Identification and Analysis of MicroRNAs Associated with Wing Polyphenism in the Brown Planthopper, Nilaparvata lugens

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Results
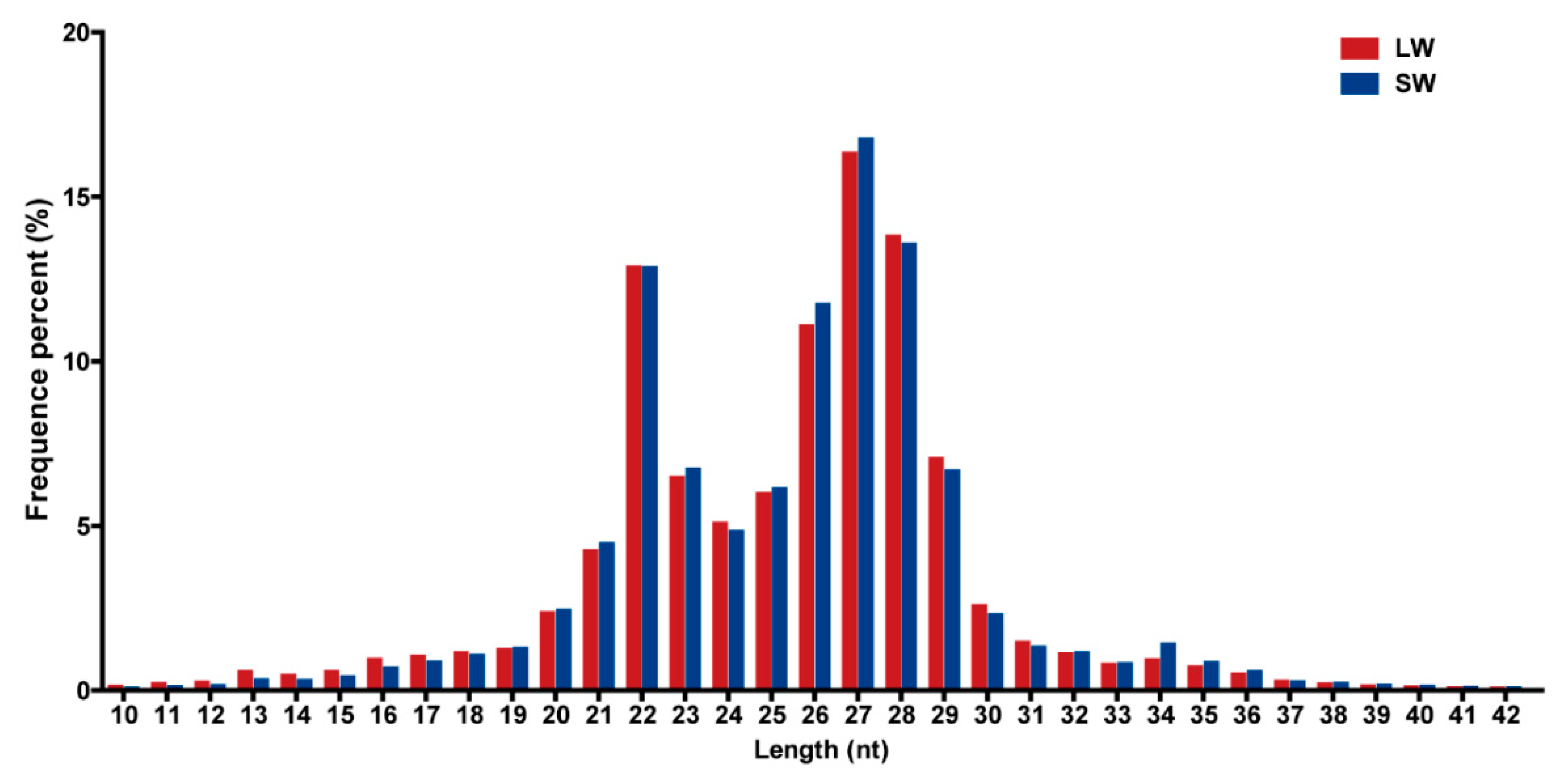
2.1. Identification of miRNAs in BPH
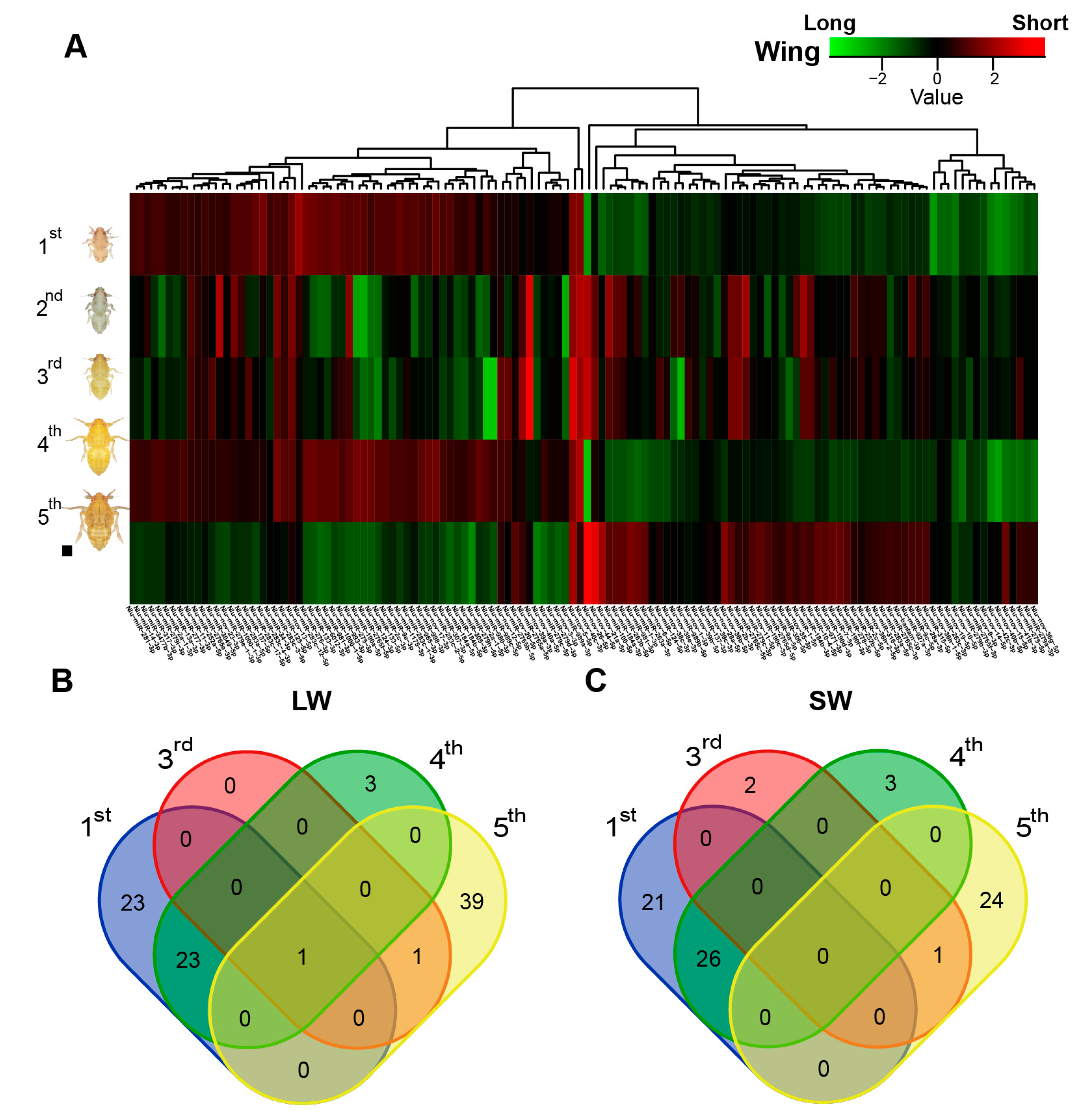
2.2. Identification of Wing Morph-Related miRNAs in BPH
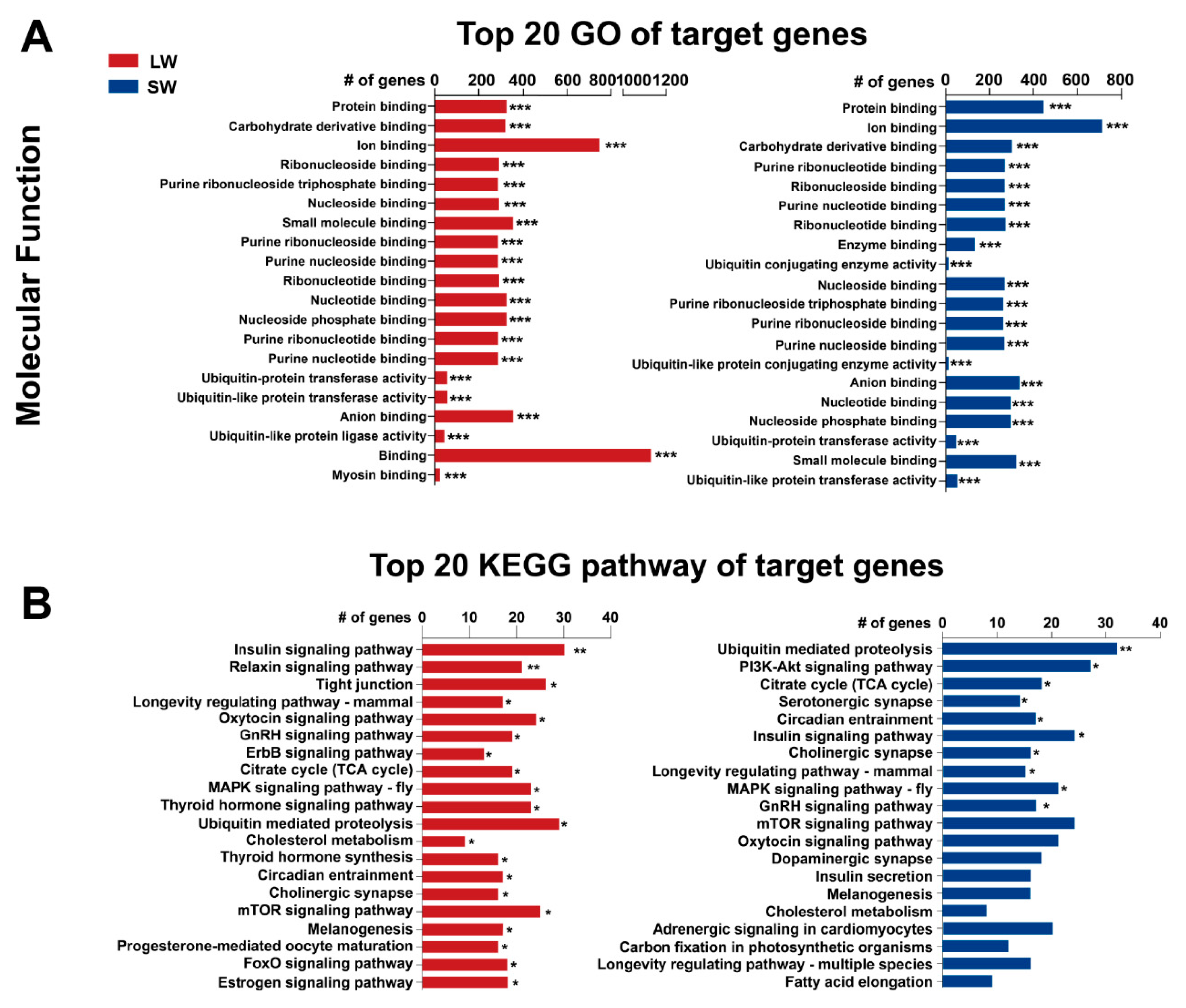
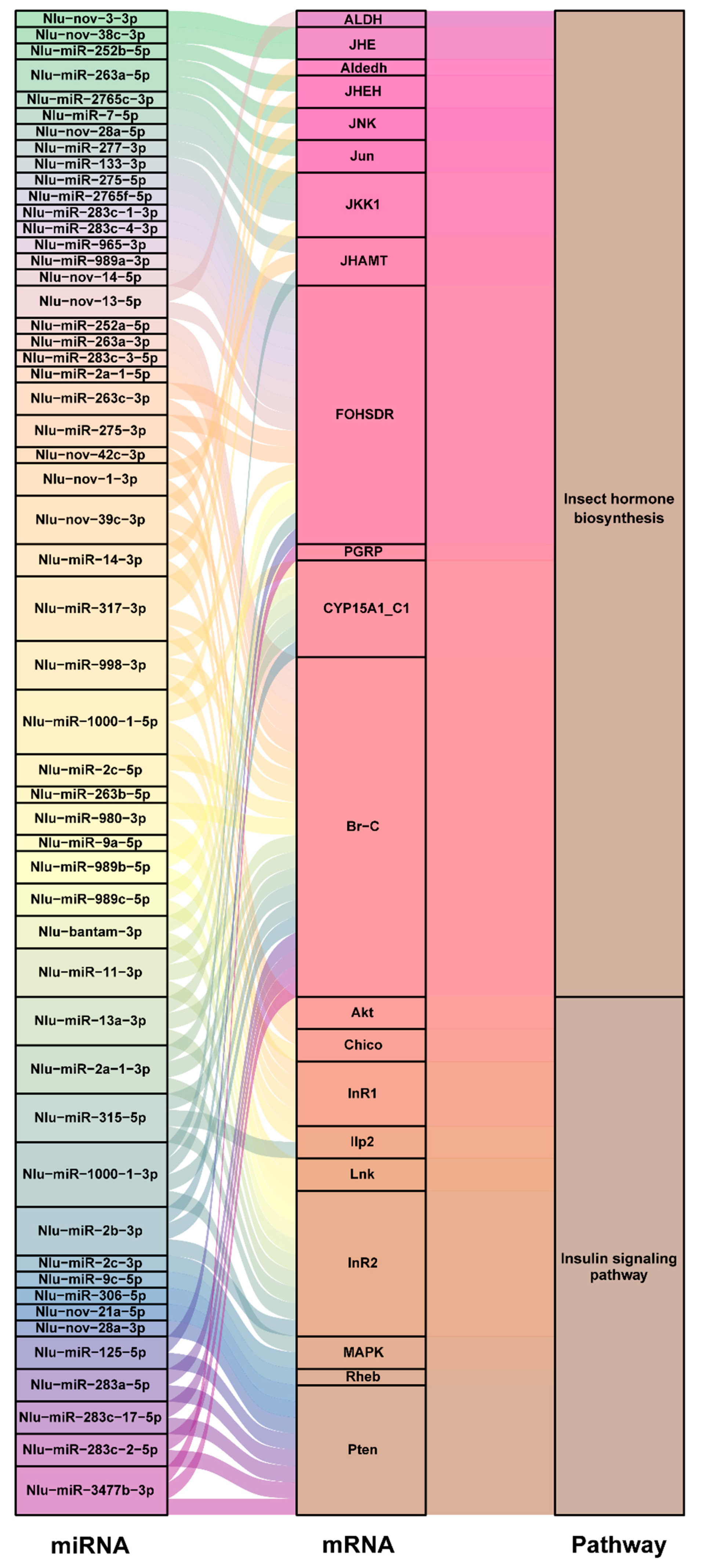
2.3. Functional Analysis of Differentially Expressed miRNAs
2.4. Validation of miRNAs and Target Gene
3. Discussion
4. Materials and Methods
4.1. Insect Rearing
4.2. Small RNA Library Preparation and Sequencing
4.3. miRNA Prediction
4.4. Differential Expression Analysis
4.5. Target Gene Prediction and GO and KEGG Enrichment Analysis
4.6. In Vitro Luciferase Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LW | Long-wing |
| SW | Short-wing |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| BPH | Brown planthopper |
| NCBI | National Center for Biotechnology Information |
| Akt | Serine/threonine-specific protein kinase |
| Aldedh | Aldehyde dehydrogenase cytosolic 2-like |
| ALDH | Aldehyde dehydrogenase dimeric NADP preferring |
| Br-C | Broad-complex core protein |
| Chico | Insulin receptor substrate |
| CYP15A1_C1 | Methyl farnesoate epoxidase-like |
| FOHSDR | Farnesol dehydrogenase-like |
| GnRH | Gonadotropin-releasing hormone |
| IIS | Insulin/insulin-like growth factor signaling |
| Ilp2 | Insulin-like peptide |
| InR1 | Insulin receptor 1 |
| InR2 | Insulin receptor 2 |
| JH | Juvenile hormone |
| JHAMT | Juvenile hormone-III synthase |
| JHE | Juvenile hormone esterase |
| JHEH | Juvenile hormone epoxide hydrolase |
| JKK1 | Dual specificity mitogen-activated protein kinase kinase 7-like |
| JNK | Mitogen-activated protein kinase 8/9/10 (c-Jun N-terminal kinase) |
| JUN | Transcription factor AP-1 |
| Lnk | SH2B adapter protein 1-like protein |
| MAPK | Mitogen-activated protein kinase |
| PGRP | Peptidoglycan recognition protein |
