Dihydrotanshinone, a Natural Diterpenoid, Preserves Blood-Retinal Barrier Integrity via P2X7 Receptor


 , , ,
, , , 
Abstract
:1. Introduction
2. Results
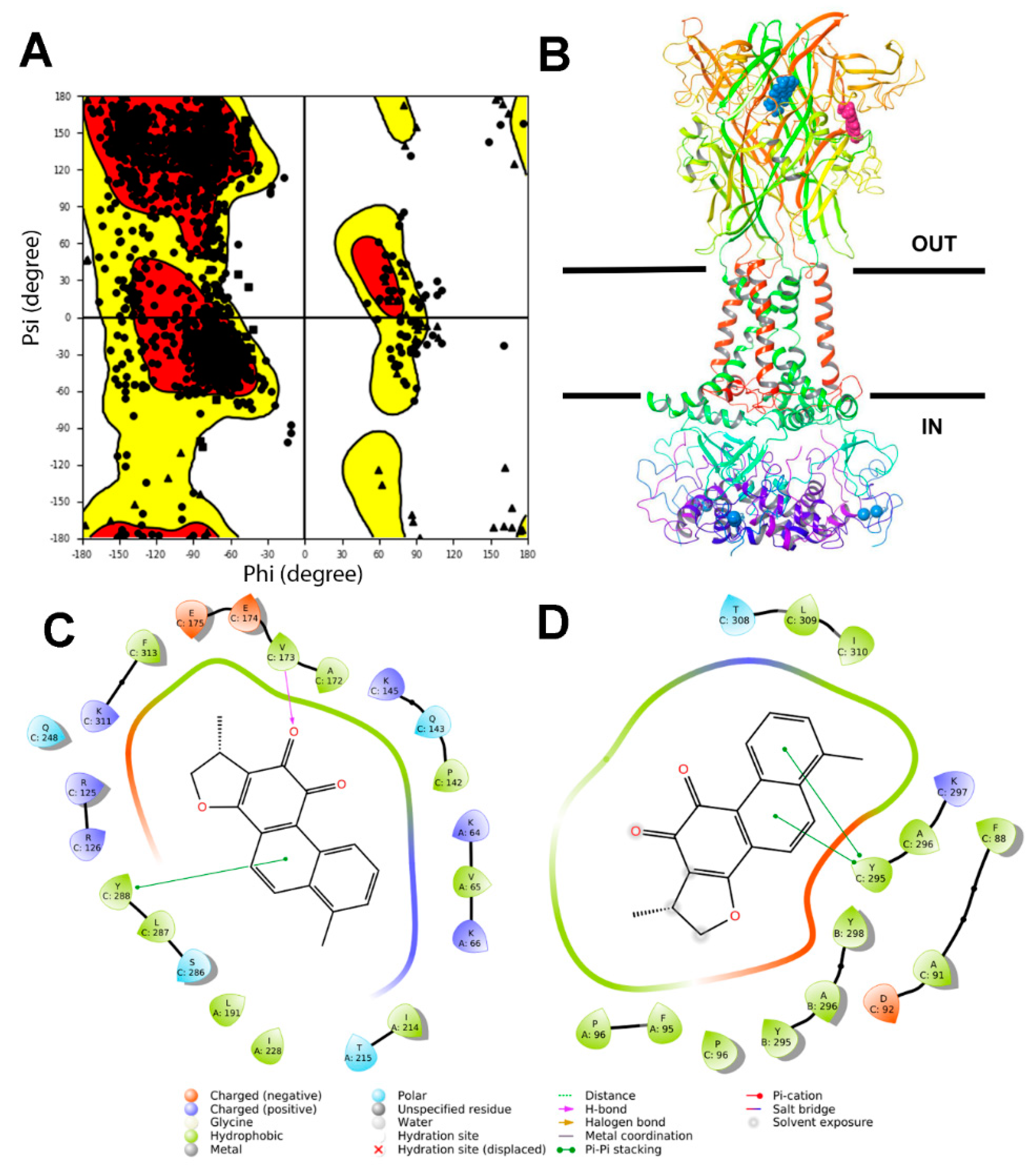
2.1. Virtual Screening at P2X7R
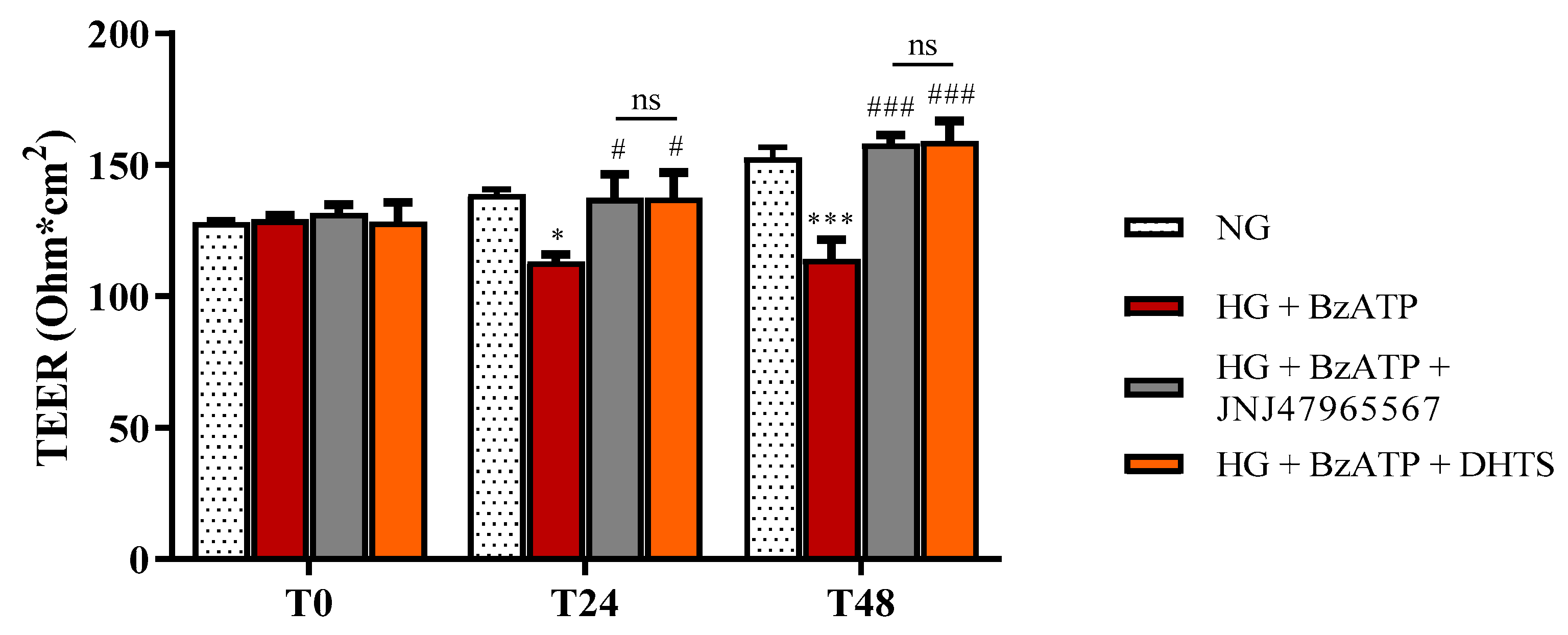
2.2. BRB Integrity
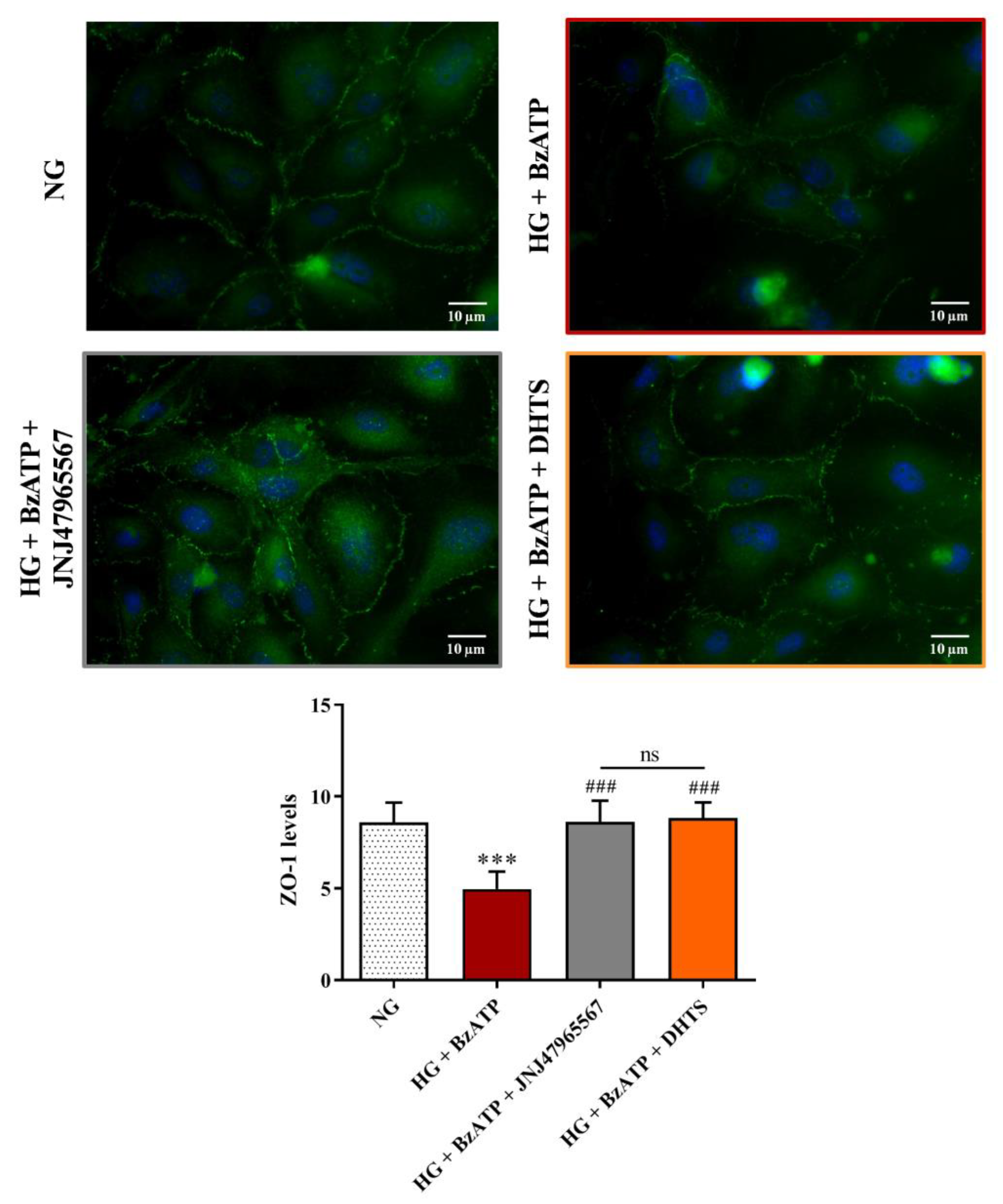
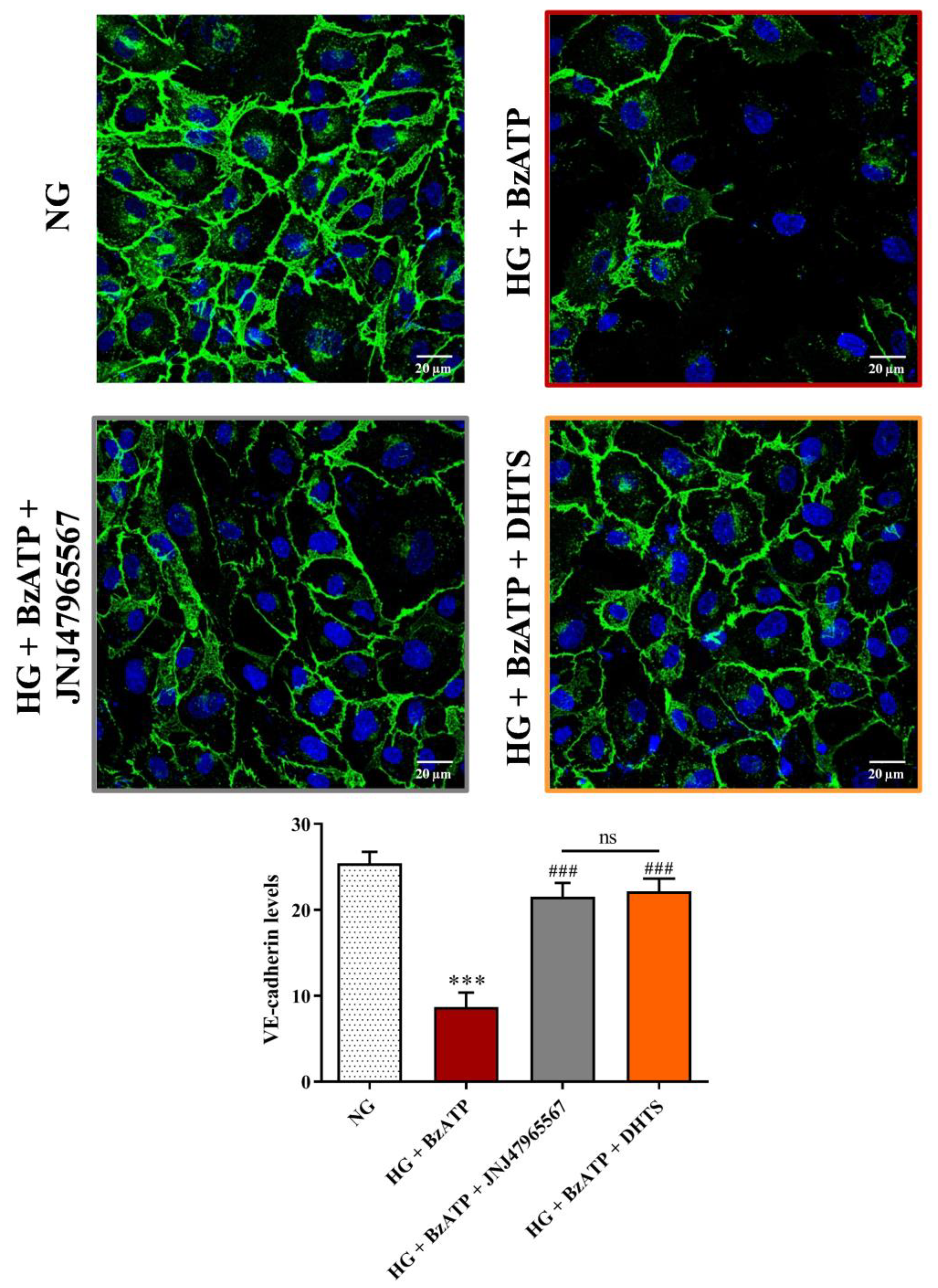
2.3. Junctional Proteins
2.4. ROS Production
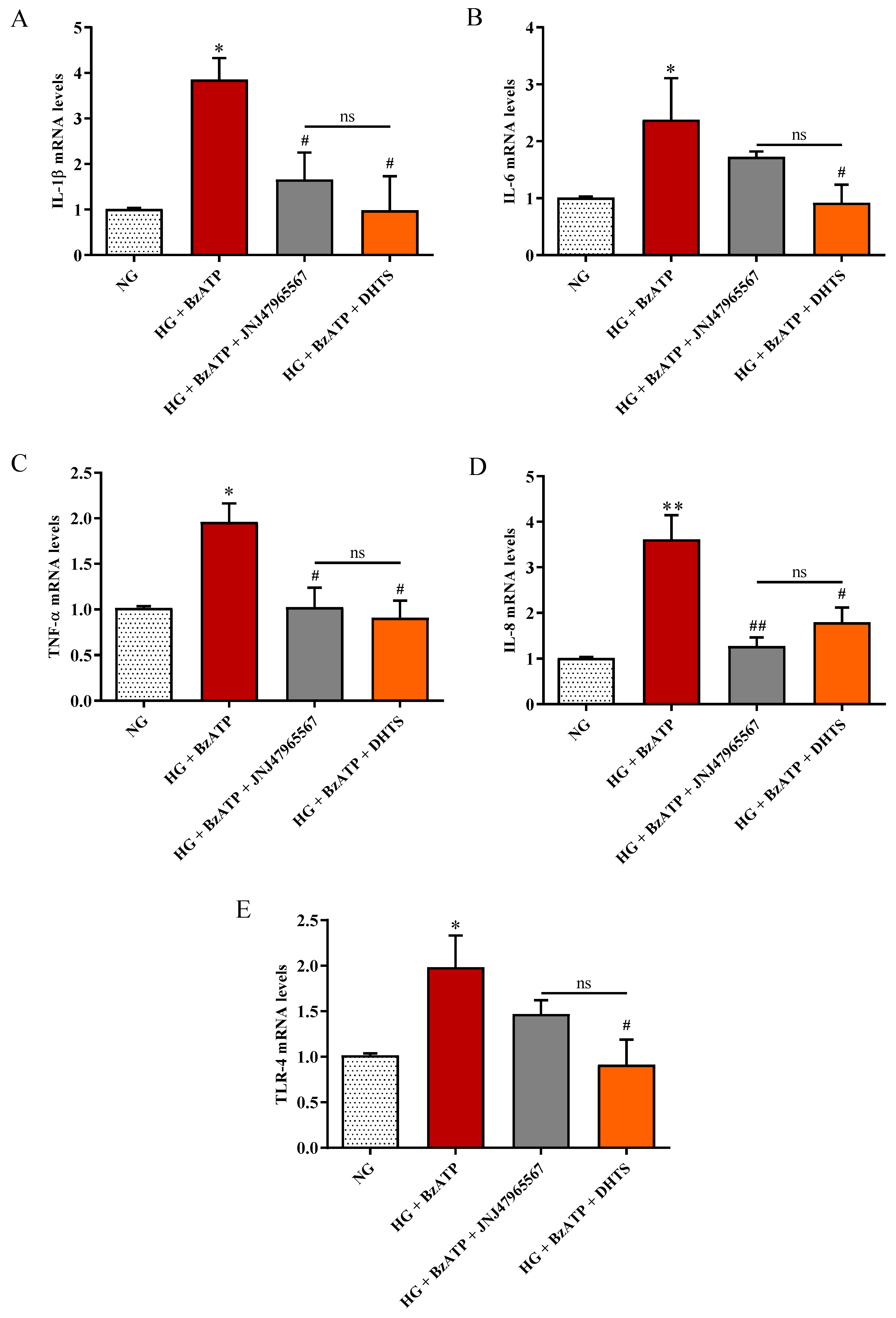
2.5. Inflammatory Biomarkers
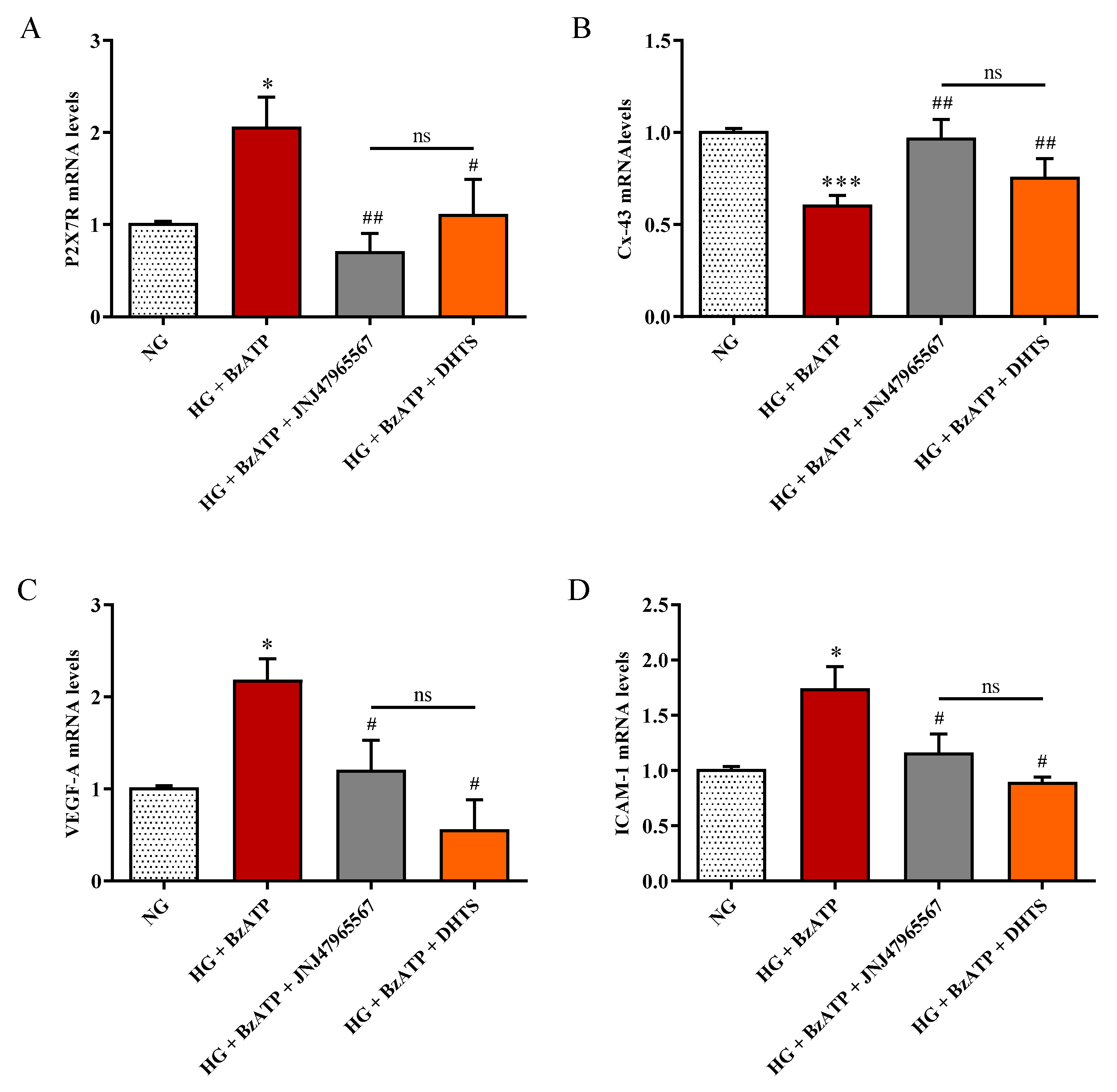
2.6. P2X7R, Cx-43, VEFG-A, and ICAM-1 Expression
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Molecular Modeling and Molecular Docking
4.3. Cell Culture Protocol and Treatment
4.4. BRB Integrity Assessment
4.5. Immunocytochemistry
4.6. Measurement of ROS Production
4.7. qRT-PCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Klein, B.E. Overview of epidemiologic studies of diabetic retinopathy. Ophthalmic Epidemiol. 2007, 14, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Ting, D.S.; Cheung, G.C.; Wong, T.Y. Diabetic retinopathy: Global prevalence, major risk factors, screening practices and public health challenges: a review. Clin. Exp. Ophthalmol. 2016, 44, 260–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.H.; Zhang, Y.; Zhang, X.M.; Li, X.R. Prevalence of diabetic retinopathy, proliferative diabetic retinopathy and non-proliferative diabetic retinopathy in asian t2dm patients: A systematic review and meta-analysis. Int. J. Ophthalmol. 2019, 12, 302–311. [Google Scholar] [PubMed]
- Mizutani, M.; Kern, T.S.; Lorenzi, M. Accelerated death of retinal microvascular cells in human and experimental diabetic retinopathy. J. Clin. Investig. 1996, 97, 2883–2890. [Google Scholar] [CrossRef]
- De Vriese, A.S.; Verbeuren, T.J.; Van de Voorde, J.; Lameire, N.H.; Vanhoutte, P.M. Endothelial dysfunction in diabetes. Br. J. Pharmacol. 2000, 130, 963–974. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, E.; Alam, N.M.; Prusky, G.T.; Sagdullaev, B.T. Blood-retina barrier failure and vision loss in neuron-specific degeneration. JCI. Insight 2019, 5, e126747. [Google Scholar] [CrossRef] [Green Version]
- Shin, E.S.; Sorenson, C.M.; Sheibani, N. Diabetes and retinal vascular dysfunction. J. Ophthalmic. Vis. Res. 2014, 9, 362–373. [Google Scholar]
- Cunha-Vaz, J.; Bernardes, R.; Lobo, C. Blood-retinal barrier. Eur. J. Ophthalmol. 2011, 21 (Suppl 6), S3–S9. [Google Scholar] [CrossRef]
- Runkle, E.A.; Antonetti, D.A. The blood-retinal barrier: Structure and functional significance. Methods Mol. Biol. 2011, 686, 133–148. [Google Scholar]
- Cholkar, K.; Dasari, S.R.; Pal, D.; Mitra, A.K. Eye: Anatomy, physiology and barriers to drug delivery. Ocul. Transp. Recept. 2013, 1–36. [Google Scholar] [CrossRef]
- Thumann, G.; Hoffmann, S.; Hinton, D.R. Cell biology of the retinal pigment epithelium. Retina 2006, 1, 137–152. [Google Scholar]
- Osanai, M.; Takasawa, A.; Murata, M.; Sawada, N. Claudins in cancer: Bench to bedside. Pflugers Arch. 2017, 469, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Campbell, H.K.; Maiers, J.L.; DeMali, K.A. Interplay between tight junctions & adherens junctions. Exp. Cell Res. 2017, 358, 39–44. [Google Scholar] [PubMed]
