Genome-Wide Characterization and Expression Profiling of GASA Genes during Different Stages of Seed Development in Grapevine (Vitis vinifera L.) Predict Their Involvement in Seed Development

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Protein Features of GASA Genes in Grapevine
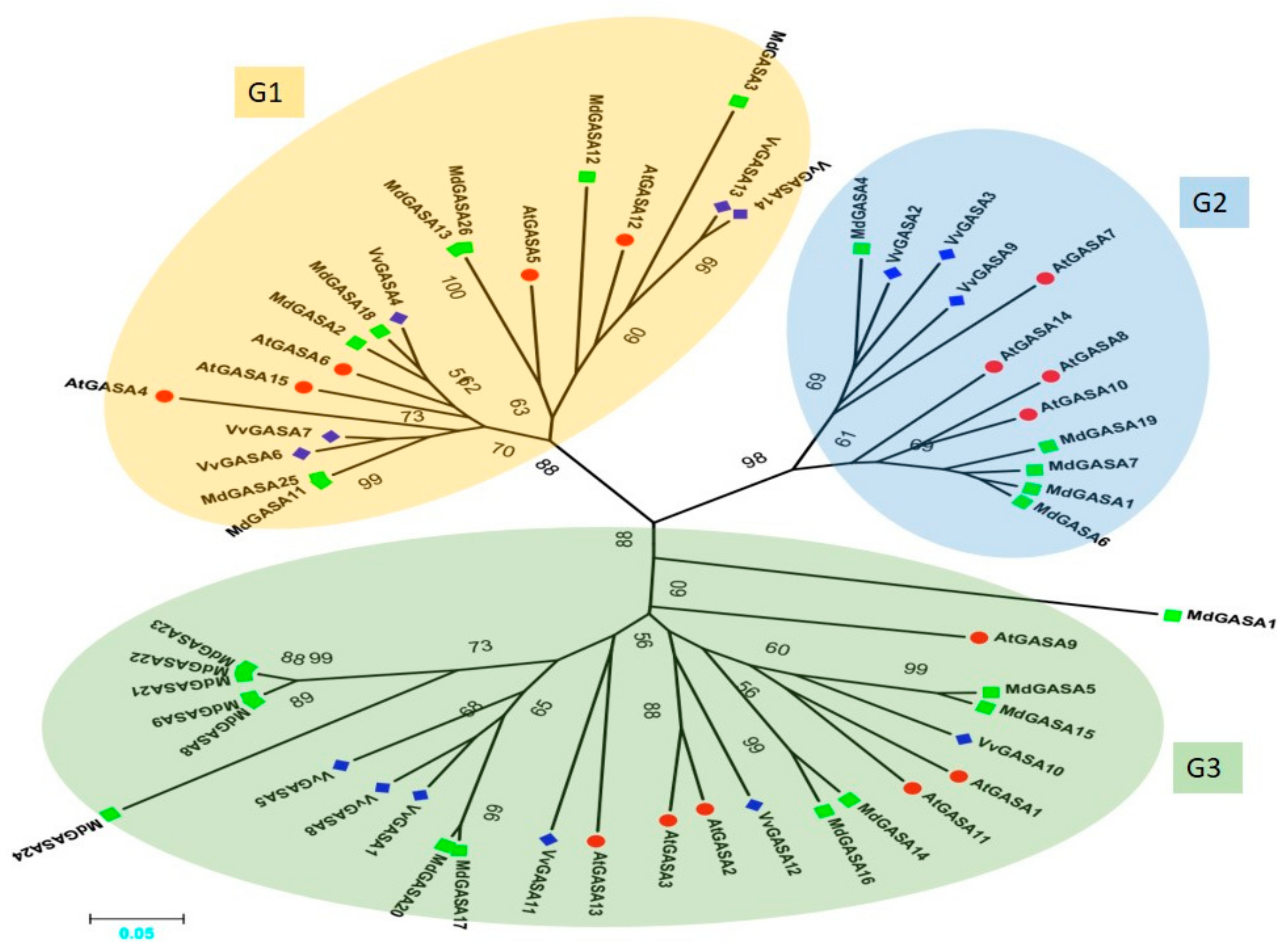
2.2. Phylogenetic Analysis of GASA Genes from Grape, Apple and Arabidopsis
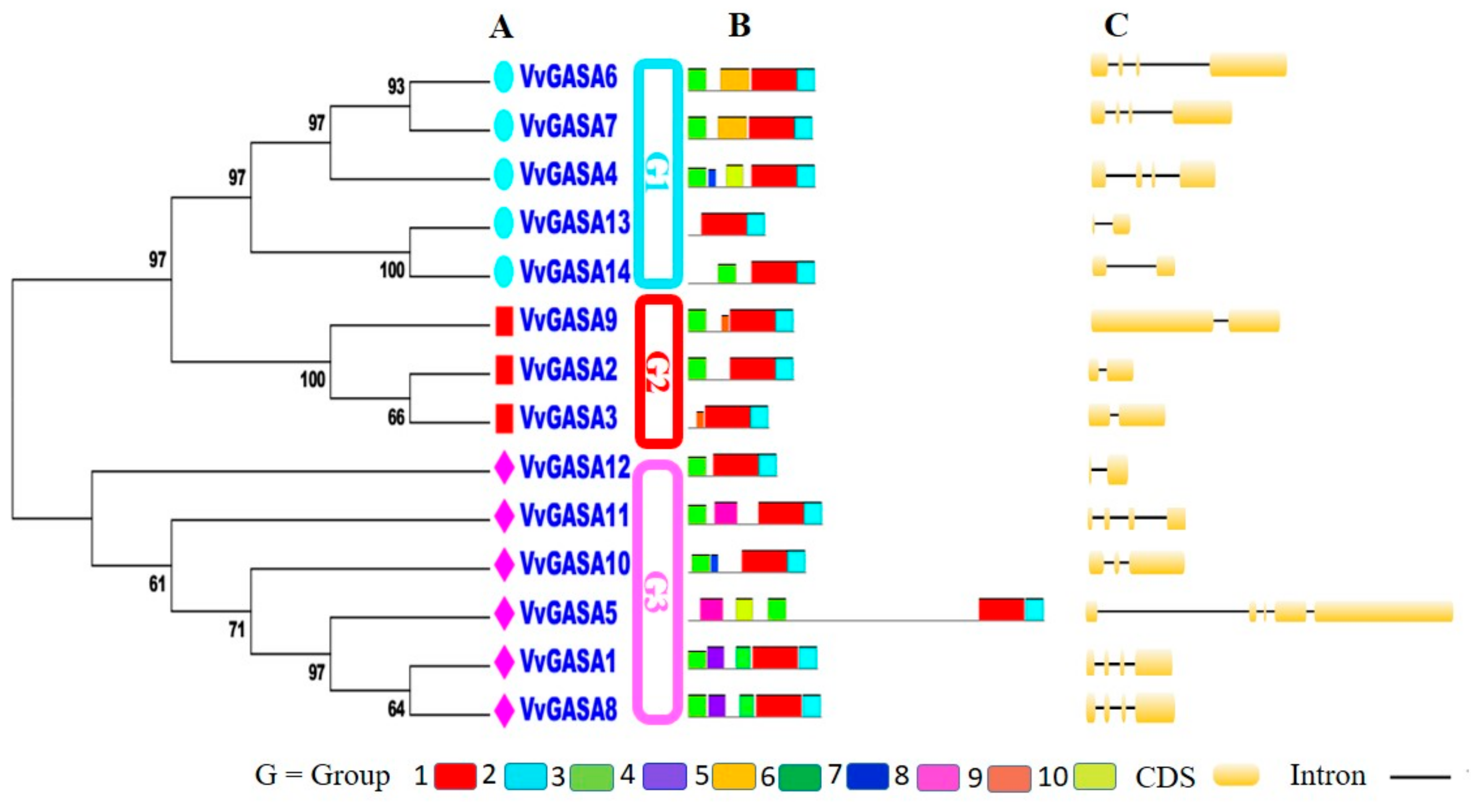
2.3. Analysis of Conserved Motifs, Domain Architecture, and Gene Structure
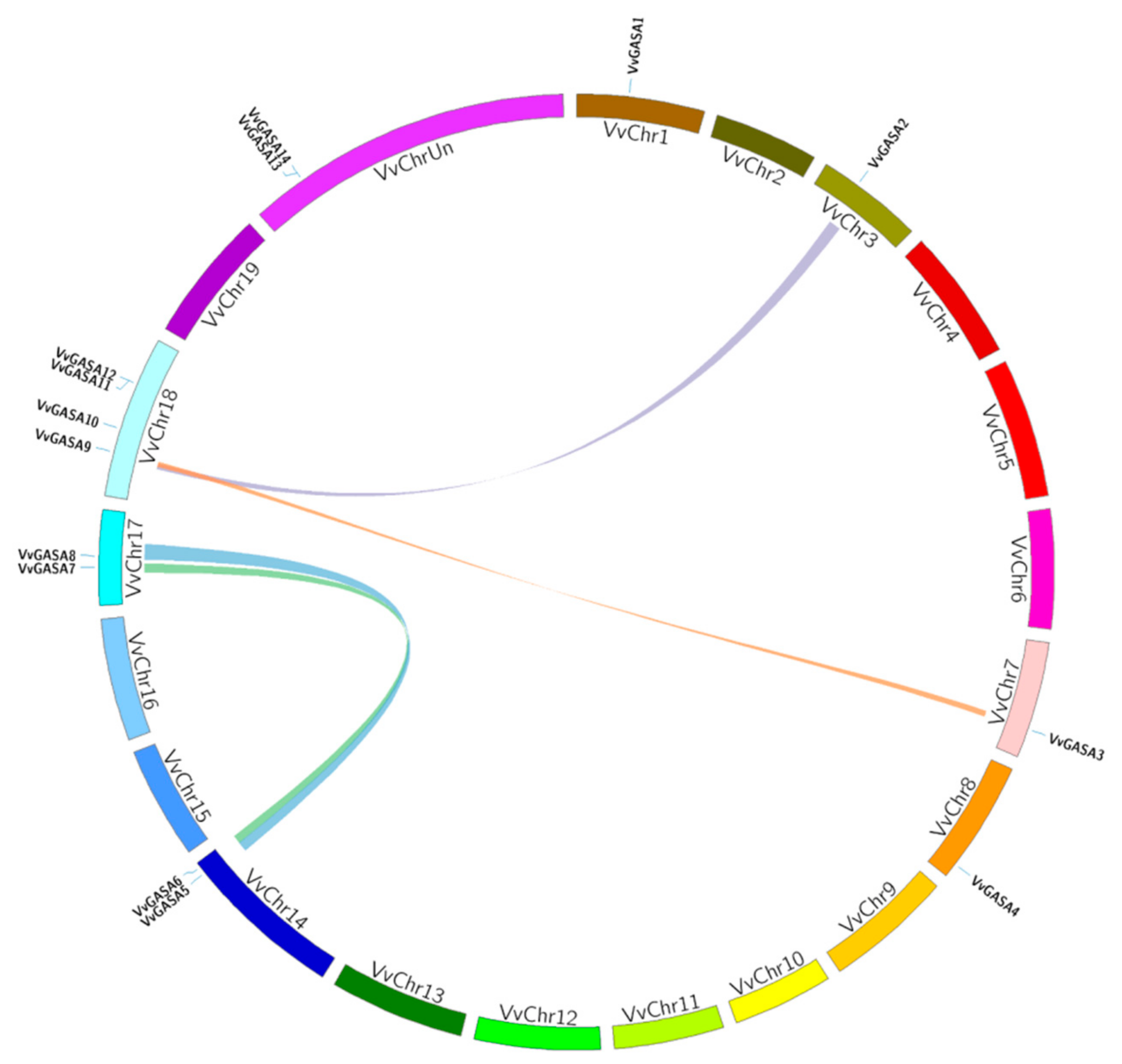
2.4. Grapevine Genes Duplication and Evolutionary Analysis
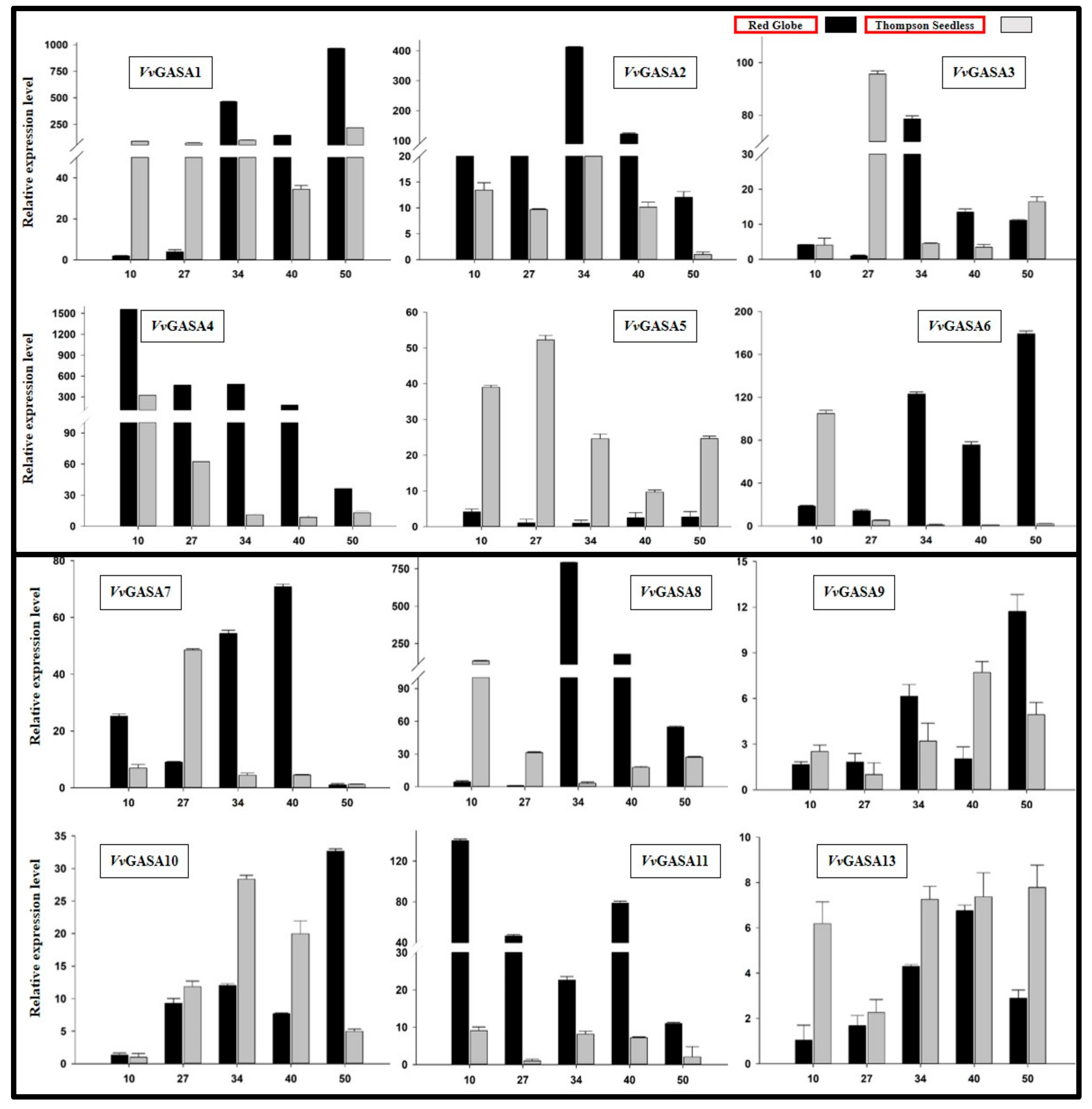
2.5. GASA Genes Expression Profiling During Seed Development
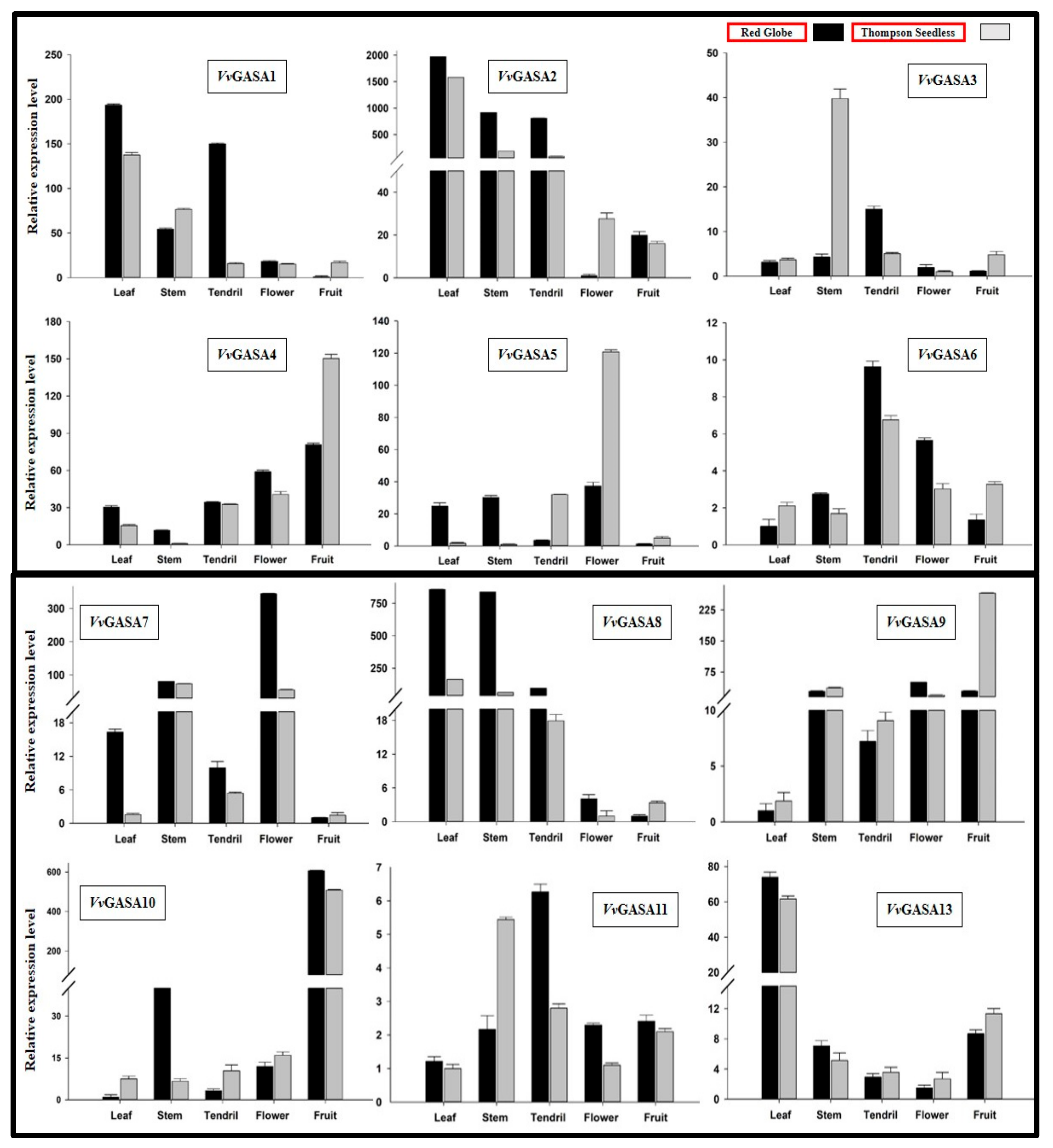
2.6. Tissue Specific Expression Profiling of Grapevine GASA Candidates
2.7. Promoter Analysis of GASA Genes
3. Discussion
4. Materials and Methods
4.1. Annotation and Identification of Putative Grapevine GASA Genes
4.2. Physicochemical Properties and Phylogeny Analysis
4.3. Exon–Intron, Gene Structures, Conserved Motif, and Promoter Analysis
4.4. Synteny Analysis and Calculation of Ka/Ks Ratio for Duplicated Genes
4.5. Plant Materials
4.6. Total RNA Extraction and Expression Analysis by RT-PCR
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Oliveira-Lima, M.; Benko-Iseppon, A.M.; Neto, J.R.; Rodriguez-Decuadro, S.; Kido, E.A.; Crovella, S.; Pandolfi, V. Snakin: Structure, roles and applications of a plant antimicrobial peptide. Curr. Protein Pept. Sci. 2017, 18, 368–374. [Google Scholar] [CrossRef]
- Shi, L.; Olszewski, N.E. Gibberellin and abscisic acid regulate GAST1 expression at the level of transcription. Plant Mol. Biol. 1998, 38, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Aubert, D.; Chevillard, M.; Dorne, A.M.; Arlaud, G.; Herzog, M. Expression patterns of GASA genes in Arabidopsis thaliana: the GASA4 gene is up-regulated by gibberellins in meristematic