Sensitivity of Rodent Microglia to Kynurenines in Models of Epilepsy and Inflammation In Vivo and In Vitro: Microglia Activation Is Inhibited by Kynurenic Acid and the Synthetic Analogue SZR104

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Seizure Symptoms
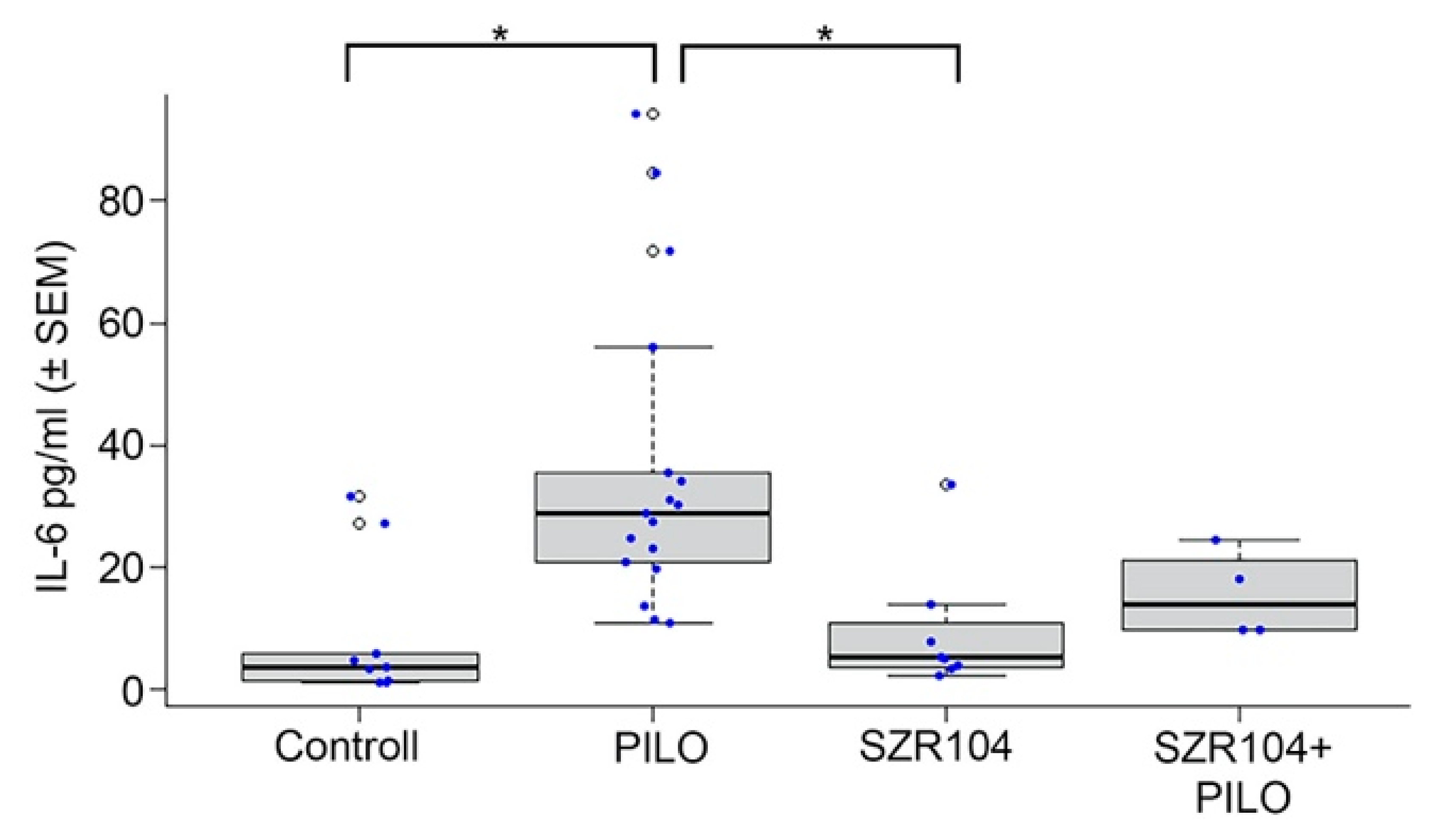
2.2. IL-6 Levels in Blood
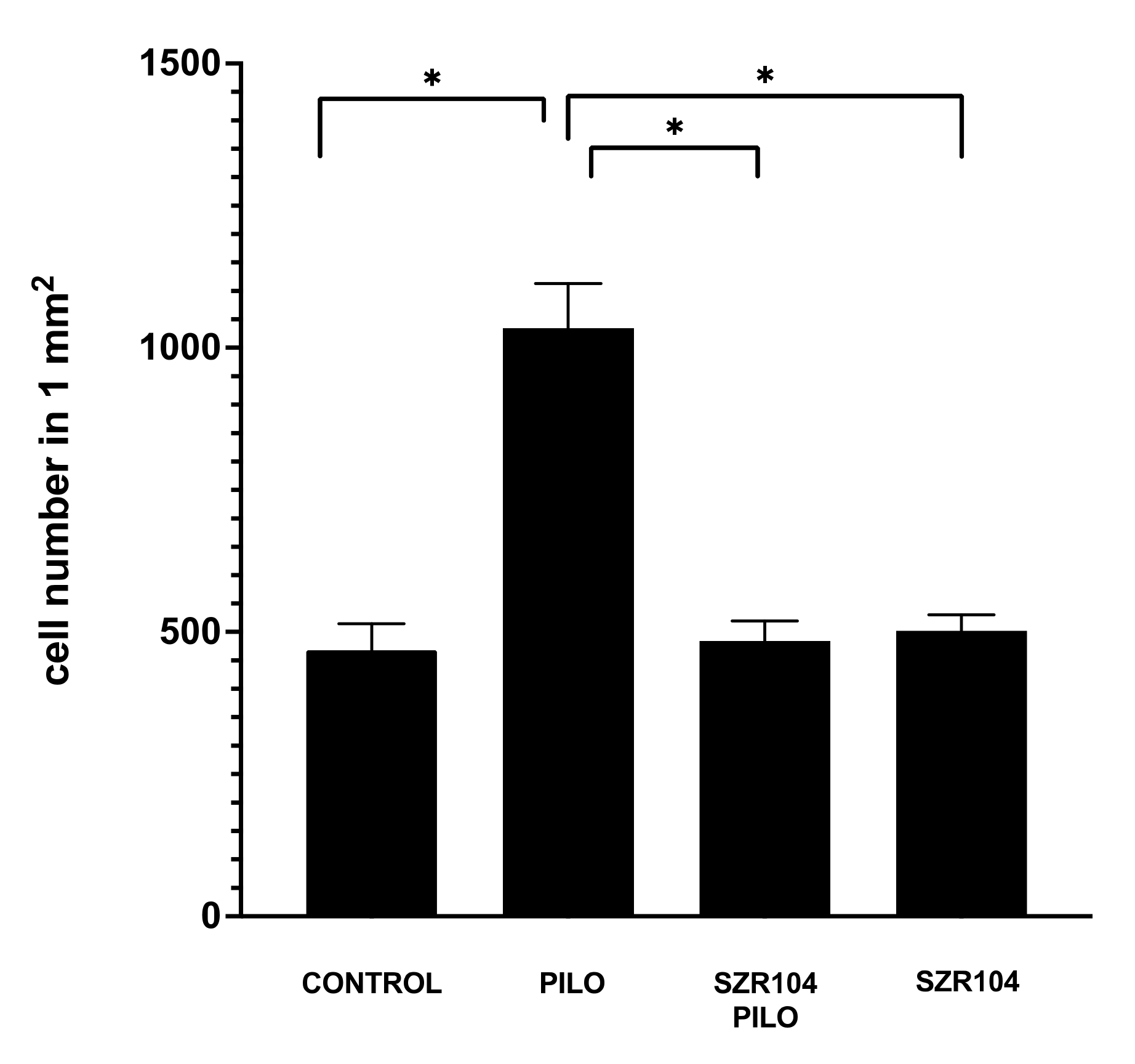
