TGFβ1 Suppressed Matrix Mineralization of Osteoblasts Differentiation by Regulating SMURF1–C/EBPβ–DKK1 Axis

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. TGFβ1 Suppressed Extracellular Matrix Mineralization of Osteoblast Differentiation, but Not Matrix Maturation
2.2. TGFβ1 Downregulated C/EBPβ and DKK1 Expression during Osteoblast Differentiation
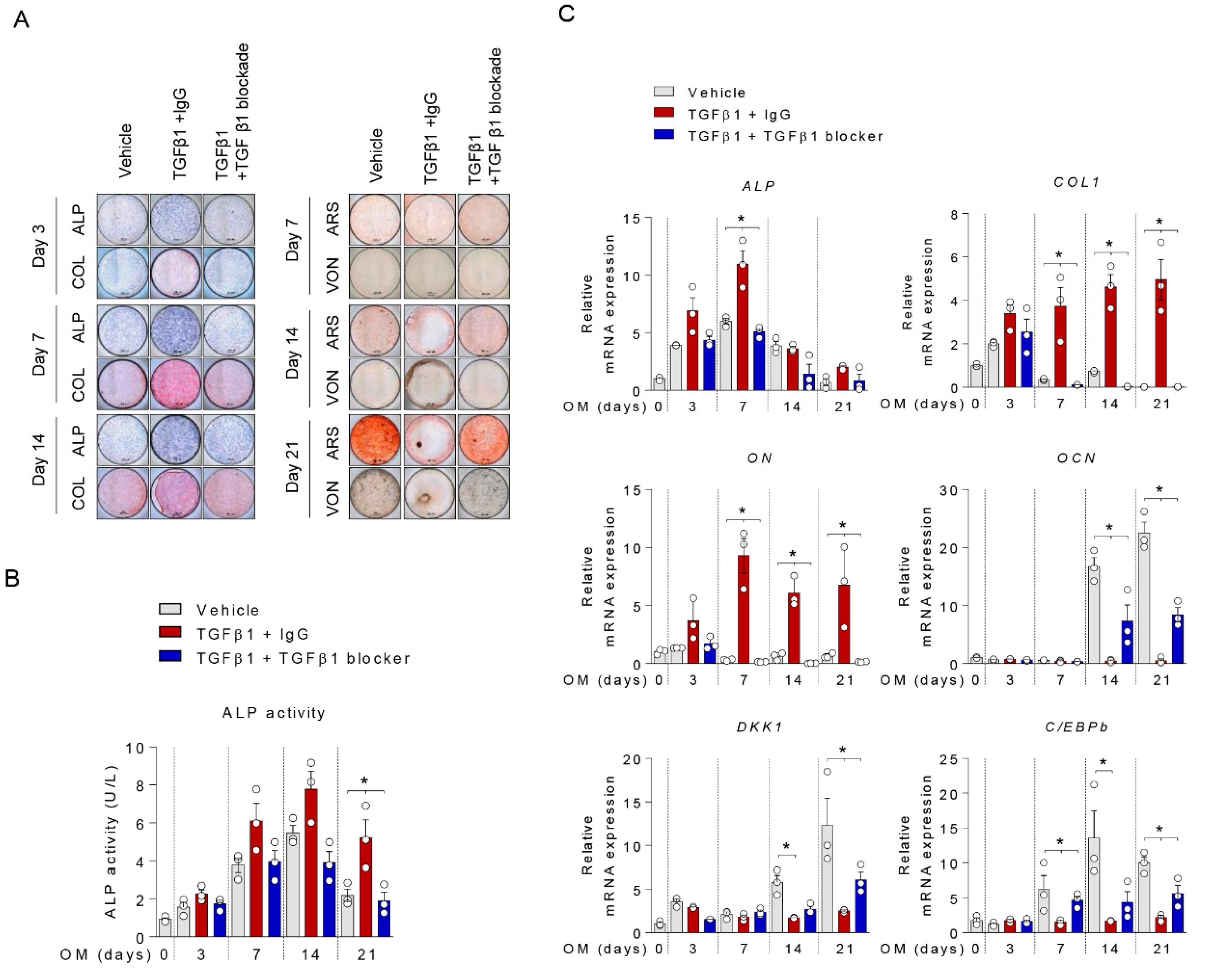
2.3. Anti-TGFβ1 Antibody Reversed TGFβ1-Mediated Suppression of Mineralization
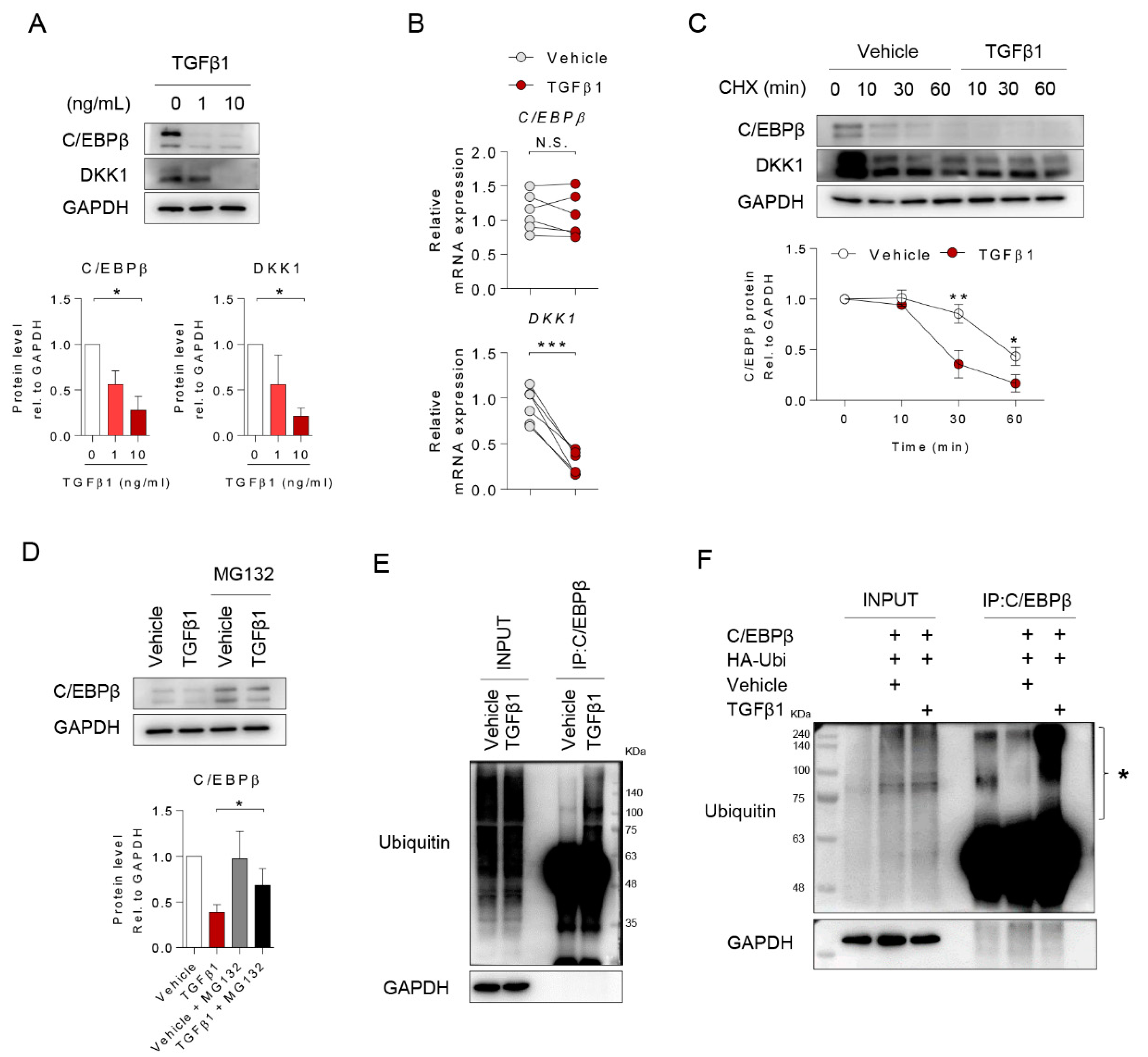
2.4. TGFβ1 Inhibited DKK1 Expression by Inducing Ubiquitination of C/EBPβ Protein
2.5. TGFβ1 Induced SMURF1 to Regulate C/EBPβ-DKK1 Expressions
3. Discussion
4. Materials and Methods
4.1. Human Bone Sample Collection
4.2. Isolation of Human Primary Osteoprogenitors and Differentiation of Osteoblasts
4.3. Assessment of Osteoblast Differentiation
4.4. Total Collagen Assay
4.5. Luciferase Assay
4.6. Constructs, Transfection, and Reagents
4.7. Immunoblot and RT-qPCR
4.8. Immunofluorescence (IF)
4.9. Immunoprecipitation (IP)
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TGFβ1 | Transforming growth factor β1 |
| DKK1 | Dickkopf 1 |
| C/EBPβ | CCAAT/enhancer-binding protein β |
| SMURF1 | Smad ubiquitin regulatory factor 1 |
| SMURF2 | Smad ubiquitin regulatory factor 2 |
| ALP | Alkaline phosphates |
| BSP | Bone sialoprotein |
| COL1 | Type 1 collagen |
| ON | Osteonectin |
| OCN | Osteocalcin |
| RANKL | Receptor activator of nuclear factor-κB ligand |
| RUNX2 | Runt-related transcription factor 2 |
| OA | Osteoarthritis |
| PBS | Phosphate buffered saline |
| DMEM | Dulbecco’s modified eagle medium |
| FBS | Fetal bovine serum |
| AA | Ascorbic acid |
| ARS | Alizarin red S |
| HA | Hydroxyapatite |
| BF | Bright Field |
| siRNA | Small interfering RNA |
| Actino D | Actinomycin D |
| CHX | Cycloheximide |
References
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, G.R., Jr.; Sullivan, E.C.; Moran, E.; Zerler, B. Relationship between alkaline phosphatase levels, osteopontin expression, and mineralization in differentiating MC3T3-E1 osteoblasts. J. Cell. Biochem. 1998, 68, 269–280. [Google Scholar] [CrossRef]
- Beck, G.R., Jr.; Zerler, B.; Moran, E. Phosphate is a specific signal for induction of osteopontin gene expression. Proc. Natl. Acad. Sci. USA 2000, 97, 8352–8357. [Google Scholar] [CrossRef] [Green Version]
- Twine, N.A.; Chen, L.; Pang, C.N.; Wilkins, M.R.; Kassem, M. Identification of differentiation-stage specific markers that define the ex vivo osteoblastic phenotype. Bone 2014, 67, 23–32. [Google Scholar] [CrossRef]
