Extracellular Vesicles Mediated Early Embryo–Maternal Interactions



Department of Veterinary Medicine, Faculty of Animal Sciences and Food Engineering, University of São Paulo, 13635-900 Pirassununga, SP, Brazil
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2020, 21(3), 1163; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031163
Submission received: 13 December 2019
/
Revised: 31 December 2019
/
Accepted: 6 January 2020
/
Published: 10 February 2020
(This article belongs to the Special Issue Embryo-Maternal Interactions Underlying Reproduction in Mammals)
Abstract
:Embryo–maternal crosstalk is an important event that involves many biological processes, which must occur perfectly for pregnancy success. This complex communication starts from the zygote stage within the oviduct and continues in the uterus up to the end of pregnancy. Small extracellular vesicles (EVs) are part of this communication and carry bioactive molecules such as proteins, lipids, mRNA, and miRNA. Small EVs are present in the oviductal and uterine fluid and have important functions during fertilization and early embryonic development. Embryonic cells are able to uptake oviductal and endometrium-derived small EVs. Conversely, embryo-derived EVs might modulate oviductal and uterine function. In this review, our aim is to demonstrate the role of extracellular vesicles modulating embryo–maternal interactions during early pregnancy.
1. Introduction
In mammals, the perfect embryo–maternal communication is necessary to allow the establishment and maintenance of pregnancy. For this, oocyte maturation occurs in the preovulatory follicle, followed by the fertilization in the oviduct, and early development in the oviduct and uterus during the luteal phase [1]. Upon ovulation, the oocyte starts its journey through the oviduct, is fertilized, and embryo development begins, followed by its first cleavages [2]. Importantly, the major embryonic gene activation (EGA) occurs during the embryo passage through the oviduct [3]. During this phase, the embryo starts to transcribe more actively, decreasing its dependency on the maternal mRNA stock. The epithelial line of the oviduct is formed by ciliated and secretory cells. These cells play roles in gamete transport, capacitation and fertilization [4,5], and on early embryonic development [6,7], mostly by oviductal secretions [8]. Small extracellular vesicles (EVs) are present in oviductal fluid content and participate in these important reproductive events [5,7].
The embryo enters the uterus 4 to 5 days after oocyte fertilization, in bovine [2,9]. During embryonic development, the embryo reaches the blastocyst stage composed of two cell lines: the inner cell mass that will originate the embryo proper and the trophectoderm cell monolayer, which ultimately will form the fetal adnexa/placenta [2,10]. Fetal appendages will establish contact with the endometrium, and thus establish the interchange interface between embryo/fetus and maternal tissues. However, in order to allow embryo development and placentation, uterine epithelial and glandular cells secrete the uterine fluid that is very important for embryo nutrition because it contains proteins, lipids, amino acids, growth factors [11], and small EVs [12,13,14].
In most domestic mammalian species, the embryo—when in the uterus—increases the secretion of biological molecules related to the maternal recognition of pregnancy (MRP). These molecules have luteotrophic and anti-luteolytic actions, guaranteeing the corpus luteum (CL)’s capability to secrete the progesterone necessary to establish and sustain pregnancy [15,16]. Additionally, different biological molecules are secreted by the embryos of several mammalian species during MRP as interferon-tau (IFNT) in ruminant [17], estradiol in pigs [18], and chorionic gonadotropin in humans [19]. However, recent reports indicate that the embryo is also able to secrete EVs [20,21,22], but their role in MRP is still elusive.
Therefore, in mammals, the embryos and the female tract (oviduct and endometrium) are able to secrete extracellular vesicles during the pre-implantation period. Extracellular vesicles are cell-secreted vesicles that are classified according the their size, biogenesis, and secretion, as exosomes, microvesicles, and apoptotic bodies [23]. Several cellular types can secrete EVs including follicular [24,25], oviductal, and endometrial cells [13,21,26], as well as in vitro and in vivo produced embryos [20,21,22,27]. Extracellular vesicles carry bioactive molecule as miRNAs, mRNAs [28], proteins [29], and lipids [30], which modulate various reproductive processes such as ovarian follicular development, oocyte maturation, embryonic development, maternal–embryonic communication, and the establishment of mammalian gestation. There is experimental evidence that EVs participate in intercellular communication in essential reproductive events related to the establishment of pregnancy, such as cell proliferation, crosstalk between the maternal organism and conceptus, as well as during embryonic implantation [31]. In early stages of pregnancy, the communication between the conceptus and maternal organism is necessary and the evidence of the participation of EVs of maternal or embryonic origin is increasing [14].
Herein, we will review the current knowledge on embryo–maternal interactions, with special emphasis on the roles of EVs during the early crosstalk between embryonic and maternal tissues.
2. Embryo–Maternal Interactions Mediated by Embryotropins
Embryotropins are bioactive molecules such as proteins, lipids, and miRNAs secreted from both in vivo—or in vitro—produced mammalian embryos [32]. These molecules may act in autocrine and/or paracrine ways, modulating the embryo’s development (in vitro culture) and the maternal endometrial cells, respectively [32].
In domestic ruminants, the mostly known and well-characterized embryotropin is the IFNT. IFNT is a cytokine secreted by the trophoblastic cells of the ruminant embryo and conceptus that can act in a paracrine and endocrine way. In the uterus, IFNT acts in a paracrine way, decreasing estrogen and oxytocin receptor expression in the endometrium, which is an essential step to maintaining a viable corpus luteum and producing progesterone [33]. Besides antiluteolytic function, IFNT has endocrine effects, stimulating the expression of IFN-stimulated genes (ISGs) in the endometrium [34], in the corpus luteum [35,36,37], in white blood cells [38], and in the liver [39]. The effects of IFNT secreted by the bovine embryo in the maternal organism can be detected as early as day 7 of development [40,41].
The IFNT function on the maternal recognition of pregnancy is well established for ruminants. There are parallels for such a response in other species. One example is the conserved response of interferon-stimulated gene 15 (ISG15), which is stimulated by IFNs and other cytokines. ISG15 is up-regulated in the endometrium of ruminants [34], primates [42], and mice [43] during early pregnancy. However, its function during the maternal recognition of pregnancy in non-ruminant species is not well established. Mouse knockout for Isg15 results in 50% fetal loss, which can be explained by change decidual gene expression that is functionally related to cell survival and adhesion pathways [44].
Besides IFNT, day 13 bovine embryos can also secrete prostaglandins, such as prostaglandin F2 alpha (PGF), prostaglandin E2 (PGE2), and prostaglandin I2 (PGI2) [45]. These prostaglandins act in a paracrine way in the endometrium, increasing ISGs’ expression and function, which can be important for uterine receptivity as well as conceptus growth and development during early pregnancy [45]. In addition, PGE2 and PGI2 can modulate blastocyst implantation, decidualization, and endometrial vascular permeability during early pregnancy in mice and rats [46]. In large domestic species, PGE2 is secreted by the endometrium and embryo, showing an important role as the local antiluteolytic factor (reviewed by [47]). PGE2 secreted by equine viable embryos during early embryonic development is involved with the initial oviductal transport of the embryo and opening the uterutubal papilla to allow the embryo to enter the uterus. This mechanism may explain why unfertilized oocytes or degenerate embryos are not able to gain the uterine lumen in equine [48,49]. Additionally, the lysophosphatidic acid (LPA) is secreted by the bovine embryo and endometrium and may act as a luteotropic factor by stimulating PGE2 synthesis in stromal cells during early pregnancy [50].
