1. Introduction
With the widespread use of antibiotics in clinical practice in recent years, the resistance rate of bacteria to traditional antibiotics is increasing. The development of antibacterial reagents with new antibacterial mechanisms is imminent [
1]. As an important part of innate immunity, antimicrobial peptides (AMPs) form the first line of defense against antimicrobial infections, as found in many microorganisms including insects, plants, and animals [
2]. Furthermore, AMPs are known for their anti-inflammatory properties, anti-biofilm formation, promotion of tissue and injury repair, and anti-cancer effects [
2,
3,
4]. Compared to traditional antibiotics, AMPs exert an antibacterial effect through non-receptor-mediated membrane permeability. AMPs are expected to be alternatives to antibiotics because of their unique action mechanism [
5,
6,
7]. Although more than 3000 AMPs were found, their application as therapeutic agents is seriously hindered due to some limiting factors including manufacturing costs, cytotoxicity, and poor stability [
8].
The heptad repeat sequence is a repeat sequence composed of seven amino acids, where the “a” and “d” positions are occupied by hydrophobic amino acids [
9]. It was found that natural AMPs possessing heptad repeat sequences have great antibacterial and anti-inflammatory activity, but their application is hindered by high cytotoxicity [
10,
11]. Previous studies mostly focused on how different hydrophobic amino acids at the “a” and “d” positions influence antibacterial activity and cytotoxicity of AMPs, but few studies used this sequence element to design new AMPs [
12,
13]. Some papers showed that symmetric AMPs centered on Gly–Gly (GG) or
d-Pro–Gly (pG) have good antibacterial activity and high cell selectivity [
14,
15,
16,
17]. Therefore, we wondered if the same effect could be achieved when GG or pG was introduced into two symmetric heptad repeat sequences.
In this study, two symmetric heptad repeat sequences were connected by a short loop, and the sequence was designed as XXRXXRRzzRRXXRXX-NH
2, where X represents Phe (F), Ile (I), and Leu (L), while zz represents Gly–Gly (GG) or
d-Pro–Gly (pG). The net charge was +6 and the hydrophobicity was 40%–60%, which is consistent with the statistical information on natural AMPs [
2,
18]. Hydrophobic amino acids F, I, and L were used to damage microbial cell membranes [
19]. Flexible (GG) and rigid (pG) loops were selectively inserted into the middle of two symmetric heptad repeat sequences to enhance cell selectivity [
17,
20]. In addition, Arg (R) was selectively introduced into the sequence template. The guanidine side chain of R allows it to form strong bidentate H-bonds with the phosphate moiety of lipid head groups, thereby enhancing the electrostatic action between peptides and anionic bacterial membranes [
20]. To determine the role of loops in the designed peptides, a control peptide was designed as a heptad repeat sequence as follows: the central pG loop of peptide LR
