.; et al. Measurement of aberrant glycosylation of prostate specific antigen can improve specificity in early detection of prostate cancer. Biochem. Biophys. Res. Commun. 2014, 448, 390–396. [Google Scholar] [CrossRef]
- Scott, E.; Munkley, J. Glycans as Biomarkers in Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 1389. [Google Scholar] [CrossRef] [Green Version]
- Epstein, J.I. An update of the Gleason grading system. J. Urol. 2010, 183, 433–440. [Google Scholar] [CrossRef]
- Epstein, J.I.; Walsh, P.C.; Carmichael, M.; Brendler, C.B. Pathologic and clinical findings to predict tumor extent of nonpalpable (stage T1c) prostate cancer. JAMA 1994, 271, 368–374. [Google Scholar] [CrossRef]
- McCarthy, C.; Saldova, R.; O’Brien, M.E.; Bergin, D.A.; Carroll, T.P.; Keenan, J.; Meleady, P.; Henry, M.; Clynes, M.; Rudd, P.M.; et al. Increased outer arm and core fucose residues on the N-glycans of mutated alpha-1 antitrypsin protein from alpha-1 antitrypsin deficient individuals. J. Proteome Res. 2014, 13, 596–605. [Google Scholar] [CrossRef]
- Banerjee, D.K. N-glycans in cell survival and death: Cross-talk between glycosyltransferases. Biochim Biophys. Acta 2012, 1820, 1338–1346. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Hatakeyama, S.; Yoneyama, T.; Tobisawa, Y.; Ishibashi, Y.; Yamamoto, H.; Yoneyama, T.; Hashimoto, Y.; Ito, H.; Nishimura, S.I.; et al. Serum N-glycan profiling is a potential biomarker for castration-resistant prostate cancer. Sci. Rep. 2019, 9, 16761. [Google Scholar] [CrossRef]
- Sethi, M.K.; Thaysen-Andersen, M.; Smith, J.T.; Baker, M.S.; Packer, N.H.; Hancock, W.S.; Fanayan, S. Comparative N-glycan profiling of colorectal cancer cell lines reveals unique bisecting GlcNAc and alpha-2,3-linked sialic acid determinants are associated with membrane proteins of the more metastatic/aggressive cell lines. J. Proteome Res. 2014, 13, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Kaprio, T.; Satomaa, T.; Heiskanen, A.; Hokke, C.H.; Deelder, A.M.; Mustonen, H.; Hagstrom, J.; Carpen, O.; Saarinen, J.; Haglund, C. N-glycomic profiling as a tool to separate rectal adenomas from carcinomas. Mol. Cell Proteom. 2015, 14, 277–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyalwidhe, J.O.; Betesh, L.R.; Powers, T.W.; Jones, E.E.; White, K.Y.; Burch, T.C.; Brooks, J.; Watson, M.T.; Lance, R.S.; Troyer, D.A.; et al. Increased bisecting N-acetylglucosamine and decreased branched chain glycans of N-linked glycoproteins in expressed prostatic secretions associated with prostate cancer progression. Proteom. Clin. Appl. 2013, 7, 677–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, T.; Ullrich, S.; Muller, I.; Nentwich, M.F.; Stubke, K.; Feldhaus, S.; Knies, C.; Hellwinkel, O.J.; Vessella, R.L.; Abramjuk, C.; et al. Human prostate cancer in a clinically relevant xenograft mouse model: Identification of beta(1,6)-branched oligosaccharides as a marker of tumor progression. Clin. Cancer Res. 2012, 18, 1364–1373. [Google Scholar] [CrossRef] [Green Version]
- Kyselova, Z.; Mechref, Y.; Al Bataineh, M.M.; Dobrolecki, L.E.; Hickey, R.J.; Vinson, J.; Sweeney, C.J.; Novotny, M.V. Alterations in the serum glycome due to metastatic prostate cancer. J. Proteome Res. 2007, 6, 1822–1832. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, Y.; Tobisawa, Y.; Hatakeyama, S.; Ohashi, T.; Tanaka, M.; Narita, S.; Koie, T.; Habuchi, T.; Nishimura, S.; Ohyama, C.; et al. Serum tri- and tetra-antennary N-glycan is a potential predictive biomarker for castration-resistant prostate cancer. Prostate 2014, 74, 1521–1529. [Google Scholar] [CrossRef]
- Saldova, R.; Huffman, J.E.; Adamczyk, B.; Muzinic, A.; Kattla, J.J.; Pucic, M.; Novokmet, M.; Abrahams, J.L.; Hayward, C.; Rudan, I.; et al. Association of medication with the human plasma N-glycome. J. Proteome Res. 2012, 11, 1821–1831. [Google Scholar] [CrossRef]
- Hamfjord, J.; Saldova, R.; Stöckmann, H.; Sandhu, V.; Bowitz, L.I.M.; Buanes, T.; Lingjærde, O.C.; Labori, K.J.; Rudd, P.M.; Kure, E.H. Serum N-Glycome Characterization in Patients with Resectable Periampullary Adenocarcinoma. J. Proteome Res. 2015, 14, 5144–5156. [Google Scholar] [CrossRef]
- Aitchison, J. The statistical analysis of compositional data. J. R. Stat. Soc. Ser. B 1982, 44, 139–177. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 24 November 2020).