regions. Plant Mol. Biol. 1998, 36, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Trapalis, M.; Li, S.F.; Parish, R.W. The Arabidopsis GASA10 gene encodes a cell wall protein strongly expressed in developing anthers and seeds. Plant Sci. 2017, 260, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Sakaguchi, N.; Shimada, H. Two OsGASR genes, rice GAST homologue genes that are abundant in proliferating tissues, show different expression patterns in developing panicles. Genes Genet. Syst. 2006, 81, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.L.; Bai, X.; Li, Y.; Cai, H.; Ji, W.; Tang, L.L.; Wen, Y.D.; Zhu, Y.M. GsGASA1 mediated root growth inhibition in response to chronic cold stress is marked by the accumulation of DELLAs. J. Plant Physiol. 2011, 168, 2153–2560. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.; Xu, Y.Y.; Joo, S.H.; Kim, S.K.; Xue, Z.; Xu, Z.H.; Wang, Z.Y.; Chong, K. OsGSR1 is involved in crosstalk between gibberellins and brassinosteroids in rice. Plant J. 2009, 57, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, K.A.; Moskal, W.A.; Wu, H.C.; Underwood, B.A.; Graham, M.A.; Town, C.D.; Vanden Bosch, K.A. Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants. Plant J. 2007, 51, 262–280. [Google Scholar] [CrossRef]
- Zhang, S.C.; Wang, X.J. Expression pattern of GASA, downstream genes of DELLA, in Arabidopsis. Chin. Sci. Bull. 2008, 53, 3839–3846. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yang, C.; Peng, J.; Sun, S.; Wang, X. GASA5, a regulator of flowering time and stem growth in Arabidopsis Thaliana. Plant Mol. Biol. 2009, 69, 745–759. [Google Scholar] [CrossRef]
- Roxrud, I.; Lid, S.E.; Fletcher, J.C.; Schmidt, E.D.; Opsahl-Sorteberg, H.G. GASA4, one of the 14-member Arabidopsis GASA family of small polypeptides, regulates flowering and seed development. Plant cell physiol. 2007, 48, 471–483. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Wang, Q.; Zhang, X.; Zhang, B.; Luo, H.; He, C. Comprehensive transcriptional and functional analyses of HbGASA genes reveal their roles in fungal pathogen resistance in Hevea brasiliensis. Tree Genet. Genom. 2018, 14, 41. [Google Scholar] [CrossRef]