pG with highest cell selectivity was removed, and Leu located at the N-terminus and C-terminus of the peptide was exchanged with Gly and
d-Pro, respectively. The positive charge and hydrophobicity were consistent with LR
pG. Finally, by amidating the C-terminus of the peptides, the charge of the AMPs was increased, and the stability of the structure was improved [
21,
22,
23]. Using this method of designing and optimizing AMPs, we obtained LR
pG (X: L, zz: pG) with high cell selectivity and good conditional stability. The antibacterial mechanism study demonstrated that LR
pG played the role of an anti-Gram-negative bacterial agent through destroying the cell membrane.
3. Discussion
Most AMPs have unique mechanisms of killing bacteria, mainly involving membrane breaking, which endows them with the potential to be alternatives to antibiotics [
2]. However, the progress of natural AMPs for therapeutic application is seriously hindered by their inherent defects (such as manufacturing costs, cytotoxicity, poor stability etc.) [
2,
8]. To overcome these inherent shortcomings, in this study, six symmetric heptad repeat sequences were connected by a short loop, and the sequence was designed as XXRXXRRzzRRXXRXX-NH
2 (X represents F, I, and L; zz represents GG or pG).
The results of MICs indicated that the designed peptides had good antibacterial activity toward Gram-negative bacteria (
Table 2). Compared to the single membrane and thick peptidoglycan layer of Gram-positive bacteria, the double-membrane structure of Gram-negative bacteria represents a weaker barrier; thus, Gram-negative bacteria are more sensitive to AMPs [
17,
21,
24]. The antibacterial activity of AMPs can be affected by many factors. Previous studies demonstrated that a proper positive charge (+6 to +7) was essential for antibacterial activity, but antibacterial activity was no longer increased when the positive charge of AMPs was beyond the threshold [
17,
20]. In this study, the positive charge of the designed peptides was set to +6. Based on their hydrophobicity, the theoretical antibacterial activity of the designed peptides would be IR
ZZ > FR
ZZ > LR
ZZ = LRα; however, the result was LRα > LR
ZZ > FR
ZZ > IR
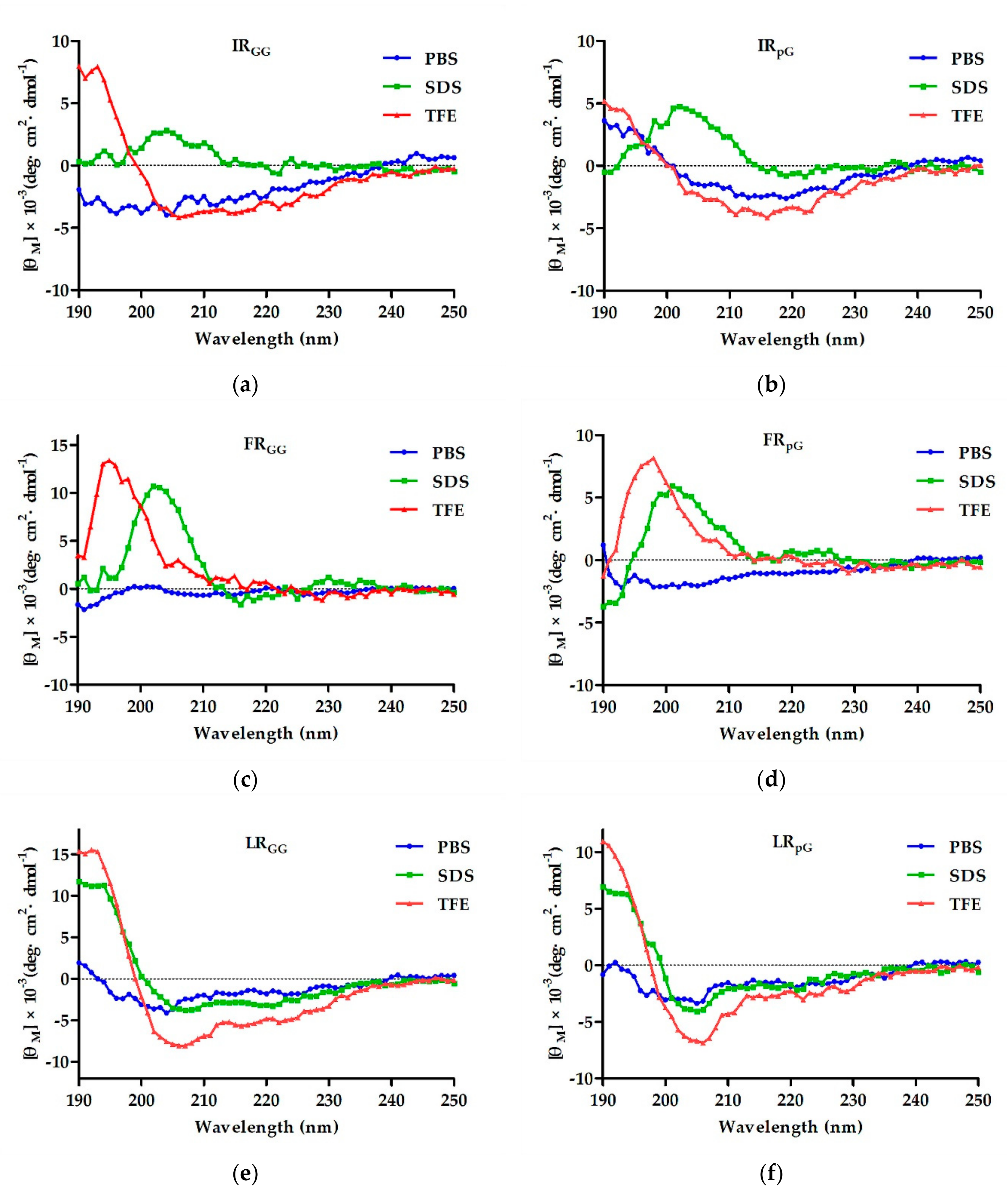
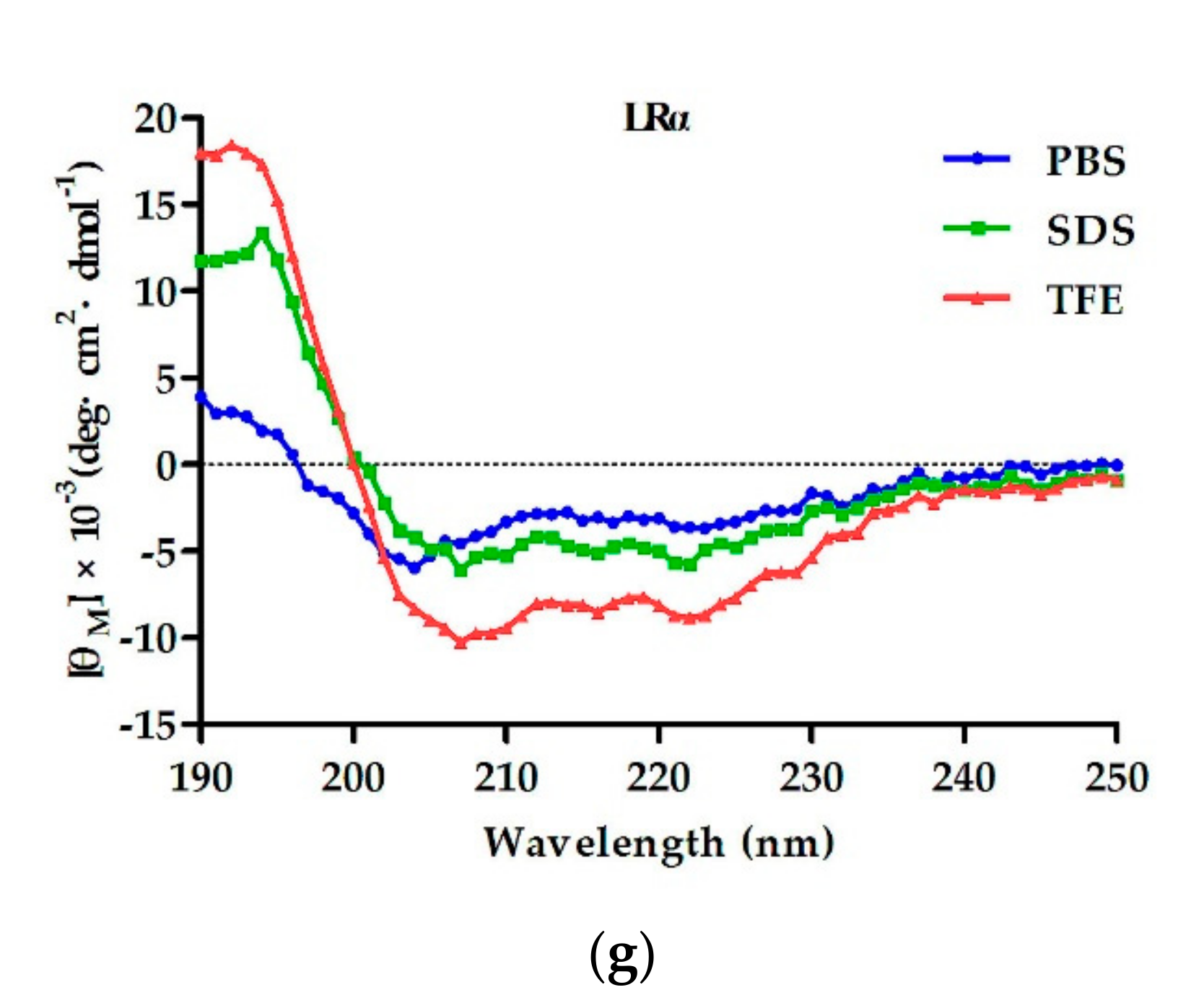
ZZ. Combined with the results of CD, although both LR
ZZ and LRα had the tendency to form an α-helical structure in membrane-mimetic environments, LRα’s tendency was stronger. One possible explanation is that the rigid loop (pG) of LR
ZZ disrupted the α-helical structure. Furthermore, we found that both FR
ZZ and IR
ZZ exhibited disordered conformations in membrane-mimetic environments. The result further confirmed the previous perspectives that the antibacterial activity of designed peptides is not only connected to the hydrophobicity of the peptides but also to the spatial stability of the secondary structure [
25]. In addition, the antibacterial activity of XR
pG was higher than that of XR
GG. Some papers confirmed that the rigid loop (pG), which promotes tightness in the hydrophobic center, has a better effect than the flexible loop (GG) [
20].
