Structural Diversity and Highly Specific Host-Pathogen Transcriptional Regulation of Defensin Genes Is Revealed in Tomato

 ,
,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
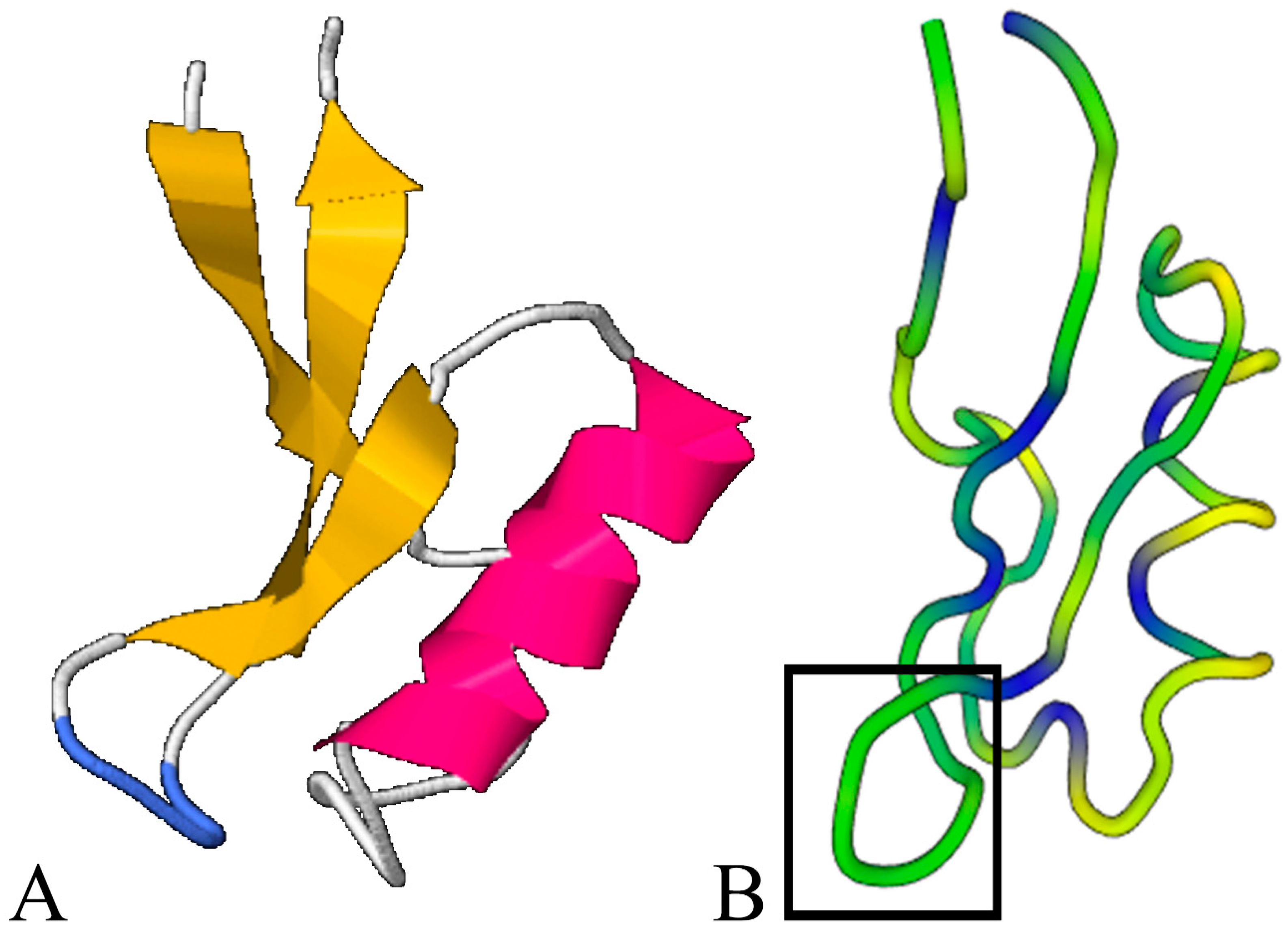
2.1. Defensin Structure Analysis
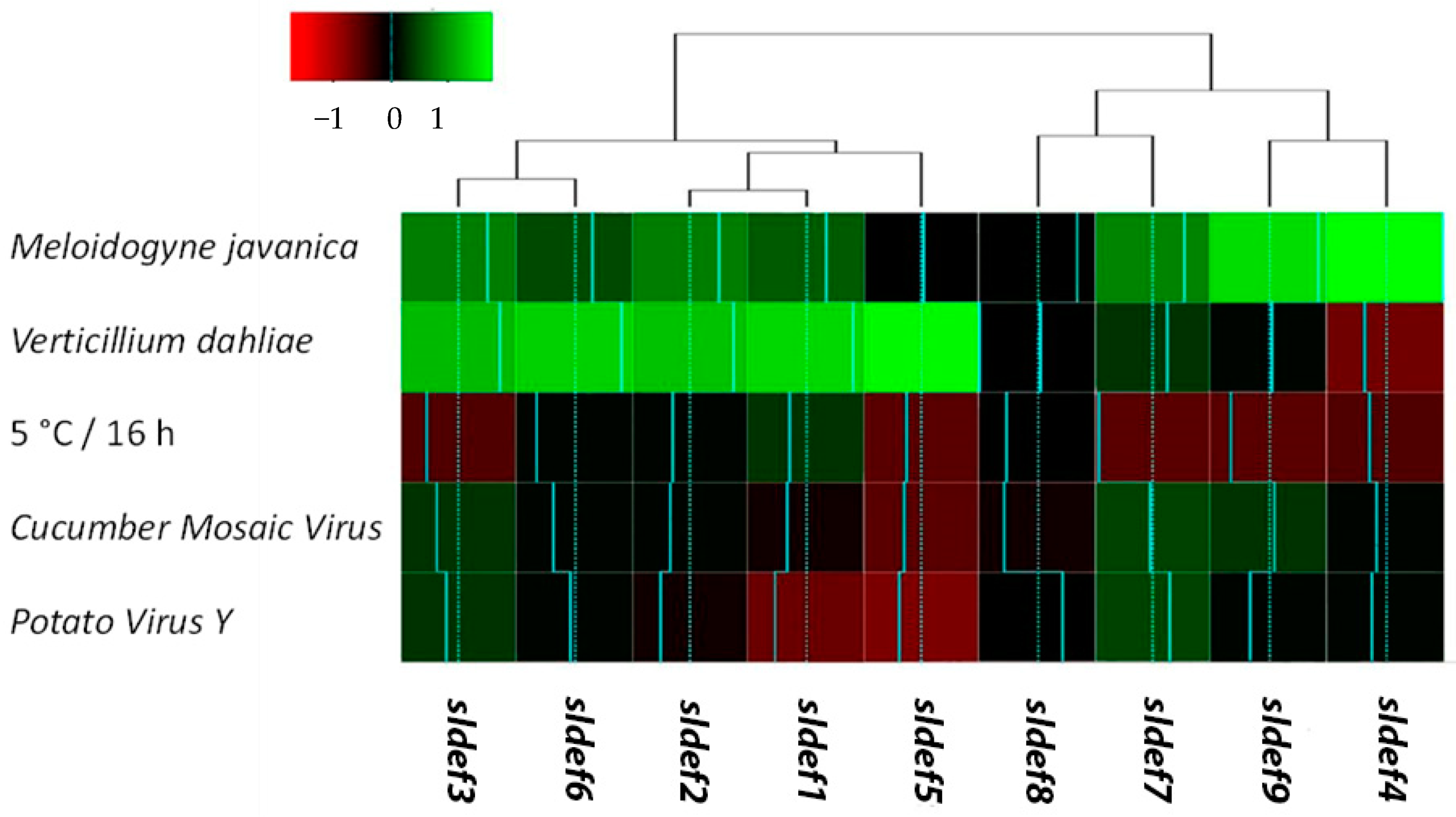
2.2. Defensin Responses Depend on Stimulus
3. Discussion
3.1. Structural Analysis
3.2. Defensins Respond Differentially to Diverse Stimuli
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Mechanical Inoculations with CMV and PVY Viruses
4.3. Nematode Inoculation
4.4. Verticillium Dahliae Preparation and Plant Inoculation
4.5. Verticillium Wilt Disease Assessment and Pathogen Re-Isolation
4.6. In Silico Analyses
4.7. RNA Extraction, Reverse Transcription, qPCR Amplification
4.8. Statistical Design and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CMV | Cucumber Mosaic Virus |
| DAMP | Damage-Associated Molecular Patterns |
| MAMPs | Microbe-Associated Molecular Patterns |
| PVY | Potato Virus Y |
References
- Schneider, D.S.; Ayres, J.S. Two ways to survive infection: What resistance and tolerance can teach us about treating infectious diseases. Nat. Rev. Immunol. 2008, 8, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Meents, A.K.; Mithöfer, A. Plant-plant communication: Is there a role for volatile damage-associated molecular patterns? Front. Plant Sci. 2020, 11, 583275. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, W.F.; Terras, F.R.G.; Cammue, B.P.A.; Osborn, R.W. Plant defensins: Novel antimicrobial peptides as components of the host defense system. Plant Physiol. 1995, 108, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Penninckx, I.A.M.A.; Thomma, B.P.H.J.; Buchala, A.; Métraux, J.-P.; Broekaert, W.F. Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell 1998, 10, 2103–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagan, I.; Garcia-Arenal, F. Tolerance of plants to pathogens: A unifying view. Annu. Rev. Phytopathol. 2020, 58, 77–96. [Google Scholar] [CrossRef]
- Newman, M.A.; Sundelin, T.; Nielsen, J.T.; Erbs, G. MAMP (microbe-associated molecular pattern) triggered immunity in plants. Front. Plant Sci. 2013, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Erbs, G.; Newman, M.A. The role of lipopolysaccharide and peptidoglycan, two glycosylated bacterial microbe-associated molecular patterns (MAMPs), in plant innate immunity. Mol. Plant Pathol. 2012, 13, 95–104. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.-M. Receptor kinases in plant-pathogen interactions: More than pattern recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [Green Version]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K.S. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Sher Khan, R.; Iqbal, A.; Malak, R.; Shehryar, K.; Attia, S.; Ahmed, T.; Ali Khan, M.; Arif, M.; Mii, M. Plant defensins: Types, mechanism of action and prospects of genetic engineering for enhanced disease resistance in plants. 3 Biotech 2019, 9, 192. [Google Scholar] [CrossRef]
