Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells

 , , ,
, , , 
Abstract
:1. Introduction
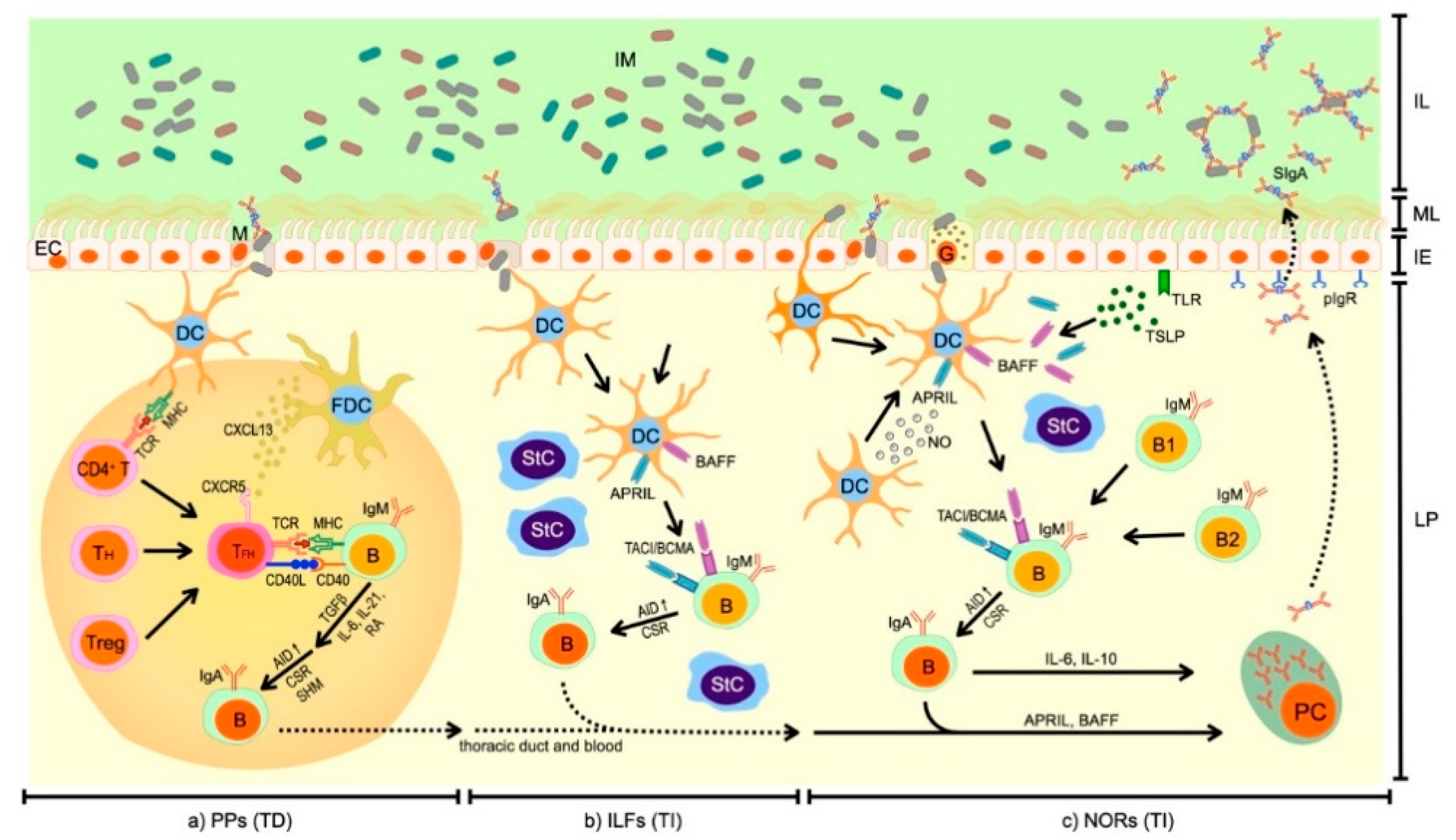
2. Synthesis of SIgA in the Intestines
2.1. T Cell-Dependent Synthesis of SIgA
2.2. T Cell-Independent Synthesis of SIgA
2.3. Migration of IgA+ B Cells and Differentiation into IgA+ PCs in LP
3. Functions of SIgA in Gut Mucosal Secretions
3.1. Defense against Pathogens
3.2. Regulating Gut Microbiota Composition, Gut Immune Homeostasis and Oral Tolerance
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schroeder, H.W.J.; Cavacini, L. Structure and Function of Immunoglobulins (author manuscript). J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [Green Version]
- Woof, J.M.; Ken, M.A. The function of immunoglobulin A in immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woof, J.M.; Kerr, M.A. IgA function—Variations on a theme. Immunology 2004, 113, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Fagarasan, S.; Honjo, T. Intestinal IgA synthesis: Regulation of front-line body defences. Nat. Rev. Immunol. 2003, 3, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Woof, J.M.; Russell, M.W. Structure and function relationships in IgA. Mucosal Immunol. 2011, 4, 590–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbons, D.L.; Spencer, J. Mouse and human intestinal immunity: Same ballpark, different players; Different rules, same score. Mucosal Immunol. 2011, 4, 148–157. [Google Scholar] [CrossRef]
- Sánchez-Salguero, E.; Mondragón-Ramírez, G.K.; Alcántara-Montiel, J.C.; Cérbulo-Vázquez, A.; Villegas-Domínguez, X.; Contreras-Vargas, V.M.; del Thompson-Bonilla, M.R.; Romero-Ramírez, H.; Santos-Argumedo, L. Infectious episodes during pregnancy, at particular mucosal sites, increase specific IgA1 or IgA2 subtype levels in human colostrum. Matern. Health Neonatol. Perinatol. 2019, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Fagarasan, S. How host-bacterial interactions lead to IgA synthesis in the gut. Trends Immunol. 2008, 29, 523–531. [Google Scholar] [CrossRef]
- Cerutti, A.; Rescigno, M. The Biology of Intestinal Immunoglobulin A Responses. Immunity 2008, 28, 740–750. [Google Scholar] [CrossRef] [Green Version]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.; Reese, T.A. Making Mouse Models That Reflect Human Immune Responses. Trends Immunol. 2017, 38, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beura, L.K.; Hamilton, S.E.; Bi, K.; Schenkel, J.M.; Odumade, O.A.; Casey, K.A.; Thompson, E.A.; Fraser, K.A.; Rosato, P.C.; Filali-Mouhim, A.; et al. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 2016, 532, 512–516. [Google Scholar] [CrossRef]
- Reese, T.A.; Bi, K.; Kambal, A.; Filali-Mouhim, A.; Beura, L.K.; Bürger, M.C.; Pulendran, B.; Sekaly, R.P.; Jameson, S.C.; Masopust, D.; et al. Sequential Infection with Common Pathogens Promotes Human-like Immune Gene Expression and Altered Vaccine Response. Cell Host Microbe 2016, 19, 713–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, K.J.; Crooks, G.M. Immune-Cell Lineage Commitment: Translation from Mice to Humans. Immunity 2007, 26, 674–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversen, R.; Snir, O.; Stensland, M.; Kroll, J.E.; Steinsbø, Ø.; Korponay-Szabó, I.R.; Lundin, K.E.A.; de Souza, G.A.; Sollid, L.M. Strong Clonal Relatedness between Serum and Gut IgA despite Different Plasma Cell Origins. Cell Rep. 2017, 20, 2357–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutteh, W.H.; Prince, S.J.; Mestecky, J. Tissue origins of human polymeric and monomeric IgA. J. Immunol. 1982, 128, 990–995. [Google Scholar]