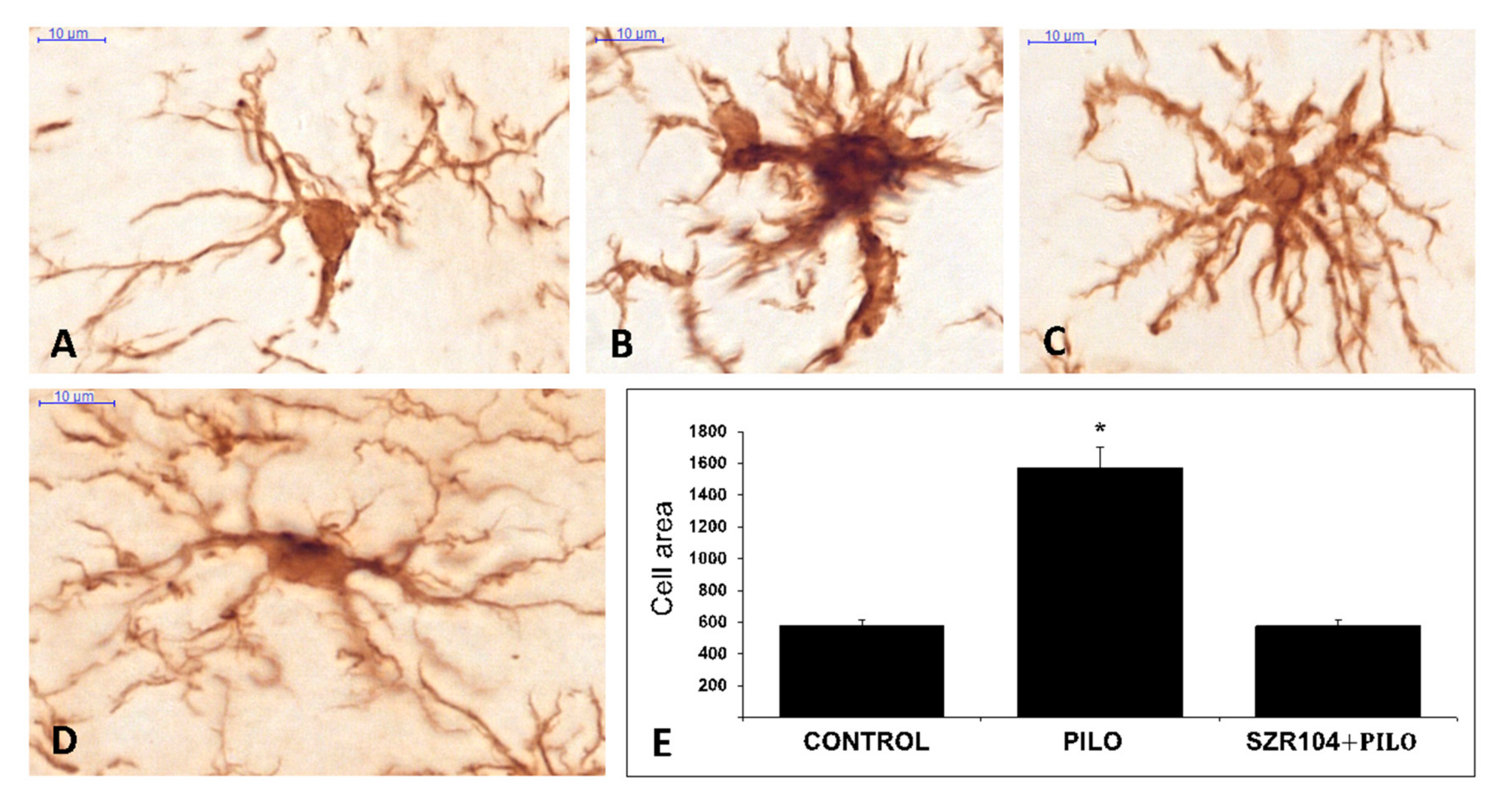
2.3. Iba1 Immunohistochemistry
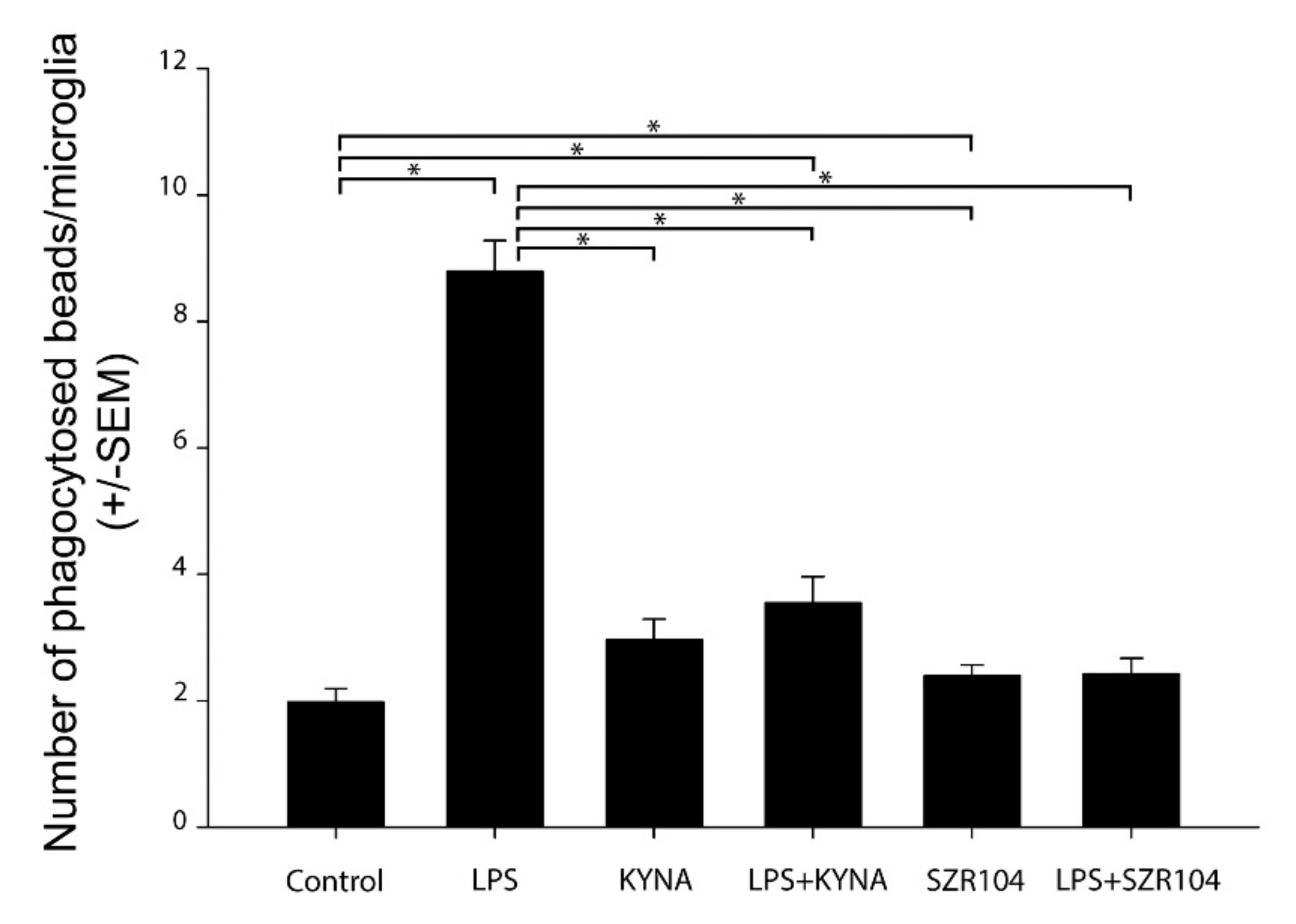
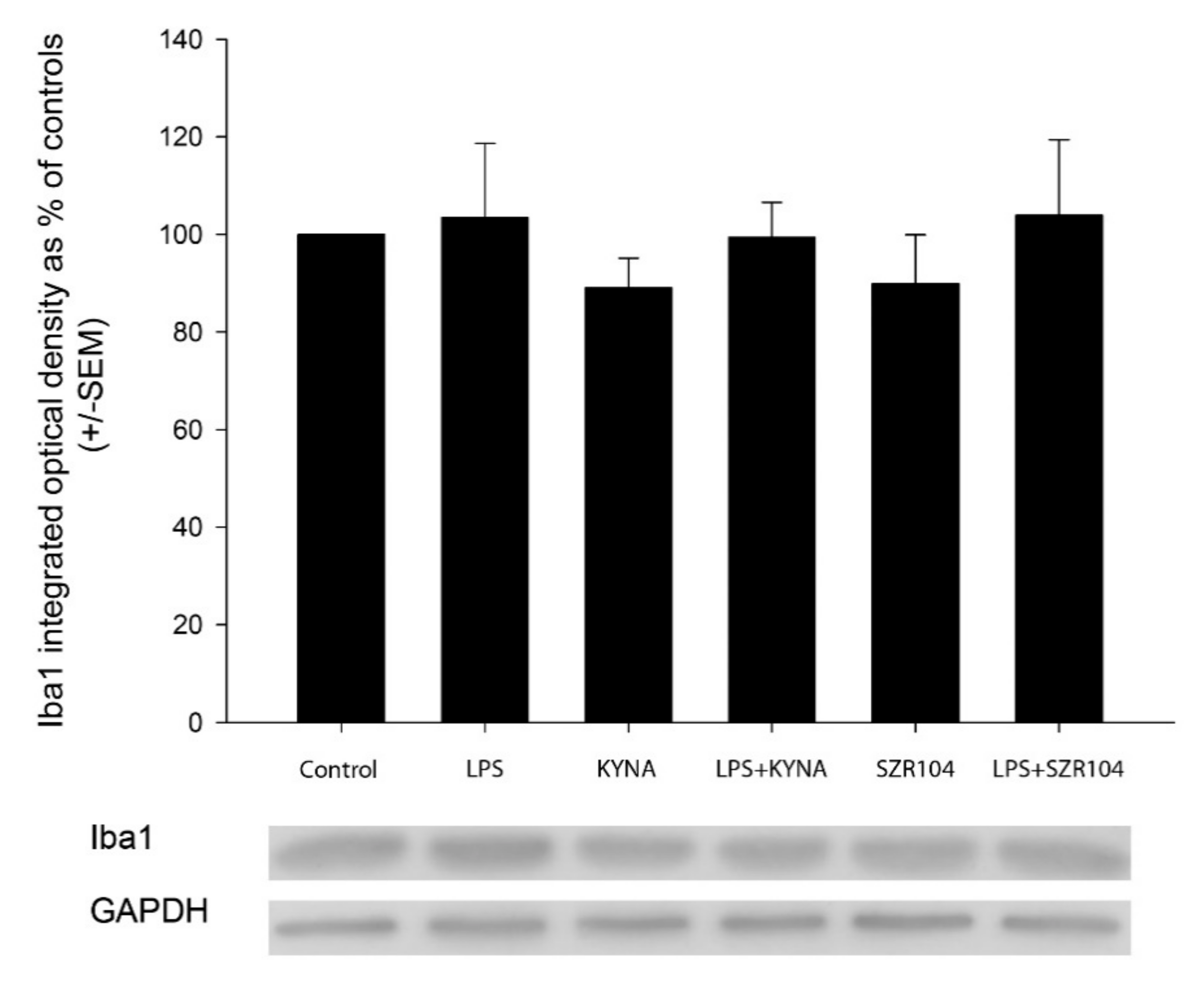
2.4. Effect of KYNA and SZR104 on Microglia Phagocytosis and Iba1 Immunoreactivity
3. Discussion
3.1. KYNA is an Endogenous Regulator in the CNS
3.2. Blood IL-6 Level Alterations in Seizure
3.3. Postepileptic Transformation of Hippocampal Microglia
3.4. Inhibition of Phagocytosis In Vitro by KYNA and SZR104
4. Materials and Methods
4.1. Kynurenines Used
4.2. In Vivo Experiments
4.3. Analysis of Blood Samples
4.4. Iba1 Immunohistochemistry
4.5. Evaluation of the Immunohistochemical Data
4.6. Cell Culture Experiments
4.7. In Vitro Phagocytosis Assay
4.8. Western Blot Analysis
4.9. Image Analysis
5. Conclusions
- The action on AHRs could explain the inhibitory effect of SZR104 on microglia activation and proliferation. The phenotype change of the microglia could be the consequence of the genomic actions of the AHR complex [28], through a molecular scenario including the decrease in TNFα secretion and increasing tumor necrosis factor-stimulated gene-6 (TSG-6) expression [25]. The depletion of TNFα may affect multi-protein complex containing mammalian target of rapamycin (mTOR) protein (mTORC1) activity and the nuclear translocation of mTOR [34]. The depletion of TNFα and mTORC1 activation may arrest the cell cycle in the late G1 phase through metabolic checkpoints [35], thereby preventing microglia proliferation.