- Kovaleva, V.; Bukhteeva, I.; Kit, O.Y.; Nesmelova, I.V. Plant defensins from a structural perspective. Int. J. Mol. Sci. 2020, 21, 5307. [Google Scholar] [CrossRef] [PubMed]
- Berkut, A.A.; Usmanova, D.R.; Peigneur, S.; Oparin, P.B.; Mineev, K.S.; Odintsova, T.I.; Tytgat, J.; Arseniev, A.S.; Grishin, E.V.; Vassilevski, A.A. Structural similarity between defense peptide from wheat and scorpion neurotoxin permits rational functional design. J. Biol. Chem. 2014, 289, 14331–14340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stotz, H.U.; Spence, B.; Wang, Y. A defensin from tomato with dual function in defense and development. Plant Mol. Biol. 2009, 71, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Jiang, N.; Meng, J.; Hou, X.; Yang, G.; Luan, Y. Identification and characterization of defensin genes conferring Phytophthora infestans resistance in tomato. Physiol. Mol. Plant Pathol. 2018, 103, 28–35. [Google Scholar] [CrossRef]
- Stotz, H.U.; Thomson, J.G.; Wang, Y. Plant defensins: Defense, development and application. Plant Signal. Behav. 2009, 4, 1010–1012. [Google Scholar] [CrossRef] [Green Version]
- Lacerda, A.F.; Vasconcelos, E.A.R.; Pelegrini, P.B.; Grossi de Sa, M.F. Antifungal defensins and their role in plant defense. Front. Microbiol. 2014, 5, 116. [Google Scholar] [CrossRef] [Green Version]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118. [Google Scholar] [CrossRef]
- Mitter, N.; Kazan, K.; Way, H.M.; Broekaert, W.F.; Manners, J.M. Systemic induction of an Arabidopsis plant defensin gene promoter by tobacco mosaic virus and jasmonic acid in transgenic tobacco. Plant Sci. 1998, 136, 169–180. [Google Scholar] [CrossRef]
- Saitoh, H.; Kiba, A.; Nishihara, M.; Yamamura, S.; Suzuki, K.; Terauchi, R. Production of antimicrobial defensin in Nicotiana benthamiana with a potato virus X vector. Mol. Plant Microbe Interact. 2001, 14, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Siddique, S.; Wieczorek, K.; Szakasits, D.; Kreil, D.P.; Bohlmann, H. The promoter of a plant defensin gene directs specific expression in nematode-induced syncytia in Arabidopsis roots. Plant Physiol. Biochem. 2011, 49, 1100–1107. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. Proteom. Protoc. Handb. 2005, 571–607. [Google Scholar]