- Bunker, J.J.; Flynn, T.M.; Koval, J.C.; Shaw, D.G.; Meisel, M.; McDonald, B.D.; Ishizuka, I.E.; Dent, A.L.; Wilson, P.C.; Jabri, B.; et al. Innate and Adaptive Humoral Responses Coat Distinct Commensal Bacteria with Immunoglobulin A. Immunity 2015, 43, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Cerutti, A.; Chen, K.; Chorny, A. Immunoglobulin Responses at the Mucosal Interface. Annu. Rev. Immunol. 2011, 29, 273–293. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Nakajima, A. New aspects of IgA synthesis in the gut. Int. Immunol. 2014, 26, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Nochi, T.; Denton, P.W.; Wahl, A.; Garcia, J.V. Cryptopatches Are Essential for the Development of Human GALT. Cell Rep. 2013, 3, 1874–1884. [Google Scholar] [CrossRef] [Green Version]
- Spencer, J.; Sollid, L.M. The human intestinal B-cell response. Mucosal Immunol. 2016, 9, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Fagarasan, S. Diverse regulatory pathways for IgA synthesis in the gut. Mucosal Immunol. 2009, 2, 468–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tezuka, H.; Ohteki, T. Regulation of IgA production by intestinal dendritic cells and related cells. Front. Immunol. 2019, 10, 1891. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.B.; Fagarasan, S. IgA synthesis: A form of functional immune adaptation extending beyond gut. Curr. Opin. Immunol. 2012, 24, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Fagarasan, S.; Muramatsu, M.; Suzuki, K.; Nagaoka, H.; Hiai, H.; Honjo, T. Critical roles of activation-induced cytidine deaminase in the homeostasis of gut flora. Science 2002, 298, 1424–1427. [Google Scholar] [CrossRef] [PubMed]
- Fagarasan, S.; Kawamoto, S.; Kanagawa, O.; Suzuki, K. Adaptive Immune Regulation in the Gut: T Cell–Dependent and T Cell–Independent IgA Synthesis. Annu. Rev. Immunol. 2010, 28, 243–273. [Google Scholar] [CrossRef] [PubMed]
- Reboldi, A.; Cyster, J.G. Peyer’s patches: Organizing B-cell responses at the intestinal frontier. Immunol. Rev. 2016, 271, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Waseem, T.C.; Sahoo, A.; Bieerkehazhi, S.; Zhou, H.; Galkina, E.V.; Nurieva, R. Insights into the molecular mechanisms of T follicular helper-mediated immunity and pathology. Front. Immunol. 2018, 9, 1884. [Google Scholar] [CrossRef]
- Bergqvist, P.; Gärdby, E.; Stensson, A.; Bemark, M.; Lycke, N.Y. Gut IgA Class Switch Recombination in the Absence of CD40 Does Not Occur in the Lamina Propria and Is Independent of Germinal Centers. J. Immunol. 2006, 177, 7772–7783. [Google Scholar] [CrossRef] [Green Version]
- Berkowska, M.A.; Driessen, G.J.A.; Bikos, V.; Grosserichter-Wagener, C.; Stamatopoulos, K.; Cerutti, A.; He, B.; Biermann, K.; Lange, J.F.; Van Der Burg, M.; et al. Human memory B cells originate from three distinct germinal center-dependent and -independent maturation pathways. Blood 2011, 118, 2150–2158. [Google Scholar] [CrossRef] [Green Version]