- Henriquez, B.; Hepp, M.; Merino, P.; Sepulveda, H.; van Wijnen, A.J.; Lian, J.B.; Stein, G.S.; Stein, J.L.; Montecino, M. C/EBPbeta binds the P1 promoter of the Runx2 gene and up-regulates Runx2 transcription in osteoblastic cells. J. Cell. Physiol. 2011, 226, 3043–3052. [Google Scholar] [CrossRef] [Green Version]
- Iyer, V.V.; Kadakia, T.B.; McCabe, L.R.; Schwartz, R.C. CCAAT/enhancer-binding protein-beta has a role in osteoblast proliferation and differentiation. Exp. Cell Res. 2004, 295, 128–137. [Google Scholar] [CrossRef]
- Tominaga, H.; Maeda, S.; Hayashi, M.; Takeda, S.; Akira, S.; Komiya, S.; Nakamura, T.; Akiyama, H.; Imamura, T. CCAAT/enhancer-binding protein beta promotes osteoblast differentiation by enhancing Runx2 activity with ATF4. Mol. Biol. Cell 2008, 19, 5373–5386. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, V.; Bryant, H.U.; Macdougald, O.A. Regulation of bone mass by Wnt signaling. J. Clin. Investig. 2006, 116, 1202–1209. [Google Scholar] [CrossRef]
- Li, X.; Liu, P.; Liu, W.; Maye, P.; Zhang, J.; Zhang, Y.; Hurley, M.; Guo, C.; Boskey, A.; Sun, L.; et al. Dkk2 has a role in terminal osteoblast differentiation and mineralized matrix formation. Nat. Genet. 2005, 37, 945–952. [Google Scholar] [CrossRef]
- Jo, S.; Yoon, S.; Lee, S.Y.; Kim, S.Y.; Park, H.; Han, J.; Choi, S.H.; Han, J.S.; Yang, J.H.; Kim, T.H. DKK1 Induced by 1,25D3 Is Required for the Mineralization of Osteoblasts. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, M.E.; Roberts, A.B.; Sporn, M.B.; Bolander, M.E. Transforming growth factor-beta and the initiation of chondrogenesis and osteogenesis in the rat femur. J. Cell Biol. 1990, 110, 2195–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, M.; Camilliere, J.J. In vivo stimulation of bone formation by transforming growth factor-beta. Endocrinology 1989, 124, 2991–2994. [Google Scholar] [CrossRef] [PubMed]
- Janssens, K.; ten Dijke, P.; Janssens, S.; Van Hul, W. Transforming growth factor-beta1 to the bone. Endocr. Rev. 2005, 26, 743–774. [Google Scholar] [CrossRef] [Green Version]
- Alliston, T.; Choy, L.; Ducy, P.; Karsenty, G.; Derynck, R. TGF-beta-induced repression of CBFA1 by Smad3 decreases cbfa1 and osteocalcin expression and inhibits osteoblast differentiation. EMBO J. 2001, 20, 2254–2272. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Deng, C.; Li, Y.P. TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [Green Version]
