Nuclear Integrants of Organellar DNA Contribute to Genome Structure and Evolution in Plants


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Characterization of Organellar DNA-Derived Sequences in Plants
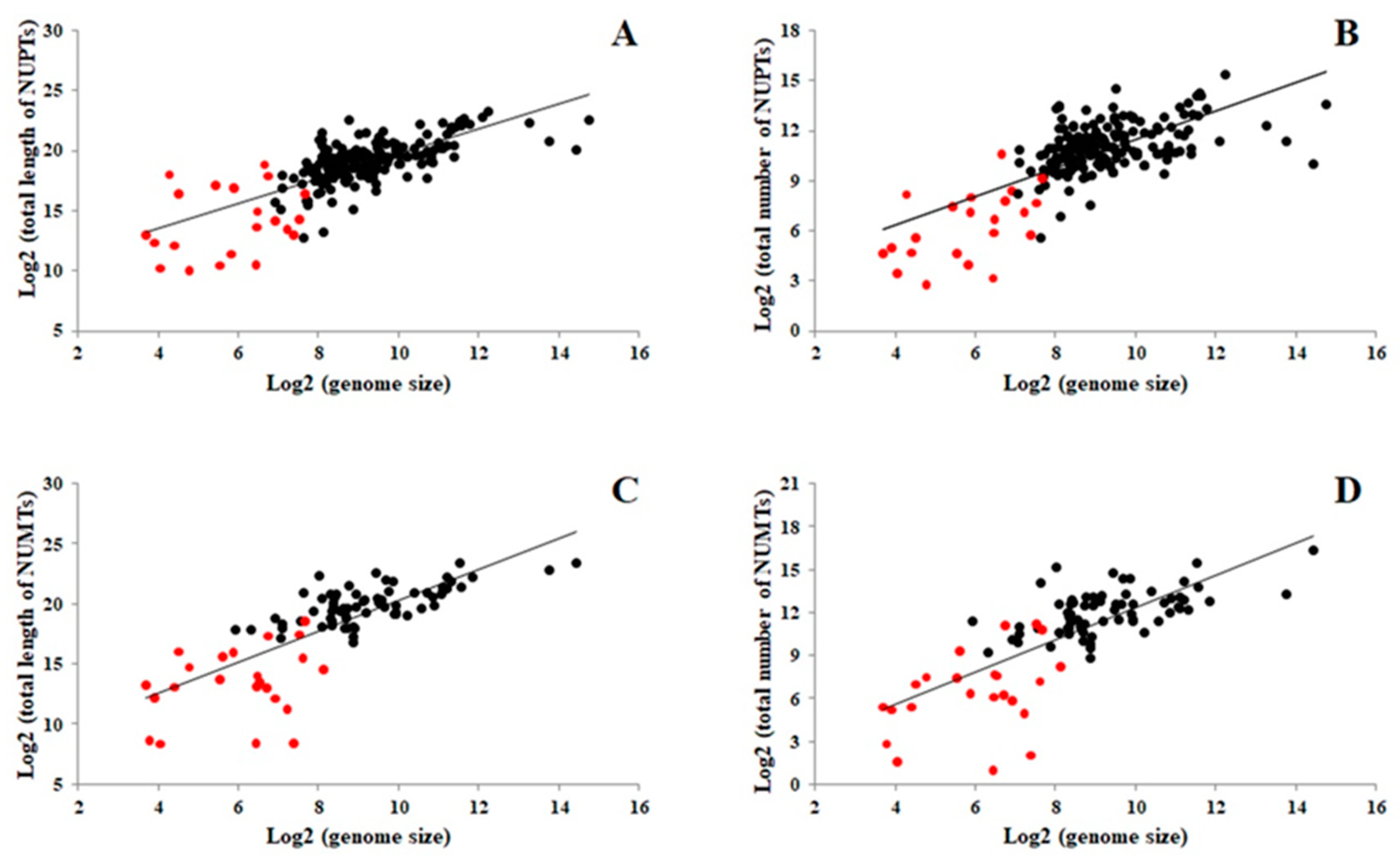
2.1. Number and Size Distribution
2.2. Organization and Distribution Patterns
2.3. Modification Pattern
2.4. Transfer Rate of Organellar DNA to Nucleus
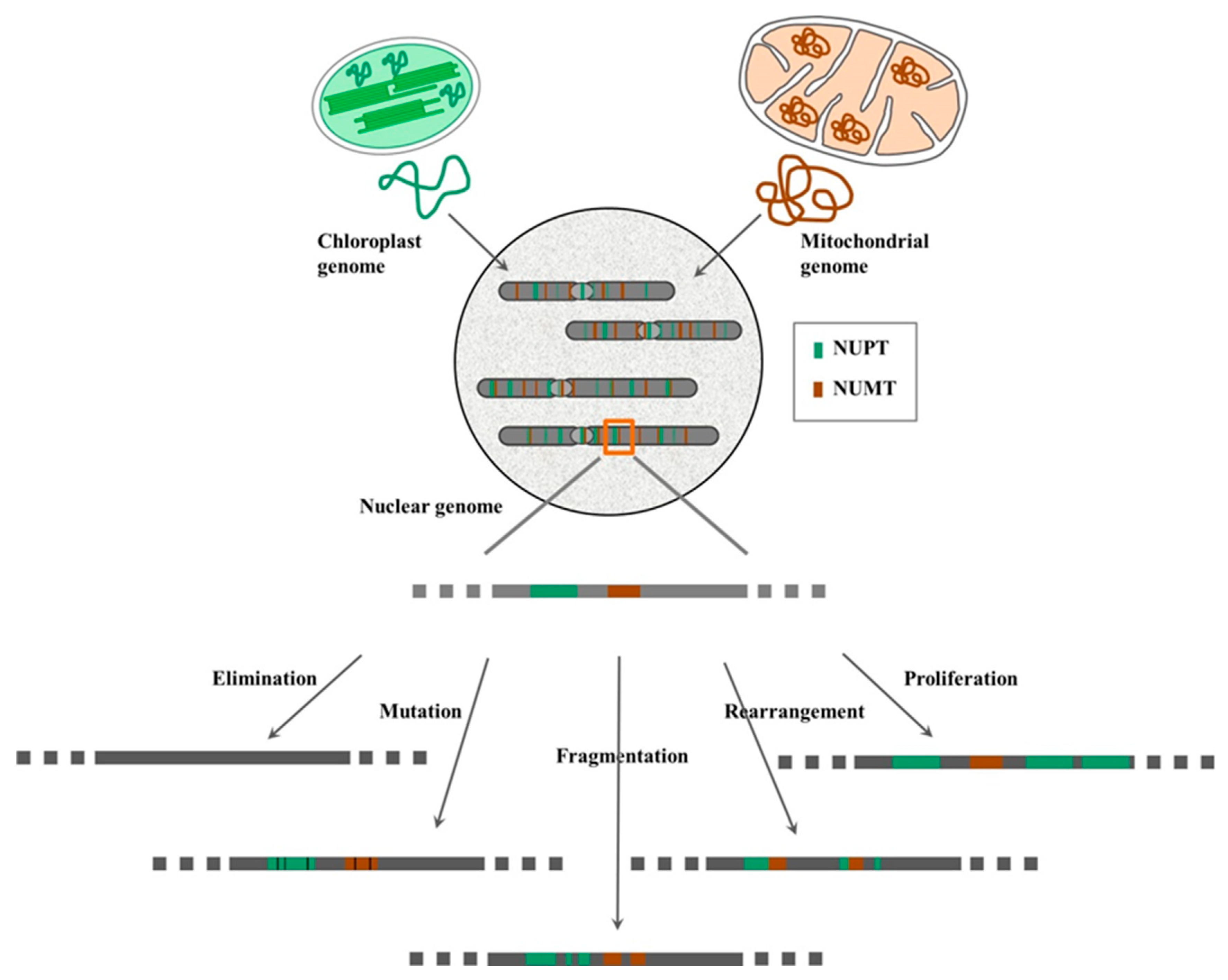
3. Genetic Consequence and Fate of NUPTs/NUMTs in Plants
3.1. Elimination
3.2. Mutation
3.3. Fragmentation
3.4. Rearrangement
3.5. Proliferation
4. Effects of Organelle DNA-Derived Sequences on the Nuclear Genome Structure and Evolution
4.1. Contribution to the Genetic Diversity
4.2. Effects on Gene Structure and Evolution
4.3. Roles Played in the Sex Chromosome Evolution
5. Mechanisms of Organellar DNA Integration
6. Conclusions and Perspective
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Douglas, S.E. Plastid evolution: Origins, diversity, trends. Curr. Opin. Genet. Dev. 1998, 8, 655–661. [Google Scholar] [CrossRef]
- Gray, M.W. The evolutionary origins of organelles. Trends Genet. 1989, 5, 294–299. [Google Scholar] [CrossRef]