| Pten | Phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase |
| Rheb | Ras homolog enriched in brain |
References
- Simpson, S.J.; Sword, G.A.; Lo, N. Polyphenism in insects. Curr. Biol. 2011, 21, R738–R749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brakefield, P.M.; Frankino, W.A. Polyphenisms in Lepidoptera: Multidisciplinary approaches to studies of evolution and development. In Phenotypic Plasticity of Insects: Mechanisms and Consequences, 1st ed.; Whitman, D.W., Ananthakrishnan, T.N., Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 337–368. [Google Scholar]
- Huber, F. New Observations on the Natural History of Bees, 3rd ed.; Pub. W. & C. Tait: Edinburgh, UK, 1821. [Google Scholar]
- Haydak, M.H. Honey bee nutrition. Annu. Rev. Entomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism: An update. Adv. Insect Physiol. 2009, 36, 1–272. [Google Scholar]
- Wang, X.; Kang, L. Molecular mechanisms of phase change in locusts. Annu. Rev. Entomol. 2014, 59, 225–244. [Google Scholar] [CrossRef] [Green Version]
- Zera, A.J. Wing polymorphism in Gryllus (Orthoptera: Gryllidae): Proximate endocrine, energetic and biochemical mechanisms underlying morph specialization for flight vs. reproduction. In Phenotypic Plasticity of Insects: Mechanisms and Consequences, 1st ed.; Whitman, D.W., Ananthakrishnan, T.N., Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 609–653. [Google Scholar]
- Zera, A.J.; Denno, R.F. Physiology and ecology of dispersal polymorphism in insects. Annu. Rev. Entomol. 1997, 42, 207–230. [Google Scholar] [CrossRef] [Green Version]
- Roff, D.A. The evolution of wing dimorphism in insects. Evolution 1986, 40, 1009–1020. [Google Scholar] [CrossRef]
- Zhang, C.X.; Brisson, J.A.; Xu, H.J. Molecular mechanisms of wing polymorphism in insects. Annu. Rev. Entomol. 2019, 64, 297–314. [Google Scholar] [CrossRef]