- Brückner, B.R.; Janshoff, A. Importance of integrity of cell-cell junctions for the mechanics of confluent mdck ii cells. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rajamani, U.; Jialal, I. Hyperglycemia induces toll-like receptor-2 and -4 expression and activity in human microvascular retinal endothelial cells: Implications for diabetic retinopathy. J. Diabetes Res. 2014, 2014, 790902. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.A.; Mohr, S. Inhibition of caspase-1/interleukin-1β signaling prevents degeneration of retinal capillaries in diabetes and galactosemia. Diabetes 2007, 56, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joussen, A.M.; Doehmen, S.; Le, M.L.; Koizumi, K.; Radetzky, S.; Krohne, T.U.; Poulaki, V.; Semkova, I.; Kociok, N. Tnf-α mediated apoptosis plays an important role in the development of early diabetic retinopathy and long-term histopathological alterations. Mol. Vis. 2009, 15, 1418–1428. [Google Scholar]
- Adams, D.H.; Shaw, S. Leucocyte-endothelial interactions and regulation of leucocyte migration. Lancet 1994, 343, 831–836. [Google Scholar] [CrossRef]
- Miyamoto, K.; Khosrof, S.; Bursell, S.E.; Rohan, R.; Murata, T.; Clermont, A.C.; Aiello, L.P.; Ogura, Y.; Adamis, A.P. Prevention of leukostasis and vascular leakage in streptozotocin-induced diabetic retinopathy via intercellular adhesion molecule-1 inhibition. Proc. Natl. Acad. Sci. USA 1999, 96, 10836–10841. [Google Scholar] [CrossRef] [Green Version]
- Khalfaoui, T.; Lizard, G.; Ouertani-Meddeb, A. Adhesion molecules (icam-1 and vcam-1) and diabetic retinopathy in type 2 diabetes. J. Mol. Histol. 2008, 39, 243–249. [Google Scholar] [CrossRef]
- Koopman, W.J.; Nijtmans, L.G.; Dieteren, C.E.; Roestenberg, P.; Valsecchi, F.; Smeitink, J.A.; Willems, P.H. Mammalian mitochondrial complex i: Biogenesis, regulation, and reactive oxygen species generation. Antioxid. Redox Signal. 2010, 12, 1431–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecilia, O.-M.; José Alberto, C.-G.; José, N.-P.; Ernesto Germán, C.-M.; Ana Karen, L.-C.; Luis Miguel, R.-P.; Ricardo Raúl, R.-R.; Adolfo Daniel, R.-C. Oxidative stress as the main target in diabetic retinopathy pathophysiology. J. Diabetes Res. 2019, 2019, 8562408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Coránguez, M.; Ramos, C.; Antonetti, D.A. The inner blood-retinal barrier: Cellular basis and development. Vis. Res. 2017, 139, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Singh, R.P. The role of anti-vascular endothelial growth factor (anti-vegf) in the management of proliferative diabetic retinopathy. Drugs Context 2018, 7, 212532. [Google Scholar] [CrossRef]
- Shibuya, M. Differential roles of vascular endothelial growth factor receptor-1 and receptor-2 in angiogenesis. J. Biochem. Mol. Biol. 2006, 39, 469–478. [Google Scholar] [CrossRef]
- Arulkumaran, N.; Unwin, R.J.; Tam, F.W. A potential therapeutic role for p2x7 receptor (p2x7r) antagonists in the treatment of inflammatory diseases. Expert Opin. Investig. Drugs 2011, 20, 897–915. [Google Scholar] [CrossRef] [Green Version]