- Ferrè, F.; Clote, P. DiANNA 1.1: An extension of the DiANNA web server for ternary cysteine classification. Nucleic Acids Res. 2006, 34, W182–W185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafee, T.M.A.; Lay, F.T.; Hulett, M.D.; Anderson, M.A. The defensins consist of two independent, convergent protein superfamilies. Mol. Biol. Evol. 2016, 33, 2345–2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lay, F.T.; Anderson, M.A. Defensins—Components of the innate immune system in plants. Curr. Protein Pept. Sci. 2005, 6, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Lay, F.T.; Poon, S.; McKenna, J.A.; Connelly, A.A.; Barbeta, B.L.; McGinness, B.S.; Fox, J.L.; Daly, N.L.; Craik, D.J.; Heath, R.L.; et al. The C-terminal propeptide of a plant defensin confers cytoprotective and subcellular targeting functions. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berrocal-Lobo, M.; Segura, A.; Moreno, M.; López, G.; García-Olmedo, F.; Molina, A. Snakin-2, an antimicrobial peptide from potato whose gene is locally induced by wounding and responds to pathogen infection. Plant Physiol. 2002, 128, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahirñak, V.; Almasia, N.I.; Hopp, H.E.; Vazquez-Rovere, C. Snakin/GASA proteins: Involvement in hormone crosstalk and redox homeostasis. Plant Signal. Behav. 2012, 7, 1004–1008. [Google Scholar] [CrossRef] [Green Version]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A. Defensins of Grasses: A Systematic Review. Biomolecules 2020, 10, 1029. [Google Scholar] [CrossRef]
- Janssen, B.J.C.; Schirra, H.J.; Lay, F.T.; Anderson, M.A.; Craik, D.J. Structure of Petunia hybrida defensin 1, a novel plant defensin with five disulfide bonds. Biochemistry 2003, 42, 8214–8222. [Google Scholar] [CrossRef]
- Ojeda, P.G.; Chan, L.Y.; Poth, A.G.; Wang, C.K.; Craik, D.J. The role of disulfide bonds in structure and activity of chlorotoxin. Future Med. Chem. 2014, 6, 1617–1628. [Google Scholar] [CrossRef] [Green Version]
- Terras, F.R.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; Van Leuven, F.; Vanderleyden, J.; et al. Small cysteine-rich antifungal proteins from radish: Their role in host defense. Plant Cell 1995, 7, 573–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Weerden, N.; Bleackley, M.; Anderson, M. Properties and mechanisms of action of naturally occurring antifungal peptides. Cell. Mol. Life Sci. 2013, 70, 3545–3570. [Google Scholar] [CrossRef] [PubMed]
- Spelbrink, R.G.; Dilmac, N.; Allen, A.; Smith, T.J.; Shah, D.M.; Hockerman, G.H. Differential antifungal and calcium channel-blocking activity among structurally related plant defensins. Plant Physiol. 2004, 135, 2055–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahzad, Z.; Ranwez, V.; Fizames, C.; Marquès, L.; Le Martret, B.; Alassimone, J.; Godé, C.; Lacombe, E.; Castillo, T.; Saumitou-Laprade, P.; et al. Plant defensin type 1 (PDF1): Protein promiscuity and expression variation within the Arabidopsis genus shed light on zinc tolerance acquisition in Arabidopsis halleri. New Phytol. 2013, 200, 820–833. [Google Scholar] [CrossRef]