- Datto, M.; Wang, X.F. Ubiquitin-mediated degradation a mechanism for fine-tuning TGF-beta signaling. Cell 2005, 121, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Hayashi, M.; Komiya, S.; Imamura, T.; Miyazono, K. Endogenous TGF-beta signaling suppresses maturation of osteoblastic mesenchymal cells. EMBO J. 2004, 23, 552–563. [Google Scholar] [CrossRef] [Green Version]
- Gordon, K.J.; Blobe, G.C. Role of transforming growth factor-beta superfamily signaling pathways in human disease. Biochim. Biophys. Acta 2008, 1782, 197–228. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Xie, Z.; Ma, Y.; Pan, X.; Wang, J.; Chen, Z.; Shi, P. TGF-beta inhibits osteogenesis by upregulating the expression of ubiquitin ligase SMURF1 via MAPK-ERK signaling. J. Cell. Physiol. 2018, 233, 596–606. [Google Scholar] [CrossRef]
- Iyengar, P. Regulation of Ubiquitin Enzymes in the TGF-β Pathway. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assoian, R.K.; Komoriya, A.; Meyers, C.A.; Miller, D.M.; Sporn, M.B. Transforming growth factor-beta in human platelets. Identification of a major storage site, purification, and characterization. J. Biol. Chem. 1983, 258, 7155–7160. [Google Scholar] [PubMed]
- Letterio, J.J.; Roberts, A.B. Regulation of immune responses by TGF-beta. Annu. Rev. Immunol. 1998, 16, 137–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepper, M.S. Transforming growth factor-beta: Vasculogenesis, angiogenesis, and vessel wall integrity. Cytokine Growth Factor Rev. 1997, 8, 21–43. [Google Scholar] [CrossRef]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-beta and the TGF-beta Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Oh, Y.; Jo, S.; Kim, T.H.; Ji, J.D. A dual role of TGF-beta in human osteoclast differentiation mediated by Smad1 versus Smad3 signaling. Immunol. Lett. 2019, 206, 33–40. [Google Scholar] [CrossRef]
- Matsunobu, T.; Torigoe, K.; Ishikawa, M.; de Vega, S.; Kulkarni, A.B.; Iwamoto, Y.; Yamada, Y. Critical roles of the TGF-beta type I receptor ALK5 in perichondrial formation and function, cartilage integrity, and osteoblast differentiation during growth plate development. Dev. Biol. 2009, 332, 325–338. [Google Scholar] [CrossRef] [Green Version]
- Sarahrudi, K.; Thomas, A.; Mousavi, M.; Kaiser, G.; Kottstorfer, J.; Kecht, M.; Hajdu, S.; Aharinejad, S. Elevated transforming growth factor-beta 1 (TGF-beta1) levels in human fracture healing. Injury 2011, 42, 833–837. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, X.; Zhao, D.; Liu, B.; Wang, B.; Yu, W.; Li, J.; Yu, X.; Cao, F.; Zheng, G.; et al. TGFbeta1 promotes the osteoinduction of human osteoblasts via the PI3K/AKT/mTOR/S6K1 signalling pathway. Mol. Med. Rep. 2019, 19, 3505–3518. [Google Scholar] [CrossRef]
