Processed Food Additive Microbial Transglutaminase and Its Cross-Linked Gliadin Complexes Are Potential Public Health Concerns in Celiac Disease


Abstract
:1. Introduction
1.1. Microbial Transglutaminase: Features and Functions in Food Industries
1.2. Enteric Luminal Sources of Microbial Transglutaminase
1.3. Celiac Disease in a Nutshell
2. Microbial Transglutaminase-Gliadin Cross-Linked Complexes Are Immunogenic in Celiac Disease
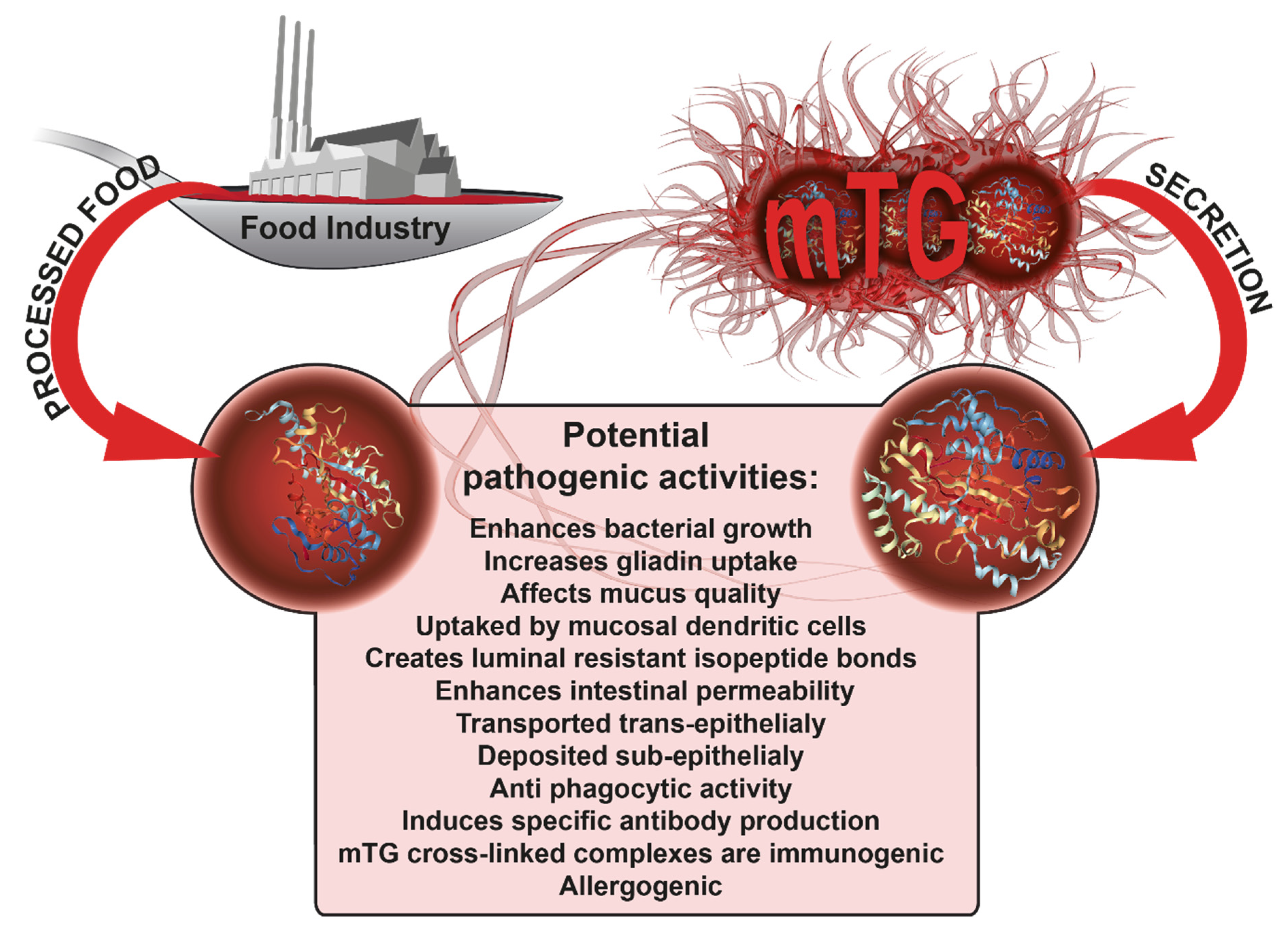
3. Microbial Transglutaminase Is Potentially Pathogenic to CD Patients
3.1. Enhancing Intestinal Permeability.
3.2. Suppression of Mechanical and Immunological Enteric Protective Barriers
3.3. Enhancing Bacterial Growth
3.4. Increasing Uptake of Gliadin Peptide
3.5. mTG and Gliadin Molecules are Cotranscytosed through the Enterocytes
3.6. Potential mTG–Gliadin Uptake by Mucosal Dendritic Cells
3.7. mTG Treated Wheat Products Are Immunoreactive
4. Should Microbial Transglutaminase Be Declared as a Public Health Caveat in Celiac Disease?
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| mTG | microbial transglutaminase |
| tTG | tissue transglutaminase |
| CD | celiac disease |
| GRAS | generally recognized as safe, |
| GFD | gluten-free diet |
References
- Hartung, T. Rebooting the generally recognized as safe (GRAS) approach for food additive safety in the US. ALTEX Altern. Anim. Exp. 2018, 35, 3–25. [Google Scholar] [CrossRef]
- Hartung, T.; Koeter, H. Food for thought… on food safety testing. ALTEX 2008, 25, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://www.gao.gov/products/GAO-10-246 (accessed on 5 March 2010).
- Trasande, L.; Shaffer, R.M.; Sathyanarayana, S.; Council on Environmental Health. Food Additives and Child Health. Pediatrics 2018, 142, e20181408. [Google Scholar] [CrossRef] [Green Version]
- Trasande, L.; Shaffer, R.M.; Sathyanarayana, S.; Council on Environmental Health. Food Additives and Child Health. Pediatrics 2018, 142, e20181410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun. Rev. 2015, 14, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T. Microbial transglutaminase: A new potential player in celiac disease. Clin. Immunol. 2019, 199, 37–43. [Google Scholar] [CrossRef]
- Kieliszek, M.; Misiewicz, A. Microbial transglutaminase and its application in the food industry. A review. Folia Microbiol. (Praha) 2014, 59, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, I.M.; Matos, M.; Costa, R.; Silva, F.; Pascoal, A.; Estevinho, L.M.; Choupina, A.B. Transglutaminases: Recent achievements and new sources. Appl. Microbiol. Biotechnol. 2014, 98, 6957–6964. [Google Scholar] [CrossRef]
- Strop, P. Versatility of microbial transglutaminase. Bioconjug. Chem. 2014, 25, 855–862. [Google Scholar] [CrossRef]
- Santhi, D.; Kalaikannan, A.; Malairaj, P.; Arun Prabhu, S. Application of microbial transglutaminase in meat foods: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 2071–2076. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Possible association between celiac disease and bacterial transglutaminase in food processing: A hypothesis. Nutr. Rev. 2015, 73, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Matthias, T. Microbial Transglutaminase is Beneficial to Food Industries but a Caveat to Public Health. Med. One 2019, 4, e190001. [Google Scholar] [CrossRef]
