Overexpression of Soybean-Derived Lunasin in Wheat and Assessment of Its Anti-Proliferative Activity in Colorectal Cancer HT-29 Cells

 ,
,
Abstract
:1. Introduction
2. Results
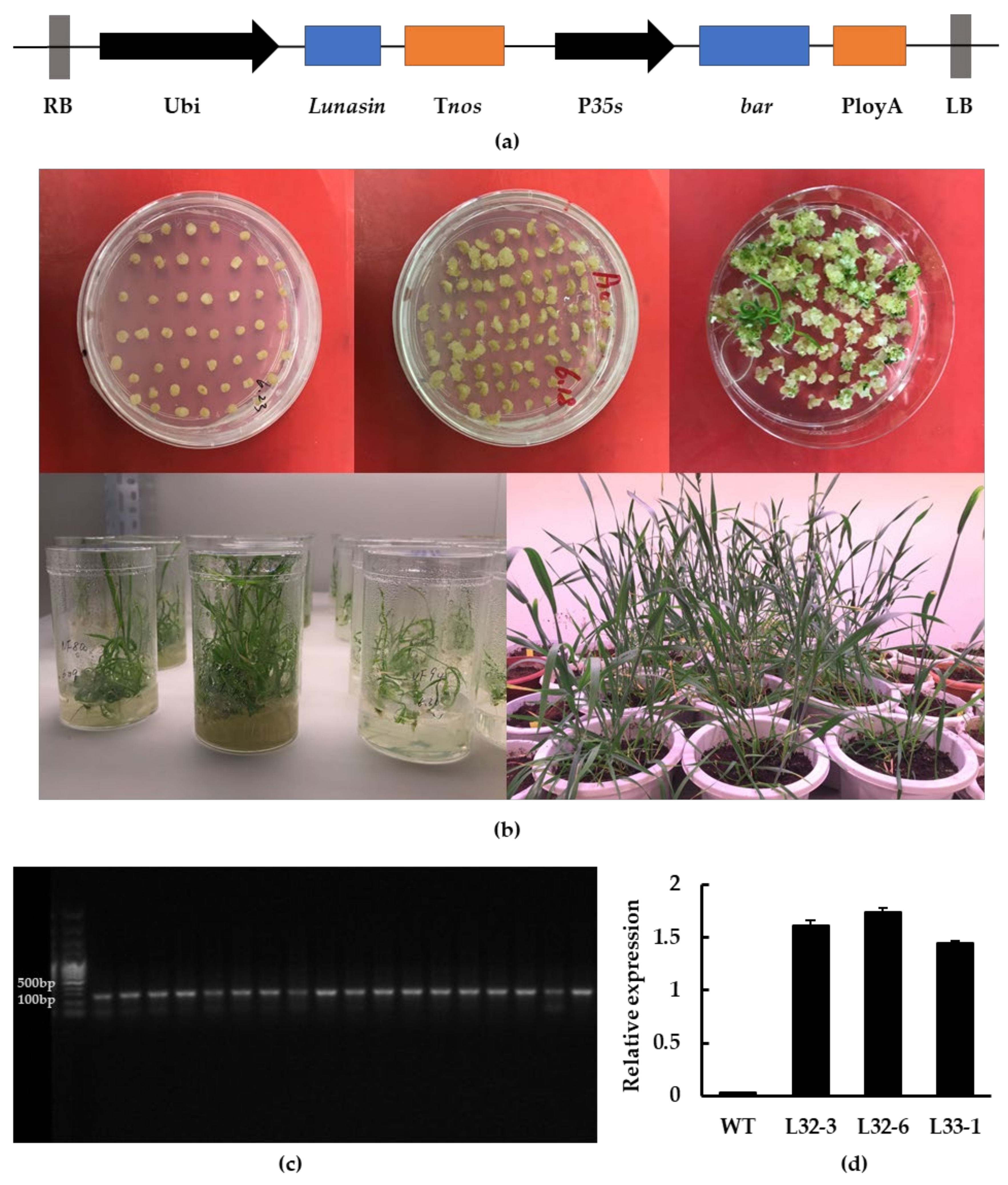
2.1. Molecular Analysis of Transgenic Wheat
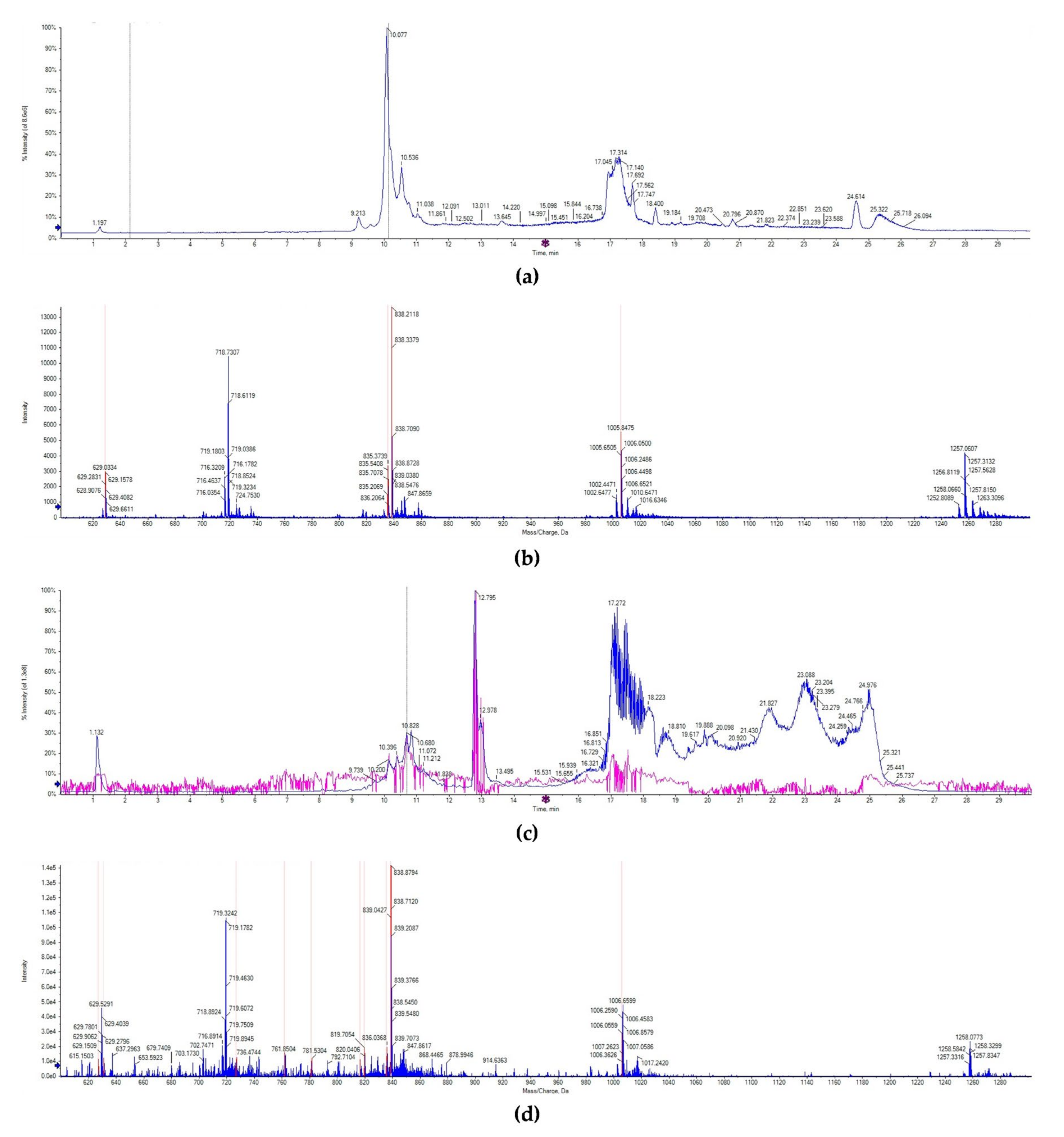
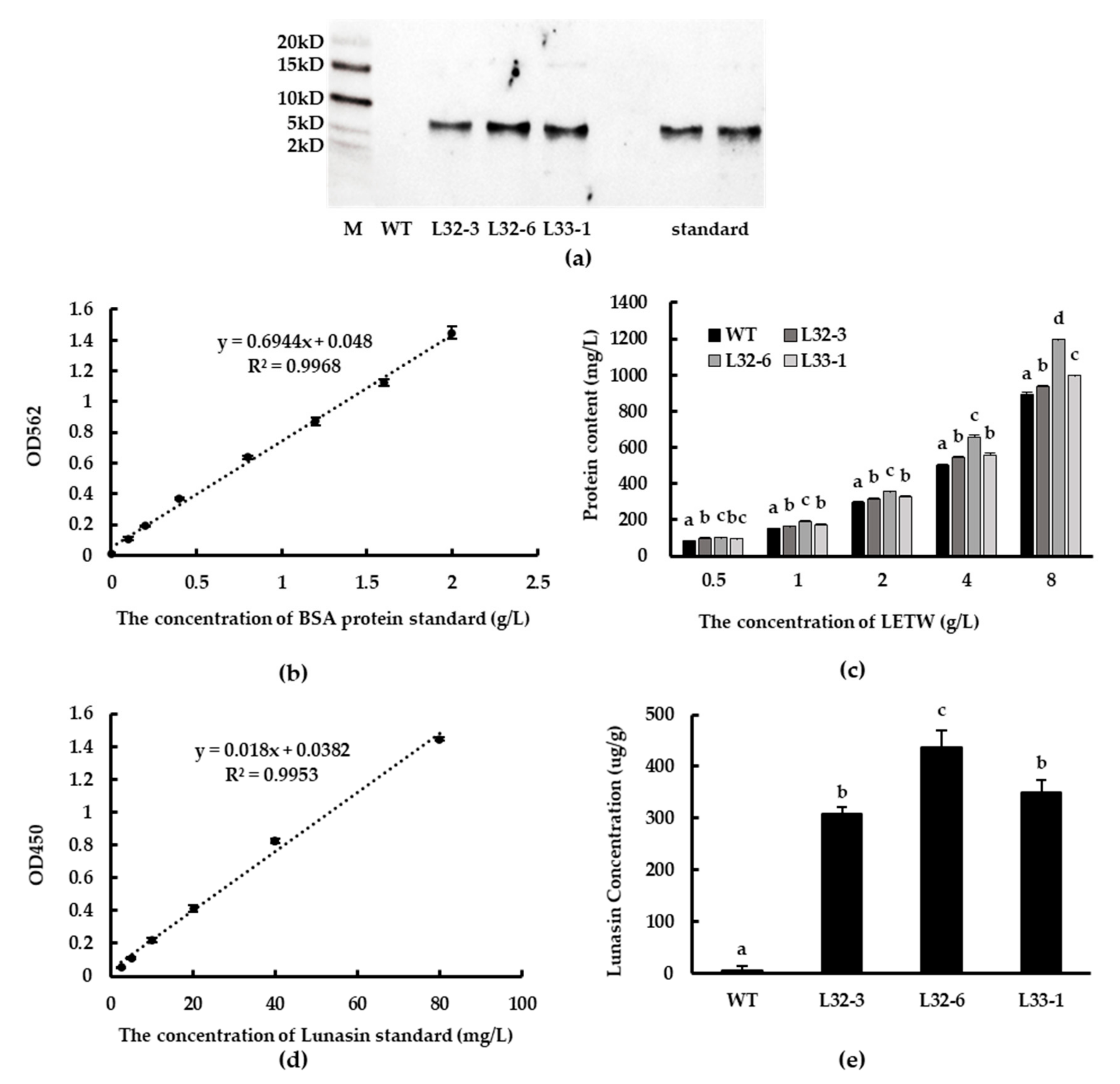
2.2. Identification and Content of Lunasin in Transgenic Wheat
2.3. Effects of LETW on Cell Proliferation in HT-29
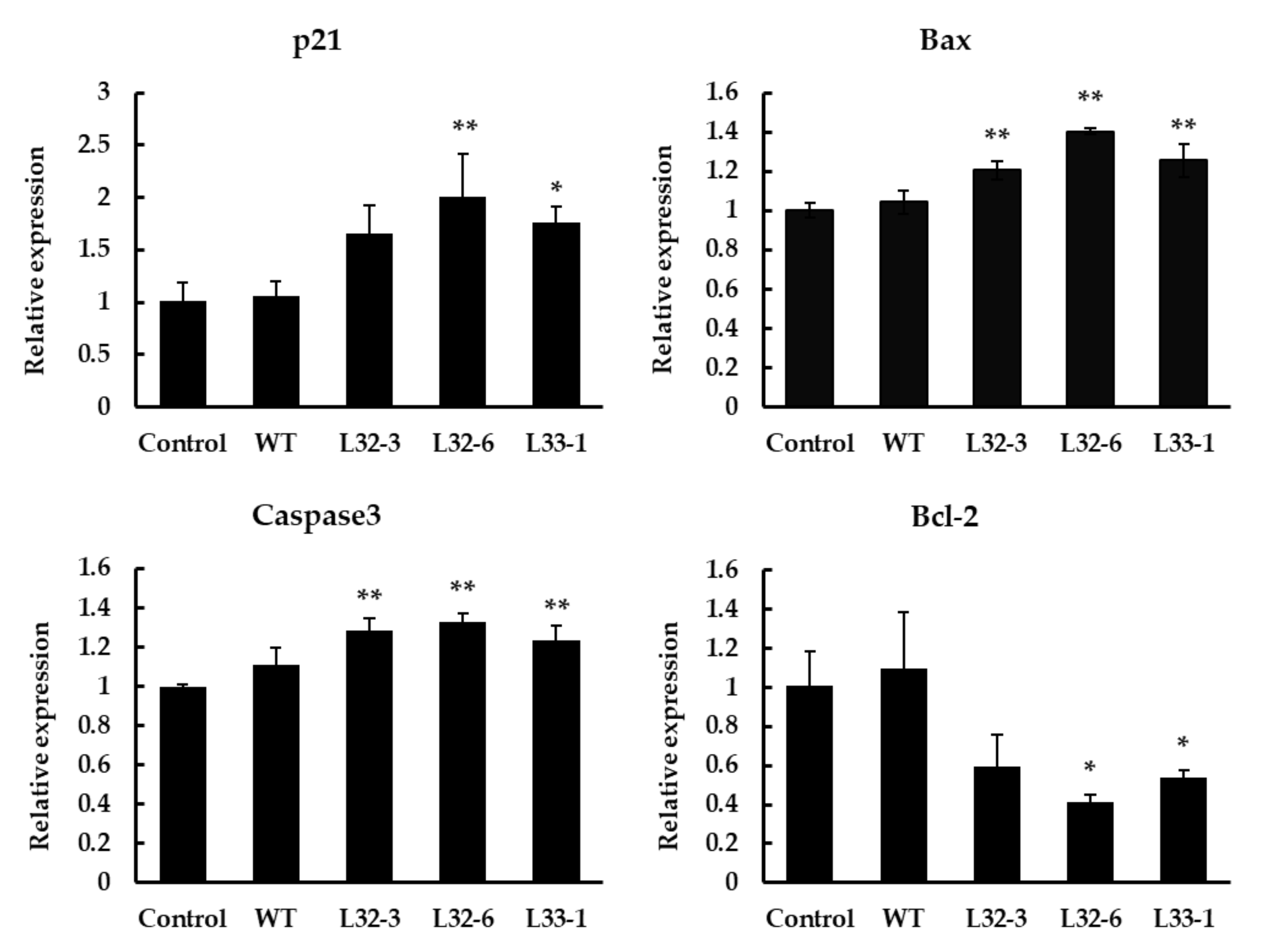
2.4. Expression Analysis of Apoptosis-Related Genes under LETW Treatment in HT-29
3. Discussion
4. Materials and Methods
4.1. Regents
4.2. Construction of the Expression Vector
4.3. Agrobacterium-Mediated Transformation of Wheat Immature Embryos
4.4. Screening for Transgenic Wheat
4.5. Isolation, Purification, and Identification of Lunasin from Wheat
4.6. UPLC-MS/MS Analysis
4.7. Lunasin Quantification by ELISA
4.8. Cell Proliferation Assay
4.9. Real-Time Quantitative PCR (RT-qPCR)
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jeong, H.J.; Jeong, J.B.; Kim, D.S.; Park, J.H.; Lee, J.B.; Kweon, D.-H.; Chung, G.Y.; Seo, E.-W.; De Lumen, B.O. The cancer preventive peptide lunasin from wheat inhibits core histone acetylation. Cancer Lett. 2007, 255, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Hsieh, C.-C.; De Lumen, B.O. Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 390, 803–808. [Google Scholar] [CrossRef] [PubMed]