- McBride, H.M. Mitochondria and endomembrane origins. Curr. Biol. 2018, 28, R367–R372. [Google Scholar] [CrossRef] [Green Version]
- Kleine, T.; Maier, U.G.; Leister, D. DNA transfer from organelles to the nucleus: the idiosyncratic genetics of endosymbiosis. Annu. Rev. Plant Biol. 2009, 60, 115–138. [Google Scholar] [CrossRef] [Green Version]
- Ma, P.F.; Zhang, Y.X.; Guo, Z.H.; Li, D.Z. Evidence for horizontal transfer of mitochondrial DNA to the plastid genome in a bamboo genus. Sci. Rep. 2015, 5, 11608. [Google Scholar] [CrossRef]
- Pinard, D.; Myburg, A.A.; Mizrachi, E. The plastid and mitochondrial genomes of Eucalyptus grandis. BMC Genom. 2019, 20, 132. [Google Scholar] [CrossRef]
- Timmis, J.N.; Ayliffe, M.A.; Huang, C.Y.; Martin, W.F. Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 2004, 5, 123–135. [Google Scholar] [CrossRef]
- Ayliffe, M.A.; Scott, N.S.; Timmis, J.N. Analysis of plastid DNA-like sequences within the nuclear genomes of higher plants. Mol. Biol. Evol. 1998, 15, 738–745. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Hill, J.; Hsiao, J.; Moffat, K.; Ouyang, S.; Cheng, Z.; Jiang, J.; Buell, C. Genome sequencing of a 239-kb region of rice chromosome 10L reveals a high frequency of gene duplication and a large chloroplast DNA insertion. Mol. Genet. Genom. 