- The inhibition of microglia phagocytosis by SZR104 and KYNA could also utilize AHR pathways, causing the decrease in TLR-4 expression on the cell surface [28], the induction of cytoskeletal changes [32] or the deregulation of the nuclear factor kappa-light-chain-enhances of activated B cells (NF-κB) pathway [28].
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AHR | Aryl hydrocarbon receptor |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazole |
| ANOVA | Analysis of variance |
| AOI | Area of interest |
| BBB | Blood–brain barrier |
| CNS | Central nervous system |
| ELISA | Enzyme-linked immunosorbent assay |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GYKI52466 | a 2,3-benzodiazepine, which is non-competitive AMPA receptor antagonist |
| Iba1 | Ionized calcium binding adaptor molecule 1 |
| iGluR | Ionotropic glutamate receptor |
| IL-6 | Interleukin-6 |
| KYNA | Kynurenic acid |
| KP | Kynurenine pathway |
| LPS | Lipopolysaccharide, endotoxin |
| mTOR | Mammalian target of rapamycin |
| mTORC1 | Multi-protein complex containing mTOR protein |
| NF-κB | Nuclear factor kappa-light-chain-enhances of activated B cells; protein complex regulating cellular responses |
| NMDA | N-methyl-D-aspartic acid |
| PILO | Pilocarpine |
| RT-PCR | Reverse transcription polymerase chain reaction |
| QUIN | Quinolinic acid |
| SEM | Standard error of the mean |
| SZR104 | N-(2-(dimethylamino)ethyl)-3-(morpholinomethyl)-4-hydroxyquinoline-2-carboxamide |
| TLR4 | Toll-like receptor4 |
| TNFα | Tumor necrosis factor-alpha |
| TRPA1,4 | Transient receptor potential ankyrin 1,4 receptors |
References
- Biernacki, T.; Sandi, D.; Bencsik, K.; Vécsei, L. Kynurenines in the pathogenesis of multiple sclerosis: Therapeutic perspectives. Cells 2020, 9, 1564. [Google Scholar] [CrossRef] [PubMed]
- Vécsei, L.; Szalárdy, L.; Fülöp, F.; Toldi, J. Kynurenines in the CNS: Recent advances and new questions. Nat. Rev. Drug Discov. 2013, 12, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Fülöp, F.; Szatmári, I.; Vámos, E.; Zádori, D.; Toldi, J.; Vécsei, L. Syntheses, transformations and pharmacological applications of kynurenic acid derivatives. Curr. Med. Chem. 2009, 16, 4828–4842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeter, I.; Nagy, K.; Gellért, L.; Vécsei, L.; Fülöp, F.; Toldi, J. A novel kynurenic acid analog (SZR104) inhibits pentylenetetrazole-induced epileptiform seizures. An electrophysiological study. J. Neural Transm. 2012, 119, 151–154. [Google Scholar] [CrossRef]
- Kovács, A.; Mihály, A.; Komáromi, A.; Gyengési, E.; Szente, M.; Weiczner, R.; Krisztin-Péva, B.; Szabó, G.; Telegdy, G. Seizure, neurotransmitter release, and gene expression are closely related in the striatum of 4-aminopyridine-treated rats. Epilepsy Res. 2003, 55, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Weiczner, R.; Krisztin-Péva, B.; Mihály, A. Blockade of AMPA receptors attenuates 4-aminopyridine seizures, decreases the activation of inhibitory neurons but is ineffective against seizure-related astrocytic swelling. Epilepsy Res. 2008, 78, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Wirthgen, E.; Hoeflich, A.; Rebl, A.; Günther, J. Kynurenic acid: The Janus-faced role of an immunomodularory tryptophan metabolite and its link to pathological conditions. Front. Immunol. 2017, 8, 1957. [Google Scholar] [CrossRef] [Green Version]
- Routy, J.-P.; Routy, B.; Graziani, G.M.; Mehraj, V. The kynurenine pathway is a double-edged sword in immune-privileged sites and in cancer: Implications of immunotherapy. Int. J. Tryptophan Res. 2016, 9, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef]
- Priller, J.; Prinz, M. Targeting microglia in brain disorders. Science 2019, 365, 32–33. [Google Scholar] [CrossRef]