Hence, embryotropins are signaling molecules acting in the maternal tissues to improve embryo–maternal recognition in mammals. Several embryotropins are secreted by different species with the same aim, which is to inform embryo presence to the mother and to maintain a functional CL, progesterone synthesis, and the maintenance of pregnancy. However, other mechanisms can be involved in the maternal recognition of pregnancy. Recently, EVs were introduced as new players in the embryo–maternal communication.
3. Extracellular Vesicles Derived from Female Reproductive Tract and Embryo
Extracellular vesicles (EVs) are formed by a phospholipid bilayer and secreted by different cell types [51]. EVs were discovered approximately 40 years ago, and they were first characterized as “small trash bags”, due to the possibility of carrying cellular material to the extracellular environment [52]. Currently, these EVs have been found in different body fluids and can be divided into exosomes, microvesicles, and apoptotic bodies [23] according to their size, biosynthesis, and contents.
Exosomes are nanovesicles between 30 and 150 nm in size [53] that carry bioactive material such as lipids [30], proteins [29], RNAs, and miRNAs [28], which are able to control regulatory pathways associated with physiological or pathological functions among neighboring or distant cells through the extracellular environment [13]. The biogenesis of small EVs initiates with plasma membrane endocytosis, which gives rise to multivesicular bodies (MVBs). Inside MVBs intraluminal vesicles are formed, which are released out of the cell when the MVB fuses with the cell plasma membrane [54]. The released vesicles are called exosomes (small extracellular vesicles).
Cellular communication mediated by small EVs can happen by fusion between the vesicle membrane and plasma membrane of the target cell, leading to the release of EVs contents into the target cell cytoplasm [13]. In addition to cell fusion, it is suggested that specific receptors present in the membrane of the target cells are involved in EVs’ endocytosis [55,56].
Small EVs have also been identified in many body fluids as follicular fluid [24], blood [57], cerebrospinal fluid [58], urine [59], oviduct [7], and uterine fluids [13,21]. In addition to body fluids, embryos cultured in vitro can secrete small EVs in the culture medium [20]. There are different methods to isolate EVs from body fluids and culture medium such as ultracentrifugation, filtration, size-exclusion chromatography, polymer precipitation, immunoaffinity, and microfluidic techniques [60]. Thus, based on the research and type sample, the best isolation method can be chosen [60]. This decision is normally based on the molecule of interest since each method can yield different concentrations and purity. For example, our laboratory has used filtration and ultracentrifugation to isolate small EVs from different body fluids such as follicular fluid [25,61], polymer precipitation, and ultracentrifugation to separate small EVs from an in vitro embryo-conditioned medium [62]. Similarly, other studies have isolated EVs from embryos or conceptus-conditioned culture media using polymer-based precipitation [21] and ultracentrifugation [20,63].
Small EVs are involved in several reproductive events related to oocyte maturation, fertilization, early embryonic development, and crosstalk between embryo and maternal organism (Table 1). One of the first studies that identified the presence of EVs associated with reproductive events used equine ovarian follicular fluid to investigate the role of extracellular vesicles during follicular development. The authors demonstrated that EVs from equine ovarian follicular fluid carried miRNAs and proteins, and were uptaken by granulosa cells in vitro and in vivo, suggesting a new cell-to-cell communication inside the ovarian follicles [24]. Another study demonstrated that oviductal microvesicles were uptaken by oocyte cytoplasm after 72 h of maturation in bitch, demonstrating the role of EVs in improving the oocyte maturation rate [64]. Furthermore, another study using murine oviductal EVs demonstrated that EVs carrying plasma membrane Ca2+- ATPase 4 (PMCA4) were uptaken by sperm, thus inducing sperm capacitation prior to fertilization [5].
4. Embryo–Maternal Interactions through Oviductal EVs
Oviduct used to be considered just a tubular connection between the ovary and the uterus where the oocyte and sperm passed through [73]. However, the oviduct is composed by ciliated and secretory cells that secrete oviductal fluid [8]. Several studies demonstrated the important biological role of the oviductal fluid during sperm capacitation [74], fertilization [4], and the outset of embryonic development [6]. Additionally, oviductal fluid contents include extracellular vesicles that have an important role during oocyte fertilization [5] and early embryonic development [7].
Recently, the functional effects of the EVs derived from the oviduct on gametes and embryos were summarized by Almiñana and Bauersachs (2019) [75]. In this review, we highlight the key findings related to EVs derived from oviductal cells and their effects in early embryonic development.
EVs from bovine oviduct epithelial cells (BOEC) were used in in vitro embryo production and demonstrated to improve embryo quality based on the increase in the number in trophectoderm and total cells and survival after vitrification [7]. In addition, space-specific EVs secreted from isthmus oviductal fluid were able to increase the survival rate and improve the development as well as the quality of in vitro produced blastocysts [67]. In vitro embryos were able to uptake EVs derived from in vivo oviduct epithelial cells, and this communication benefits the embryo blastocyst rate, survival, and quality [66]. Moreover, EVs secreted by donor oviductal cells increase birth rates after embryo transfer in mice due to decreased apoptosis and improved cellular differentiation in embryos [68]. Altogether, these data show the importance of embryo–maternal interactions mediated by EVs derived from the oviduct during early embryonic development, leading to improved embryo quality and successful pregnancy.
An important problem that may occur during the passage of the zygote through the oviduct is ectopic pregnancy (EP), which occurs when the embryo after fertilization implants outside of the uterine cavity due to structural abnormalities in the fallopian tubes, for example [76,77]. Approximately 1.5%–2% of all the pregnancies are ectopic [78]; 97% are in the fallopian tube (oviduct) (reviewed by [79]). Currently, two diagnosis methods are used to detect EP: measurements of human chorionic gonadotropin(hCG)and progesterone in the serum [77]. The serial serum hCG measurement with intervals of 48 h is needed for the diagnostic; however, during this timecourse, tubal rupture might occur in patients, leading to possible complications in clinical status [77,80]. Therefore, only these two biomarkers are not precise and efficient to detect EP. Recently, new approaches showed that circulating miRNA miR-323-3p can be associated with serum hCG and progesterone to improve EP diagnostic [81]. Thus, the presence of miR-323-3p in serum could serve as a marker for EP. However, more studies are necessary to demonstrate if this miRNA is carried by small EVs or not, in order to become a reliable diagnostic marker.
5. Embryo–Maternal Interactions between Uterus and Embryo Mediated by EVs
The mammalian uterus is designed to allow sperm transport [82] as well as embryonic and fetal development [12]. In bovine, the morula stage embryo will arrive at the uterus at the uterotubal junction portion around Day 5 of embryo development [2]. Uterine fluid, termed histotroph in ruminants, is the result of glandular cells secretion inside of the uterine lumen [11,83]. This fluid is very important for the nutrition of the embryo since it contains proteins, lipids, amino acids, growth factors [11], and recently described extracellular vesicles that carry bioactive substances that are important for the early embryonic development [12,13,14]. During early embryo development, intense crosstalk starts between the embryo and the maternal uterine environment. This communication is necessary to induce the maternal recognition of pregnancy; thus, it is important to understand the role of extracellular vesicles in the embryo–maternal interface.