- Matthias, T.; Lerner, A. Microbial transglutaminase is immunogenic and potentially pathogenic in pediatric celiac disease. Front. Pediatr. 2018, 6, 389. [Google Scholar] [CrossRef]
- Lerner, A.; Ramesh, A.; Matthias, T. David and Goliath war revival in the enteric viruses and microbiota struggle. Potential implication for celiac disease. Microorganisms 2019, 7, 173. [Google Scholar]
- Makarova, K.S.; Aravind, L.; Koonin, E.V. A superfamily of archaeal, bacterial, and eukaryotic proteins homologous to animal transglutaminases. Protein Sci. 1999, 8, 1714–1719. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, V.; Koonin, E.V.; Aravind, L. Peptide-N-glycanases and DNA repair proteins, Xp-C/Rad4, are, respectively, active and inactivated enzymes sharing a common transglutaminase fold. Hum. Mol. Genet. 2001, 10, 1627–1630. [Google Scholar] [CrossRef] [Green Version]
- Iyer, L.M.; Balaji, S.; Koonin, E.V.; Aravind, L. Evolutionary genomics of nucleo-cytoplasmic large DNA viruses. Virus Res. 2006, 117, 156–184. [Google Scholar] [CrossRef] [Green Version]
- Devoto, A.E.; Santini, J.M.; Olm, M.R.; Anantharaman, K.; Munk, P.; Tung, J.; Archie, E.A.; Turnbaugh, P.J.; Seed, K.D.; Blekhman, R.; et al. Megaphages infect Prevotella and variants are widespread in gut microbiomes. Nat. Microbiol. 2019, 4, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Fatima, S.W.; Khare, S.K. Current insight and futuristic vistas of microbial transglutaminase in nutraceutical industry. Microbiol. Res. 2018, 215, 7–14. [Google Scholar] [CrossRef]
- Yokoyama, K.; Nio, N.; Kikuchi, Y. Properties and applications of microbial transglutaminase. Appl. Microbiol. Biotechnol. 2004, 64, 447–454. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, Y.; Chen, J. Microbial transglutaminase production: Understanding the mechanism. Biotechnol. Genet. Eng. Rev. 2010, 26, 205–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malandain, H. Transglutaminases: A meeting point for wheat allergy, celiac disease, and food safety. Eur. Ann. Allergy Clin. Immun. 2005, 37, 397–403. [Google Scholar]
- Lerner, A.; Aminov, R.; Matthias, T. Dysbiosis may trigger autoimmune diseases via inappropriate posttranslational modification of host proteins. Front. Microbiol. 2016, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Aminov, R.; Matthias, T. Intestinal dysbiotic transglutaminases are potential environmental drivers of systemic autoimmunogenesis. Front. Microbiol. 2017, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Aminov, R.; Matthias, T. Potential effects of horizontal gene exchange in the human gut. Front. Immunol. 2017, 8, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Shoenfeld, Y.; Matthias, T. Probiotics: If it does not help it does not do any harm. Really? Microorganisms 2019, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- Del Duca, S.; Serafini-Fracassini, D.; Cai, G. Senescence and programmed cell death in plants: Polyamine action mediated by transglutaminase. Front. Plant Sci. 2014, 5, 120. [Google Scholar] [CrossRef] [Green Version]
- Del Duca, S.; Bonner, P.L.R.; Aloisi, I.; Serafini-Fracassini, D.; Cai, G. Determination of Transglutaminase Activity in Plants. Methods Mol. Biol. 2018, 1694, 173–200. [Google Scholar] [CrossRef]
- Skovbjerg, H.; Norén, O.; Anthonsen, D.; Moller, J.; Sjöström, H. Gliadin is a good substrate of several transglutaminases: Possible implication in the pathogenesis of coeliac disease. Scand. J. Gastroenterol. 2002, 37, 812–817. [Google Scholar] [CrossRef]