- Hickman, S.; Izzy, S.; Seu, P.; Morsett, L.; El Khoury, J. Microglia in neurodegeneration. Nat. Neurosci. 2018, 21, 1359–1369. [Google Scholar] [CrossRef]
- Lee, M. Neurotransmitters and microglial-mediated neuroinflammation. Curr. Protein Pept. Sci. 2013, 14, 21–32. [Google Scholar] [CrossRef]
- Deczkowska, A.; Keren-Shaul, H.; Weiner, A.; Colonna, M.; Schwartz, M.; Amit, I. Disease-associated microglia: A universal immune sensor of neurodegeneration. Cell 2018, 173, 1073–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirdajova, D.B.; Kriska, J.; Tureckova, J.; Anderova, M. Ischemia-triggered glutamate excitotoxicity from the perspective of glial cells. Front. Cell. Neurosci. 2020, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pocock, J.M.; Kettenmann, H. Neurotransmitter receptors on microglia. Trends Neurosci. 2007, 30, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Sapp, E.; Kegel, K.B.; Aronin, N.; Hashikawa, T.; Uchiyama, Y.; Tohyama, K.; Bhide, P.G.; Vonsattel, J.P.; DiFiglia, M. Early and progressive accumulation of reactive microglia in the Huntington disease brain. J. Neuropathol. Exp. Neurol. 2001, 60, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Hovens, I.B.; Nyakas, C.; Shoemaker, R.G. A novel method for evaluating microglial activation using ionized calcium-binding adaptor protein-1 staining: Cell body to cell size ratio. Neuroimmunol. Neuroinflamm. 2014, 1, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Benson, M.J.; Manzanero, S.; Borges, K. Complex alterations in microglial M1/M2 markers during the development of epilepsy in two mouse models. Epilepsia 2015, 56, 895–905. [Google Scholar] [CrossRef]
- Diaz-Aparicio, I.; Beccari, S.; Abiega, O.; Sierra, A. Clearing the corpses: Regulatory mechanisms, novel tool, and therapeutic potential of harnessing microglial phagocytosis in the diseased brain. Neural Regen. Res. 2016, 11, 1533–1539. [Google Scholar] [CrossRef]
- Kata, D.; Földesi, I.; Fehér, Z.L.; Hackler, L., Jr.; Puskás, L.G.; Gulya, K. Rosuvastatin enhances anti-inflammatory and inhibits pro-inflammatory functions in cultured microglial cells. Neuroscience 2016, 314, 47–63. [Google Scholar] [CrossRef] [Green Version]
- Szabó, M.; Gulya, K. Development of the microglial phenotype in culture. Neuroscience 2013, 241, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Kata, D.; Földesi, I.; Fehér, L.Z.; Hackler, L., Jr.; Puskás, L.G.; Gulya, K. A novel pleiotropic effect of aspirin: Beneficial regulation of pro- and anti-inflammatory mechanisms in microglial cells. Brain Res. Bull. 2017, 132, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.W. Does kynurenic acid act on nicotinic receptors? An assessment of the evidence. J. Neurochem. 2020, 152, 627–649. [Google Scholar] [CrossRef] [Green Version]
- Szakács, R.; Weiczner, R.; Mihály, A.; Krisztin-Péva, B.; Zádor, Z.; Zádor, E. Non-competitive NMDA receptor-antagonists moderate seizure-induced c-fos expression in the rat cerebral cortex. Brain Res. Bull. 2003, 59, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Mándi, Y.; Endrész, V.; Mosolygó, T.; Burián, K.; Lantos, I.; Fülöp, F.; Szatmári, I.; Lőrinczi, B.; Balog, A.; Vécsei, L. The opposite effect of kynurenic acid and different kynurenic acid analogs on tumor necrosis factor-α (TNF-α) production and tumor necrosis factor-stimulated gene-6 (TSG-6) expression. Front. Immunol. 2019, 10, 1406. [Google Scholar] [CrossRef]
- Curia, G.; Longo, D.; Biagini, G.; Jones, R.S.G.; Avoli, M. The pilocarpine model of temporal lobe epilepsy. J. Neurosci. Methods 2008, 172, 143–157. [Google Scholar] [CrossRef]