During maternal recognition of the pregnancy period, EVs were isolated within the uterine flushing of ewes on day 14 of the estrus cycle were fluorescently labeled with PKH67 and observed inside conceptus trophectoderm cells. This finding demonstrates that EVs are involved in paracrine communication between the endometrium and conceptus during the early pregnancy period [21]. In addition to that, sheep endometrial epithelium can secrete exosomes containing ovine endogenous jaagsiekte retroviruses (enJSRV) mRNA, which acts on trophectoderm via toll-like receptors (TLR) to induce IFNT production [69]. Trophoblast cells from the conceptus at day 15 and 17 secrete EVs containing IFNT that are able to stimulate ISGs’ expression in endometrial cell culture [22,70]. Furthermore, macrophage-capping protein (CAPG) and aldo-keto reductase family 1, member B1 protein (AKR1B1) proteins are present in EVs isolated from the uterine flushing of pregnant cows on days 15 and 17 of gestation [22]. Besides that, EVs isolated from uterine flushing in the pre-implantation period increase the expression of apoptotic genes (BAX, CASP3, TNFA, and TP53 transcripts) in endometrial cells [70]. In addition to that, endometrial cells treated with EVs from the post-implantation induced and increase in vascular cell adhesion molecule 1 (VCAM) transcript, indicating the modulation of adhesion molecules [70]. Furthermore, exosomes isolated from uterine flushing obtained from pregnant cows on days 17, 20, and 22 were used to treat trophoblast CT-1 cells and did not induce changes in IFNT and CDX2 mRNA expression, suggesting that the pregnancy period may influence EVs’ contents [70]. Together, this information highlights the EVs biological role during the period of maternal recognition of pregnancy, which may enhance embryo–maternal communication and consequently contribute to the maintenance of pregnancy.
Steroid hormones, such as progesterone and estradiol, can induce changes in the EVs secretion in human endometrial cells [26]. Progesterone, which is secreted by corpus luteum, is necessary to the establishment and maintenance of pregnancy and acts in the elongation and survival of the conceptus [84]. Progesterone induces myometrium relaxation and stimulates the production of mucin 1 (MUC-1), which is a protein that prevents conceptus adhesion to endometrium; thus, it can continue elongating and producing IFNT [85] as well as stimulating histotroph production by endometrial glands [11]. On day 10 to 14 of the estrous cycle in ovine, an increase in EVs secretion by endometrial luminal cells occurs, suggesting that progesterone is responsible for this event [71]. Besides that, EVs from the uterine lumen had miRNAs upregulated by progesterone that were predicted to modulate phosphoinositide 3-kinase/ Serine/threonine kinase 1 (PI3K/AKT), bone morphogenetic protein (BMP), and post-transcriptional silencing by small RNA pathways [71]. These results reinforce that progesterone is very important during the onset of pregnancy because it can modulate the endometrial function and consequently contribute to embryo development.
Extracellular vesicles are also secreted by endometrium and chorioallantoic membrane cells as well as trophectoderm and maternal endothelial cells from sows on day 20 of pregnancy [72]. MiRNAs and proteins within EVs were able to modulate the angiogenesis pathway within trophectoderm and maternal endothelial cells [72]. Moreover, EVs derived from the porcine trophectoderm are uptaken by maternal endothelial cells and stimulate the cellular proliferation of these cells [72]. Together, these results demonstrate that EVs have an important biological role in conceptus–endometrium crosstalk during the establishment of pregnancy in porcine.
Local vascularization between the uterine horn ipsilateral and the corpus luteum is more prominent during the estrous cycle luteal phase than the contralateral phase, suggesting that the oviduct and uterine could signal to CL and adjacent tissue for future pregnancy and/or luteal vascularization maintenance [86]. This elevated vascularization on the ipsilateral horn could be involved with the early onset of pregnancy recognition. As an example, exosomal miRNAs were identified in serum samples of nonpregnant and pregnant mares on days 9, 11, or 13 postovulation [87]. These miRNAs were increased in nonpregnant mares and predicted to target the pathway of focal adhesion molecules (FAMs) in the endometrium [87], which are involved in the regulation of the extracellular matrix [88]. These data suggest that in pregnant mares, FAMs are normally abundant, which suggests that exosomal miRNAs are less necessary to modulate focal adhesion pathway mRNAs in the endometrium, allowing the embryo to move inside the uterus, which contributes to the maternal recognition of pregnancy [87].
In bovine, 27 miRNAs were highly abundant in the serum small EVs of cows with embryonic mortality compared to the pregnant group on day 17 [89]. These miRNAs modulate pathways associated with many important processes such as inflammation, cell proliferation, endometriosis, cell cycle progression, contraction, infection, late-onset preeclampsia, apoptosis, differentiation, uterine leiomyoma, ovarian endometriosis, and cell viability [89]. However, in a retrospective study examining EVs isolated from the blood plasma of pregnant cows on day 21 of gestation, a low abundance of 27 miRNAs was identified in samples from initial somatic cell nuclear transfer (SCNT) embryonic loss compared with full-term SCNT pregnancies and full-term artificial insemination pregnancies. These miRNAs modulate the pathways associated with pregnancy establishment as well as cell proliferation, differentiation, apoptosis, angiogenesis, and uterus embryonic development [90]. In addition, 29 miRNAs from serum small EVs were differently detected in the 30 days of pregnancy group compared to the normal group [91]. Different pathways involved in metabolism are modulated by these 21 up-regulated miRNAs and eight down-regulated miRNAs in pregnant cows [91].
In Table 2, we itemized the publications about EVs carrying miRNAs in the pregnant female blood. On day 9, in pregnant mares, miRNA-29c and miRNA-101 are down-regulated in circulation exosomes [87]; however, on day 21, in bovine, these miRNAs are more abundant [90]. On day 17, in bovine, miRNA-15a, miRNA-15b, miRNA-101, miRNA-106b, miRNA-652, miRNA-143, and miRNA-148a presented decreased expression [89]; however, on day 21, miRNA-15a, miRNA-15b, miRNA-101, miRNA-106b, and miRNA-652 [90], and on day 30, miRNA-143 and miRNA-148a [91] expression increased in the serum-derived small EVs of pregnant cows. MiRNA-148b is increased in blood exosomes on days 21 and 30 of bovine pregnancy [90,91]. However, on day 21, miRNA-193b is more abundant, but on day 30, miRNA-193b is down-regulated in pregnant cows [90,91].
These data suggest that there is EV-mediated communication between the uterus and peripheral circulation. This communication can be direct and modulated by biological factors secreted by the embryo, or it can be indirect, where the embryo stimulates an endometrium response. Thus, extracellular vesicles can be part of this intricate mechanism improving embryo–maternal interactions and consequently pregnancy success in mammals. Moreover, these studies demonstrate the potential role of circulating exosomal miRNAs as biomarkers in early embryonic mortality or early pregnancy diagnosis.
In conclusion, small EVs carry important bioactive molecules that are involved in embryo–maternal crosstalk (Figure 1) through the modulation of important signaling pathways such as angiogenesis, apoptosis, interferon-tau, adhesion, proliferation, and cell survival. The above-mentioned studies demonstrate that EVs participate in embryo–maternal interactions during early embryonic development and the maternal recognition of pregnancy in mammals.