- De Mejia, E.G.; Dia, V.P. Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides 2009, 30, 2388–2398. [Google Scholar] [CrossRef] [PubMed]
- Achuthan, R.; Bell, S.M.; Roberts, P.; Leek, J.P.; Horgan, K.; Markham, A.F.; MacLennan, K.A.; Speirs, V. Genetic events during the transformation of a tamoxifen-sensitive human breast cancer cell line into a drug-resistant clone. Cancer Genet. Cytogenet. 2001, 130, 166–172. [Google Scholar] [CrossRef]
- Galvez, A.F.; De Lumen, B.O. A soybean cDNA encoding a chromatin-binding peptide inhibits mitosis of mammalian cells. Nat. Biotechnol. 1999, 17, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.; Galvez, A.; De Lumen, B.O. Lunasin™ Suppresses E1A-Mediated Transformation of Mammalian Cells but Does Not Inhibit Growth of Immortalized and Established Cancer Cell Lines. Nutr. Cancer 2003, 47, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Park, J.H.; Lam, Y.; De Lumen, B.O. Characterization of Lunasin Isolated from Soybean. J. Agric. Food Chem. 2003, 51, 7901–7906. [Google Scholar] [CrossRef] [PubMed]
- Dia, V.P.; De Mejia, E.G. Lunasin promotes apoptosis in human colon cancer cells by mitochondrial pathway activation and induction of nuclear clusterin expression. Cancer Lett. 2010, 295, 44–53. [Google Scholar] [CrossRef]