{kind=link}
{kind=link}
{kind=link}
| Full Cohort (n = 117) | Indolent (n = 41) | Significant (n = 32) | Aggressive (n = 44) | |
|---|---|---|---|---|
| Age (y) | ||||
| Mean (median) | 60.6 (60) | 59.1 (59) | 59.9 (59.5) | 62.6 (62.5) |
| Range | 49–74 | 49–70 | 49–73 | 52–74 |
| Pre-Op PSA (ng/mL) | ||||
| Mean (median) | 7.05 (6.5) | 6.5 (6) | 7.11 (6.3) | 7.51 (6.7) |
| Range | 0.7–18.7 | 0.7–12.3 | 2.8–14.7 | 3.1–18.7 |
| Clinical Stage-DRE (%) | ||||
| T1c | 61 (52) | 24 (58) | 19 (59) | 18 (41) |
| T2a | 21 (18) | 4 (10) | 6 (19) | 11 (25) |
| Not described | 35 (30) | 13 (32) | 7 (22) | 15 (34) |
| Biopsy GS (%) | ||||
| ≤6 | 57 (49) | 38 (93) | 11 (34) | 8 (18) |
| 3 + 4 = 7 | 35 (30) | 3 (7) | 16 (50) | 16 (36) |
| 4 + 3 = 7 | 13 (11) | 0 (0) | 4 (13) | 9 (20) |
| ≥8 | 11 (9) | 0 (0) | 1 (3) | 10 (22) |
| Not described | 1 (1) | 0 (0) | 0 (0) | 1 (2) |
| Prostatectomy Stage | ||||
| OC | 73 (62) | 41 (100) | 32 (100) | 0 (0) |
| NOC | 44 (38) | 0 (0) | 0 (0) | 44 (100) |
| Prostatectomy GS (%) | ||||
| ≤6 | 41 (35) | 41 (100) | 0 (0) | 0 (0) |
| 3 + 4 = 7 | 45 (48) | 0 (0) | 27 (84) | 18 (41) |
| 4 + 3 = 7 | 21 (18) | 0 (0) | 2 (6) | 19 (43) |
| ≥8 | 10 (9) | 0 (0) | 3 (10) | 7 (16) |
| N-glycans Total | # Features | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glycan label | GU | Glycans | S0 | S1 | S2 | S3 | S4 | G0 | G1 | G2 | G3 | G4 | A1 | A2 | A3 | A4 | High M | CoreF | Outer arm F |
| GP1 | 4.85 | A1 | |||||||||||||||||
| GP2 | 5.34 | M4 | |||||||||||||||||
| 5.41 | FA1 | ||||||||||||||||||
| 5.41 | A2 | ||||||||||||||||||
| GP3 | 5.61 | A1(6)G1 | |||||||||||||||||
| GP4 | 5.78 | A2B | |||||||||||||||||
| 5.78 | A1(3)G1 | ||||||||||||||||||
| GP5 | 5.88 | FA2 | |||||||||||||||||
| GP6 | 6.18 | M5 | 50% | 50% | 50% | 50% | |||||||||||||
| 6.24 | FA2B | ||||||||||||||||||
| 6.24 | FA1(6)G1 | ||||||||||||||||||
| 6.24 | A2(6)G1 | ||||||||||||||||||
| 6.38 | FA1(3)G1 | ||||||||||||||||||
| 6.38 | A2(3)G1 | ||||||||||||||||||
| GP7 | 6.55 | A2(6)BG1 | |||||||||||||||||
| GP8 | 6.71 | A2(3)BG1 | |||||||||||||||||
| 6.71 | FA2(6)G1 | ||||||||||||||||||
| 6.71 | M4A1G1 | ||||||||||||||||||
| GP9 | 6.84 | FA2(3)G1 | |||||||||||||||||
| GP10 | 6.95 | FA2(6)BG1 | |||||||||||||||||
| GP11 | 7.08 | FA2(3)BG1 | |||||||||||||||||
| 7.08 | M6 D1,D2 | ||||||||||||||||||
| 7.08 | M6 D3 | ||||||||||||||||||
| GP12 | 7.20 | A1(3)G1S(3)1 | 50% | 50% | 50% | 50% | 50% | 50% | |||||||||||
| 7.20 | A2G2 | ||||||||||||||||||
| GP13 | 7.38 | A1(3)G1S(6)1 | |||||||||||||||||