- Nahirñak, V.; Rivarola, M.; Gonzalez de Urreta, M.; Paniego, N.; Hopp, H.E.; Almasia, N.I.; Vazquez-Rovere, C. Genome-wide Analysis of the Snakin/GASA Gene Family in Solanum tuberosum cv. Kennebec. Am. J. Potato Res. 2016, 93, 172–188. [Google Scholar] [CrossRef]
- Herzog, M.; Dorne, A.M.; Grellet, F. GASA, a gibberellin-regulated gene family from Arabidopsis thaliana related to the tomato GAST1 gene. Plant Mol. Biol. 1995, 27, 743–752. [Google Scholar] [CrossRef]
- Ben-Nissan, G.; Lee, J.Y.; Borohov, A.; Weiss, D. GIP, a Petunia Hybrida GA induced cysteine-rich protein: A possible role in shoot elongation and transition to flowering. Plant J. 2004, 37, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Geng, X.L.; Zhang, H.Y.; Zhou, C.L.; Zhao, A.J.; Wang, F.; Zhao, Y.; Tian, X.J.; Hu, Z.R.; Xin, M.M.; et al. Isolation and characterization of heat-responsive gene TaGASR1 from wheat (Triticum aestivum L.). J. Plant Biol. 2017, 60, 57–65. [Google Scholar] [CrossRef]
- Fan, S.; Zhang, D.; Xing, L.; Qi, S.; Du, L.; Wu, H.; Shao, H.; Li, Y.; Ma, J.; Han, M. Comprehensive analysis of GASA family members in the Malus domestica genome: Identifcation, characterization, and their expressions in response to apple flower induction. BMC Genom. 2017, 18, 827. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Sana, A.; Jamil, A.; Nasir, J.A.; Ahmed, S.; Hameed, M.U.; Abdullah. A genome-wide approach to the comprehensive analysis of GASA gene family in Glycine max. Plant Mol. Biol. 2019, 100, 607. [Google Scholar] [CrossRef]