- Jira, W.; Schwägele, F. A sensitive high performance liquid chromatography-tandem mass spectrometry method for the detection of microbial transglutaminase in different types of restructured meat. Food Chem. 2017, 221, 1970–1978. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, A.; Koppel, R.; Widmer, M. Determination of microbial transglutaminase in meat and meat products. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Iranzo, M.; Aguado, C.; Pallotti, C.; Cañizares, J.V.; Mormeneo, S. Transglutaminase activity is involved in Saccharomyces cerevisiae wall construction. Microbiology 2002, 148, 1329–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, L.W.; Onishi, M.; Pringle, J.R.; Lord, M. Fission yeast Cyk3p is a transglutaminase-like protein that participates in cytokinesis and cell morphogenesis. Mol. Biol. Cell 2012, 23, 2433–2444. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Boorsma, A.; De Groot, P.W. Cell wall construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Reyna-Beltrán, E.; Iranzo, M.; Calderón-González, K.G.; Mondragón-Flores, R.; Labra-Barrios, M.L.; Mormeneo, S.; Luna-Arias, J.P. The Candida albicans ENO1 gene encodes a transglutaminase involved in growth, cell division, morphogenesis, and osmotic protection. J. Biol. Chem. 2018, 293, 4304–4323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corouge, M.; Loridant, S.; Fradin, C.; Salleron, J.; Damiens, S.; Moragues, M.D.; Souplet, V.; Jouault, T.; Robert, R.; Dubucquoi, S.; et al. Humoral immunity links Candida albicans infection and celiac disease. PLoS ONE 2015, 10, e0121776. [Google Scholar] [CrossRef]
- Caio, G.; Volta, U.; Sapone, A.; Leffler, D.A.; De Giorgio, R.; Catassi, C.; Fasano, A. Celiac disease: A comprehensive current review. BMC Med. 2019, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Reif, S.; Lerner, A. Tissue transglutaminase—The key player in celiac disease: A review. Autoimmun. Rev. 2004, 3, 40–45. [Google Scholar] [CrossRef]
- Lerner, A.; Neidhöfer, S.; Matthias, T. Transglutaminase 2 and anti transglutaminase 2 autoantibodies in celiac disease and beyond: Part A: TG2 double-edged sword: Gut and extraintestinal involvement. Immunome Res. 2015, 11, 101–105. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Agmon-Levin, N.; Shapira, Y.; Gilburd, B.; Reuter, S.; Lavi, I.; Shoenfeld, Y. The thrombophilic network of autoantibodies in celiac disease. BMC Med. 2013, 11, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciacci, C.; Ciclitira, P.; Hadjivassiliou, M.; Kaukinen, K.; Ludvigsson, J.F.; McGough, N.; Sanders, D.S.; Woodward, J.; Leonard, J.N.; Swift, G.L. The gluten-free diet and its current application in coeliac disease and dermatitis herpetiformis. United Eur. Gastroenterol. J. 2015, 3, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Samasca, G.; Lerner, A.; Sur, G.; Lupan, L.; Makovicky, P.; Matthias, T.; Freeman, H.J. Challenging in Gluten-free diet in celiac disease: Prague consensus. Eur. J. Clin. Investig. 2017, 47, 394–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T. Gluten free diet-tough alley in torrid time. Int. J. Celiac Dis. 2017, 5, 50–55. [Google Scholar]