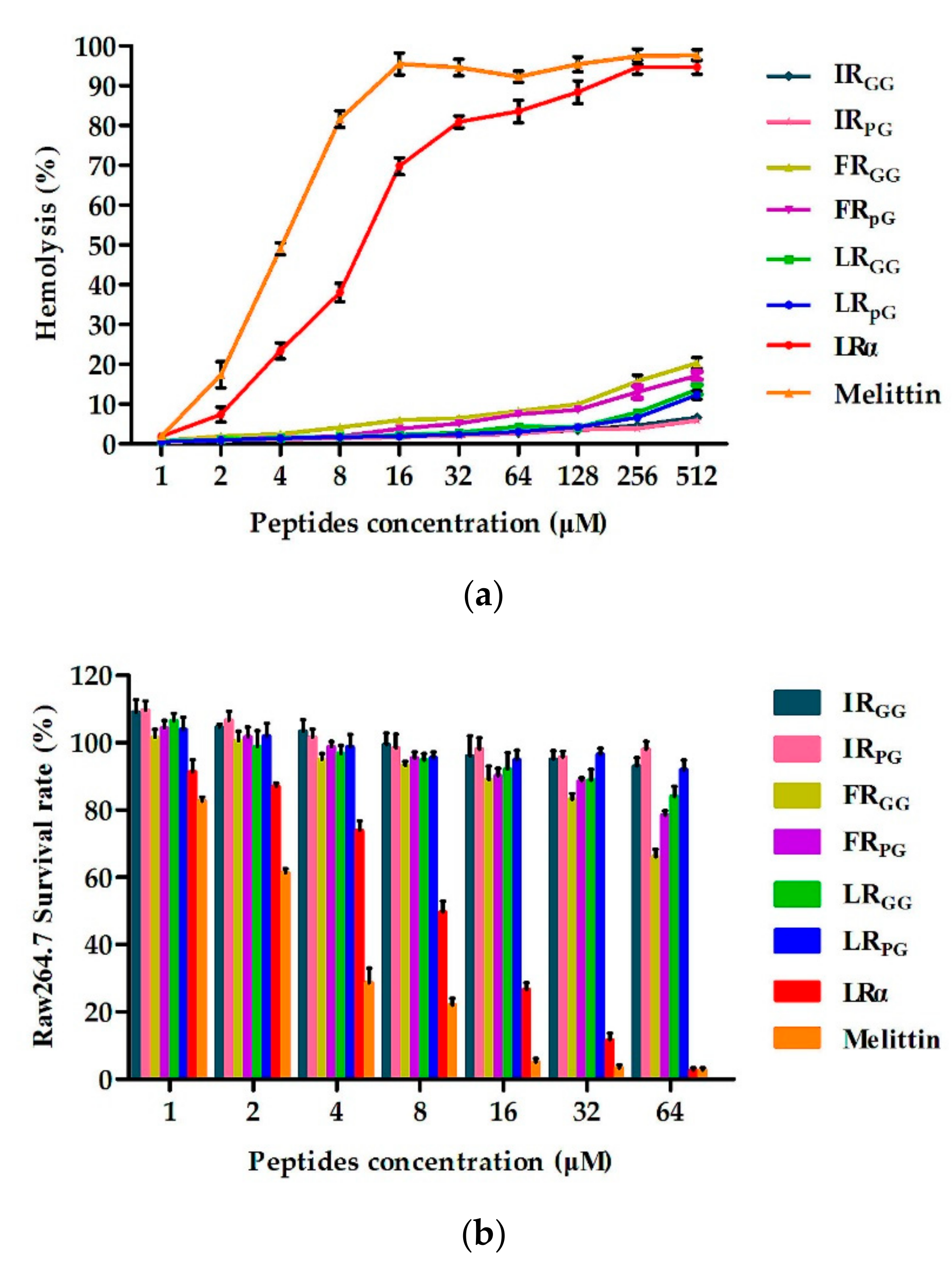
However, the antimicrobial activity of AMPs is not the only criterion when evaluating their potential for clinical application. The hemolytic activity and cytotoxicity to mammalian cells are also major limiting factors [
26,
27]. Therefore, the cytotoxicity and hemolytic activity of the designed peptides were tested. At all examined concentrations, the designed peptides had much lower hemolysis and cytotoxicity (
Figure 2), showing better cell selectivity compared to LRα. This result may be due to the insertion of the central loop as previous papers reported. The cell selectivity of XR
pG was better than that of XR
GG, which may be due to the rigid pG having a better effect on promoting cell selectivity than flexible GG [
20]. As shown in
Table 2, LR
pG had a great balance between antibacterial activity and cytotoxicity, and it had higher cell selectivity.
In addition to the cell selectivity assays of AMPs, stability is also a major reason impeding AMPs from becoming therapeutic agents [
2,
28,
29]. In this study, the effects of the physiological concentrations of salts, temperature, and pH on the antibacterial activity of the designed peptides were investigated. As shown in
Table 3, in salt ions, the antimicrobial activity of LR
pG against
E. coli ATCC25922 was negligibly affected by all tested cations except for Na
+. Previous reports suggested that a stable α-helical structure helps salt tolerance [
30]. However, the insertion of the loop (pG) structure into the sequence template still resulted in good salt stability in this study. The result was due to the loop (pG) promoting hydrogen bonding and hydrophobic interactions, thereby achieving the effect of salt tolerance [
31]. The antibacterial activity of LR
pG against
E. coli ATCC25922 decreased because of the disruption of electrostatic interaction between LR
pG and the cell membrane in the presence of 150 mM Na
+ [
32]. For the clinical application of AMPs, thermal stability is very important since many foods and feed products need to be heated during processing [
33]. LR
pG exerted good thermal stability (
Table 4) and could maintain antibacterial activity in different pH conditions.
Furthermore, LR
pG exhibited a synergistic effect with streptomycin and additive effects with other antibiotics against
E. coli ATCC25922 (
Table 5). Some studies reported that the antibacterial effect of streptomycin and chloramphenicol involved acting on ribosomes, while ciprofloxacin acted on DNA gyrase, and cefotaxime acted on the cell membrane. In this study, LR
pG acted on cell membranes to increase membrane permeability and promote antibiotic entry into cells [
34,
35]. The combination of LR
pG and antibiotics improved the binding efficiency and ultimately enhanced the therapeutic efficacy. Therefore, LR
pG can serve as an additive to decrease the dose of antibiotics.