- Ling, H.Q.; Zhao, S.; Liu, D.; Wang, J.; Sun, H.; Zhang, C.; Fan, H.; Li, D.; Dong, L.; Tao, Y. Draft genome of the wheat A-genome progenitor Triticumurartu. Nature 2013, 496, 487–490. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Wang, F.; Liu, T.; Dong, Z.; Li, A.; Jing, R.; Mao, L.; Li, Y.; Liu, X.; Zhang, K.; et al. Natural variation of TaGASR7-1 A1affects grain length in common wheat under multiple cultivation conditions. Mol. Breed. 2014, 34, 937. [Google Scholar] [CrossRef]
- Muhammad, I.; Li, W.Q.; Jinga, X.Q.; Zhoua, M.R.; Shalmania, A.; Ali, M.; Weia, X.Y.; Sharif, R.; Liua, W.T.; Chen, K.M. A systematic in silico prediction of Gibberellic acid stimulated GASA family members: a novel small peptide contributes to floral architecture and transcriptomic changes induced by external stimuli in rice. J. Plant Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- FAO STAT 2018. Available online: http://www.fao.org/land-water/databases-and-software/crop-information/grape/en/ (accessed on 30 December 2019).
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends. Genet. 2006, 22, 511–519. [Google Scholar] [CrossRef] [PubMed]
- FAO; OIV. FAO-OIV Focus Table and Dried Grapes; Non-Alcoholic Products of the Vitivini Cultural Sector Intended for Human Consumption; FAO: Rome, Italy, 2016; Volume I7042, p. 64. Available online: www.oiv.int/public/medias/5268/fao-oiv-focus-2016 (accessed on 30 December 2019).
- Zhang, S.; Wang, L.; Sun, X.; Li, Y.; Yao, J.; Nocker, S.v.; Wang, X. Genome-Wide Analysis of the YABBY Gene Family in Grapevine and Functional Characterization of VvYABBY4. Front. Plant Sci. 2019, 10, 1207. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.D.; Zhang, S.L.; Dong, R.Z.; Wang, L.; Yao, J.; Nocker, S.V.; Wang, X. The grapevine homeobox gene VvHB58 infuences seed and fruit development through multiple hormonal signaling pathways. BMC Plant Biol. 2019, 19, 523. [Google Scholar] [CrossRef] [Green Version]
- Li, W.H.; Gojobori, T.; Nei, M. Pseudogenes as a paradigm of neutral evolution. Nature 1981, 292, 237–239. [Google Scholar] [CrossRef]