- Kiss, E.A.; Vonarbourg, C.; Kopfmann, S.; Hobeika, E.; Finke, D.; Esser, C.; Diefenbach, A. Natural aryl hydrocarbon receptor ligands control organogenesis of intestinal lymphoid follicles. Science 2011, 334, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Tamayo, E.; Alvarez, P.; Merino, R. TGFβ superfamily members as regulators of B cell development and function—implications for autoimmunity. Int. J. Mol. Sci. 2018, 19, 3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castigli, E.; Scott, S.; Dedeoglu, F.; Bryce, P.; Jabara, H.; Bhan, A.K.; Mizoguchi, E.; Geha, R.S. Impaired IgA class switching inn APRIL-deficient mice. Proc. Natl. Acad. Sci. USA 2004, 101, 3903–3908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Bülow, G.U.; Van Deursen, J.M.; Bram, R.J. Regulation of the T-independent humoral response by TACI. Immunity 2001, 14, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Castigli, E.; Wilson, S.A.; Garibyan, L.; Rachid, R.; Bonilla, F.; Schneider, L.; Geha, R.S. TACI is mutant in common variable immunodeficiency and IgA deficiency. Nat. Genet. 2005, 37, 829–834. [Google Scholar] [CrossRef]
- Gutzeit, C.; Magri, G.; Cerutti, A. Intestinal IgA production and its role in host-microbe interaction. Immunol. Rev. 2014, 260, 76–85. [Google Scholar] [CrossRef]
- Xiong, N.; Hu, S. Regulation of intestinal IgA responses. Cell. Mol. Life Sci. 2015, 72, 2645–2655. [Google Scholar] [CrossRef]
- Arnold, J.N.; Wormald, M.R.; Sim, R.B.; Rudd, P.M.; Dwek, R.A. The impact of glycosylation on the biological function and structure of human immunoglobulins. Annu. Rev. Immunol. 2007, 25, 21–50. [Google Scholar] [CrossRef]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Melo-Gonzalez, F.; Kammoun, H.; Evren, E.; Dutton, E.E.; Papadopoulou, M.; Bradford, B.M.; Tanes, C.; Fardus-Reid, F.; Swann, J.R.; Bittinger, K.; et al. Antigen-presenting ILC3 regulate T cell-dependent IgA responses to colonic mucosal bacteria. J. Exp. Med. 2019, 216, 728–742. [Google Scholar] [CrossRef]
- Bollinger, R.R.; Everett, M.L.; Palestrant, D.; Love, S.D.; Lin, S.S.; Parker, W. Human secretory immunoglobulin A may contribute to biofilm formation in the gut. Immunology 2003, 109, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, P.; Ding, H.; Canales-Herrerias, P.; Pasrich, P.J.; Sonnenburg, J.L.; Peterson, D.A. Intestinal IgA regulates expression of a fructan polysaccharide utilization locus in colonizing gut commensal Bacteroides thetaiotaomicron. MBio 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrier, C.; Sprenger, N.; Corthésy, B. Glycans on secretory component participate in innate protection against mucosal pathogens. J. Biol. Chem. 2006, 281, 14280–14287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uren, T.K.; Wijburg, O.L.C.; Simmons, C.; Johansen, F.E.; Brandtzaeg, P.; Strugnell, R.A. Vaccine-induced protection against gastrointestinal bacterial infections in the absence of secretory antibodies. Eur. J. Immunol. 2005, 35, 180–188. [Google Scholar] [CrossRef]
- Forbes, S.J.; Bumpus, T.; McCarthy, E.A.; Corthésy, B.; Mantis, N.J. Transient suppression of shigella flexneri type 3 secretion by a protective O-antigen-specific monoclonal igA. MBio 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Pabst, O.; Slack, E. IgA and the intestinal microbiota: The importance of being specific. Mucosal Immunol. 2020, 13, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Lycke, N.Y.; Bemark, M. The regulation of gut mucosal IgA B-cell responses: Recent developments. Mucosal Immunol. 2017, 10, 1361–1374. [Google Scholar] [CrossRef]