- Parodi, J.; Flores, C.; Aguayo, C.; Rudolph, M.I.; Casanello, P.; Sobrevia, L. Inhibition of nitrobenzylthioinosine-sensitive adenosine transport by elevated d-glucose involves activation of p2y2 purinoceptors in human umbilical vein endothelial cells. Circ. Res. 2002, 90, 570–577. [Google Scholar] [CrossRef] [Green Version]
- Trueblood, K.E.; Mohr, S.; Dubyak, G.R. Purinergic regulation of high-glucose-induced caspase-1 activation in the rat retinal müller cell line rmc-1. Am. J. Physiol. Cell. Physiol. 2011, 301, C1213–C1223. [Google Scholar] [CrossRef]
- Sathanoori, R.; Swärd, K.; Olde, B.; Erlinge, D. The atp receptors p2x7 and p2x4 modulate high glucose and palmitate-induced inflammatory responses in endothelial cells. PLoS ONE 2015, 10, e0125111. [Google Scholar]
- Shibata, M.; Ishizaki, E.; Zhang, T.; Fukumoto, M.; Barajas-Espinosa, A.; Li, T.; Puro, D.G. Purinergic vasotoxicity: Role of the pore/oxidant/katp channel/ca2+ pathway in p2x7-induced cell death in retinal capillaries. Vision 2018, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, T. Role of p2x7 receptors in the development of diabetic retinopathy. World. J. Diabetes 2014, 5, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Platania, C.B.M.; Lazzara, F.; Fidilio, A.; Fresta, C.G.; Conti, F.; Giurdanella, G.; Leggio, G.M.; Salomone, S.; Drago, F.; Bucolo, C. Blood-retinal barrier protection against high glucose damage: The role of p2x7 receptor. Biochem. Pharmacol. 2019, 168, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Fresta, C.G.; Fidilio, A.; Caruso, G.; Caraci, F.; Giblin, F.J.; Leggio, G.M.; Salomone, S.; Drago, F.; Bucolo, C. A new human blood–retinal barrier model based on endothelial cells, pericytes, and astrocytes. Int. J. Mol. Sci. 2020, 21, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, A.E.; Yoshioka, C.; Mansoor, S.E. Full-length p2x(7) structures reveal how palmitoylation prevents channel desensitization. Cell 2019, 179, 659–670. [Google Scholar] [CrossRef]
- Platania, C.B.M.; Pittalà, V.; Pascale, A.; Marchesi, N.; Anfuso, C.D.; Lupo, G.; Cristaldi, M.; Olivieri, M.; Lazzara, F.; Di Paola, L.; et al. Novel indole derivatives targeting hur-mrna complex to counteract high glucose damage in retinal endothelial cells. Biochem. Pharmacol. 2020, 175, 113908. [Google Scholar] [CrossRef]
- Platania, C.B.M.; Giurdanella, G.; Di Paola, L.; Leggio, G.M.; Drago, F.; Salomone, S.; Bucolo, C. P2x7 receptor antagonism: Implications in diabetic retinopathy. Biochem. Pharmacol. 2017, 138, 130–139. [Google Scholar] [CrossRef]
- Eshaq, R.S.; Aldalati, A.M.Z.; Alexander, J.S.; Harris, N.R. Diabetic retinopathy: Breaking the barrier. Pathophysiology 2017, 24, 229–241. [Google Scholar] [CrossRef]
- Wisniewska-Kruk, J.; Hoeben, K.A.; Vogels, I.M.; Gaillard, P.J.; Van Noorden, C.J.; Schlingemann, R.O.; Klaassen, I. A novel co-culture model of the blood-retinal barrier based on primary retinal endothelial cells, pericytes and astrocytes. Exp. Eye Res. 2012, 96, 181–190. [Google Scholar] [CrossRef]