- Dia, V.P.; De Mejia, E.G. Lunasin induces apoptosis and modifies the expression of genes associated with extracellular matrix and cell adhesion in human metastatic colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 623–634. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Xu, F.; Han, Y.; Hernández-Ledesma, B.; Xiao, H. Inhibitory Effects of Peptide Lunasin in Colorectal Cancer HCT-116 Cells and Their Tumorsphere-Derived Subpopulation. Int. J. Mol. Sci. 2020, 21, 537. [Google Scholar] [CrossRef] [Green Version]
- Dinelli, G.; Bregola, V.; Bosi, S.; Fiori, J.; Gotti, R.; Simonetti, E.; Trozzi, C.; Leoncini, E.; Prata, C.; Massaccesi, L.; et al. Lunasin in wheat: A chemical and molecular study on its presence or absence. Food Chem. 2014, 151, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Pigolev, A.V.; Miroshnichenko, D.; Pushin, A.; Terentyev, V.V.; Boutanaev, A.M.; Dolgov, S.; Savchenko, T.V. Overexpression of Arabidopsis OPR3 in Hexaploid Wheat (Triticum aestivum L.) Alters Plant Development and Freezing Tolerance. Int. J. Mol. Sci. 2018, 19, 3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 Transcription Factor Enhances Heat and Drought Stress Tolerance in Wheat (Triticum aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayadi, M.; Brini, F.; Masmoudi, K. Overexpression of a Wheat Aquaporin Gene, TdPIP2;1, Enhances Salt and Drought Tolerance in Transgenic Durum Wheat cv. Maali. Int. J. Mol. Sci. 2019, 20, 2389. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, Y.; Sun, F.; Li, X.; Wang, P.; Chang, J.; Wang, Y.; Yang, G.; He, G. Co-expression of high-molecular-weight glutenin subunit 1Ax1 and Puroindoline a (Pina) genes in transgenic durum wheat (Triticum turgidum ssp. durum) improves milling and pasting quality. BMC Plant Biol. 2019, 19, 126. [Google Scholar] [CrossRef]
- Du, X.; Wei, J.; Luo, X.; Liu, Z.; Qian, Y.; Zhu, B.; Weng, Q.; Tang, H. Low-molecular-weight glutenin subunit LMW-N13 improves dough quality of transgenic wheat. Food Chem. 2020, 327, 127048. [Google Scholar] [CrossRef]
- Chen, H.; Li, S.; Liu, Y.; Liu, J.; Ma, X.; Du, L.; Wang, K.; Ye, X. Effects of 1Dy12 subunit silencing on seed storage protein accumulation and flour-processing quality in a common wheat somatic variation line. Food Chem. 2021, 335, 127663. [Google Scholar] [CrossRef]
- Abouseadaa, H.; Osman, G.; Ramadan, A.M.; Hassan, S.M.; Abdelsattar, M.; Morsy, Y.B.; Alameldin, H.F.; El-Ghareeb, D.K.; Nour-Eldin, H.A.; Salem, R.; et al. Development of transgenic wheat (Triticum aestivum L.) expressing avidin gene conferring resistance to stored product insects. BMC Plant Biol. 2015, 15, 183. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Gao, C.; Chen, C.; Zhang, W.; Huang, X.-Y.; Zhao, F.-J. Overexpression of Rice OsHMA3 in Wheat Greatly Decreases Cadmium Accumulation in Wheat Grains. Environ. Sci. Technol. 2020, 54, 10100–10108. [Google Scholar] [CrossRef]
- Liu, C.-F.; Pan, T.-M. Recombinant expression of bioactive peptide lunasin in Escherichia coli. Appl. Microbiol. Biotechnol. 2010, 88, 177–186. [Google Scholar] [CrossRef]