In this study, we also compared a natural AMP (melittin) to LR
pG in the GM
MIC, MHC
10, GM
SI, and stability results. Although it exhibited a slightly better antibacterial activity than LR
pG, melittin had a strong hemolysis effect. By calculating the GM
SI values (MHC
10/GM
MIC), we found that the GM
SI of LR
pG was 584 times higher than that of melittin. Furthermore, LR
pG exerted a good conditional stability and additive effects with conventional antibiotics. Based on the results above, LR
pG overcomes the main limiting factors of natural AMPs (such as melittin) and has the potential to develop into a therapeutic agent. Therefore, we further studied the antibacterial mechanism of LR
pG. Previous studies demonstrated that cationic AMPs combine with anions on the surface of bacteria via electrostatic action, and their hydrophobic amino acids are inserted into the bacterial cell membrane. Once the critical concentration is exceeded, the membrane breaks, and this ultimately leads to death of the bacteria [
21,
22]. LR
pG firstly bound to the LPS of the outer membrane of
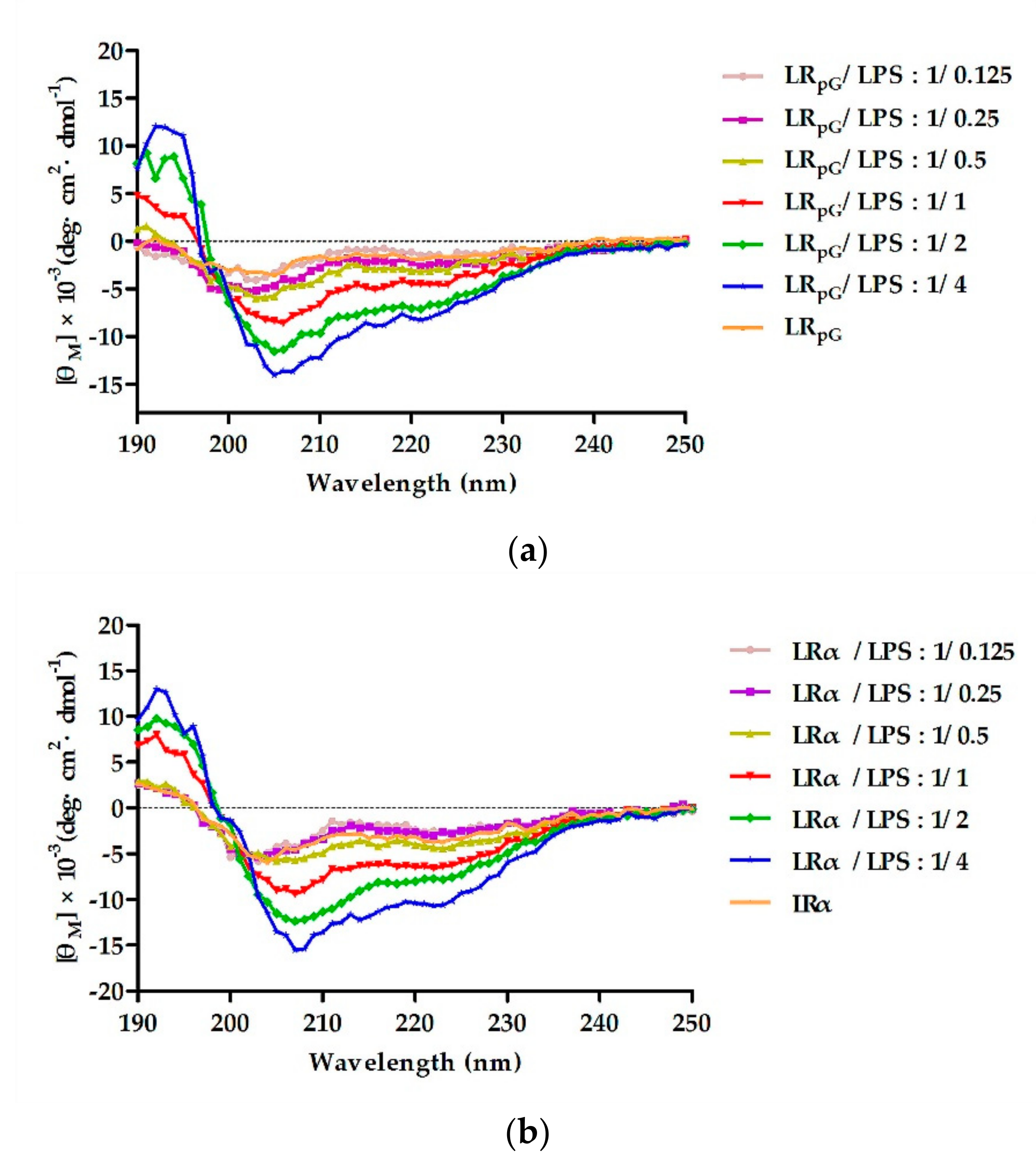
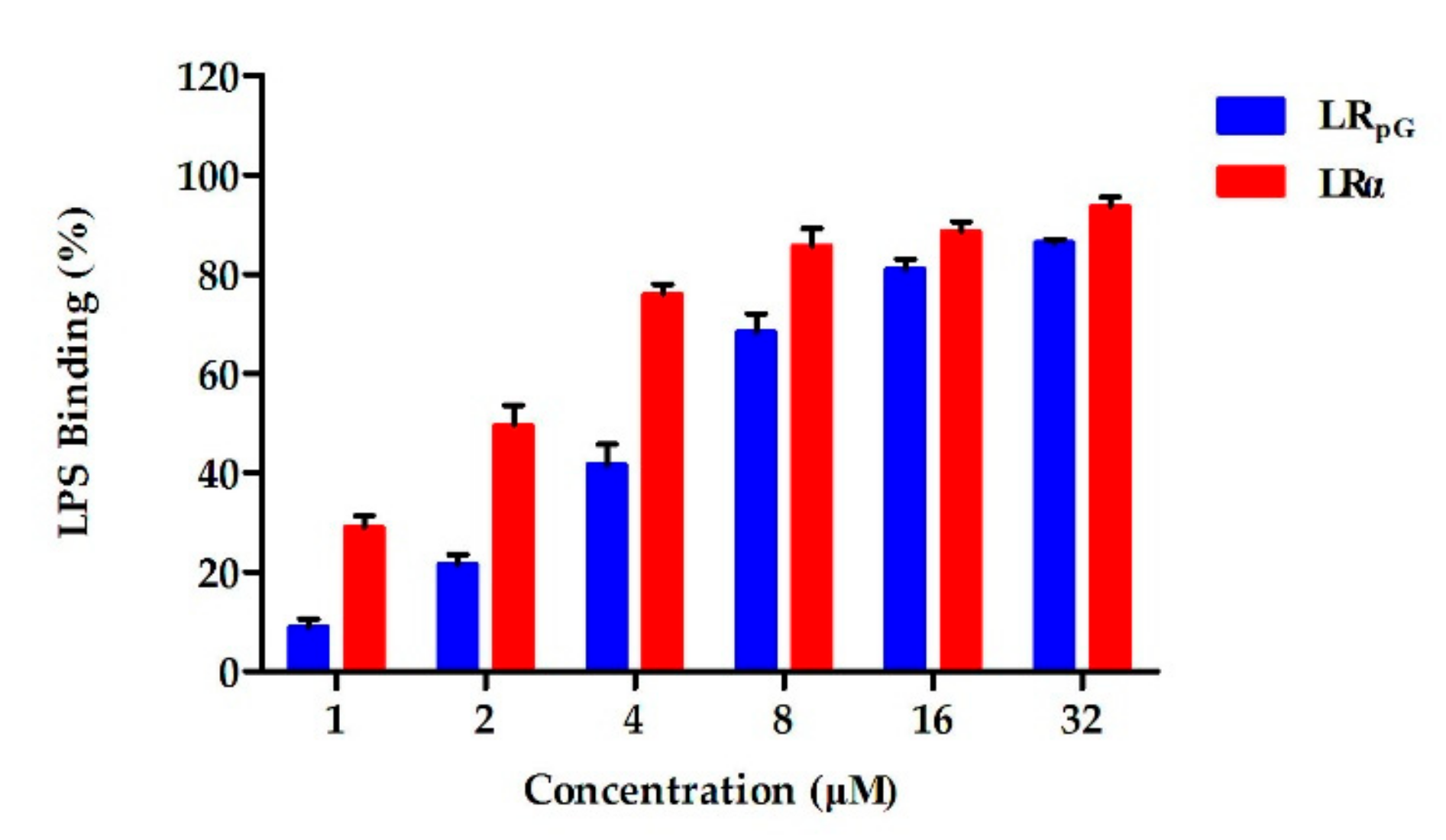
E. coli ATCC25922 through electrostatic action (