- Ragelle, H.; Goncalves, A.; Kustermann, S.; Antonetti, D.A.; Jayagopal, A. Organ-on-a-chip technologies for advanced blood-retinal barrier models. J. Ocul. Pharmacol. Ther. 2020, 36, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Musso, N.; Costantino, A.; Lazzarino, G.; Tascedda, F.; Gulisano, M.; Lunte, S.M.; Caraci, F. Microfluidics as a novel tool for biological and toxicological assays in drug discovery processes: Focus on microchip electrophoresis. Micromachines 2020, 11, 593. [Google Scholar] [CrossRef]
- Giuliani, A.L.; Sarti, A.C.; Falzoni, S.; Di Virgilio, F. The p2x7 receptor-interleukin-1 liaison. Front. Pharmacol. 2017, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boss, J.D.; Singh, P.K.; Pandya, H.K.; Tosi, J.; Kim, C.; Tewari, A.; Juzych, M.S.; Abrams, G.W.; Kumar, A. Assessment of neurotrophins and inflammatory mediators in vitreous of patients with diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5594–5603. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Hwang, D.K.; Song, X.; Tao, Y. Association between aqueous cytokines and diabetic retinopathy stage. J. Ophthalmol. 2017, 2017, 9402198. [Google Scholar] [CrossRef] [PubMed]
- Bucolo, C.; Gozzo, L.; Longo, L.; Mansueto, S.; Vitale, D.C.; Drago, F. Long-term efficacy and safety profile of multiple injections of intravitreal dexamethasone implant to manage diabetic macular edema: A systematic review of real-world studies. J. Pharmacol. Sci. 2018, 138, 219–232. [Google Scholar] [CrossRef]
- Sugiyama, T.; Kawamura, H.; Yamanishi, S.; Kobayashi, M.; Katsumura, K.; Puro, D.G. Regulation of p2x7-induced pore formation and cell death in pericyte-containing retinal microvessels. Am. J. Physiol. Cell Physiol. 2005, 288, C568–C576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawamura, H.; Sugiyama, T.; Wu, D.M.; Kobayashi, M.; Yamanishi, S.; Katsumura, K.; Puro, D.G. Atp: A vasoactive signal in the pericyte-containing microvasculature of the rat retina. J. Physiol. 2003, 551, 787–799. [Google Scholar] [CrossRef]
- Clapp, C.; Diaz-Lezama, N.; Adan-Castro, E.; Ramirez-Hernandez, G.; Moreno-Carranza, B.; Sarti, A.C.; Falzoni, S.; Solini, A.; Di Virgilio, F. Pharmacological blockade of the p2x7 receptor reverses retinal damage in a rat model of type 1 diabetes. Acta Diabetol. 2019, 56, 1031–1036. [Google Scholar] [CrossRef]
- Sanderson, J.; Dartt, D.A.; Trinkaus-Randall, V.; Pintor, J.; Civan, M.M.; Delamere, N.A.; Fletcher, E.L.; Salt, T.E.; Grosche, A.; Mitchell, C.H. Purines in the eye: Recent evidence for the physiological and pathological role of purines in the rpe, retinal neurons, astrocytes, müller cells, lens, trabecular meshwork, cornea and lacrimal gland. Exp. Eye Res. 2014, 127, 270–279. [Google Scholar] [CrossRef]
- Križaj, D.; Ryskamp, D.A.; Tian, N.; Tezel, G.; Mitchell, C.H.; Slepak, V.Z.; Shestopalov, V.I. From mechanosensitivity to inflammatory responses: New players in the pathology of glaucoma. Curr. Eye Res. 2014, 39, 105–119. [Google Scholar] [CrossRef]
- Beckel, J.M.; Argall, A.J.; Lim, J.C.; Xia, J.; Lu, W.; Coffey, E.E.; Macarak, E.J.; Shahidullah, M.; Delamere, N.A.; Zode, G.S.; et al. Mechanosensitive release of adenosine 5’-triphosphate through pannexin channels and mechanosensitive upregulation of pannexin channels in optic nerve head astrocytes: A mechanism for purinergic involvement in chronic strain. Glia 2014, 62, 1486–1501. [Google Scholar] [CrossRef]