- Benckert, J.; Schmolka, N.; Kreschel, C.; Zoller, M.J.; Sturm, A.; Wiedenmann, B.; Wardemann, H. The majority of intestinal IgA+ and IgG+ plasmablasts in the human gut are antigen-specific. J. Clin. Investig. 2011, 121, 1946–1955. [Google Scholar] [CrossRef] [Green Version]
- Tsui, F.P.; Egan, W.; Summers, M.F.; Andrew Byrd, R.; Schneerson, R.; Robbins, J.B. Determination of the structure of the Escherichia coli K100 capsular polysaccharide, cross-reactive with the capsule from type b Haemophilus influenzae. Carbohydr. Res. 1988, 173, 65–74. [Google Scholar] [CrossRef]
- Crivelli, C.; Demarta, A.; Peduzzi, R. Intestinal secretory immunoglobulin A (sIgA) response to Aeromonas exoproteins in patients with naturally acquired Aeromonas diarrhea. FEMS Immunol. Med. Microbiol. 2001, 30, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Winsor, D.K.; Mathewson, J.J.; DuPont, H.L. Western blot analysis of intestinal secretory immunoglobulin a response to Campylobacter jejuni antigens in patients with naturally acquired Campylobacter enteritis. Gastroenterology 1986, 90, 1217–1222. [Google Scholar] [CrossRef]
- Winsor, D.K.; Mathewson, J.J.; Dupont, H.L.; Winsor, D.K. Comparison of serum and fecal antibody responses of patients with naturally acquired shigella sonnei infection. J. Infect. Dis. 1988, 158, 1108–1112. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Sasaki, T.; Satoh-Takayama, N.; Kanaya, T.; Kato, T.; Takikawa, Y.; Takahashi, M.; Tachibana, N.; Kim, K.S.; Surh, C.D.; et al. Dietary Antigens Induce Germinal Center Responses in Peyer’s Patches and Antigen-Specific IgA Production. Front. Immunol. 2019, 10, 2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blutt, S.E.; Miller, A.D.; Salmon, S.L.; Metzger, D.W.; Conner, M.E. IgA is important for clearance and critical for protection from rotavirus infection. Mucosal Immunol. 2012, 5, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, A.J.; Köller, Y.; McCoy, K.D. The bilateral responsiveness between intestinal microbes and IgA. Trends Immunol. 2015, 36, 460–470. [Google Scholar] [CrossRef]
- Bowes, T.; Wagner, E.R.; Boffey, J.; Nicholl, D.; Cochrane, L.; Benboubetra, M.; Conner, J.; Furukawa, K.; Furukawa, K.; Willison, H.J. Tolerance to self gangliosides is the major factor restricting the antibody response to lipopolysaccharide core oligosaccharides in Campylobacter jejuni strains associated with Guillain-Barré syndrome. Infect. Immun. 2002, 70, 5008–5018. [Google Scholar] [CrossRef] [Green Version]
- Turula, H.; Wobus, C.E. The Role of the Polymeric Immunoglobulin Receptor and Secretory Immunoglobulins during Mucosal Infection and Immunity. Viruses 2018, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jin, L.; Chen, T.; Pirozzi, C.J. The Effects of Secretory IgA in the Mucosal Immune System. Biomed. Res. Int. 2020, 2020. [Google Scholar] [CrossRef]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut microbiota metabolism and interaction with food components. Int. J. Mol. Sci. 2020, 21, 3688. [Google Scholar] [CrossRef]
- Meffre, E.; Wardemann, H. B-cell tolerance checkpoints in health and autoimmunity. Curr. Opin. Immunol. 2008, 20, 632–638. [Google Scholar] [CrossRef]
- Wu, H.Y.; Weiner, H.L. Oral Tolerance. Immunol. Res. 2003, 28, 265–284. [Google Scholar] [CrossRef]