- Lerner, A.; Shoenfeld, Y.; Matthias, T. Adverse effects of gluten ingestion and advantages of gluten withdrawal in nonceliac autoimmune disease. Nutr. Rev. 2017, 75, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; O’Bryan, T.; Matthias, T. Navigating the gluten-free diet boom: The dark side of gluten free diet. Front. Pediatr. 2019, 7, 414. [Google Scholar] [CrossRef] [Green Version]
- Ludvigsson, J.F.; Green, P.H. The missing environmental factor in celiac disease. N. Engl. J. Med. 2014, 371, 1341–1343. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Jeremias, P.; Matthias, T. The world incidence of celiac disease is increasing: A review. Int. J. Recent Sci. Res. 2015, 7, 5491–5496. [Google Scholar]
- Lerner, A.; Jeremias, P.; Matthias, T. The world incidence and prevalence of autoimmune diseases is increasing: A review. Int. J. Celiac Dis. 2015, 3, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Jeremias, P.; Neidhöfer, S.; Matthias, T. Comparison of the reliability of 17 celiac disease associated bio-markers to reflect intestinal damage. J. Clin. Cell Immunol. 2017, 8, 486. [Google Scholar] [CrossRef] [Green Version]
- Agardh, D.; Matthias, T.; Wusterhausen, P.; Neidhöfer, S.; Heller, A.; Lerner, A. Antibodies against neo-epitope of microbial and human transglutaminases’ complexes as biomarkers of childhood celiac disease. Clin. Exp. Immunol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Ramesh, A.; Matthias, T. Serological diagnosis of celiac disease: New biomarkers. Gastroenterol. Clin. N. Am. 2019, 48, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Matthias, T. Gluten and autoimmunogenesis. In Musaic of Autoimmunity, 2nd ed.; Shoenfield, Y., Perricone, C., Eds.; the Novel Factors of Autoimmune Diseases Revisited; Elsevier: Amsterdam, The Netherlands, 2019; pp. 315–321. [Google Scholar]
- Lerner, A.; Jeremias, P.; Neidhöfer, S.; Matthias, T. Antibodies against neo-epitope tTg complexed to gliadin are different and more reliable then anti-tTg for the diagnosis of pediatric celiac disease. J. Immunol. Methods 2016, 429, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T. Microbial transglutaminase should be considered as an environmental inducer of celiac disease. World J. Clin. Cases 2019, 7, 3912–3914. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Don’t forget the exogenous microbial transglutaminases: It is immunogenic and potentially pathogenic. AIMS Biophys. 2016, 3, 546–552. [Google Scholar] [CrossRef]
- Matthias, T.; Jeremias, P.; Neidhöfer, S.; Lerner, A. The industrial food additive microbial transglutaminase, mimics the tissue transglutaminase and is immunogenic in celiac disease patients. Autoimmun. Rev. 2016, 15, 1111–1119. [Google Scholar] [CrossRef]
- Stricker, S.; de Laffolie, J.; Rudloff, S.; Komorowski, L.; Zimmer, K.P. Intracellular Localization of Microbial Transglutaminase and Its Influence on the Transport of Gliadin in Enterocytes. J. Pediatr. Gastroenterol. Nutr. 2019, 68, e43–e50. [Google Scholar] [CrossRef]
- Fasano, A. Zonolin and its regulation of intestinal barrier function: The biological door to inflammation, autoimmunity, and cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef] [Green Version]
- Hollon, J.; Puppa, E.L.; Greenwald, B.; Goldberg, E.; Guerrerio, A.; Fasano, A. Effect of gliadin on permeability of intestinal biopsy explants from celiac disease patients and patients with non-celiac gluten sensitivity. Nutrients 2015, 7, 1565–1576. [Google Scholar] [CrossRef]