- Romano, G.L.; Amato, R.; Lazzara, F.; Porciatti, V.; Chou, T.H.; Drago, F.; Bucolo, C. P2x7 receptor antagonism preserves retinal ganglion cells in glaucomatous mice. Biochem. Pharmacol. 2020, 180, 114199. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A. Recent advances in cns p2x7 physiology and pharmacology: Focus on neuropsychiatric disorders. Front. Pharmacol. 2018, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Agostino, V.G.; Lal, P.; Mantelli, B.; Tiedje, C.; Zucal, C.; Thongon, N.; Gaestel, M.; Latorre, E.; Marinelli, L.; Seneci, P.; et al. Dihydrotanshinone-i interferes with the rna-binding activity of hur affecting its post-transcriptional function. Sci. Rep. 2015, 5, 16478. [Google Scholar]
- Bharadwaj, S.; Lee, K.E.; Dwivedi, V.D. Discovery of ganoderma lucidum triterpenoids as potential inhibitors against dengue virus ns2b-ns3 protease. Sci. Rep. 2019, 9, 19059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, Y.; Suzuki, Y.; Nakamura, A.; Watanabe, Y.; Sekiya, M.; Roppongi, S.; Kushibiki, C.; Iizuka, I.; Tani, O.; Sakashita, H.; et al. Fragment-based discovery of the first nonpeptidyl inhibitor of an s46 family peptidase. Sci. Rep. 2019, 9, 13587. [Google Scholar] [CrossRef]
- Pittalà, V.; Vanella, L.; Maria Platania, C.B.; Salerno, L.; Raffaele, M.; Amata, E. Synthesis, in vitro and in silico studies of ho-1 inducers and lung antifibrotic agents. Future Med. Chem. 2019, 11, 1523–1536. [Google Scholar] [CrossRef]
- Adamczyk, M.; Griffiths, R.; Dewitt, S.; Knäuper, V.; Aeschlimann, D. P2x7 receptor activation regulates rapid unconventional export of transglutaminase-2. J. Cell Sci. 2015, 128, 4615–4628. [Google Scholar] [CrossRef] [Green Version]
- Di Virgilio, F.; Giuliani, A.L.; Vultaggio-Poma, V.; Falzoni, S.; Sarti, A.C. Non-nucleotide agonists triggering p2x7 receptor activation and pore formation. Front. Pharmacol. 2018, 9, 39. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Gagliano, C.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. Vip family members prevent outer blood retinal barrier damage in a model of diabetic macular edema. J. Cell. Physiol. 2017, 232, 1079–1085. [Google Scholar] [CrossRef]
- Lobo, C.L.; Bernardes, R.C.; Cunha-Vaz, J.G. Alterations of the blood-retinal barrier and retinal thickness in preclinical retinopathy in subjects with type 2 diabetes. Arch. Ophthalmol. 2000, 118, 1364–1369. [Google Scholar] [CrossRef] [Green Version]
- Umeda, K.; Matsui, T.; Nakayama, M.; Furuse, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Establishment and characterization of cultured epithelial cells lacking expression of zo-1. J. Biol. Chem. 2004, 279, 44785–44794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeil, E.; Capaldo, C.T.; Macara, I.G. Zonula occludens-1 function in the assembly of tight junctions in madin-darby canine kidney epithelial cells. Mol. Biol. Cell. 2006, 17, 1922–1932. [Google Scholar] [CrossRef] [Green Version]