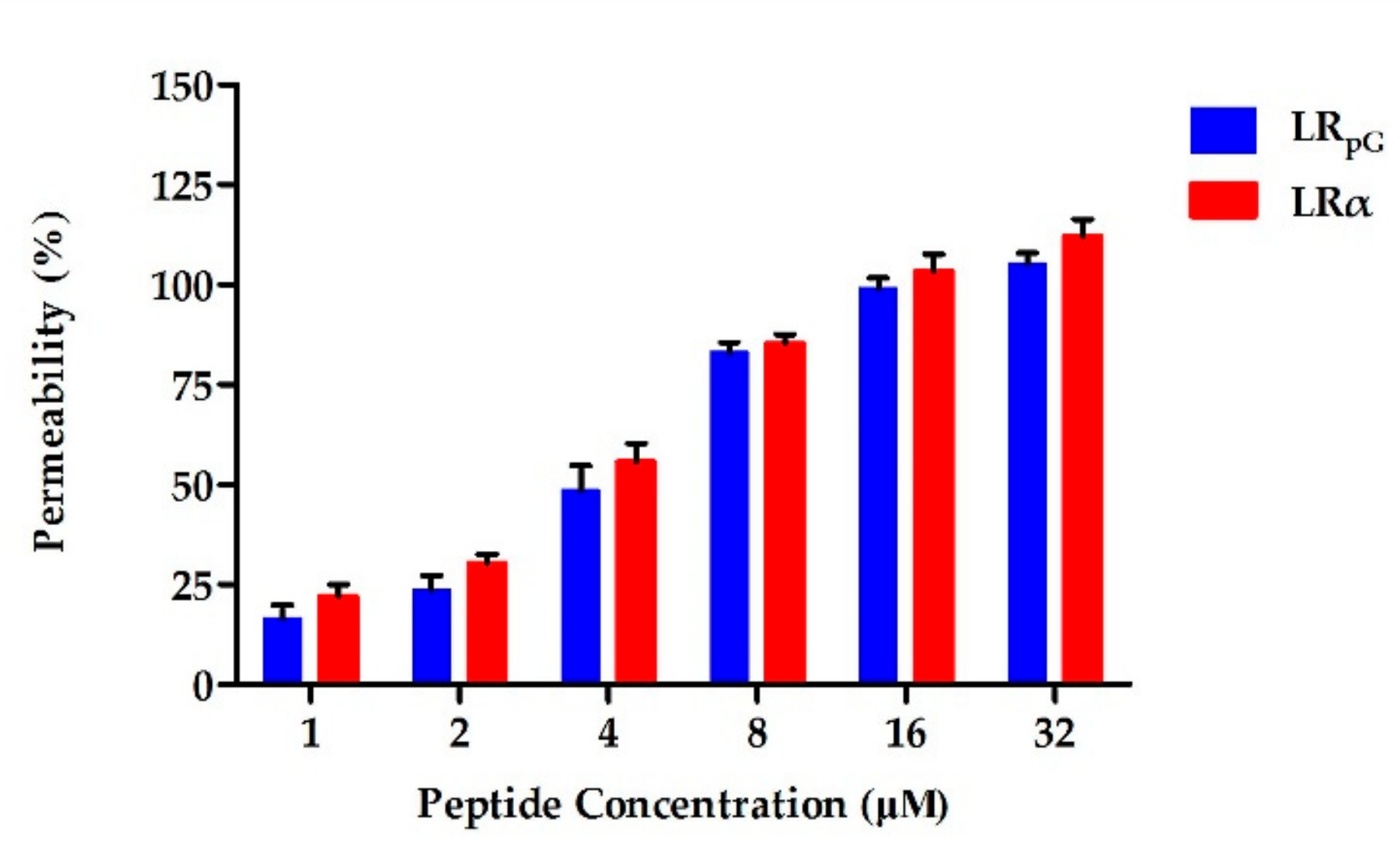
Figure 8), and then destroyed the cell membrane. Compared to the outer membrane, LR
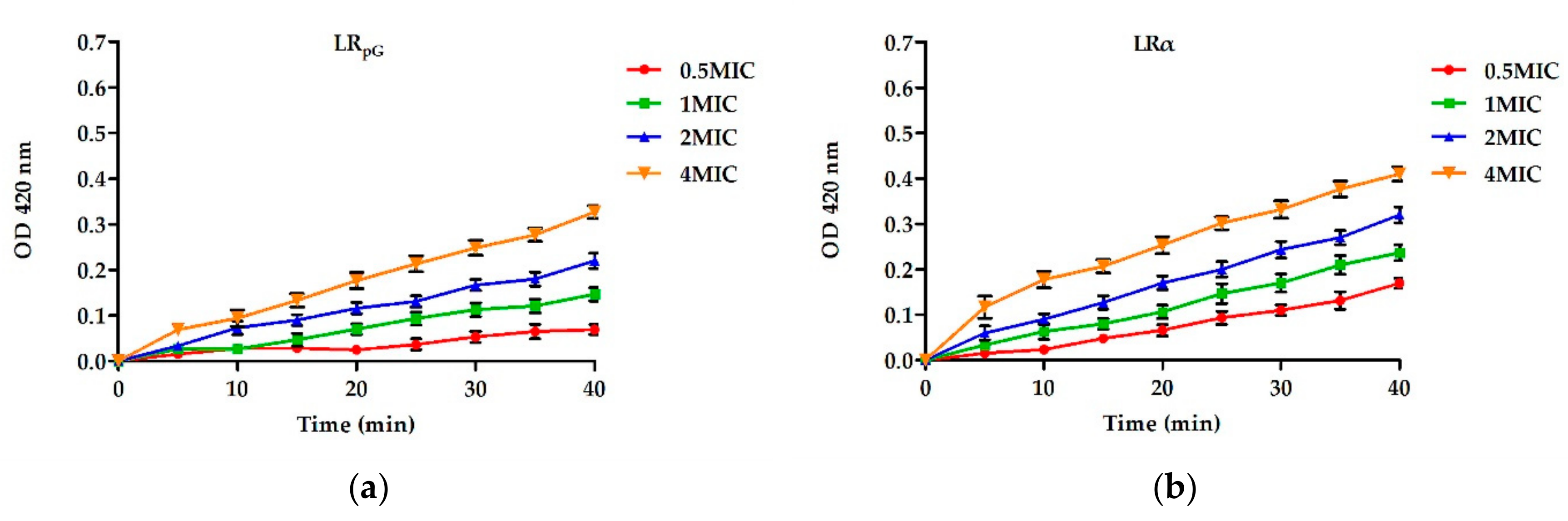
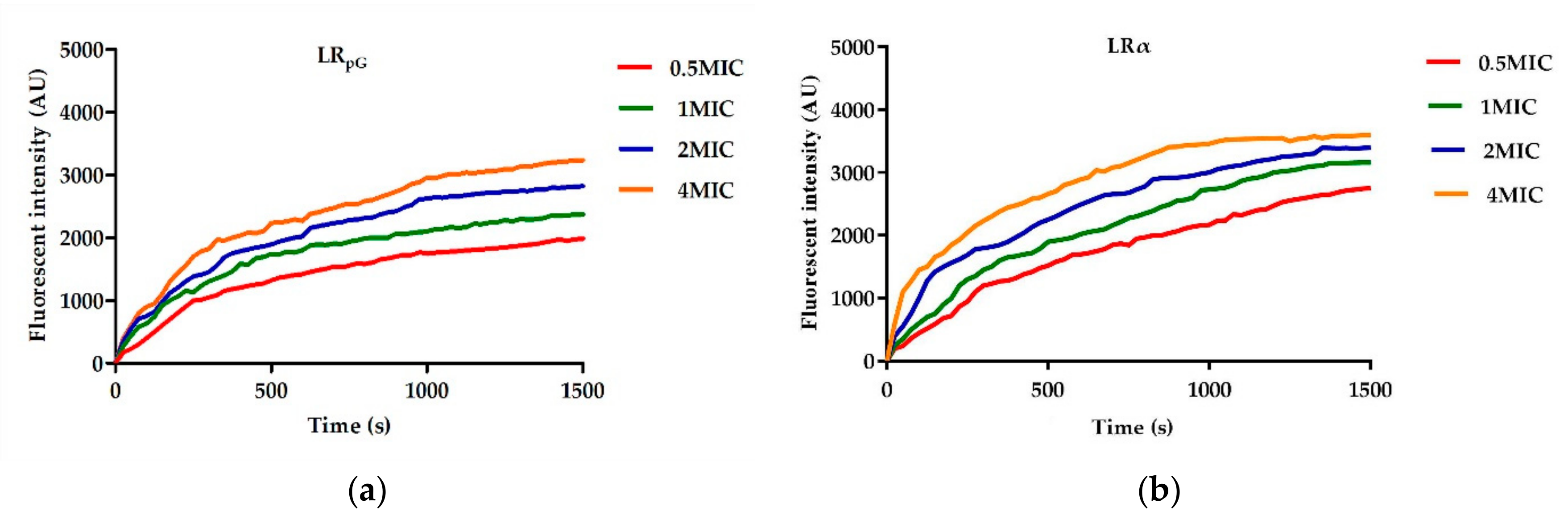
pG had a much weaker permeability to the inner membrane than LRα (
Figure 3 and
Figure 4). This result might be due to the decreased helical tendency of the LR
pG as a result of the loop (pG), which was consistent with the previous papers showing that the helical tendency of AMPs was closely related to the permeability of bacterial inner membrane [
20]. After breaking the impermeable barrier, LR
pG interacted with the cytoplasmic membrane, resulting in pore and ion channel formation (
Figure 5). SEM results directly confirmed that LR
pG killed
E. coli (
Figure 6) by destroying the bacterial cell membranes and causing intracellular lysate leakage. In addition to the membrane mechanisms mentioned above, LR
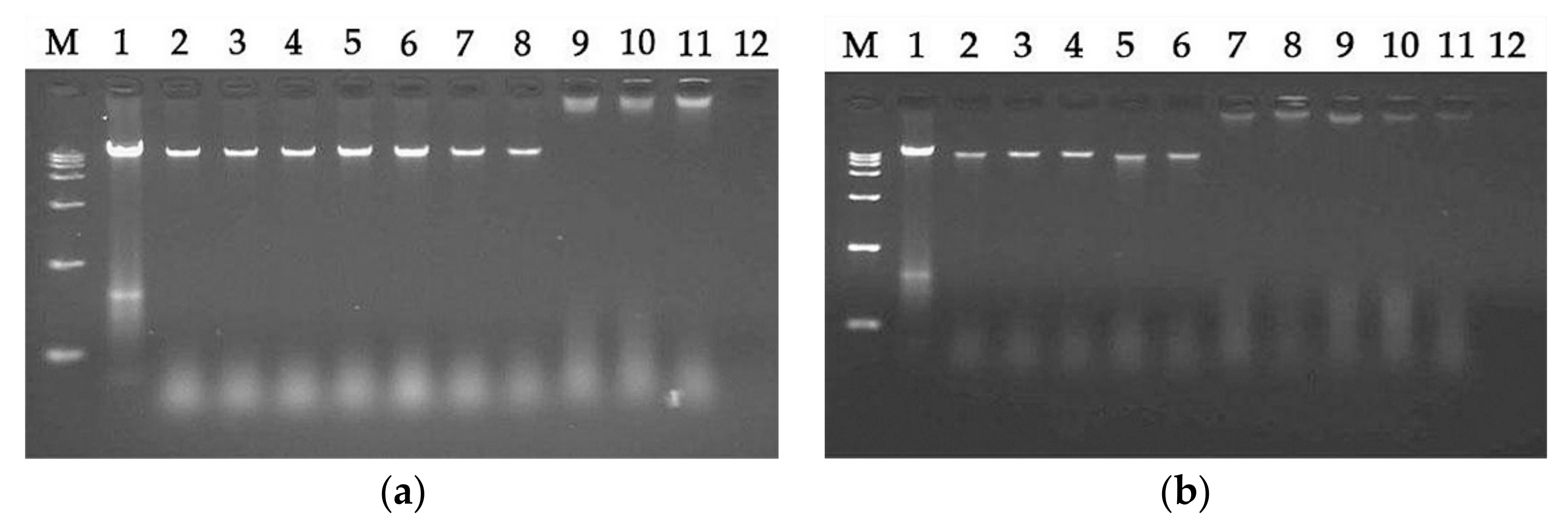
pG can bind to intracellular material DNA at a concentration much higher than the MIC (
Figure 7), suggesting that LR
pG cannot kill bacteria by DNA binding at the MIC. Furthermore, we found that the DNA binding ability of LR