- Cardoso-Silva, D.; Delbue, D.; Itzlinger, A.; Moerkens, R.; Withoff, S.; Branchi, F.; Schumann, M. Intestinal Barrier Function in Gluten-Related Disorders. Nutrients 2019, 11, 2325. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Ren, J.; Zhao, M.; Cui, C.; He, P. Emulsifying properties of the transglutaminase-treated crosslinked product between peanut protein and fish (Decapterus maruadsi) protein hydrolysates. J. Sci. Food Agric. 2011, 91, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhao, M.; Sun, W.; Zhao, G.; Ren, J. Effects of microfluidization treatment and transglutaminase cross-linking on physicochemical, functional, and conformational properties of peanut protein isolate. J. Agric. Food Chem. 2011, 59, 8886–8894. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.S.; Hu, X.C.; Zhang, K.; Xu, X.K.; Zhang, H.M.; Huang, H. Characterization of spray-dried microalgal oil encapsulated in cross-linked sodium caseinate matrix induced by microbial transglutaminase. J. Food Sci. 2011, 76, E112–E118. [Google Scholar] [CrossRef] [PubMed]
- Brenzetti, S.; Dal Bello, F.; Arendt, E.K. Microstructural, fundamental rheology, and baking characteristics of batters and breads from different gluten-free flours treated with a microbial transglutaminase. J. Cereal Sci. 2008, 48, 33–45. [Google Scholar] [CrossRef]
- Li, F.; Lu, J.; Kong, X.; Hyeon, T.; Ling, D. Dynamic Nanoparticle Assemblies for Biomedical Applications. Adv. Mater. 2017, 29, 1605897. [Google Scholar] [CrossRef]
- Fuchs, S.; Kutscher, M.; Hertel, T.; Winter, G.; Pietzsch, M.; Coester, C. Transglutaminase: New insights into gelatin nanoparticle cross-linking. J. Microencapsul. 2010, 27, 747–754. [Google Scholar] [CrossRef]
- Groh, K.J.; Geueke, B.; Muncke, J. Food contact materials and gut health: Implications for toxicity assessment and relevance of high molecular weight migrants. Food Chem. Toxicol. 2017, 109 Pt 1, 1–18. [Google Scholar] [CrossRef]
- Li, N.; Neu, J. Glutamine deprivation alters intestinal tight junctions via a PI3-K/Akt mediated pathway in Caco-2 cells. J. Nutr. 2009, 139, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Neidhöfer, S.; Matthias, T. The gut microbiome feelings of the brain: Perspective for Non-Microbiologists. Microorganisms 2017, 5, 66. [Google Scholar] [CrossRef]
- Boukhettala, N.; Claeyssens, S.; Bensifi, M.; Maurer, B.; Abed, J.; Lavoinne, A.; Déchelotte, P.; Coëffier, M. Effects of essential amino acids or glutamine deprivation on intestinal permeability and protein synthesis in HCT-8 cells: Involvement of GCN2 and mTOR pathways. Amino Acids 2012, 42, 375–383. [Google Scholar] [CrossRef]
- Kimura, H. Histone modifications for human epigenome analysis. J. Hum. Genet. 2013, 58, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Nam, K.H.; Kwon, O.S.; Kim, I.G.; Bustin, M.; Choy, H.E.; Park, S.C. Histone cross-linking by transglutaminase. Biochem. Biophys. Res. Commun. 2002, 293, 1453–1457. [Google Scholar] [CrossRef]
- Kocsis, D.; Béres, N.; Veres, G.; Szabó, D.; Müller, K.E.; Arató, A.; Juhász, M. Genetic and epigenetic aspects of celiac disease. Orv. Hetil. 2014, 155, 83–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, A.S.; Baird, A.M.; Gray, S.G. Epigenetic Methodologies for the Study of Celiac Disease. Methods Mol. Biol. 2015, 1326, 131–158. [Google Scholar] [CrossRef] [PubMed]