- Eberl, G. Immunity by equilibrium. Nat. Rev. Immunol. 2016, 16, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M.; Lopatin, U.; Chieppa, M. Interactions among dendritic cells, macrophages, and epithelial cells in the gut: Implications for immune tolerance. Curr. Opin. Immunol. 2008, 20, 669–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Sonnenberg, G.F. Essential immunologic orchestrators of intestinal homeostasis. Sci. Immunol. 2018, 3, eaao1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, A.; Durant, L.; Hoyles, L.; McCartney, A.L.; Man, R.; Segal, J.; Costello, S.P.; Hendy, P.; Reddi, D.; Bouri, S.; et al. Deficient Resident Memory T Cell and CD8 T Cell Response to Commensals in Inflammatory Bowel Disease. J. Crohn’s Colitis 2020, 14, 525–537. [Google Scholar] [CrossRef]
- Idzko, M.; Hammad, H.; Van Nimwegen, M.; Kool, M.; Willart, M.A.M.; Muskens, F.; Hoogsteden, H.C.; Luttmann, W.; Ferrari, D.; Di Virgilio, F.; et al. Extracellular ATP triggers and maintains asthmatic airway inflammation by activating dendritic cells. Nat. Med. 2007, 13, 913–919. [Google Scholar] [CrossRef]
- Mandapathil, M.; Hilldorfer, B.; Szczepanski, M.J.; Czystowska, M.; Szajnik, M.; Ren, J.; Lang, S.; Jackson, E.K.; Gorelik, E.; Whiteside, T.L. Generation and accumulation of immunosuppressive adenosine by human CD4+CD25highFOXP3+ regulatory T Cells. J. Biol. Chem. 2010, 285, 7176–7186. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhao, W.; Wang, Y.; Jin, L.; Jin, G.; Sun, X.; Wang, W.; Wang, K.; Xu, X.; Hao, J.; et al. A wave of Foxp3+ regulatory T cell accumulation in the neonatal liver plays unique roles in maintaining self-tolerance. Cell. Mol. Immunol. 2020, 17, 507–518. [Google Scholar] [CrossRef]
- Bos, N.A.; Kimura, H.; Meeuwsen, C.G.; De Visser, H.; Hazenberg, M.P.; Wostmann, B.S.; Pleasants, J.R.; Benner, R.; Marcus, D.M. Serum immunoglobulin levels and naturally occurring antibodies against carbohydrate antigens in germ-free BALB/c mice fed chemically defined ultrafiltered diet. Eur. J. Immunol. 1989, 19, 2335–2339. [Google Scholar] [CrossRef]
- Yang, C.; Mogno, I.; Contijoch, E.; Borgerding, J.; Aggarwala, V.; Li, Z.; Grasset, E.; Helmus, D.; Dubinsky, M.; Mehandru, S.; et al. Strain-level differences in gut microbiome composition determine fecal IgA levels and are modifiable by gut microbiota manipulation. bioRxiv 2019, 544015. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Shinkura, R.; Doi, Y.; Maruya, M.; Fagarasan, S.; Honjo, T. Mice carrying a knock-in mutation of Aicda resulting in a defect in somatic hypermutation have impaired gut homeostasis and compromised mucosal defense. Nat. Immunol. 2011, 12, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, S.; Tran, T.H.; Maruya, M.; Suzuki, K.; Doi, Y.; Tsutsui, Y.; Kato, L.M.; Fagarasan, S. The inhibitory receptor PD-1 regulates IgA selection and bacterial composition in the gut. Science 2012, 336, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.I.; Pedron, T.; Tournebize, R.; Olivo-Marin, J.C.; Sansonetti, P.J.; Phalipon, A. Anti-inflammatory role for intracellular dimeric immunoglobulin A by neutralization of lipopolysaccharide in epithelial cells. Immunity 2003, 18, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Gratz, I.K.; Campbell, D.J. Organ-specific and memory Treg cells: Specificity, development, function, and maintenance. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuczma, M.P.; Szurek, E.A.; Cebula, A.; Chassaing, B.; Jung, Y.J.; Kang, S.M.; Fox, J.G.; Stecher, B.; Ignatowicz, L. Commensal epitopes drive differentiation of colonic Tregs. Sci. Adv. 2020, 6, eaaz3186. [Google Scholar] [CrossRef] [Green Version]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic T reg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; Van Der Veeken, J.; Deroos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.C.A.M.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur. J. Pharmacol. 2018, 831, 52–59. [Google Scholar] [CrossRef]
- Kelly, D.; Campbell, J.I.; King, T.P.; Grant, G.; Jansson, E.A.; Coutts, A.G.P.; Pettersson, S.; Conway, S. Commensal anaerobic gut bacteria attenuate inflammation by regulating nuclear-cytoplasmic shutting of PPAR-γ and ReIA. Nat. Immunol. 2004, 5, 104–112. [Google Scholar] [CrossRef]
- Kelly, D.; Conway, S.; Aminov, R. Commensal gut bacteria: Mechanisms of immune modulation. Trends Immunol. 2005, 26, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.A.; McNulty, N.P.; Guruge, J.L.; Gordon, J.I. IgA Response to Symbiotic Bacteria as a Mediator of Gut Homeostasis. Cell Host Microbe 2007, 2, 328–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, D.A.; Planer, J.D.; Guruge, J.L.; Xue, L.; Downey-Virgin, W.; Goodman, A.L.; Seedorf, H.; Gordon, J.I. Characterizing the interactions between a naturally primed immunoglobulin a and its conserved Bacteroides thetaiotaomicron species-specific epitope in gnotobiotic mice. J. Biol. Chem. 2015, 290, 12630–12649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, D.B.; Suzuki, K.; Fagarasan, S. Fostering of advanced mutualism with gut microbiota by Immunoglobulin A. Immunol. Rev. 2016, 270, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Reboldi, A.; Arnon, T.I.; Rodda, L.B.; Atakilit, A.; Sheppard, D.; Cyster, J.G. Mucosal immunology: IgA production requires B cell interaction with subepithelial dendritic cells in Peyer’s patches. Science 2016, 352, aaf4822. [Google Scholar] [CrossRef] [Green Version]
- Palm, N.W.; De Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W.; et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef] [Green Version]
- Viladomiu, M.; Kivolowitz, C.; Abdulhamid, A.; Dogan, B.; Victorio, D.; Castellanos, J.G.; Woo, V.; Teng, F.; Tran, N.L.; Sczesnak, A.; et al. IgA-coated E. Coli enriched in Crohn’s disease spondyloarthritis promote TH17-dependent inflammation. Sci. Transl. Med. 2017, 9, eaaf9655. [Google Scholar] [CrossRef]
- D’Auria, G.; Peris-Bondia, F.; Džunková, M.; Mira, A.; Collado, M.C.; Latorre, A.; Moya, A. Active and secreted IgA-coated bacterial fractions from the human gut reveal an under-represented microbiota core. Sci. Rep. 2013, 3, 3515. [Google Scholar] [CrossRef] [Green Version]
- Catanzaro, J.R.; Strauss, J.D.; Bielecka, A.; Porto, A.F.; Lobo, F.M.; Urban, A.; Schofield, W.B.; Palm, N.W. IgA-deficient humans exhibit gut microbiota dysbiosis despite secretion of compensatory IgM. Sci. Rep. 2019, 9, 13574. [Google Scholar] [CrossRef]