| GP14 | 7.38 | A2BG2 | |||||||||||||||||
| GP15 | 7.62 | A2(3)G1S(3)1 | 50% | 50% | 50% | ||||||||||||||
| 7.62 | A2(3)G1S(6)1 | ||||||||||||||||||
| 7.62 | FA1G1S(3)1 | ||||||||||||||||||
| 7.62 | FA1G1S(6)1 | ||||||||||||||||||
| GP16 | 7.62 | M5A1G1 | |||||||||||||||||
| 7.62 | FA2G2 | ||||||||||||||||||
| GP17 | 7.76 | FA2BG2 | |||||||||||||||||
| 7.76 | M7 D3 | ||||||||||||||||||
| 7.76 | A2(6)BG1S(3)1 | ||||||||||||||||||
| 7.76 | A2(6)BG1S(6)1 | ||||||||||||||||||
| GP18 | 7.92 | A2(3)BG1S(3)1 | |||||||||||||||||
| 7.92 | A2(3)BG1S(6)1 | ||||||||||||||||||
| 7.92 | FA2(6)G1S(3)1 | ||||||||||||||||||
| 7.92 | FA2(6)G1S(6)1 | ||||||||||||||||||
| 7.92 | M4A1G1S(3)1 | ||||||||||||||||||
| 7.92 | M4A1G1S(6)1 | ||||||||||||||||||
| 7.92 | M7 D1 | ||||||||||||||||||
| GP19 | 8.03 | FA2(3)G1S(3)1 | |||||||||||||||||
| 8.03 | FA2(3)G1S(6)1 | ||||||||||||||||||
| 8.03 | FA2(6)BG1S(3)1 | ||||||||||||||||||
| 8.03 | FA2(6)BG1S(6)1 | ||||||||||||||||||
| 8.20 | FA2(3)BG1S(3)1 | ||||||||||||||||||
| 8.20 | FA2(3)BG1S(6)1 | ||||||||||||||||||
| GP20 | 8.38 | A2G2S(3)1 | |||||||||||||||||
| 8.38 | A2G2S(6)1 | ||||||||||||||||||
| 8.38 | A3G3 | ||||||||||||||||||
| GP21 | 8.53 | A2BG2S(3)1 | |||||||||||||||||
| 8.53 | A2BG2S(6)1 | ||||||||||||||||||
| GP22 | 8.63 | M5A1G1S(3)1 | 50% | 50% | 50% | 50% | |||||||||||||
| 8.63 | M5A1G1S(6)1 | ||||||||||||||||||
| 8.63 | FA3G3 | ||||||||||||||||||
| 8.63 | M8 D2,D3 | ||||||||||||||||||
| 8.63 | * A2G2S(3)1 | ||||||||||||||||||
| GP23 | 8.80 | FA2G2S(3)1 | |||||||||||||||||
| 8.80 | FA2G2S(6)1 | ||||||||||||||||||
| GP24 | 8.80 | M8 D1,D3 | |||||||||||||||||
| GP25 | 9.02 | FA2BG2S(3)1 | |||||||||||||||||
| 9.02 | FA2BG2S(6)1 | ||||||||||||||||||
| GP26 | 9.21 | * A2G2S(3,3)2 | |||||||||||||||||
| 9.21 | * A2G2S(3,6)2 | ||||||||||||||||||
| 9.21 | * FA2G2S(3,3)2 | ||||||||||||||||||
| GP27 | 9.21 | A2F1G2S(3)1 | |||||||||||||||||
| 9.21 | A2F1G2S(6)1 | ||||||||||||||||||
| GP28 | 9.43 | A3G3S(3)1 | |||||||||||||||||
| 9.43 | A3G3S(6)1 | ||||||||||||||||||
| 9.43 | M9 | ||||||||||||||||||
| 9.62 | A3BG3S(3)1 | ||||||||||||||||||
| 9.62 | A3BG3S(6)1 | ||||||||||||||||||
| 9.62 | A2G2S(3,3)2 | ||||||||||||||||||
| 9.62 | A2G2S(3,6)2 | ||||||||||||||||||
| 9.62 | A2G2S(6,6)2 | ||||||||||||||||||
| GP29 | 9.79 | FA3G3S(3)1 | |||||||||||||||||
| 9.79 | FA3G3S(6)1 | ||||||||||||||||||
| 9.79 | FA3BG3S(3)1 | ||||||||||||||||||
| 9.79 | A2BG2S(3,3)2 | ||||||||||||||||||
| 9.79 | A2BG2S(3,6)2 | ||||||||||||||||||
| 9.79 | A2BG2S(6,6)2 | ||||||||||||||||||