- Recktenwald, C.V.; Hansson, G.C. The Reduction-insensitive Bonds of the MUC2 Mucin Are Isopeptide Bonds. J. Biol. Chem. 2016, 291, 13580–13590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, R.U.; Mehta, K. Transglutaminases, thioredoxins and protein disulphide isomerase: Diverse enzymes with a common goal of cross-linking proteins in lower organisms. Indian J. Exp. Biol. 2004, 42, 235–243. [Google Scholar]
- Tagami, U.; Shimba, N.; Nakaamura, M.; Yokoyama, K.; Suzuki, E.; Hirokawa, T. Substrate specificity of microbial transglutaminase as revealed by three-dimentional docking simulation and mutagenesis. Protein Eng. Des. Sel. 2009, 22, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Pian, Y.; Wang, P.; Liu, P.; Zheng, Y.; Zhu, L.; Wang, H.; Xu, B.; Yuan, Y.; Jiang, Y. Proteomics identification of novel fibrinogen-binding proteins of Streptococcus suis contributing to antiphagocytosis. Front. Cell. Infect. Microbiol. 2015, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Zhang, P.; Li, W.; Liu, R.; Tang, J.; Fan, H. hsdS, Belonging to the Type I Restriction-Modification System, Contributes to the Streptococcus suis Serotype 2 Survival Ability in Phagocytes. Front. Microbiol. 2017, 8, 1524. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Qin, W.; Zhu, H.; Wang, X.; Jiang, J.; Hu, J. How Streptococcus suis serotype 2 attempts to avoid attack by host immune defenses. J. Microbiol. Immunol. Infect. 2019, 52, 516–525. [Google Scholar] [CrossRef]
- Zhao, J.; Pan, S.; Lin, L.; Fu, L.; Yang, C.; Xu, Z.; Wei, Y.; Jin, M.; Zhang, A. Streptococcus suis serotype 2 strains can induce the formation of neutrophil extracellular traps and evade trapping. FEMS Microbiol. Lett. 2015, 362, fnv022. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.Y.; Chen, J.; Li, Y. Heterologous leaky production of transglutaminase in Lactococcus lactis significantly enhances the growth performance of the host. Appl. Environ. Microbiol. 2005, 71, 8911–8919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.Y.; Chen, J.; Li, Y. Influence of expression of transglutaminase on the growth of Lactococcus lactis. Wei Sheng Wu Xue Bao 2005, 45, 510–515. [Google Scholar] [PubMed]
- Lebreton, C.; Menard, S.; Abed, J.; Moura, I.C.; Coppo, R.; Dugave, C.; Monteiro, R.C.; Fricot, A.; GarfaTraore, M.; Griffin, M.; et al. Interactions among secretory immunoglobulin A, CD71, and transglutaminase-2 affect permeability of intestinal epithelial cells to gliadin peptides. Gastroenterology 2012, 143, 698–707. [Google Scholar] [CrossRef]
- Salmi, T.T.; Collin, P.; Jarvinen, O.; Haimila, K.; Partanen, J.; Laurila, K.; Korponay-Szabo, I.R.; Huhtala, H.; Reunala, T.; Maki, M.; et al. Immunoglobulin A autoantibodies against transglutaminase 2 in the small intestinal mucosa predict forthcoming coeliac disease. Aliment. Pharmacol. Ther. 2006, 24, 541–552. [Google Scholar] [CrossRef]
- Borrelli, M.; Maglio, M.; Korponay-Szabó, I.R.; Vass, V.; Mearin, M.L.; Meijer, C.; Niv-Drori, H.; Ribes-Koninckx, C.; Roca, M.; Shamir, R.; et al. Intestinal anti-transglutaminase 2 immunoglobulin A deposits in children at risk for coeliac disease (CD): Data from the PreventCD study. Clin. Exp. Immunol. 2018, 191, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Killinger, B.A.; Labrie, V. Vertebrate food products as a potential source of prion-like α-synuclein. NPJ Parkinsons Dis. 2017, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Owen, J.L.; Mohamadzadeh, M. Microbial activation of gut dendritic cells and the control of mucosal immunity. J. Interferon Cytokine Res. 2013, 33, 619–631. [Google Scholar] [CrossRef] [Green Version]