- Kyle, S.; James, K.A.; McPherson, M.J. Recombinant production of the therapeutic peptide lunasin. Microb. Cell Fact. 2012, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Nadia, E.; Yao, Y.; Shi, Z.; Ren, G. Tandem repeated expression of lunasin gene in Pichia pastoris and its anti-inflammatory activity in vitro. J. Biosci. Bioeng. 2018, 126, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Hao, Y.; Zhu, Y.; Shi, Z.; Zhao, G. Expression of Bioactive Lunasin Peptide in Transgenic Rice Grains for the Application in Functional Food. Molecules 2018, 23, 2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.; Fan, X.; Guo, H.; Yao, Y.; Ren, G.; Lv, X.; Yang, X. Overexpression of the bioactive lunasin peptide in soybean and evaluation of its anti-inflammatory and anti-cancer activities in vitro. J. Biosci. Bioeng. 2020, 129, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, H.; Du, L.; Ye, X. Generation of marker-free transgenic hexaploid wheat via anAgrobacterium-mediated co-transformation strategy in commercial Chinese wheat varieties. Plant Biotechnol. J. 2017, 15, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.; Sung, C.K.; Im, Y.J.; Chun, C. Activation of JNK and p38 in MCF-7 Cells and the In Vitro Anticancer Activity of Alnus hirsuta Extract. Molecules 2020, 25, 1073. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Youle, R.J. The Role of Mitochondria in Apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-P.; Yu, X.