| GP30 | 10.04 | A3F1G3S(3)1 | |||||||||||||||||
| 10.04 | FA2G2S(3,3)2 | ||||||||||||||||||
| 10.04 | FA2G2S(3,6)2 | ||||||||||||||||||
| 10.04 | FA2G2S(6,6)2 | ||||||||||||||||||
| GP31 | 10.17 | FA2BG2S(3,3)2 | |||||||||||||||||
| 10.17 | FA2BG2S(3,6)2 | ||||||||||||||||||
| 10.17 | FA2BG2S(6,6)2 | ||||||||||||||||||
| 10.17 | A2F1G2S(3,3)2 | ||||||||||||||||||
| 10.17 | A2F1G2S(3,6)2 | ||||||||||||||||||
| 10.17 | A2F1G2S(6,6)2 | ||||||||||||||||||
| 10.17 | M9Glc | ||||||||||||||||||
| GP32 | 10.31 | A3G3S(3,3)2 | |||||||||||||||||
| 10.31 | A3G3S(3,6)2 | ||||||||||||||||||
| 10.31 | A3G3S(6,6)2 | ||||||||||||||||||
| GP33 | 10.43 | A3BG3S(3,3)2 | |||||||||||||||||
| 10.43 | A3BG3S(3,6)2 | ||||||||||||||||||
| 10.43 | A3BG3S(6,6)2 | ||||||||||||||||||
| GP34 | 10.60 | A4G4S(3)1 | |||||||||||||||||
| GP35 | 10.77 | FA3G3S(3,3)2 | |||||||||||||||||
| 10.77 | FA3G3S(3,6)2 | ||||||||||||||||||
| 10.77 | FA3G3S(6,6)2 | ||||||||||||||||||
| 10.77 | * A3G3S(3,3)2 | ||||||||||||||||||
| GP36 | 10.96 | * A3G3S(3,3,3)3 | |||||||||||||||||
| 10.96 | A3F1G3S(3,3)2 | ||||||||||||||||||
| 10.96 | A4G4S(6)1 | ||||||||||||||||||
| GP37 | 11.14 | * A3G3S(3,3,6)3 | |||||||||||||||||
| 11.14 | * A3G3S(3,6,6)3 | ||||||||||||||||||
| 11.14 | * A3BG3S(3,3,3)3 | ||||||||||||||||||
| 11.14 | * A3BG3S(3,3,6)3 | ||||||||||||||||||
| 11.14 | * A3BG3S(3,6,6)3 | ||||||||||||||||||
| GP38 | 11.28 | * FA3G3S(3,3,3)3 | |||||||||||||||||
| 11.28 | * FA3BG3S(3,3,3)3 | ||||||||||||||||||
| GP39 | 11.54 | A4G4S(3,6)2 | |||||||||||||||||
| 11.54 | A3G3S(3,3,3)3 | ||||||||||||||||||
| 11.54 | A3G3S(3,3,6)3 | ||||||||||||||||||
| 11.54 | A3G3S(3,6,6)3 | ||||||||||||||||||
| 11.54 | A3G3S(6,6,6)3 | ||||||||||||||||||
| 11.72 | A3BG3S(3,3,3)3 | ||||||||||||||||||
| 11.72 | A3BG3S(3,3,6)3 | ||||||||||||||||||
| 11.72 | A3BG3S(6,6,6)3 | ||||||||||||||||||
| GP40 | 11.89 | FA3G3S(3,3,3)3 | |||||||||||||||||
| 11.89 | FA3G3S(3,3,6)3 | ||||||||||||||||||
| 11.89 | FA3G3S(3,6,6)3 | ||||||||||||||||||
| 11.89 | FA3G3S(6,6,6)3 | ||||||||||||||||||
| GP41 | 12.03 | * A3G3S(3,3,3)3 | |||||||||||||||||
| 12.03 | * A3G3S(3,3,6)3 | ||||||||||||||||||
| 12.03 | * A3G3S(3,6,6)3 | ||||||||||||||||||
| 12.03 | A3F1G3S(3,3,3)3 | ||||||||||||||||||
| 12.03 | A3F1G3S(3,3,6)3 | ||||||||||||||||||
| 12.03 | FA3BG3S(3,3,3)3 | ||||||||||||||||||
| 12.03 | FA3BG3S(6,6,6)3 | ||||||||||||||||||
| GP42 | 12.15 | A4G4S(3,3,3)3 | |||||||||||||||||
| 12.15 | A3F1G3S(3,6,6)3 | ||||||||||||||||||
| GP43 | 12.33 | A4G4S(3,3,6)3 | |||||||||||||||||