- Deissler, H.L.; Deissler, H.; Lang, G.K.; Lang, G.E. Vegf but not plgf disturbs the barrier of retinal endothelial cells. Exp. Eye Res. 2013, 115, 162–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.S.; Kim, Y.G.; Cho, H.J.; Park, J.; Jeong, H.; Lee, S.E.; Lee, S.P.; Kang, H.J.; Kim, H.S. Dipeptidyl peptidase-4 inhibitor increases vascular leakage in retina through ve-cadherin phosphorylation. Sci. Rep. 2016, 6, 29393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Wang, H.; Liu, Y.; Li, W.; Kim, D.; Huang, H. Blockade of vascular endothelial growth factor receptor 1 prevents inflammation and vascular leakage in diabetic retinopathy. J. Ophthalmol. 2015, 2015, 605946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Kharashi, A.S. Role of oxidative stress, inflammation, hypoxia and angiogenesis in the development of diabetic retinopathy. Saudi J. Ophthalmol. 2018, 32, 318–323. [Google Scholar] [CrossRef]
- Rivera, J.C.; Dabouz, R.; Noueihed, B.; Omri, S.; Tahiri, H.; Chemtob, S. Ischemic retinopathies: Oxidative stress and inflammation. Oxid. Med. Cell Longev. 2017, 2017, 3940241. [Google Scholar] [CrossRef] [Green Version]
- El-Remessy, A.B.; Tawfik, H.E.; Matragoon, S.; Pillai, B.; Caldwell, R.B.; Caldwell, R.W. Peroxynitrite mediates diabetes-induced endothelial dysfunction: Possible role of rho kinase activation. Exp. Diabetes. Res. 2010, 2010, 247861. [Google Scholar] [CrossRef]
- Sorrentino, F.S.; Matteini, S.; Bonifazzi, C.; Sebastiani, A.; Parmeggiani, F. Diabetic retinopathy and endothelin system: Microangiopathy versus endothelial dysfunction. Eye (Lond.) 2018, 32, 1157–1163. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, M.; Katsnelson, M.A.; Dubyak, G.R. Neutrophil p2x7 receptors mediate nlrp3 inflammasome-dependent il-1β secretion in response to atp. Nat. Commun. 2016, 7, 10555. [Google Scholar] [CrossRef] [Green Version]
- Doktor, F.; Prager, P.; Wiedemann, P.; Kohen, L.; Bringmann, A.; Hollborn, M. Hypoxic expression of nlrp3 and vegf in cultured retinal pigment epithelial cells: Contribution of p2y(2) receptor signaling. Purinergic. Signal. 2018, 14, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.; Tsung, A.; Billiar, T. Linking oxidative stress to inflammation: Toll-like receptors. Free Radic. Biol. Med. 2010, 48, 1121–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, R.; Huang, L.; Du, L.J.; Feng, J.F.; Li, J.; Luo, Y.Y.; Xu, Q.M.; Yang, S.L.; Gao, H.; Feng, Y.L. Dihydrotanshinone exhibits an anti-inflammatory effect in vitro and in vivo through blocking tlr4 dimerization. Pharmacol. Res. 2019, 142, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Wang, Q.; Ao, H.; Shoblock, J.R.; Lord, B.; Aluisio, L.; Fraser, I.; Nepomuceno, D.; Neff, R.A.; Welty, N.; et al. Pharmacological characterization of a novel centrally permeable p2x7 receptor antagonist: Jnj-47965567. Br. J. Pharmacol. 2013, 170, 624–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronica, E.; Bauer, S.; Bozzi, Y.; Caleo, M.; Dingledine, R.; Gorter, J.A.; Henshall, D.C.; Kaufer, D.; Koh, S.; Löscher, W.; et al. Neuroinflammatory targets and treatments for epilepsy validated in experimental models. Epilepsia 2017, 58 (Suppl 3), 27–38. [Google Scholar] [CrossRef]