- Sathoff, A.E.; Velivelli, S.; Shah, D.M.; Samac, D.A. Plant defensin peptides have antifungal and antibacterial activity against human and plant pathogens. Phytopathology 2019, 109, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.-H.; Park, S.; Park, S.; Oh, B.-J.; Back, K.; Han, O.; Kim, J.-I.; Kim, Y.S. Overexpression of a defensin enhances resistance to a fruit-specific anthracnose fungus in pepper. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J.; Cammue, B.P.A.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef]
- Baxter, A.A.; Richter, V.; Lay, F.T.; Poon, I.K.; Adda, C.G.; Veneer, P.K.; Phan, T.K.; Bleackley, M.R.; Anderson, M.A.; Kvansakul, M.; et al. The tomato defensin TPP3 binds phosphatidylinositol (4,5)-bisphosphate via a conserved dimeric cationic grip conformation to mediate cell lysis. Mol. Cell. Biol. 2015, 35, 1964–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lay, F.T.; Mills, G.D.; Poon, I.K.H.; Cowieson, N.P.; Kirby, N.; Baxter, A.A.; Van Der Weerden, N.L.; Dogovski, C.; Perugini, M.A.; Anderson, M.A.; et al. Dimerization of plant defensin NaD1 enhances its antifungal activity. J. Biol. Chem. 2012, 287, 19961–19972. [Google Scholar] [CrossRef] [Green Version]
- Boller, T.; He, S.Y. Innate immunity in plants: An arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–743. [Google Scholar] [CrossRef] [Green Version]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.M.E.; Bleackley, M.R.; Wiltshire, J.L.; Anderson, M.A.; Traven, A.; Van Der Weerden, N.L. Identification and mechanism of action of the plant defensin nad1 as a new member of the antifungal drug arsenal against candida albicans. Antimicrob. Agents Chemother. 2013, 57, 3667–3675. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.S.; Wiens, M.E.; Smith, J.G. Antiviral mechanisms of human defensins. J. Mol. Biol. 2013, 425, 4965–4980. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, Y.M.; McKenna, J.A.; McGinness, B.S.; Hinch, J.; Poon, S.; Connelly, A.A.; Anderson, M.A.; Heath, R.L. Field resistance to Fusarium oxysporum and Verticillium dahliae in transgenic cotton expressing the plant defensin NaD1. J. Exp. Bot. 2014, 65, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.W.; Huffaker, A.; Crippen, D.; Robbins, R.T.; Goggin, F.L. Plant elicitor peptides promote plant defences against nematodes in soybean. Mol. Plant Pathol. 2018, 19, 858–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiegel, Y.; McClure, M.A. The surface coat of plant-parasitic nematodes: Chemical composition, origin, and biological role—A review. J. Nematol. 1995, 27, 127–134. [Google Scholar]