- Stagg, A.J. Intestinal Dendritic Cells in Health and Gut Inflammation. Front. Immunol. 2018, 9, 2883. [Google Scholar] [CrossRef] [Green Version]
- Ráki, M.; Schjetne, K.W.; Stamnaes, J.; Molberg, Ø.; Jahnsen, F.L.; Issekutz, T.B.; Bogen, B.; Sollid, L.M. Surface expression of transglutaminase 2 by dendritic cells and its potential role for uptake and presentation of gluten peptides to T cells. Scand. J. Immunol. 2007, 65, 213–220. [Google Scholar] [CrossRef]
- Hodrea, J.; Demény, M.A.; Majai, G.; Sarang, Z.; Korponay-Szabó, I.R.; Fésüs, L. Transglutaminase 2 is expressed and active on the surface of human monocyte-derived dendritic cells and macrophages. Immunol. Lett. 2010, 130, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Berti, C.; Roncoroni, L.; Falini, M.L.; Caramanico, R.; Dolfini, E.; Bardella, M.T.; Elli, L.; Terrani, C.; Forlani, F. Celiac-related properties of chemically and enzymatically modified gluten proteins. J. Agric. Food Chem. 2007, 55, 2482–2488. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Cha’vez, F.; Rouzaud-Sa´ndez, O.; Sotelo-Cruz, N.; Calderón de la Barca, A.M. Bovine milk caseins and transglutaminase-treated cereal prolamines are differentially recognized by IgA of celiac disease patients according to age. J. Agric. Food Chem. 2009, 57, 3754–3759. [Google Scholar] [CrossRef] [PubMed]
- Elli, L.; Roncoroni, L.; Hils, M.; Pasternack, R.; Barisani, D.; Terrani, C.; Vaira, V.; Ferrero, S.; Bardella, M.T. Immunological effects of transglutaminase-treated gluten in coeliac disease. Hum. Immunol. 2012, 73, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Cha´vez, F.; Rouzaud-Sa´ndez, O.; Sotelo-Cruz, N.; Calderón de la Barca, A.M. Transglutaminase treatment of wheat and maize prolamines of bread increase the serum IgA reactivity of celiac disease patients. J. Agric. Food Chem. 2008, 56, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Falini, M.L.; Elli, L.; Caramanico, R.; Bardella, M.T.; Terrani, C.; Roncoroni, L.; Doneda, L.; Forlani, F. Immunoreactivity of antibodies against transglutaminase-deamidated gliadins in adult celiac disease. Dig. Dis. Sci. 2008, 53, 2697–2701. [Google Scholar] [CrossRef] [PubMed]
- Heredia-Sandoval, N.G.; Islas-Rubio, A.R.; Cabrera-Cha´vez, F.; de la Barca, A.M.C. Transamidation of gluten proteins during the bread-making process of wheat flour to produce breads with less immunoreactive gluten. Food Funct. 2014, 5, 1813–1818. [Google Scholar] [CrossRef]
- Dekking, E.H.A.; Van Veblen, P.A.; de Ru, A.; Kooy-Winkelaar, E.M.C.; Gröneveld, T.; Nieuwenhuizen, W.F.; Koning, F. Microbial transglutaminases generate T cell stimulatory epitopes involved in celiac disease. J. Cereal Sci. 2008, 47, 339–346. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Food Industrial Microbial Transglutaminase in Celiac Disease: Treat or Trick. Int. J. Celiac Dis. 2015, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Kooy-Winkelaar, Y.M.; Cordfunke, R.A.; Dragan, I.; Thompson, A.; Drijfhout, J.W.; Peter, A.; Chen, V.V.H.; Koning, F. Abrogation of Immunogenic Properties of Gliadin Peptides through Transamidation by Microbial Transglutaminase Is Acyl-Acceptor Dependent. J. Agric. Food Chem. 2017, 65, 7542–7552. [Google Scholar] [CrossRef]