- Bassett, D.E.; Eisen, M.B.; Boguski, M.S. Gene expression informatics–it’s all in your mine. Nat. Genet. 1999, 21, 51–55. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, X. One new kind of phytohormonal signaling integrator: Up-and-coming GASA family genes. Plant Signal. Behav. 2017, 12, e1226453-2. [Google Scholar] [CrossRef] [Green Version]
- Rubinovich, L.; Weiss, D. The Arabidopsis cysteine-rich protein GASA4 promotes GA responses and exhibits redox activity in bacteria and in planta. Plant J. 2010, 64, 1018–1027. [Google Scholar] [CrossRef]
- Qu, J.; Kang, S.G.; Hah, C.; Jang, J.C. Molecular and cellular characterization of GA-Stimulated Transcripts GASA4 and GASA6 in Arabidopsis thaliana. Plant Sci. 2016, 246, 1–10. [Google Scholar] [CrossRef]
- Yang, Q.; Niu, Q.; Tang, Y.; Ma, Y.; Yan, X.; Li, J.; Tian, J.; Bai, S.; Teng, Y. PpyGAST1 is potentially involved in bud dormancy release by integrating the GA biosynthesis and ABA signaling in ‘Suli’ pear (Pyruspyrifolia White Pear Group). Environ. Exp. Bot. 2019, 162, 302–312. [Google Scholar] [CrossRef]
- Moyano-Cañete, E.; Bellido, M.L.; García-Caparrós, N.; Medina-Puche, L.; AmilRuiz, F.; González-Reyes, J.A.; Caballero, J.L.; Mun˜oz-Blanco, J.; Blanco-Portales, R. FaGAST2, a strawberry ripening related gene, acts together with faGAST1 to determine cell size of the fruit receptacle. Plant Cell Physiol. 2013, 54, 218–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, A.H.; Freeling, M.; Tang, H.; Wang, X. Insights from the comparison of plant genome sequences. Annu Rev. Plant Biol. 2010, 61, 349–372. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, C.C.; Loughran, N.B.; Walsh, T.A.; Harrison, A.J.; O’Connell, M.J. Positive selectionneighboring functionally essential sites and disease-implicated regions of mammalian reproductiveproteins. BMC Evol. Biol. 2010, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhao, Y.; Wei, X.; Li, C.; Wang, A.; Zhao, Q.; Li, W.; Guo, Y.; Deng, L.; Zhu, C. Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm. Nat. Genet. 2012, 44, 32–39. [Google Scholar] [CrossRef]