pG was relatively weaker than that of LRα. One possible reason is that the α-helical forming ability of LR
pG was weaker than that of LRα. In short, LR
pG exerted its antibacterial effect by destroying the integrity of the bacterial cell membrane, causing leakage of the cellular content and binding to DNA through the cell membrane.
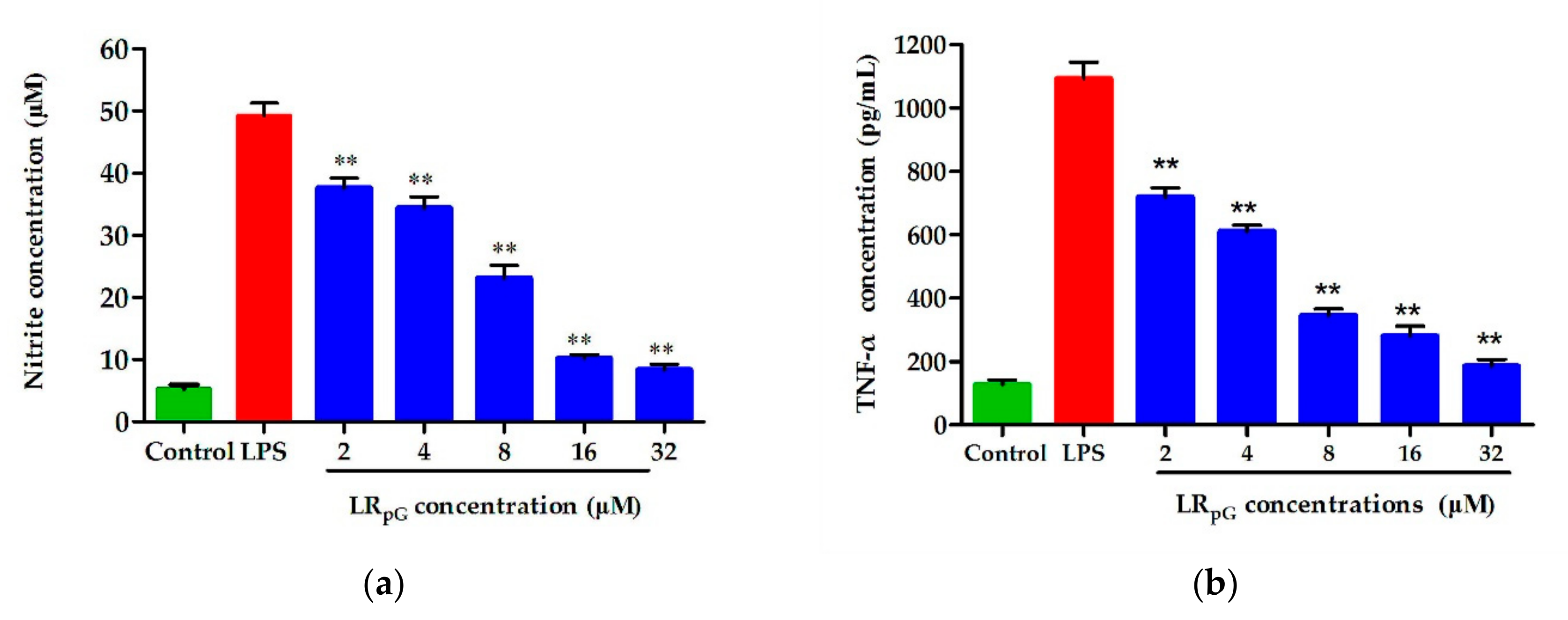
LPS is a crucial component of the outer membrane in Gram-negative bacteria [
36]. Following bacterial death under the action of antibacterial agents, a large amount of LPS detaches from the cells and enters into the blood circulation of the body, activates the inflammatory signaling pathway, and releases inflammatory mediators such as NO and TNF-α [
37,
38]. Previous studies showed that positively charged amphipathic AMPs had a strong LPS neutralization ability and were ideal candidates for anti-inflammatory drugs [
39,
40]. The LPS-induced inflammatory mediators NO and TNF-α were remarkably inhibited by LR
pG in a concentration-dependent manner (
Figure 10). Based on this study, LR
pG has the potential to develop into an anti-inflammatory agent by blocking LPS-mediated inflammatory mediators.