- Xu, H.J.; Zhang, C.X. Insulin receptors and wing dimorphism in rice planthoppers. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20150489. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.J.; Xue, J.; Lu, B.; Zhang, X.C.; Zhuo, J.C.; He, S.F.; Ma, X.F.; Jiang, Y.Q.; Fan, H.W.; Xu, J.Y.; et al. Two insulin receptors determine alternative wing morphs in planthoppers. Nature 2015, 519, 464–467. [Google Scholar] [CrossRef]
- Lin, X.; Xu, Y.; Jiang, J.; Lavine, M.; Lavine, L.C. Host quality induces phenotypic plasticity in a wing polyphenic insect. Proc. Natl. Acad. Sci. USA 2018, 115, 7563–7568. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Yao, Y.; Wang, B.; Lavine, M.D.; Lavine, L.C. FOXO links wing form polyphenism and wound healing in the brown planthopper, Nilaparvata lugens. Insect Biochem Mol. Biol. 2016, 70, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, X.; Zhao, M.; Guo, M.; Han, K.; Dong, X.; Zhao, J.; Cai, W.; Zhang, Q.; Hua, H. Ultrabithorax is a key regulator for the dimorphism of wings, a main cause for the outbreak of planthoppers in rice. Natl. Sci. Rev. 2020, 7, 1181–1189. [Google Scholar] [CrossRef]
- Iwanaga, K.; Tojo, S. Effects of juvenile-hormone and rearing density on wing dimorphism and oocyte development in the brown planthopper, Nilaparvata lugens. J. Insect Physiol. 1986, 32, 585–590. [Google Scholar] [CrossRef]
- Vellichirammal, N.N.; Madayiputhiya, N.; Brisson, J.A. The genomewide transcriptional response underlying the pea aphid wing polyphenism. Mol. Ecol. 2016, 25, 4146–4160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vellichirammal, N.N.; Gupta, P.; Hall, T.A.; Brisson, J.A. Ecdysone signaling underlies the pea aphid transgenerational wing polyphenism. Proc. Natl. Acad. Sci. USA 2017, 114, 1419–1423. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, X.D. Transcriptomic analysis suggests genes expressed stage-independently and stage-dependently modulating the wing dimorphism of the brown planthopper. Genes 2019, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. Metazoan microRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Kloosterman, W.P.; Plasterk, R.H.A. The diverse functions of microRNAs in animal development and disease. Dev. Cell 2006, 11, 441–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, R.S. MicroRNA function: Multiple mechanisms for a tiny rna? RNA 2005, 11, 1753–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasudevan, S.; Tong, Y.C.