| 12.33 | A4G4S(3,6,6)3 | ||||||||||||||||||
| GP44 | 12.48 | A4F1G3S(3,3,3)3 | |||||||||||||||||
| 12.48 | A3F2G3S(3,3,3)3 | ||||||||||||||||||
| 12.48 | A4F1G3S(3,3,6)3 | ||||||||||||||||||
| 12.48 | A4F1G3S(3,6,6)3 | ||||||||||||||||||
| GP45 | 12.67 | A3F2G3S(3,3,6)3 | 33% | ||||||||||||||||
| 12.67 | A4F2G3S(3,3,3)3 | ||||||||||||||||||
| 12.67 | A4F2G3S(3,3,6)3 | ||||||||||||||||||
| 12.67 | * A4G4S(3,3,3,3)4 | ||||||||||||||||||
| 12.78 | A4G4S(3,3,3,3)4 | ||||||||||||||||||
| GP46 | 12.96 | A4G4S(3,3,3,6)4 | |||||||||||||||||
| GP47 | 13.27 | * A4G4S(3,3,3,6)4 | |||||||||||||||||
| 13.27 | A4G4S(3,3,6,6)4 | ||||||||||||||||||
| 13.27 | A4G4S(3,6,6,6)4 | ||||||||||||||||||
| GP48 | 13.47 | * A4G4S(3,3,3,3)4 | 50% | ||||||||||||||||
| 13.47 | FA4G4S(3,3,3,3)4 | ||||||||||||||||||
| 13.47 | FA4G4S(3,3,3,6)4 | ||||||||||||||||||
| 13.47 | A4BG4S(3,3,6,6)4 | ||||||||||||||||||
| GP49 | 13.82 | A4F1G4S(3,3,3,3)4 | |||||||||||||||||
| 13.82 | A4F1G4S(3,3,3,6)4 | ||||||||||||||||||
| 13.82 | A4F1G4S(3,3,6,6)4 | ||||||||||||||||||
| 13.82 | A4F1G4S(3,6,6,6)4 | ||||||||||||||||||
| GP50 | 13.99 | A4G4LacS(3,3,3,3)4 | 33% | ||||||||||||||||
| 13.99 | A4G4LacS(3,3,3,6)4 | ||||||||||||||||||
| 13.99 | A4F2G4S(3,3,3,3)4 | ||||||||||||||||||
| 13.99 | A4F2G4S(3,3,6,6)4 | ||||||||||||||||||
| 14.43 | A4F3G4S(3,3,3,3)4 | ||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilgunn, S.; Murphy, K.; Stöckmann, H.; Conroy, P.J.; Murphy, T.B.; Watson, R.W.; O’Kennedy, R.J.; Rudd, P.M.; Saldova, R. Glycosylation in Indolent, Significant and Aggressive Prostate Cancer by Automated High-Throughput N-Glycan Profiling. Int. J. Mol. Sci. 2020, 21, 9233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239233
Gilgunn S, Murphy K, Stöckmann H, Conroy PJ, Murphy TB, Watson RW, O’Kennedy RJ, Rudd PM, Saldova R. Glycosylation in Indolent, Significant and Aggressive Prostate Cancer by Automated High-Throughput N-Glycan Profiling. International Journal of Molecular Sciences. 2020; 21(23):9233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239233
Chicago/Turabian StyleGilgunn, Sarah, Keefe Murphy, Henning Stöckmann, Paul J. Conroy, T. Brendan Murphy, R. William Watson, Richard J. O’Kennedy, Pauline M. Rudd, and Radka Saldova. 2020. "Glycosylation in Indolent, Significant and Aggressive Prostate Cancer by Automated High-Throughput N-Glycan Profiling" International Journal of Molecular Sciences 21, no. 23: 9233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239233