-D.; Sun, Y.-W.; Jones, H.D.; Xia, L. Generation of Marker- and/or Backbone-Free Transgenic Wheat Plants via Agrobacterium-Mediated Transformation. Front. Plant Sci. 2016, 7, 1324. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.A.; Lovegrove, A.; Shewry, P.R. Lunasin in cereal seeds: What is the origin? J. Cereal Sci. 2013, 57, 267–269. [Google Scholar] [CrossRef] [Green Version]
- Alaswad, A.A.; Krishnan, H.B. Immunological Investigation for the Presence of Lunasin, a Chemopreventive Soybean Peptide, in the Seeds of Diverse Plants. J. Agric. Food Chem. 2016, 64, 2901–2909. [Google Scholar] [CrossRef]
- Johnson, I. Anticarcinogenic effects of diet-related apoptosis in the colorectal mucosa. Food Chem. Toxicol. 2002, 40, 1171–1178. [Google Scholar] [CrossRef]
- De Mejía, E.G.; Wang, W.; Dia, V.P. Lunasin, with an arginine-glycine-aspartic acid motif, causes apoptosis to L1210 leukemia cells by activation of caspase-3. Mol. Nutr. Food Res. 2009, 54, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Tsunashima, M.; Hiei, Y.; Komari, T. Wheat (Triticum aestivum L.) Transformation Using Immature Embryos. Bioinform. MicroRNA Res. 2015, 1223, 189–198. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Gene | Sequence (5′—3′) |
|---|---|---|
| Lunasin—F | Lunasin | ATGTCCAAGTGGCAGCACCAG |
| Lunasin—R | Lunasin | TCAGTCATCGTCATCGTCGTCAT |
| Lunasin—QF | Lunasin | AGCACCAGCAGCAGGATT |