- Yu, L.; Qian, J. Dihydrotanshinone i alleviates spinal cord injury via suppressing inflammatory response, oxidative stress and apoptosis in rats. Med. Sci. Monit. 2020, 26, e920738. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, H.; Li, W.; Wu, H.; Yang, Y. Highly-expressed p2x7 receptor promotes growth and metastasis of human hos/mnng osteosarcoma cells via pi3k/akt/gsk3β/β-catenin and mtor/hif1α/vegf signaling. Int. J. Cancer. 2019, 145, 1068–1082. [Google Scholar] [CrossRef] [Green Version]
- Qaum, T.; Xu, Q.; Joussen, A.M.; Clemens, M.W.; Qin, W.; Miyamoto, K.; Hassessian, H.; Wiegand, S.J.; Rudge, J.; Yancopoulos, G.D.; et al. Vegf-initiated blood-retinal barrier breakdown in early diabetes. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2408–2413. [Google Scholar]
- Ahuja, S.; Saxena, S.; Akduman, L.; Meyer, C.H.; Kruzliak, P.; Khanna, V.K. Serum vascular endothelial growth factor is a biomolecular biomarker of severity of diabetic retinopathy. Int. J. Retin. Vitr. 2019, 5, 29. [Google Scholar] [CrossRef]
- Bobbie, M.W.; Roy, S.; Trudeau, K.; Munger, S.J.; Simon, A.M.; Roy, S. Reduced connexin 43 expression and its effect on the development of vascular lesions in retinas of diabetic mice. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3758–3763. [Google Scholar] [CrossRef]
- Tien, T.; Barrette, K.F.; Chronopoulos, A.; Roy, S. Effects of high glucose-induced cx43 downregulation on occludin and zo-1 expression and tight junction barrier function in retinal endothelial cells. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6518–6525. [Google Scholar] [CrossRef] [PubMed]
- Kamiuchi, K.; Hasegawa, G.; Obayashi, H.; Kitamura, A.; Ishii, M.; Yano, M.; Kanatsuna, T.; Yoshikawa, T.; Nakamura, N. Intercellular adhesion molecule-1 (icam-1) polymorphism is associated with diabetic retinopathy in type 2 diabetes mellitus. Diabet. Med. 2002, 19, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Platania, C.B.M.; Ronchetti, S.; Riccardi, C.; Migliorati, G.; Marchetti, M.C.; Di Paola, L.; Lazzara, F.; Drago, F.; Salomone, S.; Bucolo, C. Effects of protein-protein interface disruptors at the ligand of the glucocorticoid-induced tumor necrosis factor receptor-related gene (gitr). Biochem. Pharmacol. 2020, 178, 114110. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Chuaqui, C.; Singh, J. Structural interaction fingerprint (sift): A novel method for analyzing three-dimensional protein-ligand binding interactions. J. Med. Chem. 2004, 47, 337–344. [Google Scholar] [CrossRef]
- Giurdanella, G.; Lazzara, F.; Caporarello, N.; Lupo, G.; Anfuso, C.D.; Eandi, C.M.; Leggio, G.M.; Drago, F.; Bucolo, C.; Salomone, S. Sulodexide prevents activation of the pla2/cox-2/vegf inflammatory pathway in human retinal endothelial cells by blocking the effect of age/rage. Biochem. Pharmacol. 2017, 142, 145–154. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. Imagej2: Imagej for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Caruso, G.; Fresta, C.G.; Fidilio, A.; O’Donnell, F.; Musso, N.; Lazzarino, G.; Grasso, M.; Amorini, A.M.; Tascedda, F.; Bucolo, C.; et al. Carnosine decreases pma-induced oxidative stress and inflammation in murine macrophages. Antioxidants 2019, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Torrisi, S.A.; Geraci, F.; Tropea, M.R.; Grasso, M.; Caruso, G.; Fidilio, A.; Musso, N.; Sanfilippo, G.; Tascedda, F.; Palmeri, A.; et al. Fluoxetine and vortioxetine reverse depressive-like phenotype and memory deficits induced by abeta1-42 oligomers in mice: A key role of transforming growth factor-beta1. Front. Pharmacol. 2019, 10, 693. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Docking Score | ΔGbinding Allosteric (kcal/mol) | ΔGbinding Orthosteric (kcal/mol) | ΔGbinding Cytosolic (kcal/mol) | |
|---|---|---|---|---|
| DHTS | −7.0 | −59 | −49 | −30 |
| JNJ47965567 | −8.0 | −86 | N.A. | N.A. |
| A438079 | −7.5 | −56 | N.A. | N.A. |
| Quercetin | −6.0 | −30 | −20 | −30 |
| Official Name # | Official Symbol | Alternative Titles/Symbols | Detected Transcript | Amplicon Length | Cat. No. § |
|---|---|---|---|---|---|
| interleukin 1, beta | IL1B | IL-1, IL1F2, IL1beta, IL1-BETA | NM_000576, XM_006712496 | 117 bp 117 bp | QT00021385 |
| interleukin 6 | IL6 | CDF, HGF, HSF, BSF2, IL-6, BSF-2, IFNB2, IFN-beta-2 | NM_000600, XM_005249745 | 107 bp 107 bp | QT00083720 |
| tumor necrosis factor | TNF | DIF, TNFA, TNFSF2, TNLG1F, TNF-alpha | NM_000594 | 98 bp | QT00029162 |
| vascular endothelial growth factor A | VEGFA | VPF, VEGF, MVCD1 | NM_001025366, NM_001025367, NM_001025368, NM_001033756, NM_001171623, NM_001171624, NM_001171625, NM_001171626, NM_001171629, NM_003376, NM_001287044 | 273 204 150 150 273 222 204 150 150 222 150 | QT01010184 |
| chemokine (C-X-C motif) ligand 8 | CXCL8 | GCP-1, GCP1, IL8, LECT, LUCT, LYNAP, MDNCF, MONAP, NAF, NAP-1, NAP1, SCYB8 | NM_000584 | 102 bp | QT00000322 |
| cytochrome b-245 beta chain | CYBB | CGD, NOX2, IMD34, AMCBX2, GP91-1, GP91PHOX, p91-PHOX, GP91-PHOX | NM_000397 | 124 bp | QT00029533 |
| transforming growth factor beta 1 | TGFB1 | CED, LAP, DPD1, TGFB, IBDIMDE, TGFbeta, TGF-beta1 | NM_000660 | 108 bp | QT00000728 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fresta, C.G.; Caruso, G.; Fidilio, A.; Platania, C.B.M.; Musso, N.; Caraci, F.; Drago, F.; Bucolo, C. Dihydrotanshinone, a Natural Diterpenoid, Preserves Blood-Retinal Barrier Integrity via P2X7 Receptor. Int. J. Mol. Sci. 2020, 21, 9305. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239305
Fresta CG, Caruso G, Fidilio A, Platania CBM, Musso N, Caraci F, Drago F, Bucolo C. Dihydrotanshinone, a Natural Diterpenoid, Preserves Blood-Retinal Barrier Integrity via P2X7 Receptor. International Journal of Molecular Sciences. 2020; 21(23):9305. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239305
Chicago/Turabian StyleFresta, Claudia Giuseppina, Giuseppe Caruso, Annamaria Fidilio, Chiara Bianca Maria Platania, Nicolò Musso, Filippo Caraci, Filippo Drago, and Claudio Bucolo. 2020. "Dihydrotanshinone, a Natural Diterpenoid, Preserves Blood-Retinal Barrier Integrity via P2X7 Receptor" International Journal of Molecular Sciences 21, no. 23: 9305. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239305