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Sun, Y.; Xiao, H.; Ge, C.; Li, F.; Han, Z. Multiple miRNAs jointly regulate the biosynthesis of ecdysteroid in the holometabolous insects, Chilo Suppressalis. RNA 2017, 23, 1817–1833. [Google Scholar] [CrossRef] [Green Version]
- Song, J.S.; Zhou, S.T. Post-transcriptional regulation of insect metamorphosis and oogenesis. Cell Mol. Life Sci. 2020, 77, 1893–1909. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, Y.; Jiang, F.; Song, T.; Wang, H.; Liu, Q.; Zhang, J.; Zhang, J.; Kang, L. miR-71 and miR-263 jointly regulate target genes chitin synthase and chitinase to control locust molting. PLoS Genet. 2016, 12, e1006257. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Wei, Y.; Jiang, F.; Wang, Y.; Guo, X.; He, J.; Kang, L. MicroRNA-133 inhibits behavioral aggregation by controlling dopamine synthesis in locusts. PLoS Genet. 2014, 10, e1004206. [Google Scholar] [CrossRef] [Green Version]
- Shang, F.; Niu, J.Z.B.; Ding, Y.; Zhang, W.; Wei, D.D.; Wei, D.; Jiang, H.B.; Wang, J.J. The miR-9b microRNA mediates dimorphism and development of wing in aphids. Proc. Natl. Acad. Sci. USA 2020, 117, 8404–8409. [Google Scholar] [CrossRef]
- Ye, X.; Xu, L.; Li, X.; He, K.; Hua, H.; Cao, Z.; Xu, J.; Ye, W.; Zhang, J.; Yuan, Z.; et al. miR-34 modulates wing polyphenism in planthopper. PLoS Genet. 2019, 15, e1008235. [Google Scholar] [CrossRef] [Green Version]
- Mackowiak, S.D. Identification of novel and known miRNAs in deep-sequencing data with miRDeep2. Curr. Protoc. Bioinformatics 2011, 36. [Google Scholar] [CrossRef]