- Startek, J.B.; Talavera, K.; Voets, T.; Alpizar, Y.A. Differential interactions of bacterial lipopolysaccharides with lipid membranes: Implications for TRPA1-mediated chemosensation. Nat. Sci. Rep. 2018, 8, 12010. [Google Scholar] [CrossRef]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR signaling pathways and regulatory functions. Biochim. Open 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Juricek, L.; Coumoul, X. The aryl hydrocarbon receptor and the nervous system. Int. J. Mol. Sci. 2018, 19, 2504. [Google Scholar] [CrossRef] [Green Version]
- Mulero-Navarro, S.; Fernandez-Salguero, P.M. New trends in aryl hydrocarbon receptor biology. Front. Cell Dev. Biol. 2016, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kondrikov, D.; Elmansi, A.; Bragg, R.T.; Mobley, T.; Barrett, T.; Eisa, N.; Kondrikova, G.; Schoeinlein, P.; Aguilar-Perez, A.; Shi, X.-M.; et al. Kynurenine inhibits autophagy and promotes senescence in aged bone marrow mesenchymal stem cells through the arylhydrocarbon receptor pathway. Exp. Gerontol. 2020, 130, 110805. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Shu, Q.; Sun, H.; Ma, Y.; Kang, D.; Zhao, Y.; Lu, J.; Gong, P.; Yang, F.; Wan, F. 1′H-indole-3′-carbonyl-thiazole-4-carboxylic acid methyl ester blocked human glioma cell invasion via aryl hydrocarbon receptors’ regulation of cytoskeletal contraction. Biomed Res. Int. 2020. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, S.; Saqcena, M.; Foster, D.A. Synthetic lethality in KRas-driven cancer cells created by glutamine deprivation. Oncoscience 2015, 2, 807–808. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pharmacological Treatment | Number of Surviving Animals | Experimental Procedures |
|---|---|---|
| Control animals: 0.9% NaCl i.p. injection | 12 | Blood samples (8 animals). Immunohistochemistry (4 animals). |
| Pilocarpine-treated: 190 mg/kg PILO, i.p. injection | 21 | Blood samples (18 animals). Immunohistochemistry (4 animals). |
| SZR104 solution i.p. (358 mg/kg) | 12 | Blood samples (8 animals). Immunohistochemistry (4 animals). |
| SZR104 (358 mg/kg) and PILO (190 mg/kg) i.p. injections | 8 | Blood samples (4 animals). Immunohistochemistry (4 animals). |
| Code | Molecular Structure | Chemical Name | Empirical Formula and Molecular Weight |
|---|---|---|---|
| KYNA |  | 4-hydroxyquinolin-2-carboxylic acid | C10H7NO3 189.17 |
| SZR104 |  | N-(2-(dimethylamino)ethyl)-3-(morpholinomethyl)-4-hydroxyquinoline-2-carboxamide | C19H26N4O3 358.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lajkó, N.; Kata, D.; Szabó, M.; Mátyás, A.; Dulka, K.; Földesi, I.; Fülöp, F.; Gulya, K.; Vécsei, L.; Mihály, A. Sensitivity of Rodent Microglia to Kynurenines in Models of Epilepsy and Inflammation In Vivo and In Vitro: Microglia Activation Is Inhibited by Kynurenic Acid and the Synthetic Analogue SZR104. Int. J. Mol. Sci. 2020, 21, 9333. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239333
Lajkó N, Kata D, Szabó M, Mátyás A, Dulka K, Földesi I, Fülöp F, Gulya K, Vécsei L, Mihály A. Sensitivity of Rodent Microglia to Kynurenines in Models of Epilepsy and Inflammation In Vivo and In Vitro: Microglia Activation Is Inhibited by Kynurenic Acid and the Synthetic Analogue SZR104. International Journal of Molecular Sciences. 2020; 21(23):9333. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239333
Chicago/Turabian StyleLajkó, Noémi, Diana Kata, Melinda Szabó, Adrienne Mátyás, Karolina Dulka, Imre Földesi, Ferenc Fülöp, Karoly Gulya, László Vécsei, and András Mihály. 2020. "Sensitivity of Rodent Microglia to Kynurenines in Models of Epilepsy and Inflammation In Vivo and In Vitro: Microglia Activation Is Inhibited by Kynurenic Acid and the Synthetic Analogue SZR104" International Journal of Molecular Sciences 21, no. 23: 9333. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239333