- Goverse, A.; Smant, G. The activation and suppression of plant innate immunity by parasitic nematodes. Annu. Rev. Phytopathol. 2014, 52, 243–265. [Google Scholar] [CrossRef]
- Cooper, D.; Eleftherianos, I. Parasitic nematode immunomodulatory strategies: Recent advances and perspectives. Pathogens 2016, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Roberts, K.; Love, A.J.; Laval, V.; Laird, J.; Tomos, A.D.; Hooks, M.A.; Milner, J.J. Long-distance movement of Cauliflower mosaic virus and host defence responses in Arabidopsis follow a predictable pattern that is determined by the leaf orthostichy. New Phytol. 2007, 175, 707–717. [Google Scholar] [CrossRef]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. New Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef] [Green Version]
- Calil, I.P.; Fontes, E.P.B. Plant immunity against viruses: Antiviral immune receptors in focus. Ann. Bot. 2017, 119, 711–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, L.C. Induced resistance in plants and the role of pathogenesis-related proteins. Eur. J. Plant Pathol. 1997, 103, 753–765. [Google Scholar] [CrossRef]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212–213, 29–37. [Google Scholar] [CrossRef]
- Koike, M.; Okamoto, T.; Tsuda, S.; Imai, R. A novel plant defensin-like gene of winter wheat is specifically induced during cold acclimation. Biochem. Biophys. Res. Commun. 2002, 298, 46–53. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Kasianov, A.S.; Kovtun, A.S.; Makeev, V.J.; Shcherbakova, L.A.; Kudryavtsev, A.M. Defensin-like peptides in wheat analyzed by whole-transcriptome sequencing: A focus on structural diversity and role in induced resistance. PeerJ 2019, 2019, 1–29. [Google Scholar] [CrossRef]
- Nie, X. Reverse transcription loop-mediated isothermal amplification of DNA for detection of potato virus Y. Plant Dis. 2005, 89, 605–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumford, R.A.; Barker, I.; Wood, K.R. The detection of tomato spotted wilt virus using the polymerase chain reaction. J. Virol. Methods 1994, 46, 303–311. [Google Scholar] [CrossRef]
- Faggioli, F.; Ferretti, L.; Albanese, G.; Sciarroni, R.; Pasquini, G.; Lumia, V.; Barba, M. Distribution of olive tree viruses in Italy as revealed by one-step RT-PCR. J. Plant Pathol. 2005, 87, 49–55. [Google Scholar]