| Lunasin—QR | Lunasin | GCCCTGAATCTTCTCCATGA |
| p21—F | p21 | ATGAAATTCACCCCCTTTCC |
| p21—R | p21 | AGGTGAGGGGACTCCAAAGT |
| Bax—F | Bax | CAAACTGGTGCTCAAGGCCC |
| Bax—R | Bax | CCGGAGGAAGTCCAATGTCC |
| Bcl-2—F | Bcl-2 | TGGGATTCCTGCGGATTGAC |
| Bcl-2—R | Bcl-2 | GTCTACTTCCTCTGTGATGTTGT |
| caspase-3—F | caspase-3 | AGCGAATCAATGGACTCTGG |
| caspase-3—R | caspase-3 | CCGAGATGTCATTCCAGTGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Qin, P.; Hao, Y.; Guo, H.; Blecker, C.; Everaert, N.; Ren, G. Overexpression of Soybean-Derived Lunasin in Wheat and Assessment of Its Anti-Proliferative Activity in Colorectal Cancer HT-29 Cells. Int. J. Mol. Sci. 2020, 21, 9594. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249594
Fan X, Qin P, Hao Y, Guo H, Blecker C, Everaert N, Ren G. Overexpression of Soybean-Derived Lunasin in Wheat and Assessment of Its Anti-Proliferative Activity in Colorectal Cancer HT-29 Cells. International Journal of Molecular Sciences. 2020; 21(24):9594. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249594
Chicago/Turabian StyleFan, Xin, Peiyou Qin, Yuqiong Hao, Huimin Guo, Christophe Blecker, Nadia Everaert, and Guixing Ren. 2020. "Overexpression of Soybean-Derived Lunasin in Wheat and Assessment of Its Anti-Proliferative Activity in Colorectal Cancer HT-29 Cells" International Journal of Molecular Sciences 21, no. 24: 9594. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249594