6. Conclusions
In this review, we demonstrate the biological roles of extracellular vesicles in events occurring during the onset of pregnancy and involved in the communication between the embryo and the maternal organism in different mammalian species. EVs carry important bioactive molecules that are capable of modulating key reproductive events during the early pregnancy period. Further investigations are necessary to elucidate if EVs secreted by the oviduct and endometrium as well as embryos can arrive in peripheral circulation and modulate different pathways in maternal organisms. Thus, the progression in our understanding related to this type of communication can advance the tests to detect pregnancies, abnormal pregnancies (EP), and predict pregnancy loss, as well as push the development of new technologies to modulate early embryo–maternal interactions.
Author Contributions
A.B., F.P., and J.C.d.S. revised the literature and prepared the manuscript draft. A.B. prepared the figure and tables. F.P. and J.C.d.S. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by São Paulo Research Foundation (FAPESP) grants number 2014/22887-0 (J.C.S.), 2018/13155-6 (F.P.) and 2017/19681-9 (A.B.).
Acknowledgments
The authors would like to thank the University of São Paulo, São Paulo Research Foundation (FAPESP grants 2014/22887-0 (J.C.S.), 2018/13155-6 (F.P.) and 2017/19681-9 (A.B.)), the National Council for Scientific and Technological Development–CNPq (grant number #306349/2017-5 (FP), #420152/2018-0 (J.C.S.)) and CAPES–Finance Code 001.
Conflicts of Interest
The authors declared no conflict of interest.
References
- Holm, P.; Callesen, H. In vivo versus in vitro produced bovine ova: Similarities and differences relevant for practical application. Reprod. Nutr. Dev. 1998, 38, 579–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betteridge, K.J.; Fléchon, J.E. The anatomy and physiology of pre-attachment bovine embryos. Theriogenology 1988, 29, 155–187. [Google Scholar] [CrossRef]
- Memili, E.; Dominko, T.; First, N.L. Onset of transcription in bovine oocytes and preimplantation embryos. Mol. Reprod. Dev. 1998, 51, 36–41. [Google Scholar] [CrossRef]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Dossary, A.A.; Strehler, E.E.; Martin-DeLeon, P.A. Expression and Secretion of Plasma Membrane Ca2+-ATPase 4a (PMCA4a) during Murine Estrus: Association with Oviductal Exosomes and Uptake in Sperm. PLoS ONE 2013, 8, e80181. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.M.; Wang, H.S.; Lee, C.L.; Lee, J.D.; Huang, H.Y.; Chang, F.H.; Lee, J.F.; Soong, Y.K. Insulin-like growth factor-binding proteins produced by Vero cells, human oviductal cells and human endometrial cells, and the role of insulin-like growth factor-binding protein-3 in mouse embryo co-culture systems. Hum. Reprod. 1996, 11, 1281–1286. [Google Scholar] [CrossRef] [Green Version]
- Lopera-Vásquez, R.; Hamdi, M.; Fernandez-Fuertes, B.; Maillo, V.; Beltrán-Breña, P.; Calle, A.; Redruello, A.; López-Martín, S.; Gutierrez-Adán, A.; Yañez-Mó, M.; et al. Extracellular Vesicles from BOEC in In Vitro Embryo Development and Quality. PLoS ONE 2016, 11, e0148083. [Google Scholar] [CrossRef]
- Leese, H.J.; Hugentobler, S.A.; Gray, S.M.; Morris, D.G.; Sturmey, R.G.; Whitear, S.-L.; Sreenan, J.M. Female reproductive tract fluids: Composition, mechanism of formation and potential role in the developmental origins of health and disease. Reprod. Fertil. Dev. 2007, 20, 1–8. [Google Scholar] [CrossRef]
- Spencer, T.E. Early pregnancy: Concepts, challenges, and potential solutions. Anim. Front. 2013, 3, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Guillomot, M. Cellular interactions during implantation in domestic ruminants. J. Reprod. Fertil. Suppl. 1995, 49, 39–51. [Google Scholar] [CrossRef]
- Forde, N.; Lonergan, P. Transcriptomic Analysis of the Bovine Endometrium: What is Required to Establish Uterine Receptivity to Implantation in Cattle ? J. Reprod. Dev. 2012, 58, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, Q.; Wang, H.; Duan, E. Uterine Fluid in Pregnancy: A Biological and Clinical Outlook. Trends Mol. Med. 2017, 23, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.H.; Rome, S.; Jalabert, A.; Forterre, A.; Singh, H.; Hincks, C.L.; Salamonsen, L.A. Endometrial Exosomes/Microvesicles in the Uterine Microenvironment: A New Paradigm for Embryo-Endometrial Cross Talk at Implantation. PLoS ONE 2013, 8, e58502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannetta, D.; Dragovic, R.; Alyahyaei, Z.; Southcombe, J. Extracellular vesicles and reproduction-promotion of successful pregnancy. Cell. Mol. Immunol. 2014, 11, 548–563. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Bazer, F.W. Temporal and spatial alterations in uterine estrogen receptor and progesterone receptor gene expression during the estrous cycle and early pregnancy in the ewe. Biol. Reprod. 1995, 53, 1527–1543. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.M.; Chen, Y.; Ezashi, T.; Walker, A.M. Interferons and the maternal–conceptus dialog in mammals. Semin. Cell Dev. Biol. 2008, 19, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Imakawa, K.; Anthony, R.V.; Kazemi, M.; Marotti, K.R.; Polites, H.G.; Roberts, R.M. Interferon-like sequence of ovine trophoblast protein secreted by embryonic trophectoderm. Nature 1987, 330, 377–379. [Google Scholar] [CrossRef]
- Bazer, F.W.; Ott, T.L.; Spencer, T.E. Pregnancy recognition in ruminants, pigs and horses: Signals from the trophoblast. Theriogenology 1994, 41, 79–94. [Google Scholar] [CrossRef]
- Ticconi, C.; Zicari, A.; Belmonte, A.; Realacci, M.; Rao, C.V.; Piccione, E. Pregnancy-Promoting Actions of HCG in Human Myometrium and Fetal Membranes. Placenta 2007, 28, S137–S143. [Google Scholar] [CrossRef]
- Mellisho, E.A.; Velásquez, A.E.; Nuñez, M.J.; Cabezas, J.G.; Cueto, J.A.; Fader, C.; Castro, F.O.; Rodríguez-Álvarez, L. Identification and characteristics of extracellular vesicles from bovine blastocysts produced in vitro. PLoS ONE 2017, 12, e0178306. [Google Scholar] [CrossRef]
- Burns, G.W.; Brooks, K.E.; Spencer, T.E. Extracellular Vesicles Originate from the Conceptus and Uterus During Early Pregnancy in Sheep. Biol. Reprod. 2016, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Kusama, K.; Bai, R.; Sakurai, T.; Isuzugawa, K.; Godkin, J.D.; Suda, Y.; Imakawa, K. Induction of IFNT-Stimulated Genes by Conceptus-Derived Exosomes during the Attachment Period. PLoS ONE 2016, 11, e0158278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, É.; Pap, E.; Kittel, Á.; et al. Membrane vesicles, current state-of-the-art: Emerging role of extracellular vesicles. Cell. Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [Green Version]