- Friedlander, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Assuncao, J.A.; Enright, A.J. MapMi: Automated mapping of microRNA loci. BMC Bioinf. 2010, 11, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Lu, L.; Hua, H.; Zhou, F.; Lu, L.; Lin, Y. Characterization and comparative analysis of small RNAs in three small RNA libraries of the brown planthopper (Nilaparvata lugens). PLoS ONE 2012, 7, e32860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Lv, W.; Hu, L.; Rao, W.; Zeng, Y.; Zhu, L.; He, Y.; He, G. Identification and analysis of brown planthopper-responsive microRNAs in resistant and susceptible rice plants. Sci. Rep. 2017, 7, 8712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human microRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Kruger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- Bennett, F.C.; Harvey, K.F. Fat cadherin modulates organ size in Drosophila via the salvador/warts/hippo signaling pathway. Curr. Biol. 2006, 16, 2101–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neto-Silva, R.M.; Wells, B.S.; Johnston, L.A. Mechanisms of growth and homeostasis in the Drosophila wing. Annu. Rev. Cell Dev. Biol. 2009, 25, 197–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogulja, D.; Rauskolb, C.; Irvine, K.D. Morphogen control of wing growth through the Fat signaling pathway. Dev. Cell 2008, 15, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Li, K.; Gao, Y.; Liu, X.; Chen, W.; Ge, W.; Feng, Q.; Palli, S.R.; Li, S. Antagonistic actions of juvenile hormone and 20-hydroxyecdysone within the ring gland determine developmental transitions in Drosophila. Proc. Natl. Acad. Sci. USA 2018, 115, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Dye, N.A.; Popovic, M.; Spannl, S.; Etournay, R.; Kainmuller, D.; Ghosh, S.; Myers, E.W.; Julicher, F.; Eaton, S. Cell dynamics underlying oriented growth of the Drosophila wing imaginal disc. Development 2017, 144, 4406–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zera, A.J. Evolutionary endocrinology of hormonal rhythms: Juvenile hormone titer circadian polymorphism in Gryllus firmus. Integr. Comp. Biol. 2016, 56, 159–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LEES, A.D. Action of juvenile hormone mimics on the regulation of larval–adult and alary polymorphism in aphids. Nature 1977, 267, 46–48. [Google Scholar] [CrossRef]