4. Materials and Methods
4.1. Materials
Escherichia coli (E. coli) ATCC25922, E. coli K88, Pseudomonas aeruginosa (P. aeruginosa) ATCC27853, Salmonella Pullorum (S. Pullorum) NCTC5776, Klebsiella pneumoniae (K. pneumoniae) CMCC46117, Staphylococcus aureus (S. aureus) ATCC25923, S. aureus ATCC29213, methicillin-resistant S. aureus ATCC43300, and Enterococcus faecalis (E. faecalis) ATCC29212 were obtained from the pharmacology and toxicology laboratory of the College of Animal Science and Technology, Jilin Agricultural University (Changchun, China). HEK293T and RAW 264.7 cells were obtained from the College of Animal Science and Technology, Jilin Agricultural University (Changchun, China). The hRBCs were obtained from the Jilin Agricultural University Hospital.
Mueller–Hinton broth (MHB) powder and Mueller–Hinton agar (MHA) were purchased from GL Biochem (Shanghai, China). Bovine serum albumin (BSA), Triton X-100, and phosphate-buffered saline (PBS) solution were purchased from Solarbio (Beijing, China). O-nitrophenyl-β-d-galactopyranoside (ONPG), LPS from E. coli 0111 B4, diSC3-5, polymyxin B, 4-(2-hydroxyethyl) piperazine-1-ethanesulfonic acid (HEPES), and NPN were all purchased from Sigma-Aldrich (Shanghai, China). TransDetect Cell Counting Kit-8 (CCK-8) and Dulbecco’s modified Eagle’s medium with high glucose (DMEM) were purchased from TransGen Biotech (Beijing, China). Fetal bovine serum (FBS) was purchased from Gibco (Shanghai, China).
4.2. Synthesis and Sequence Analysis of Peptides
The designed peptides were sent to GL Biochem (Shanghai, China) for synthesis and purification. The purity of the peptide was determined by reverse-phase high-performance liquid chromatography on an analytical Kromasil C
18 column (Beijing, China). The purity of the peptide was over 95% (RP-HPLC). ESI-MS analysis was used to measure the molecular weight of designed peptides, and values were very close to the calculated values. The primary physical and chemical parameters of the designed peptides were measured online at
http://web.expasy.org/compute_pi/. The hydrophobicity and relative hydrophobic moments were measured online at
http://heliquest.ipmc.cnrs.fr/.
4.3. CD Spectroscopy
The CD Spectroscopy of the designed peptides was detected on a J-810 spectropolarimeter (Jasco, Tokyo, Japan) at 25 °C. The final concentration of peptides in 10 mM PBS (pH 7.4), 50% TFE, and 30 mM SDS was 150 µM. The experimental parameters were as follows: 10 nm·min
−1 scanning rate, 1 mm path length, and 190–250 nm wavelength range. Each peptide was scanned an average of three times. The collected data were transformed to mean residue ellipticity as follows:
where θ
M represents the mean residue ellipticity (deg·cm
2·dmol
−1), θ
obs represents the ellipticity correction of that measurement buffer at a given wavelength (mdeg), l represents the path length (cm), c represents the peptide concentration (mM), and n represents the number of amino residues.
4.4. MIC Measurements
The antimicrobial activity of the designed peptides was examined using the micro-dilution method described previously [
29]. The bacteria were cultured for 16 h in MHB at 37 °C, and the microbial suspension was diluted to a final concentration of 10
5 colony-forming units (CFU)/mL. Then, 50 µL of MHB containing different concentrations peptides (0.25–256 µM) was mixed with 50 µL of bacterial solution, and the mixture was added to a 96-well plate. After incubation for 16 h, MIC was examined by measuring the OD at 492 nm (Microplate reader, TECAN GENios F129004, Tecan, Salzburg, Austria). The assays were performed three times.
4.5. Biocompatibility Assays
The hemolytic activity was examined based on the method mentioned previously [
22]. Briefly, the collected hRBCs were washed three times and diluted at 2% (
v/v) with PBS (pH 7.4). Then, 100 µL of hRBCs were put into each tube including 100 µL of two-fold diluted peptides. The mixtures were centrifuged at 1000×
g for 3 min. The OD value at 570 nm of the supernatant was measured, describing hemoglobin release (Microplate reader, TECAN GENios F129004, Tecan, Salzburg, Austria). For this assay, 1% Triton X-100-treated hRBCs were used as a positive control and untreated hRBCs were used as a negative control. MHC
10 was defined as the peptide concentration that caused 10% hemolysis. The assays were performed three times.
The cytotoxicity was detected according to a CCK-8 assay as described previously. Briefly, RAW 264.7 and HEK293T cells were placed into 96-well plates at a density of 1.0–2.0 × 105, and then cultured for 16 h under conditions of 5% CO2. Then, two-fold diluted peptides in DMEM were added to the plates. After 16 h of culturing, CCK-8 (10%, v/v) was put into each well containing cell culture, and mixtures were cultured for 2 h. The cytotoxicity assay was examined by measuring the OD of the mixtures at 450 nm (Microplate reader, TECAN GENios F129004, Tecan, Salzburg, Austria). The assays were performed three times.
4.6. Condition Sensitivity Assays
The salt, temperature, and pH sensitivities of designed peptides were determined by MIC assay as described previously [
40]. For the salt sensitivity assay, peptides were diluted in deionized water containing different salts, and mixed with
E. coli ATCC25922 (10
5 CFU/mL). The final physiological concentrations of different salts were as follows: 150 mM NaCl, 4.5 mM KCl, 6 μM NH
4Cl, 1 mM MgCl
2, 8 μM ZnCl
2, and 4 μM FeCl
3. For temperature sensitivity, peptides were cooled on ice for 10 min, and incubated at 37 °C and 100 °C for 30 min. For pH sensitivity, peptides were treated with different pH for 1 h. Briefly, peptides were diluted in deionized water, and the pH of the solution was adjusted to 4.0, 6.0, 8.0, and 10.0 with HCl or NaOH. The subsequent steps were the same as for the MIC measurements. These tests were performed three times.