- Heil, A.; Ohsam, J.; van Genugten, B.; Diez, O.; Yokoyama, k.; Kumazawa, Y.; Pasternack, R.; Hils, M. Microbial Transglutaminase Used in Bread Preparation at Standard Bakery Concentrations Does Not Increase Immunodetectable Amounts of Deamidated Gliadin. J. Agric. Food Chem. 2017, 65, 6982–6990. [Google Scholar] [CrossRef] [PubMed]
- Scarnato, L.; Gadermaier, G.; Volta, U.; De Giorgio, R.; Caio, G.; Lanciotti, R.; Del Duca, S. Immunoreactivity of Gluten-Sensitized Sera Toward Wheat, Rice, Corn, and Amaranth Flour Proteins Treated with Microbial Transglutaminase. Front. Microbiol. 2019, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, C.; Barisani, D. Food additives can act as triggering factors in celiac disease: Current knowledge based on a critical review of the literature. World J. Clin. Cases 2019, 7, 917–927. [Google Scholar] [CrossRef]
- Heil, A.; Ohsam, J.; Büchold, C.; Pasternack, R.; Yokoyama, K.; Kumazawa, Y.; Hils, M. Microbial transglutaminase has a lower deamidation preference than human tissue transglutaminase on a celiac disease relevant wheat gliadin T-cell epitope. J. Cereal Sci. 2016, 70, 47–56. [Google Scholar] [CrossRef]
- Available online: https://www.fda.gov/food-ingredients-packaging/generally-recognized-safe-gras (accessed on 6 September 2019).
- Available online: https://www.ecfr.gov/cgi-bin/text-idx/SID=e956d645a8b4e6b3e34e4e5d1b690209&184.1150 (accessed on 10 December 2019).
- Available online: https://www.fda.gov/food/food-additives-petitions (accessed on 24 October 2019).
- Herman, L.; Chemaly, M.; Cocconcelli, P.S.; Fernandez, P.; Klein, G.; Peixe, L.; Prieto, M.; Querol, A.; Suarez, J.E.; Sundh, I.; et al. The qualified presumption of safety assessment and its role in EFSA risk evaluations: 15 years past. FEMS Microbiol. Lett. 2019, 366 (Suppl. 1), i17–i23. [Google Scholar] [CrossRef]


{kind=link}
| MTG Source | Reference | |
|---|---|---|
| Extra-Intestinal | Processed food additive | [6,8,9,11,12,13,14,20] |
| Pathobionts | [24,25,26] | |
| Probiotics | [27,28] | |
| Plants | [29,30] | |
| Vegetables | [31] | |
| Meat | [32,33] | |
| Intra-Intestinal | Microbiome | [7,12,13,14,24,25] |
| Dysbiome | [7,12,13,14,24,25] | |
| Yeasts | [34,35,36] | |
| Fungi | [37,38] | |
| Viruses | [15,16,17,18] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lerner, A.; Matthias, T. Processed Food Additive Microbial Transglutaminase and Its Cross-Linked Gliadin Complexes Are Potential Public Health Concerns in Celiac Disease. Int. J. Mol. Sci. 2020, 21, 1127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031127
Lerner A, Matthias T. Processed Food Additive Microbial Transglutaminase and Its Cross-Linked Gliadin Complexes Are Potential Public Health Concerns in Celiac Disease. International Journal of Molecular Sciences. 2020; 21(3):1127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031127
Chicago/Turabian StyleLerner, Aaron, and Torsten Matthias. 2020. "Processed Food Additive Microbial Transglutaminase and Its Cross-Linked Gliadin Complexes Are Potential Public Health Concerns in Celiac Disease" International Journal of Molecular Sciences 21, no. 3: 1127. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031127