- Mirpuri, J.; Raetz, M.; Sturge, C.R.; Wilhelm, C.L.; Benson, A.; Savani, R.C.; Hooper, L.V.; Yarovinsky, F. Proteobacteria-specific IgA regulates maturation of the intestinal microbiota. Gut Microbes 2013, 5, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Baldassarre, M.E.; Di Mauro, A.; Mastromarino, P.; Fanelli, M.; Martinelli, D.; Urbano, F.; Capobianco, D.; Laforgia, N. Administration of a multi-strain probiotic product to women in the perinatal period differentially affects the breast milk cytokine profile and may have beneficial effects on neonatal gastrointestinal functional symptoms. A randomized clinical trial. Nutrients 2016, 8, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.Z.; Zhang, Y.H.; Guo, X.H.; Zhang, H.Y.; Zhang, Y. The double-edge role of B cells in mediating antitumor T-cell immunity: Pharmacological strategies for cancer immunotherapy. Int. Immunopharmacol. 2016, 36, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, Q.; Wang, J.; Wei, F.; Yang, L.; Ren, X. A new perspective: Exploring future therapeutic strategies for cancer by understanding the dual role of B lymphocytes in tumor immunity. Int. J. Cancer 2019, 144, 2909–2917. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Function | Mechanism | Reference |
|---|---|---|
| Immune exclusion | Retention of bacteria in the intestinal lumen. | [8,18,37] |
| Niche occupancy | Binding bacteria to mucus. | [18,38,39,40,41] |
| Immobilization | Impairing mobility by targeting flagella, cell agglutination. | [18] |
| Modulation of bacterial gene expression and metabolism | Targeting bacterial surface end extracellular proteins. | [42] |
| Neutralization | Targeting adhesins and toxins. | [8,18,39,43,44,45] |
| Antigen uptake | Facilitated transcytosis. | [27,39] |
| Finding | Reference |
|---|---|
| Infections and immunizations modulate maternal SIgA at the mammary gland impacting the development and protection of the newborn. | [7] |
| Dietary antigens are essential for a normal antigen-specific IgA response to bacteria. | [53] |
| TRM and its interaction with DC contribute substantially to the B cell activity. | [65] |
| The ability and localization of SIgA induction is a strain-specific bacterial trait. | [70] |
| Commensal antigens administration may help to widen the repertoire of specific Tregs for restoration and maintenance of intestinal homeostasis. | [75] |
| The gut SIgA targeting specific bacterial antigen can control specific bacteria growth influencing microbiota composition. | [42] |
| ILC3 functions as a critical regulatory checkpoint in the generation of TD colonic SIgA and maintain tissue homeostasis and mutualism with commensal microbiota. | [40] |
| SIgA plays a critical role in controlling gut microbial community composition and maintain a diverse and stable gut microbiota. | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrzak, B.; Tomela, K.; Olejnik-Schmidt, A.; Mackiewicz, A.; Schmidt, M. Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. Int. J. Mol. Sci. 2020, 21, 9254. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239254
Pietrzak B, Tomela K, Olejnik-Schmidt A, Mackiewicz A, Schmidt M. Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. International Journal of Molecular Sciences. 2020; 21(23):9254. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239254
Chicago/Turabian StylePietrzak, Bernadeta, Katarzyna Tomela, Agnieszka Olejnik-Schmidt, Andrzej Mackiewicz, and Marcin Schmidt. 2020. "Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells" International Journal of Molecular Sciences 21, no. 23: 9254. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239254