4.7. Synergy with Conventional Antibiotics
The combined antibacterial effect of peptides with other antibiotics was evaluated by checkerboard assay as described previously [
31]. Briefly, peptides and antibiotics diluted two-fold were prepared at concentrations ranging from 1/8× MIC to 2× MIC. Subsequently, each peptide at the same concentration was added to the longitudinal column of wells, and each antibiotic at the same concentration was added to the horizontal row of wells. A suspension at 10
5 CFU/mL was put into each well for 16 h at 37 °C. Each assay was performed three times. The fractional inhibitory concentration (FIC) index was calculated as follows:
where FIC ≤ 0.5 denotes synergy, 0.5 < FIC ≤ 1.0 denotes additive, and 1.0 < FIC ≤ 4.0 denotes indifferent.
4.8. Outer Membrane Permeability Assay
The outer membrane permeability was examined using fluorescent dye NPN according to the method mentioned previously [
17,
21,
41]. Briefly,
E. coli ATCC25922 cells were washed three times and diluted to 10
5 CFU/mL in 5 mM HEPES buffer (pH 7.4, containing 5 mM glucose). Then, the final concentration of 10 μM NPN was added to the suspension, and incubated at room temperature in the dark for 30 min. An equal volume of suspension and peptides at various concentrations were mixed in a black 96-well plate. The fluorescence intensities of the samples were measured (emission λ = 420 nm, excitation λ = 350 nm) with an F4500 fluorescence spectrophotometer (Hitachi, Tokyo, Japan). Polymyxin B (10 µg/mL) with strong outer membrane permeability was used as a positive control. Values were converted to percentage NPN uptake using the following equation:
where F
obs is the observed fluorescence of NPN with
E. coli ATCC25922 cells at a given peptide concentration, F
0 is the initial fluorescence of NPN with
E. coli ATCC25922 cells, and F
100 is the fluorescence of NPN with
E. coli ATCC25922 cells with the addition of 10 µg/mL polymyxin B.
4.9. Inner Membrane Permeability Assay
The inner membrane permeabilization was determined by measuring the cytoplasm β-galactosidase activity according to the method mentioned previously [
21]. In short, an equal volume of the suspension of
E. coli ATCC25922 cells was washed three times and diluted to an OD
600 of 0.05 with 10 mM PBS (containing 1.5 mM ONPG); then, the suspension was put into each well of a 96-well plate and cultured with 0.5× MIC to 4× MIC peptides at 37 °C. The OD at 420 nm (Microplate reader, TECAN GENios F129004, Tecan, Salzburg, Austria) was recorded for 40 min every 5 min eight times. The method was used to detect the permeabilization of the inner membrane through the reflection of the ONPG influx into the cells.
4.10. Cytoplasmic Membrane Depolarization Assay
DiSC
3-5, a membrane potential-sensitive fluorescent dye, was used to assess the ability of the peptides to disrupt the cytoplasmic membrane as previously described, [
42]. Briefly,
E. coli ATCC25922 in the mid-log phase was washed three times and diluted to an OD
600 of 0.05 with 5 mM HEPES buffer (containing 20 mM glucose and 100 mM KCl); then, the mixture of
E. coli ATCC25922 and 0.5× MIC to 4× MIC peptides was mixed with diSC
3-5 (final concentration of 0.4 μM) at 37 °C for 1 h. The fluorescence changes from 0 to 1500 s were recorded at the emission wavelength (λ = 670 nm) and excitation wavelength (λ = 622 nm) (Fluorescence Spectrophotometer, F4500, Hitachi, Tokyo, Japan).
4.11. Scanning Electron Microscopy
E. coli ATCC25922 cells in the mid-log phase were washed three times and diluted to 107 CFU/mL in 10 mM PBS. The bacterial suspension and peptide (1× MIC) were added to 24-well plates containing polylysine-treated glass slides. The bacteria untreated by peptide were used as a control. After incubation, the bacterial suspension was removed, and the treatment method with polylysine-glass slides was repeated. Then, the polylysine-treated glass slides were fixed with glutaraldehyde (2.5%) at 4 °C for 12 h and dehydrated with a series of graded ethanol solutions for 20 min. The polylysine-treated glass slides were transferred into a mixture (v:v = 1:1) of pure alcohol and tertiary butanol for 15 min, and then placed into pure tertiary butanol for 15 min. Finally, the samples were treated using a critical point dryer with liquid CO2, and then scanned by a S-3400N SEM (Hitachi, Tokyo, Japan) after coating with gold-palladium.
4.12. DNA Binding Assay
The DNA binding assays were determined according to gel retardation experiments described previously [
17]. Briefly, 400 ng of genomic DNA (
E. coli ATCC25922) was mixed with 0.5 to 256 µM peptide in a binding buffer (50 µg/mL BSA, 1 mM ethylene diamine tetraacetic acid (EDTA), 20 mM KCl, 10 mM Tris-HCl (pH = 8.0), 5% glycerol, and 1 mM dithiothreitol) for 60 min at 37 °C. Subsequently, the samples were examined by 1% agarose gel electrophoresis.
4.13. LPS Binding Assay
The ability of peptides to bind LPS was detected on a J-810 spectropolarimeter at 25 °C. Briefly, concentration of LPS ranged from 37.5–600 µM, diluted using the double dilution method. The final concentration of peptides in the LPS solution was 150 µM. The experimental parameters were the same as in
Section 4.3.
4.14. LAL Assay
Peptide neutralization was evaluated using a quantitative chromogenic limulus amoebocyte lysate (LAL) assay (Xiamen, China) [
43]. Briefly, 1 endotoxin unit (EU) of LPS was incubated with two-fold serial dilutions of peptides at 37 °C for 60 min; then, 100 µL of LAL reagent was added to the mixtures. After additional incubation for 15 min at 37 °C, 100 µL of chromogenic substrate solution was put into the tubes. Subsequently, the mixtures were incubated for another 10 min at 37 °C before adding 500 µL of azo reagent solutions (1, 2, and 3) in turn. The data were examined by measuring the OD value at 545 nm (Microplate reader, TECAN GENios F129004, Tecan, Salzburg, Austria).
4.15. Endotoxin Neutralization Assay
RAW 264.7 cells were put into 24-well plates at a density of 1.0–2.0 × 105 cells/well and stimulated with LPS (100 ng/mL) with (1–64 μM) or without peptide. After incubating for 24 h at 37 °C in 5% CO2, the supernatants were collected for analysis of the NO production using Griess reagent and TNF-α levels using ELISA based on the manufacturer’s protocol. The cells treated with LPS only and untreated cells were set as the positive and negative controls, respectively.
4.16. Statistical Analysis
ANOVA was used to analyze the data through the SPSS 18.0 software (IBM, Chicago, IL, USA). Quantitative data were expressed as the mean ± standard deviation. A p-value < 0.01 was considered to have statistical significance.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}