- Da Silveira, J.C.; Veeramachaneni, D.N.R.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-Secreted Vesicles in Equine Ovarian Follicular Fluid Contain miRNAs and Proteins: A Possible New Form of Cell Communication Within the Ovarian Follicle. Biol. Reprod. 2012, 86. [Google Scholar] [CrossRef]
- Andrade, G.; Meirelles, F.; Perecin, F.; da Silveira, J. Cellular and extracellular vesicular origins of miRNAs within the bovine ovarian follicle. Reprod. Domest. Anim. 2017, 52, 1036–1045. [Google Scholar] [CrossRef]
- Greening, D.W.; Nguyen, H.P.T.; Elgass, K.; Simpson, R.J.; Salamonsen, L.A. Human Endometrial Exosomes Contain Hormone-Specific Cargo Modulating Trophoblast Adhesive Capacity: Insights into Endometrial-Embryo Interactions. Biol. Reprod. 2016, 94, 38. [Google Scholar] [CrossRef]
- Kropp, J.; Salih, S.M.; Khatib, H. Expression of microRNAs in bovine and human pre-implantation embryo culture media. Front. Genet. 2014, 5, 91. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.; Lotvall, J. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.J.; Jensen, S.S.; Lim, J.W. Proteomic profiling of exosomes: Current perspectives. Proteomics 2008, 8, 4083–4099. [Google Scholar] [CrossRef]
- Subra, C.; Laulagnier, K.; Perret, B.; Record, M. Exosome lipidomics unravels lipid sorting at the level of multivesicular bodies. Biochimie 2007, 89, 205–212. [Google Scholar] [CrossRef]
- Saadeldin, I.M.; Oh, H.J.; Lee, B.C. Embryonic–maternal cross-talk via exosomes: Potential implications. Stem Cells Cloning Adv. Appl. 2015, 8, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Wydooghe, E.; Vandaele, L.; Heras, S.; De Sutter, P.; Deforce, D.; Peelman, L.; De Schauwer, C.; Van Soom, A. Autocrine embryotropins revisited: How do embryos communicate with each other in vitro when cultured in groups? Biol. Rev. 2017, 92, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.M.; Ealy, A.D.; Alexenko, A.P.; Han, C.S.; Ezashi, T. Trophoblast Interferons. Placenta 1999, 20, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Austin, K.J.; Carr, A.L.; Pru, J.K.; Hearne, C.E.; George, E.L.; Belden, E.L.; Hansen, T.R. Localization of ISG15 and Conjugated Proteins in Bovine Endometrium Using Immunohistochemistry and Electron Microscopy. Endocrinology 2004, 145, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.F.; Henkes, L.E.; Ashley, R.L.; Purcell, S.H.; Smirnova, N.P.; Veeramachaneni, D.N.R.; Anthony, R.V.; Hansen, T.R. Expression of Interferon (IFN)-Stimulated Genes in Extrauterine Tissues during Early Pregnancy in Sheep Is the Consequence of Endocrine IFN-τ Release from the Uterine Vein. Endocrinology 2008, 149, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Antoniazzi, A.Q.; Webb, B.T.; Romero, J.J.; Ashley, R.L.; Smirnova, N.P.; Henkes, L.E.; Bott, R.C.; Oliveira, J.F.; Niswender, G.D.; Bazer, F.W.; et al. Endocrine delivery of interferon tau protects the corpus luteum from prostaglandin F2 alpha-induced luteolysis in ewes. Biol Reprod 2013, 88, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridi, A.; Bertolin, K.; Rissi, V.B.; Mujica, L.K.S.; Glanzner, W.G.; Macedo, M.P.d.; Comim, F.V.; Gonçalves, P.B.D.; Antoniazzi, A.Q. Parthenogenetic bovine embryos secrete type I interferon capable of stimulating ISG15 in luteal cell culture. Anim. Reprod. 2018, 15, 1268–1277. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Austin, K.J.; Rempel, L.A.; Hansen, T.R. Low blood ISG15 mRNA and progesterone levels are predictive of non-pregnant dairy cows. J. Endocrinol. 2006, 191, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Meyerholz, M.M.; Mense, K.; Knaack, H.; Sandra, O.; Schmicke, M. Pregnancy-Induced ISG-15 and MX-1 Gene Expression is Detected in the Liver of Holstein–Friesian Heifers During Late Peri-Implantation Period. Reprod. Domest. Anim. 2016, 51, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Sponchiado, M.; Gomes, N.S.; Fontes, P.K.; Martins, T.; del Collado, M.; Pastore, A.d.A.; Pugliesi, G.; Nogueira, M.F.G.; Binelli, M. Pre-hatching embryo-dependent and -independent programming of endometrial function in cattle. PLoS ONE 2017, 12, e0175954. [Google Scholar] [CrossRef] [PubMed]
- Sponchiado, M.; Gonella-Diaza, A.M.; Rocha, C.C.; Turco, E.G.L.; Pugliesi, G.; Leroy, J.L.M.R.; Binelli, M. The pre-hatching bovine embryo transforms the uterine luminal metabolite composition in vivo. Sci. Rep. 2019, 9, 8354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bebington, C.; Doherty, F.J.; Fleming, S.D. Ubiquitin cross-reactive protein gene expression is increased in decidualized endometrial stromal cells at the initiation of pregnancy. Mol. Hum. Reprod. 1999, 5, 966–972. [Google Scholar] [CrossRef] [Green Version]
- Austin, K.J.; Bany, B.M.; Belden, E.L.; Rempel, L.A.; Cross, J.C.; Hansen, T.R. Interferon-Stimulated Gene-15 (Isg15) Expression Is Up-Regulated in the Mouse Uterus in Response to the Implanting Conceptus. Endocrinology 2003, 144, 3107–3113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashley, R.L.; Henkes, L.E.; Bouma, G.J.; Pru, J.K.; Hansen, T.R. Deletion of the Isg15 Gene Results in Up-Regulation of Decidual Cell Survival Genes and Down-Regulation of Adhesion Genes: Implication for Regulation by IL-1β. Endocrinology 2010, 151, 4527–4536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.E.; Forde, N.; Dorniak, P.; Hansen, T.R.; Romero, J.J.; Lonergan, P. Conceptus-derived prostaglandins regulate gene expression in the endometrium prior to pregnancy recognition in ruminants. Reproduction 2013, 146, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, T.; Gillio-Meina, C.; Phang, S. Prostaglandins and the initiation of blastocyst implantation and decidualization. Reproduction 2007, 134, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Ottobre, J.S.; Vincent, D.L.; Silvia, W.J.; Inskeep, E.K. Aspects of regulation of uterine secretion of prostaglandins during the oestrous cycle and early pregnancy. Anim. Reprod. Sci. 1984, 7, 75–100. [Google Scholar] [CrossRef]
- Weber, J.A.; Freeman, D.A.; Vanderwall, D.K.; Woods, G.L. Prostaglandin E2 Secretion by Oviductal Transport-Stage Equine Embryos1. Biol. Reprod. 1991, 45, 540–543. [Google Scholar] [CrossRef]
- Weber, J.A.; Freeman, D.A.; Vanderwall, D.K.; Woods, G.L. Prostaglandin E2 Hastens Oviductal Transport of Equine Embryos1. Biol. Reprod. 1991, 45, 544–546. [Google Scholar] [CrossRef]