- Li, X.; Shi, S.; Tao, Q.; Tao, Y.; Miao, J.; Peng, X.; Li, C.; Yang, Z.; Zhou, Y.; Liang, G. OsGASR9 positively regulates grain size and yield in rice (Oryza sativa). Plant Sci. 2019, 286, 17–27. [Google Scholar] [CrossRef]
- Altschul, S.F.; Warren, G.; Webb, M.; Eugene, W.M.; David, J.L. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar]
- Ahmad, B.; Zhang, S.; Yao, J.; Rahman, M.U.; Hanif, M.; Zhu, Y.; Wang, X. Genomic Organization of the B3-Domain Transcription Factor Family in Grapevine (Vitis vinifera L.) and Expression during Seed Development in Seedless and Seeded Cultivars. Int. J. Mol. Sci. 2019, 20, 4553. [Google Scholar] [CrossRef] [Green Version]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; De Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Tang, H.; Wang, X.; Paterson, A.H. PGDD: A database of gene and genome duplication in plants. Nucleic Acids Res. 2012, 41, D1152–D1158. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Juretic, N.; Hoen, D.R.; Huynh, M.L.; Harrison, P.M.; Bureau, T.E. The evolutionary fate of MULE-mediated duplications of host gene fragments in rice. Genome Res. 2005, 15, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Locus ID | Gene ID | Accession No. | Chromosome No. | Start Site | End Site | CDS (bp) | ORF (aa) |
|---|---|---|---|---|---|---|---|
| GSVIVT01020178001 | VvGASA1 | CBI32100 | 1+ | 9381743 | 9382568 | 327 | 108 |
| GSVIVT01037887001 | VvGASA2 | CBI22497 | 3- | 6715491 | 6716068 | 267 | 88 |
| GSVIVT01000168001 | VvGASA3 | CBI33733 | 7+ | 15821751 | 15822734 | 204 | 67 |
| GSVIVT01033563001 | VvGASA4 | CBI30071 | 8- | 19734994 | 19736181 | 321 | 106 |
| GSVIVT01032528001 | VvGASA5 | CBI34969 | 14+ | 28107504 | 28112390 | 897 | 298 |
| GSVIVT01011412001 | VvGASA6 | CBI22214 | 14- | 29446051 | 29447932 | 321 | 106 |