- Li, F.C.; Gu, Z.Y.; Wang, B.B.; Xie, Y.; Ma, L.; Xu, K.Z.; Ni, M.; Zhang, H.; Shen, W.D.; Li, B. Effects of the biosynthesis and signaling pathway of ecdysterone on silkworm (Bombyx mori) following exposure to titanium dioxide nanoparticles. J. Chem. Ecol. 2014, 40, 913–922. [Google Scholar] [CrossRef]
- Stanley-Samuelson, D.W.; Jurenka, R.A.; Cripp, C.; Blomquist, G.J.; de Renobal, M. Fatty acids in insects: Composition, metabolrsm, and biological significance. Arch. Insect Biochem. Physiol. 1988, 9, 1–33. [Google Scholar] [CrossRef]
- Gao, X.K.; Luo, J.Y.; Zhu, X.Z.; Wang, L.; Ji, J.C.; Zhang, L.J.; Zhang, S.; Cui, J.J. Growth and fatty acid metabolism of Aphis gossypii parasitized by the parasitic wasp Lysiphlebia japonica. J. Agr. Food Chem. 2019, 67, 8756–8765. [Google Scholar] [CrossRef]
- Richhariya, S.; Jayakumar, S.; Sukumar, S.K.; Hasan, G. dSTIM- and Ral/exocyst-mediated synaptic release from pupal dopaminergic neurons sustains Drosophila flight. Eneuro 2018, 5, e0455-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepperl, J.; Reim, G.; Luthi, U.; Kaech, A.; Hausmann, G.; Basler, K. Sphingolipid depletion impairs endocytic traffic and inhibits wingless signaling. Mech. Develop. 2013, 130, 493–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herr, D.R.; Fyrst, H.; Phan, V.; Heinecke, K.; Georges, R.; Harris, G.L.; Saba, J.D. Sply regulation of sphingolipid signaling molecules is essential for Drosophila development. Development 2003, 130, 2443–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Bao, Z.; Yang, M.; Shen, Y.; Zhang, X.; Yue, B.; Meng, Y.; Fan, Z. Identification and characterization of microRNAs in American cockroach (Periplaneta americana). Gene 2020, 743, 144610. [Google Scholar] [CrossRef] [PubMed]