- Woclawek-Potocka, I.; Brzezicka, E.; Skarzynski, D.J. Lysophosphatic Acid Modulates Prostaglandin Secretion in the Bovine Endometrial Cells Differently on Days 8-10 of the Estrous Cycle and Early Pregnancy. J. Reprod. Dev. 2009, 55, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.M.; Adam, M.; Hammond, J.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [PubMed]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569. [Google Scholar] [CrossRef]
- Morelli, A.E.; Larregina, A.T.; Shufesky, W.J.; Sullivan, M.L.G.; Stolz, D.B.; Papworth, G.D.; Zahorchak, A.F.; Logar, A.J.; Wang, Z.; Watkins, S.C.; et al. Endocytosis, intracellular sorting, and processing of exosomes by dendritic cells. Blood 2004, 104, 3257–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, E.; Guérin, C.; Hogg, N.; Amigorena, S.; Théry, C. CD8+ Dendritic Cells Use LFA-1 to Capture MHC-Peptide Complexes from Exosomes In Vivo. J. Immunol. 2007, 179, 1489–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleys, E.R.; Halleran, J.L.; McWhorter, E.; Hergenreder, J.; Enriquez, V.A.; da Silveira, J.C.; Bruemmer, J.E.; Winger, Q.A.; Bouma, G.J. Identification of microRNAs in exosomes isolated from serum and umbilical cord blood, as well as placentomes of gestational day 90 pregnant sheep. Mol. Reprod. Dev. 2014, 81, 983–993. [Google Scholar] [CrossRef]
- Street, J.M.; Barran, P.E.; Mackay, C.L.; Weidt, S.; Balmforth, C.; Walsh, T.S.; Chalmers, R.T.A.; Webb, D.J.; Dear, J.W. Identification and proteomic profiling of exosomes in human cerebrospinal fluid. J. Transl. Med. 2012, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prunotto, M.; Farina, A.; Lane, L.; Pernin, A.; Schifferli, J.; Hochstrasser, D.F.; Lescuyer, P.; Moll, S. Proteomic analysis of podocyte exosome-enriched fraction from normal human urine. J. Proteom. 2013, 82, 193–229. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Buzás, E.I.; Bemis, L.T.; Bora, A.; Lässer, C.; Lötvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J. Extracell Vesicles 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- De Ávila, A.C.F.C.M.; Bridi, A.; Andrade, G.M.; del Collado, M.; Sangalli, J.R.; Nociti, R.P.; da Silva Junior, W.A.; Bastien, A.; Robert, C.; Meirelles, F.V.; et al. Estrous cycle impacts miRNA content in extracellular vesicles that modulate bovine cumulus cell transcripts during in vitro maturation. Biol. Reprod. 2019. [Google Scholar] [CrossRef] [PubMed]
- Andrade, G.M.; Bomfim, M.M.; del Collado, M.; Meirelles, F.V.; Perecin, F.; da Silveira, J.C. Oxygen tension modulates extracellular vesicles and its miRNA contents in bovine embryo culture medium. Mol. Reprod. Dev. 2019, 86, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Qing, S.; Liu, R.; Qin, H.; Wang, W.; Qiao, F.; Ge, H.; Liu, J.; Zhang, Y.; Cui, W.; et al. Effects of embryo-derived exosomes on the development of bovine cloned embryos. PLoS ONE 2017, 12, e0174535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange-Consiglio, A.; Perrini, C.; Albini, G.; Modina, S.; Lodde, V.; Orsini, E.; Esposti, P.; Cremonesi, F. Oviductal microvesicles and their effect on in vitro maturation of canine oocytes. Reproduction 2017, 154, 167. [Google Scholar] [CrossRef]
- Saadeldin, I.M.; Kim, S.J.; Choi, Y.B.; Lee, B.C. Improvement of Cloned Embryos Development by Co-Culturing with Parthenotes: A Possible Role of Exosomes/Microvesicles for Embryos Paracrine Communication. Cell. Reprogramming 2014, 16, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Almiñana, C.; Corbin, E.; Tsikis, G.; Alcântara-Neto, A.S.; Labas, V.; Reynaud, K.; Galio, L.; Uzbekov, R.; Garanina, A.S.; Druart, X.; et al. Oviduct extracellular vesicles protein content and their role during oviduct–embryo cross-talk. Reproduction 2017, 154, 253. [Google Scholar] [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Ramírez, M.Á.; Yáñez-Mó, M.; Rizos, D. Effect of bovine oviductal extracellular vesicles on embryo development and quality in vitro. Reproduction 2017, 153, 461. [Google Scholar] [CrossRef] [Green Version]
- Qu, P.; Zhao, Y.; Wang, R.; Zhang, Y.; Li, L.; Fan, J.; Liu, E. Extracellular vesicles derived from donor oviduct fluid improved birth rates after embryo transfer in mice. Reprod. Fertil. Dev. 2019, 31, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-González, I.; Xu, J.; Wang, X.; Burghardt, R.C.; Dunlap, K.A.; Bazer, F.W. Exosomes, endogenous retroviruses and toll-like receptors: Pregnancy recognition in ewes. Reproduction 2015, 149, 281–291. [Google Scholar] [CrossRef]
- Kusama, K.; Nakamura, K.; Bai, R.; Nagaoka, K.; Sakurai, T.; Imakawa, K. Intrauterine exosomes are required for bovine conceptus implantation. Biochem. Biophys. Res. Commun. 2018, 495, 1370–1375. [Google Scholar] [CrossRef]
- Burns, G.W.; Brooks, K.E.; O’Neil, E.V.; Hagen, D.E.; Behura, S.K.; Spencer, T.E. Progesterone effects on extracellular vesicles in the sheep uterus†. Biol. Reprod. 2018, 98, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Bidarimath, M.; Khalaj, K.; Kridli, R.T.; Kan, F.W.K.; Koti, M.; Tayade, C. Extracellular vesicle mediated intercellular communication at the porcine maternal-fetal interface: A new paradigm for conceptus-endometrial cross-talk. Sci. Rep. 2017, 7, 40476. [Google Scholar] [CrossRef]
- Brower, L.K.; Anderson, E. Cytological Events Associated with the Secretory Process in the Rabbit Oviduct1. Biol. Reprod. 1969, 1, 130–148. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.W.; Plante, C.; Allan King, W.; Hansen, P.J.; Betteridge, K.J.; Suarez, S.S. Fertilizing Capacity of Bovine Sperm may be Maintained by Binding to Oviductal Epithelial Cells1. Biol. Reprod. 1991, 44, 102–107. [Google Scholar] [CrossRef]
- Almiñana, C.; Bauersachs, S. Extracellular Vesicles in the Oviduct: Progress, Challenges and Implications for the Reproductive Success. Bioengineering 2019, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Della-Giustina, D.; Denny, M. Ectopic pregnancy. Emerg. Med. Clin. N. Am. 2003, 21, 565–584. [Google Scholar] [CrossRef]
- Murray, H.; Baakdah, H.; Bardell, T.; Tulandi, T. Diagnosis and treatment of ectopic pregnancy. CMAJ 2005, 173, 905–912. [Google Scholar] [CrossRef] [Green Version]
- Pisarska, M.D.; Carson, S.A.; Buster, J.E. Ectopic pregnancy. Lancet 1998, 351, 1115–1120. [Google Scholar] [CrossRef]
- Brennan, D.F. Ectopic Pregnancy—Part I: Clinical and Laboratory Diagnosis. Acad. Emerg. Med. 1995, 2, 1081–1089. [Google Scholar] [CrossRef]
- ACOG Practice Bulletin No. 94: Medical management of ectopic pregnancy. Obstet. Gynecol. 2008, 111, 1479–1485.