| GSVIVT01008003001 | VvGASA7 | CBI15224 | 17+ | 6769396 | 6770752 | 315 | 104 |
| GSVIVT01007817001 | VvGASA8 | CBI15083 | 17+ | 8741561 | 8742409 | 336 | 111 |
| GSVIVT01009384001 | VvGASA9 | CBI19434 | 18+ | 7913344 | 7915123 | 267 | 88 |
| GSVIVT01009902001 | VvGASA10 | CBI19861 | 18- | 12275798 | 12276616 | 297 | 98 |
| GSVIVT01034477001 | VvGASA11 | CBI18167 | 18+ | 20718815 | 20720279 | 339 | 112 |
| GSVIVT01034476001 | VvGASA12 | CBI18166 | 18- | 20720304 | 20720676 | 225 | 74 |
| GSVIVT01003387001 | VvGASA13 | CBI25689 | Un- | 9775242 | 9775609 | 195 | 64 |
| GSVIVT01003388001 | VvGASA14 | CBI25690 | Un- | 9791751 | 9792551 | 321 | 106 |
| Gene | MW | PI | Major Amino Acid% | Instability Index | Aliphatic Index | GRAVY | Localization Predicted |
|---|---|---|---|---|---|---|---|
| GASA1 | 11.96 | 8.61 | C(11.9), L(8.3), R(7.3) | 36.64 | 67.06 | −0.172 | extr., vacu. |
| GASA2 | 9.71 | 9.02 | C(14.8), K(12.5), L(11.4) | 38.49 | 58.75 | −0.281 | chlo, nucl., extr |
| GASA3 | 7.28 | 8.87 | C(17.9), K(13.4), G(10.4) | 41.98 | 33.43 | −0.515 | chlo., nucl., cyto., extr. |
| GASA4 | 11.85 | 9.20 | C(11.2), P(11.2), T(9.3) | 51.40 | 45.61 | −0.421 | extr., chlo., nucl. |
| GASA5 | 31.96 | 9.64 | P(23.7), V(9.7), K(8.4) | 67.94 | 78.53 | −0.241 | cyto., ER |
| GASA6 | 11.79 | 9.30 | C(11.3), K(11.3), G(9.4) | 35.75 | 53.40 | −0.289 | extr., vacu., chlo. |
| GASA7 | 11.62 | 9.22 | C(11.4), K(11.4), P (8.6) | 38.76 | 57.62 | −0.233 | extr., vacu. |
| GASA8 | 12.27 | 8.66 | C(10.7), L(9.8), G(8.9) | 45.33 | 80.80 | 0.046 | extr., chlo., vacu. |
| GASA9 | 9.71 | 8.96 | C(13.5), K(13.5), S(10.1) | 44.76 | 54.83 | −0.206 | extr., chlo., vacu. |
| GASA10 | 10.35 | 8.50 | C(12.1), A(10.1), S(10.1) | 47.05 | 63.23 | −0.143 | extr., vacu. |
| GASA11 | 12.62 | 9.52 | S(11.5), C(10.6), K(10.6) | 50.04 | 68.23 | −0.344 | extr. |
| GASA12 | 8.36 | 9.00 | C(16), K(10.7), A(10.7) | 42.75 | 49.47 | −0.417 | mito., chlo., cyto. |