- Leevers, S.J.; Weinkove, D.; MacDougall, L.K.; Hafen, E.; Waterfield, M.D. The Drosophila phosphoinositide 3-kinase Dp110 promotes cell growth. Embo. J. 1996, 15, 6584–6594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morooka, S.; Tojo, S. Maintenance and selection of strains exhibiting specific wing form and body color under high-density conditions in the brown planthopper, Nilaparvata lugens (Homoptera, Delphacidae). Appl. Entomol. Zool. 1992, 27, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.H.; Xu, L.; Hua, H.X.; Chen, M.Y.; Guo, M.J.; He, K.; Zhao, J.; Li, F. Chromosomal-level genomes of three rice planthoppers provide new insights into sex chromosome evolution. Mol. Ecol. Resour. 2020. [Google Scholar] [CrossRef]
- Yin, C.; Shen, G.; Guo, D.; Wang, S.; Ma, X.; Xiao, H.; Liu, J.; Zhang, Z.; Liu, Y.; Zhang, Y.; et al. InsectBase: A resource for insect genomes and transcriptomes. Nucleic Acids Res. 2016, 44, D801–D807. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Sample | Raw Reads | High Quality | Clean Reads | Q20 | GC Content | 5′adapter Contamines | 3′adapter_Null or Insert_Null | with PloyA/T/G/C |
|---|---|---|---|---|---|---|---|---|---|
| 1 | LW-1st-1 | 13,625,214 | 13,614,741 | 13,330,526 | 98.25% | 48.45% | 3519 | 217,585 | 62,872 |
| 2 | LW-1st-2 | 14,390,780 | 14,380,460 | 14,079,327 | 98.33% | 47.56% | 2826 | 234,820 | 63,230 |
| 3 | LW-2nd-1 | 13,447,515 | 13,437,862 | 13,204,811 | 98.25% | 47.64% | 2869 | 162,612 | 67,300 |
| 4 | LW-2nd-2 | 12,552,925 | 12,544,080 | 12,284,953 | 98.22% | 47.33% | 2611 | 189,043 | 67,244 |
| 5 | LW-3rd-1 | 14,008,106 | 13,997,085 | 13,723,894 | 98.40% | 47.89% | 3182 | 194,273 | 75,452 |
| 6 | LW-3rd-2 | 13,190,404 | 13,178,303 | 12,975,678 | 98.22% | 48.60% | 3609 | 149,545 | 49,216 |
| 7 | LW-4th-1 | 18,344,628 | 18,328,565 | 17,708,977 | 98.13% | 47.87% | 5209 | 540,722 | 73,320 |
| 8 | LW-4th-2 | 18,656,643 | 18,637,940 | 17,960,139 | 98.11% | 48.57% | 4572 | 603,621 | 69,249 |
| 9 | LW-5th-1 | 17,176,623 | 17,165,563 | 16,607,021 | 98.32% | 47.57% | 5806 | 474,728 | 77,696 |
| 10 | LW-5th-2 | 16,751,585 | 16,739,158 | 16,254,637 | 98.12% | 47.58% | 5980 | 419,723 | 58,517 |