- Dovas, C.I.; Efthimiou, K.; Katis, N.I. Generic detection and differentiation of tobamoviruses by a spot nested RT-PCR-RFLP using dI-containing primers along with homologous dG-containing primers. J. Virol. Methods 2004, 117, 137–144. [Google Scholar] [CrossRef]
- Hussey, R.S.; Barker, K.R. A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Report. 1973, 57, 1025–1028. [Google Scholar]
- Bridge, J.; Page, S.L.J. Estimation of root-knot nematode infestation levels on roots using a rating chart. Trop. Pest Manag. 1980, 26, 296–298. [Google Scholar] [CrossRef]
- Ligoxigakis, E.K.; Markakis, E.A. Incidence and pathogenicity of races and isolates of Verticillium dahliae in Crete, southern Greece. Phytoparasitica 2012, 40, 493–506. [Google Scholar] [CrossRef]
- Pegg, G.; Brady, B. Verticillium Wilts; CABI Publishing: Egham, UK; Boston, MA, USA, 2002. [Google Scholar]
- Van den Heuvel, K.J.P.T.; Hulzink, J.M.R.; Barendse, G.W.M.; Wullems, G.J. The expression of tgas118, encoding a defensin in Lycopersicon esculentum, is regulated by gibberellin. J. Exp. Bot. 2001, 52, 1427–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm, L.; Rosenström, P. Dali server: Conservation mapping in 3D. Nucleic Acids Res. 2010, 38, W545–W549. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, K.B. GeneDoc: Analysis and visualization of genetic variation. EMBNEW News 1997, 4, 14. [Google Scholar]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Tsaniklidis, G.; Pappi, P.; Tsafouros, A.; Charova, S.N.; Nikoloudakis, N.; Roussos, P.A.; Paschalidis, K.A.; Delis, C. Polyamine homeostasis in tomato biotic/abiotic stress cross-tolerance. Gene 2020, 727. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a/a | Acronym | NCBI Accession (Nucleotide/Protein) | SOLYC Number | Motif | Theoretical PI | Cys | Predicted Connectivity | Secretion Probability |
|---|---|---|---|---|---|---|---|---|
| 1 | SlDEF1 | NM_001247943/NP_001234872 | Solyc07g007750.3.1 | C-X10-C-X5-C-X3-C-X9-C-X6-C-X1-C-X3-C | 8.96 | 8 | 1–8, 2–5, 3–6, 4–7 | 0.934 |
| 2 | SlDEF2a | XR_003246738/XP_025886757 | Solyc05g150107.1.1 | C-X5-C-X4-C-X6-C-X4-C-X1-C-X11-C-X5-C- C-X3-C-X6-C-X4-C-X1-C | 6.76 | 12 | 1–10, 2–5, 3–8, 4–7, 6–9 | 0.981 |
| 3 | SlDEF2b | XR_003246738/XP_019069532 | Solyc05g150107.1.1 | C-X5-C-X4-C-X6-C-X4-C-X1-C-X11-C-X5-C- C-X3-C-X6-C-X4-C-X1-C | 6.76 | 12 | 1–10, 2–5, 3–8, 4–7, 6–9 | 0.957 |
| 4 | SlDEF3 | NM_001328663/NP_001315592 | Solyc07g006380.3.1 | C-X10-C-X5-C-X3-C-X9-C-X6-C-X1-C-X3-C | 6.56 | 8 | 1–8, 2–5, 3–6, 4–7 | 0.989 |
| 5 | SlDEF4 | XM_004242790/XP_004242838 | Solyc07g009060.4.1 | C-X10-C-X5-C-X3-C-X9-C-X16-C-X1-C-X3-C | 9.25 | 8 | 1–2, 3–6, 4–5, 7–8 | 0.969 |
| 6 | SlDEF5 | XM_010321070/XP_010319372 | Solyc04g008470.3.1 | C-X10-C-X5-C-X3-C-X9-C-X6-C-X1-C-X3-C | 9.54 | 8 | 1–8, 2–5, 3–6, 4–7 | 0.984 |