- Zhao, Z.; Zhao, Q.; Warrick, J.; Lockwood, C.M.; Woodworth, A.; Moley, K.H.; Gronowski, A.M. Circulating MicroRNA miR-323-3p as a Biomarker of Ectopic Pregnancy. Clin. Chem. 2012, 58, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, D.; Barratt, C.L.R.; Björndahl, L.; de Jager, C.; Jequier, A.M.; Muller, C.H. What should it take to describe a substance or product as ‘sperm-safe’. Hum. Reprod. Update 2013, 19, i1–i45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazer, F.W.; Wu, G.; Johnson, G.A.; Kim, J.; Song, G. Uterine Histotroph and Conceptus Development: Select Nutrients and Secreted Phosphoprotein 1 Affect Mechanistic Target of Rapamycin Cell Signaling in Ewes1. Biol. Reprod. 2011, 85, 1094–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.E.; Bazer, F.W. Biology of progesterone action during pregnancy recognition and maintenance of pregnancy. Front. Biosci. 2002, 7, d1879–d1898. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.A.; Bazer, F.W.; Jaeger, L.A.; Ka, H.; Garlow, J.E.; Pfarrer, C.; Spencer, T.E.; Burghardt, R.C. Muc-1, Integrin, and Osteopontin Expression During the Implantation Cascade in Sheep. Biol. Reprod. 2001, 65, 820–828. [Google Scholar] [CrossRef] [Green Version]
- Ginther, O.J. INTERNAL REGULATION OF PHYSIOLOGICAL PROCESSES THROUGH LOCAL VENOARTERIAL PATHWAYS: A REVIEW. J. Anim. Sci. 1974, 39, 550–564. [Google Scholar] [CrossRef]
- Klohonatz, K.M.; Cameron, A.D.; Hergenreder, J.R.; da Silveira, J.C.; Belk, A.D.; Veeramachaneni, D.N.R.; Bouma, G.J.; Bruemmer, J.E. Circulating miRNAs as Potential Alternative Cell Signaling Associated with Maternal Recognition of Pregnancy in the Mare1. Biol. Reprod. 2016, 95, 124. [Google Scholar] [CrossRef]
- Burghardt, R.C.; Burghardt, J.R.; Taylor, J.D.; Reeder, A.T.; Nguen, B.T.; Spencer, T.E.; Bayless, K.J.; Johnson, G.A. Enhanced focal adhesion assembly reflects increased mechanosensation and mechanotransduction at maternal–conceptus interface and uterine wall during ovine pregnancy. Reproduction 2009, 137, 567–582. [Google Scholar] [CrossRef] [Green Version]
- Pohler, K.G.; Green, J.A.; Moley, L.A.; Gunewardena, S.; Hung, W.-T.; Payton, R.R.; Hong, X.; Christenson, L.K.; Geary, T.W.; Smith, M.F. Circulating microRNA as candidates for early embryonic viability in cattle. Mol. Reprod. Dev. 2017, 84, 731–743. [Google Scholar] [CrossRef]
- De Bem, T.H.C.; da Silveira, J.C.; Sampaio, R.V.; Sangalli, J.R.; Oliveira, M.L.F.; Ferreira, R.M.; Silva, L.A.; Perecin, F.; King, W.A.; Meirelles, F.V.; et al. Low levels of exosomal-miRNAs in maternal blood are associated with early pregnancy loss in cloned cattle. Sci. Rep. 2017, 7, 14319. [Google Scholar] [CrossRef]
- Markkandan, K.; Ahn, K.; Lee, D.J.; Kim, T.I.; Dang, C.; Hong, S.-E.; Yoon, H.-B.; Lim, H.-J.; Hong, C.P. Profiling and identification of pregnancy-associated circulating microRNAs in dairy cattle. Genes Genom. 2018, 40, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
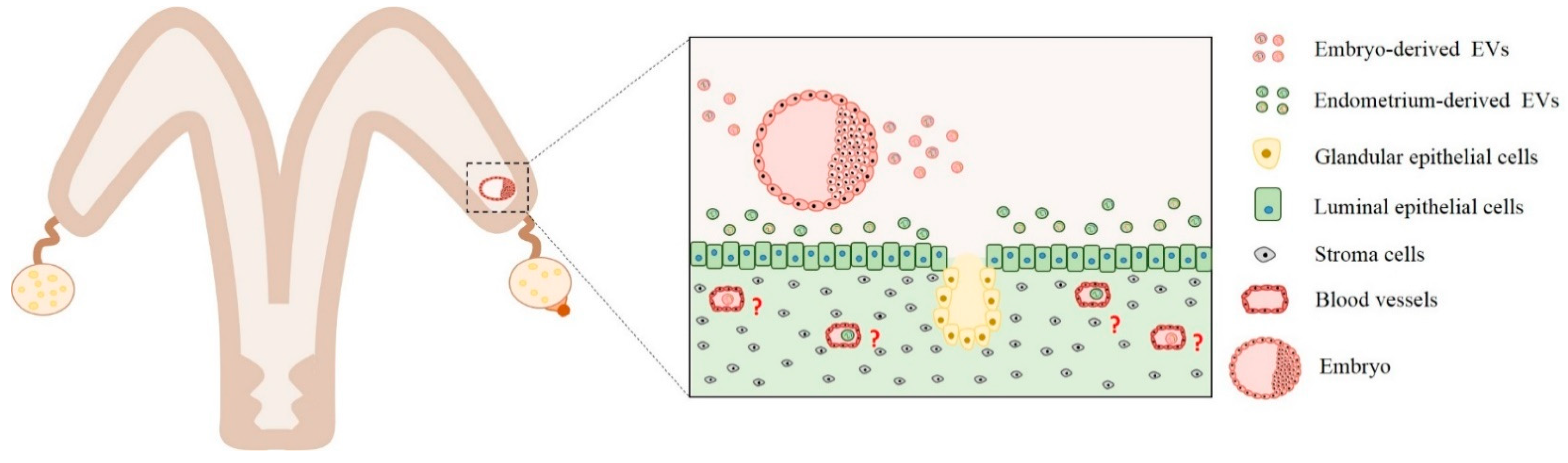
Figure 1.
Embryo–maternal interactions mediated by extracellular vesicles secreted from the embryo and endometrium cells. Extracellular vesicles secreted by embryos are uptaken by endometrial cells. Extracellular vesicles (EVs) content include miRNA, mRNAs, and proteins that act by paracrine signaling. Different experiments demonstrated that embryo-derived EVs can modulate interferon-tau, apoptosis, cell proliferation, adhesion, angiogenesis, and cell survival biological pathways. Similarly, endometrium cells can secret EVs containing proteins, miRNAs, and mRNAs. These EVs are uptaken by trophoblast cells, and their contents are predicted to regulate interferon-tau production and angiogenesis pathways. However, questions related to how embryo-derived EVs can exit the uterus and arrive in other maternal tissue cells are yet unclear (red interrogation points). Therefore, further studies are needed to understand how the factors secreted by the embryos exit the uterus and to arrive in other maternal tissue, as blood cells, to modulate biological pathways.