- Alves, R.D.; Eijken, M.; Bezstarosti, K.; Demmers, J.A.; van Leeuwen, J.P. Activin A suppresses osteoblast mineralization capacity by altering extracellular matrix (ECM) composition and impairing matrix vesicle (MV) production. Mol. Cell. Proteom. 2013, 12, 2890–2900. [Google Scholar] [CrossRef] [Green Version]
- Baroncelli, M.; Drabek, K.; Eijken, M.; van der Eerden, B.C.J.; van de Peppel, J.; van Leeuwen, J. Two-day-treatment of Activin-A leads to transient change in SV-HFO osteoblast gene expression and reduction in matrix mineralization. J. Cell. Physiol. 2020, 235, 4865–4877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducy, P.; Desbois, C.; Boyce, B.; Pinero, G.; Story, B.; Dunstan, C.; Smith, E.; Bonadio, J.; Goldstein, S.; Gundberg, C.; et al. Increased bone formation in osteocalcin-deficient mice. Nature 1996, 382, 448–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diegel, C.R.; Hann, S.; Ayturk, U.M.; Hu, J.C.W.; Lim, K.E.; Droscha, C.J.; Madaj, Z.B.; Foxa, G.E.; Izaguirre, I.; Transgenics Core, V.V.A.; et al. An osteocalcin-deficient mouse strain without endocrine abnormalities. PLoS Genet. 2020, 16, e1008361. [Google Scholar] [CrossRef] [PubMed]
- Moriishi, T.; Ozasa, R.; Ishimoto, T.; Nakano, T.; Hasegawa, T.; Miyazaki, T.; Liu, W.; Fukuyama, R.; Wang, Y.; Komori, H.; et al. Osteocalcin is necessary for the alignment of apatite crystallites, but not glucose metabolism, testosterone synthesis, or muscle mass. PLoS Genet. 2020, 16, e1008586. [Google Scholar] [CrossRef]
- Di, M.; Wang, L.; Li, M.; Zhang, Y.; Liu, X.; Zeng, R.; Wang, H.; Chen, Y.; Chen, W.; Zhang, Y.; et al. Dickkopf1 destabilizes atherosclerotic plaques and promotes plaque formation by inducing apoptosis of endothelial cells through activation of ER stress. Cell Death Dis. 2017, 8, e2917. [Google Scholar] [CrossRef]
- Liang, L.; Tu, Y.; Lu, J.; Wang, P.; Guo, Z.; Wang, Q.; Guo, K.; Lan, R.; Li, H.; Liu, P. Dkk1 exacerbates doxorubicin-induced cardiotoxicity by inhibiting the Wnt/beta-catenin signaling pathway. J. Cell Sci. 2019, 132. [Google Scholar] [CrossRef] [Green Version]
- Mikheev, A.M.; Mikheeva, S.A.; Rostomily, R.; Zarbl, H. Dickkopf-1 activates cell death in MDA-MB435 melanoma cells. Biochem. Biophys. Res. Commun. 2007, 352, 675–680. [Google Scholar] [CrossRef] [Green Version]
- Dallas, S.L.; Bonewald, L.F. Dynamics of the transition from osteoblast to osteocyte. Ann. N. Y. Acad. Sci. 2010, 1192, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.S.; Gu, Y.; Jiang, C.; Chen, L. Osteonectin regulates the extracellular matrix mineralization of osteoblasts through P38 signaling pathway. J. Cell. Physiol. 2020, 235, 2220–2231. [Google Scholar] [CrossRef]