| 7 | SlDEF6 | XM_004242803/XP_004242851 | Solyc07g009260.3.1 | C-X10-C-X5-C-X3-C-X9-C-X9-C-X1-C-X3-C | 8.99 | 8 | 1–8, 2–5, 3–6, 4–7 | 0.883 |
| 8 | SlDEF7 | NM_001310317/NP_001297246 | Solyc07g007710.4.1 | C-X10-C-X5-C-X3-C-X9-C-X6-C-X1-C-X3-C | 8.98 | 8 | 1–8, 2–5, 3–6, 4–7 | 0.954 |
| 9 | SlDEF8 | NM_001310318/NP_001297247 | Solyc07g007730.4.1 | C-X10-C-X5-C-X3-C-X9-C-X6-C-X1-C-X3-C | 8.77 | 8 | 1–8, 2–5, 3–6, 4–7 | 0.980 |
| 10 | SlDEF9 | NM_001346524/NP_001333453 | Solyc07g007755.1.1 | C-X10-C-X5-C-X3-C-X9-C-X6-C-X1-C-X3-C | 9.14 | 8 | 1–8, 2–5, 3–6, 4–7 | 0.951 |
| a/a | Code | NCBI Accession (P) | SOLYC Number | Confidence | Coverage | Template |
|---|---|---|---|---|---|---|
| 1 | SlDEF1 | NP_001234872 | Solyc07g007750.3.1 | 100% | 100% | c2lr3A 1 |
| 2 | SlDEF2a | XP_025886757 | Solyc05g150107.1.1 | 86.3% | 28% | d1f2si 2 |
| 3 | SlDEF2b | XP_019069532 | Solyc05g150107.1.1 | 85.9% | 27% | c1f2sI 2 |
| 4 | SlDEF3 | NP_001315592 | Solyc07g006380.3.1 | 99.9% | 100% | c4uj0B 1 |
| 5 | SlDEF4 | XP_004242838 | Solyc07g009060.4.1 | 96.2% | 83% | c2n2qA 1 |
| 6 | SlDEF5 | XP_010319372 | Solyc04g008470.3.1 | 99.9% | 100% | c2lr3A 1 |
| 7 | SlDEF6 | XP_004242851 | Solyc07g009260.3.1 | 100% | 98% | d1bk8a 1 |
| 8 | SlDEF7 | NP_001297246 | Solyc07g007710.4.1 | 99.9% | 100% | c2lr3A 1 |
| 9 | SlDEF8 | NP_001297247 | Solyc07g007730.4.1 | 100% | 100% | c2lr3A 1 |
| 10 | SlDEF9 | NP_001333453 | Solyc07g007755.1.1 | 100% | 100% | c2lr3A 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikoloudakis, N.; Pappi, P.; Markakis, E.A.; Charova, S.N.; Fanourakis, D.; Paschalidis, K.; Delis, C.; Tzortzakakis, E.A.; Tsaniklidis, G. Structural Diversity and Highly Specific Host-Pathogen Transcriptional Regulation of Defensin Genes Is Revealed in Tomato. Int. J. Mol. Sci. 2020, 21, 9380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249380
Nikoloudakis N, Pappi P, Markakis EA, Charova SN, Fanourakis D, Paschalidis K, Delis C, Tzortzakakis EA, Tsaniklidis G. Structural Diversity and Highly Specific Host-Pathogen Transcriptional Regulation of Defensin Genes Is Revealed in Tomato. International Journal of Molecular Sciences. 2020; 21(24):9380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249380
Chicago/Turabian StyleNikoloudakis, Nikolaos, Polyxeni Pappi, Emmanouil A. Markakis, Spyridoula N. Charova, Dimitrios Fanourakis, Konstantinos Paschalidis, Costas Delis, Emmanuel A. Tzortzakakis, and Georgios Tsaniklidis. 2020. "Structural Diversity and Highly Specific Host-Pathogen Transcriptional Regulation of Defensin Genes Is Revealed in Tomato" International Journal of Molecular Sciences 21, no. 24: 9380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249380