| GASA13 | 7.42 | 8.50 | C(17.2), K(10.9), Y(9.4) | 42.31 | 41.09 | −0.492 | nucl., cyto., mito. |
| GASA14 | 11.96 | 8.80 | C(12.3), L(12.3), K(11.3) | 45.71 | 81.79 | 0.103 | chlo., nucl., extr. |
| Gene1 | Gene2 | Ka | Ks | Ka/Ks | Selection Pressure | Gene Duplications |
|---|---|---|---|---|---|---|
| GASA7 | GASA6 | 0.156 | 1.5871 | 0.0928 | Purifying selection | Segmental |
| GASA8 | GASA5 | 0.4568 | 1 | 0.4568 | Purifying selection | Segmental |
| GASA9 | GASA2 | 0.1985 | 0.929 | 0.213 | Purifying selection | Segmental |
| GASA3 | GASA9 | 0.1781 | 1.1731 | 0.151 | Purifying selection | Segmental |
| GASA12 | GASA11 | 0.196 | 0.333 | 0.585 | Purifying selection | Tandem |
| GASA14 | GASA13 | 0.040 | 0.074 | 0.540 | Purifying selection | Tandem |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, B.; Yao, J.; Zhang, S.; Li, X.; Zhang, X.; Yadav, V.; Wang, X. Genome-Wide Characterization and Expression Profiling of GASA Genes during Different Stages of Seed Development in Grapevine (Vitis vinifera L.) Predict Their Involvement in Seed Development. Int. J. Mol. Sci. 2020, 21, 1088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031088
Ahmad B, Yao J, Zhang S, Li X, Zhang X, Yadav V, Wang X. Genome-Wide Characterization and Expression Profiling of GASA Genes during Different Stages of Seed Development in Grapevine (Vitis vinifera L.) Predict Their Involvement in Seed Development. International Journal of Molecular Sciences. 2020; 21(3):1088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031088
Chicago/Turabian StyleAhmad, Bilal, Jin Yao, Songlin Zhang, Xingmei Li, Xiuming Zhang, Vivek Yadav, and Xiping Wang. 2020. "Genome-Wide Characterization and Expression Profiling of GASA Genes during Different Stages of Seed Development in Grapevine (Vitis vinifera L.) Predict Their Involvement in Seed Development" International Journal of Molecular Sciences 21, no. 3: 1088. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031088