- Malipiero, U.; Holler, M.; Werner, U.; Fontana, A. Sequence analysis of the promoter region of the glioblastoma derived T cell suppressor factor/transforming growth factor (TGF)-beta 2 gene reveals striking differences to the TGF-beta 1 and -beta 3 genes. Biochem. Biophys. Res. Commun. 1990, 171, 1145–1151. [Google Scholar] [CrossRef]
- Seyedin, S.M.; Thomas, T.C.; Thompson, A.Y.; Rosen, D.M.; Piez, K.A. Purification and characterization of two cartilage-inducing factors from bovine demineralized bone. Proc. Natl. Acad. Sci. USA 1985, 82, 2267–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, R.; Asch, E.; Bloch, D.; Bole, G.; Borenstein, D.; Brandt, K.; Christy, W.; Cooke, T.D.; Greenwald, R.; Hochberg, M.; et al. Development of criteria for the classification and reporting of osteoarthritis. Classification of osteoarthritis of the knee. Diagnostic and Therapeutic Criteria Committee of the American Rheumatism Association. Arthritis Rheum. 1986, 29, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.A.; Gundle, R.; Beresford, J.N. Isolation and culture of bone-forming cells (osteoblasts) from human bone. Methods Mol. Med. 1996, 2, 233–262. [Google Scholar] [CrossRef] [PubMed]
- Wrobel, E.; Leszczynska, J.; Brzoska, E. The Characteristics Of Human Bone-Derived Cells (HBDCS) during osteogenesis in vitro. Cell Mol. Biol. Lett. 2016, 21, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Ecarot, B.; Glorieux, F.H. In vivo osteogenic activity of isolated human bone cells. J. Bone Miner. Res. 1991, 6, 45–51. [Google Scholar] [CrossRef]
- Jo, S.; Kang, S.; Han, J.; Choi, S.H.; Park, Y.S.; Sung, I.H.; Kim, T.H. Accelerated osteogenic differentiation of human bone-derived cells in ankylosing spondylitis. J. Bone Miner. Metab. 2018, 36, 307–313. [Google Scholar] [CrossRef]
- Jo, S.; Lee, J.K.; Han, J.; Lee, B.; Kang, S.; Hwang, K.T.; Park, Y.S.; Kim, T.H. Identification and characterization of human bone-derived cells. Biochem. Biophys. Res. Commun. 2018, 495, 1257–1263. [Google Scholar] [CrossRef]
- Park, P.-R.; Jo, S.; Jin, S.-H.; Kim, T.-J. MicroRNA-10b Plays a Role in Bone Formation by Suppressing Interleukin-22 in Ankylosing Spondylitis. J. Rheum. Dis. 2020, 27. [Google Scholar] [CrossRef] [Green Version]
- Ha, Y.J.; Choi, Y.S.; Kang, E.H.; Shin, K.; Kim, T.K.; Song, Y.W.; Lee, Y.J. SOCS1 suppresses IL-1beta-induced C/EBPbeta expression via transcriptional regulation in human chondrocytes. Exp. Mol. Med. 2016, 48, e241. [Google Scholar] [CrossRef] [Green Version]