Figure 1.
Embryo–maternal interactions mediated by extracellular vesicles secreted from the embryo and endometrium cells. Extracellular vesicles secreted by embryos are uptaken by endometrial cells. Extracellular vesicles (EVs) content include miRNA, mRNAs, and proteins that act by paracrine signaling. Different experiments demonstrated that embryo-derived EVs can modulate interferon-tau, apoptosis, cell proliferation, adhesion, angiogenesis, and cell survival biological pathways. Similarly, endometrium cells can secret EVs containing proteins, miRNAs, and mRNAs. These EVs are uptaken by trophoblast cells, and their contents are predicted to regulate interferon-tau production and angiogenesis pathways. However, questions related to how embryo-derived EVs can exit the uterus and arrive in other maternal tissue cells are yet unclear (red interrogation points). Therefore, further studies are needed to understand how the factors secreted by the embryos exit the uterus and to arrive in other maternal tissue, as blood cells, to modulate biological pathways.


{kind=link}
Table 1.
Major findings about extracellular vesicles (EVs) derived from the reproductive female tract and embryos in mammals. IFNT: interferon-tau.
Table 1.
Major findings about extracellular vesicles (EVs) derived from the reproductive female tract and embryos in mammals. IFNT: interferon-tau.
| Topic | Findings | Reference |
|---|---|---|
| EVs derived from ovarian follicles |
| Da Silveira et al., 2012 [24] |
| ||
| EVs derived from in vitro produced embryos |
| Mellisho et al., 2017 [20] |
| Saadeldin et al., 2014 [65] | |
| Qu et al., 2017 [63] | |
| Oviductal EVs |
| Lange-Consiglio et al., 2017 [64] |
| Al-Dossary et al., 2013 [5] | |
| Lopera-Vásquez et al., 2016; Almiñana et al., 2017; Lopera-Vasquez et al., 2017 [7,66,67] | |
| Qu et al., 2019 [68] | |
| Embryonic and uterine EVs |
| Burns et al., 2016 [21] |
| Ruiz-González et al., 2015 [69] | |
| Nakamura et al., 2016; Kusama et al., 2018 [22,70] | |
| Nakamura et al., 2016 [22] | |
| Burns et al., 2018 [71] | |
| ||
| Bidarimath et al., 2017 [72] |
Table 2.
The main published papers about extracellular vesicle-associated miRNAs in pregnant female blood.
Table 2.
The main published papers about extracellular vesicle-associated miRNAs in pregnant female blood.
| Species | Period of Pregnancy | Circulating Exosomal miRNAs in Pregnant Female | Pathways Modulated by miRNAs | Reference |
|---|---|---|---|---|
| Equine | Day 9 | Down-regulated: eca-miR-27a, eca-miR-29c, eca-miR-101, and eca-miR-486-5p | Extracellular matrix (ECM)–receptor interaction Proteoglycans in cancer TGF-beta Focal adhesion pathway | Klohonatz et al., 2016 [87] |
| Day 11 | Down-regulated: eca-miR-195 Up-regulated: eca-miR-767-5p | Regulating pluripotency of stem cells Fatty acid biosynthesis FoxO Focal adhesion pathway | ||
| Day 13 | Down-regulated: eca-miR-188-5p, eca-miR-653, eca-miR-874, and eca-miR-140-3p Up-regulated: eca-miR-30c and eca-miR-323-5p | Glutamatergic synapse Regulation of actin cytoskeleton Long-term potentiation Focal adhesion pathway | ||
| Bovine | Day 17 | Down-regulated: bta-let-7c, bta-mir-100, bta-mir-101-1, bta-mir-101-2, bta-mir-106b, bta-mir-125b-2, bta-mir-127, bta-mir-141, bta-mir-143, bta-mir-148a, bta-mir-15a, bta-mir-15b, bta-mir-16a, bta-mir-16b, bta-mir-1839, bta-mir-199a-1, bta-mir-199b, bta-mir-2415, bta-mir-25, bta-mir-331, bta-mir-339b, bta-mir-3596, bta-mir-3604-1, bta-mir-409b, bta-mir-451, bta-mir-652, bta-mir-99a | Cancer Connective tissue disorders Organismal injury and abnormalities Reproductive system disease Endocrine disorders | Pohler et al., 2017 [89] |
| Bovine | Day 21 | Highly abundant: bta-mir-15a, bta-mir-221, bta-mir-425-5p, bta-mir-101, bta-mir-93, bta-mir-106a, bta-mir-106b, bta-mir-22-5p, bta-mir-652, bta-mir-103, bta-mir-138, bta-mir-193b, bta-let-7f, bta-let-7g, bta-mir-15b, bta-let-7e, bta-let-7a-5p, bta-let-7d, bta-mir-660, bta-mir-29a, bta-mir-30d, bta-mir-497, bta-mir-148b, bta-mir-361, bta-mir-30a-5p, bta-mir-29c, bta-mir-29d-3p | Wnt TGF-beta Renal cell carcinoma Melanoma Colorectal cancer Glioma p53 Pancreatic cancer; Bladder cancer Dorsoventral axis formation | De Bem et al., 2017 [90] |
| Bovine | Day 30 | Down-regulated: bta-miR-193b, bta-miR-197, bta-miR-339a, bta-miR-326, bta-miR-484, bta-miR-486, bta-miR-423-3p, bta-miR-92a Up-regulated: bta-miR-146b, bta-miR-27b, bta-miR-26b, bta-miR-200a, bta-miR-450b, bta-miR-199c, bta-miR-194, bta-miR-6119-3p, bta-miR-7, bta-miR-199a-3p, bta-miR-574, bta-miR-215, bta-miR-148a, bta-let-7a-3p, bta-miR-21-5p, bta-miR-126-5p, bta-miR-148b, bta-miR-143, bta-miR-1246, bta-miR-192, bta-miR-98 | Membrane trafficking Chromosome and associated proteins Exosome G protein-coupled receptors Transcription factors Ubiquitin system Olfactory transduction Transporters Protein kinases Cytoskeleton proteins Cell adhesion molecules and their ligands Protein phosphatase and associated proteins Peptidases Pathway in cancer Messenger RNA Biogenesis CD molecules PI3K–Akt signaling pathway Human papillomavirus infection Mitochondrial biogenesis Spliceosome | Markkandan et al., 2018 [91] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bridi, A.; Perecin, F.; Silveira, J.C.d. Extracellular Vesicles Mediated Early Embryo–Maternal Interactions. Int. J. Mol. Sci. 2020, 21, 1163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031163
AMA Style
Bridi A, Perecin F, Silveira JCd. Extracellular Vesicles Mediated Early Embryo–Maternal Interactions. International Journal of Molecular Sciences. 2020; 21(3):1163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031163
Chicago/Turabian StyleBridi, Alessandra, Felipe Perecin, and Juliano Coelho da Silveira. 2020. "Extracellular Vesicles Mediated Early Embryo–Maternal Interactions" International Journal of Molecular Sciences 21, no. 3: 1163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031163
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.