| 11 | SW-1st-1 | 17,257,604 | 17,247,057 | 16,612,101 | 98.16% | 47.29% | 8058 | 505,257 | 121,319 |
| 12 | SW-1st-2 | 13,757,659 | 13,748,950 | 13,172,776 | 98.21% | 46.95% | 5890 | 466,836 | 103,188 |
| 13 | SW-2nd-1 | 15,251,104 | 15,240,012 | 14,434,758 | 98.28% | 47.38% | 3383 | 683,287 | 118,280 |
| 14 | SW-2nd-2 | 19,765,649 | 19,750,938 | 19,257,968 | 98.33% | 47.36% | 4397 | 344,244 | 143,995 |
| 15 | SW-3rd-1 | 18,780,777 | 18,766,573 | 17,960,404 | 98.41% | 48.60% | 5290 | 696,703 | 103,821 |
| 16 | SW-3rd-2 | 16,793,873 | 16,779,788 | 16,026,154 | 98.26% | 48.51% | 4287 | 666,676 | 82,349 |
| 17 | SW-4th-1 | 13,125,238 | 13,116,371 | 12,854,273 | 98.26% | 48.00% | 7247 | 142,684 | 111,947 |
| 18 | SW-4th-2 | 13,552,267 | 13,541,969 | 13,173,223 | 98.12% | 47.65% | 4728 | 229,093 | 134,671 |
| 19 | SW-5th-1 | 19,598,809 | 19,582,736 | 18,883,495 | 98.21% | 47.47% | 4252 | 567,450 | 127,157 |
| 20 | SW-5th-2 | 19,297,940 | 19,282,397 | 18,584,543 | 98.29% | 47.92% | 4342 | 573,153 | 120,010 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Zhang, J.; Zhan, A.; Wang, Y.; Ma, X.; Jie, W.; Cao, Z.; Omar, M.A.A.; He, K.; Li, F. Identification and Analysis of MicroRNAs Associated with Wing Polyphenism in the Brown Planthopper, Nilaparvata lugens. Int. J. Mol. Sci. 2020, 21, 9754. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249754
Xu L, Zhang J, Zhan A, Wang Y, Ma X, Jie W, Cao Z, Omar MAA, He K, Li F. Identification and Analysis of MicroRNAs Associated with Wing Polyphenism in the Brown Planthopper, Nilaparvata lugens. International Journal of Molecular Sciences. 2020; 21(24):9754. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249754
Chicago/Turabian StyleXu, Le, Jiao Zhang, Anran Zhan, Yaqin Wang, Xingzhou Ma, Wencai Jie, Zhenghong Cao, Mohamed A. A. Omar, Kang He, and Fei Li. 2020. "Identification and Analysis of MicroRNAs Associated with Wing Polyphenism in the Brown Planthopper, Nilaparvata lugens" International Journal of Molecular Sciences 21, no. 24: 9754. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249754