- Jo, S.; Lee, Y.Y.; Han, J.; Lee, Y.L.; Yoon, S.; Lee, J.; Oh, Y.; Han, J.S.; Sung, I.H.; Park, Y.S.; et al. CCAAT/enhancer-binding protein beta (C/EBPbeta) is an important mediator of 1,25 dihydroxyvitamin D3 (1,25D3)-induced receptor activator of nuclear factor kappa-B ligand (RANKL) expression in osteoblasts. BMB Rep. 2019, 52, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; Kim, Y.J.; Jeong, H.M.; Jin, Y.H.; Yeo, C.Y.; Lee, K.Y. Akt enhances Runx2 protein stability by regulating Smurf2 function during osteoblast differentiation. FEBS J. 2014, 281, 3656–3666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.; Lee, Y.L.; Kim, S.; Lee, H.; Chung, H. PCGF2 negatively regulates arsenic trioxide-induced PML-RARA protein degradation via UBE2I inhibition in NB4 cells. Biochim. Biophys. Acta 2016, 1863, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.G.; Jeong, B.C.; Kim, E.J.; Choi, H.; Oh, S.H.; Kim, D.K.; Koo, S.H.; Choi, H.S.; Koh, J.T. Cyclic AMP Response Element-binding Protein H (CREBH) Mediates the Inhibitory Actions of Tumor Necrosis Factor alpha in Osteoblast Differentiation by Stimulating Smad1 Degradation. J. Biol. Chem. 2015, 290, 13556–13566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.; Wang, S.E.; Lee, Y.L.; Kang, S.; Lee, B.; Han, J.; Sung, I.H.; Park, Y.S.; Bae, S.C.; Kim, T.H. IL-17A induces osteoblast differentiation by activating JAK2/STAT3 in ankylosing spondylitis. Arthritis Res. Ther. 2018, 20, 115. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, B.; Park, H.; Lee, Y.L.; Oh, Y.; Park, J.; Kim, S.Y.; Weon, S.; Choi, S.H.; Yang, J.-H.; Jo, S.; et al. TGFβ1 Suppressed Matrix Mineralization of Osteoblasts Differentiation by Regulating SMURF1–C/EBPβ–DKK1 Axis. Int. J. Mol. Sci. 2020, 21, 9771. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249771
Nam B, Park H, Lee YL, Oh Y, Park J, Kim SY, Weon S, Choi SH, Yang J-H, Jo S, et al. TGFβ1 Suppressed Matrix Mineralization of Osteoblasts Differentiation by Regulating SMURF1–C/EBPβ–DKK1 Axis. International Journal of Molecular Sciences. 2020; 21(24):9771. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249771
Chicago/Turabian StyleNam, Bora, Hyosun Park, Young Lim Lee, Younseo Oh, Jinsung Park, So Yeon Kim, Subin Weon, Sung Hoon Choi, Jae-Hyuk Yang, Sungsin Jo, and et al. 2020. "TGFβ1 Suppressed Matrix Mineralization of Osteoblasts Differentiation by Regulating SMURF1–C/EBPβ–DKK1 Axis" International Journal of Molecular Sciences 21, no. 24: 9771. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249771