2002, 267, 713–720. [Google Scholar] [CrossRef]
- Huang, C.Y.; Ayliffe, M.A.; Timmis, J.N. Direct measurement of the transfer rate of chloroplast DNA into the nucleus. Nature 2003, 422, 72–76. [Google Scholar] [CrossRef]
- Huang, C.Y.; Ayliffe, M.A.; Timmis, J.N. Simple and complex nuclear loci created by newly transferred chloroplast DNA in tobacco. Proc. Natl. Acad. Sci. USA 2004, 101, 9710–9715. [Google Scholar] [CrossRef] [Green Version]
- Stegemann, S.; Hartmann, S.; Ruf, S.; Bock, R. High-frequency gene transfer from the chloroplast genome to the nucleus. Proc. Natl. Acad. Sci. USA 2003, 100, 8828–8833. [Google Scholar] [CrossRef] [Green Version]
- Roark, L.; Hui, A.; Donnelly, L.; Birchler, J.; Newton, K. Recent and frequent insertions of chloroplast DNA into maize nuclear chromosomes. Cytogenet. Genome Res. 2010, 129, 17–23. [Google Scholar] [CrossRef]
- Sloan, D.B.; Warren, J.M.; Williams, A.M.; Wu, Z.; Abdel-Ghany, S.E.; Chicco, A.J.; Havird, J.C. Cytonuclear integration and co-evolution. Nat. Rev. Genet. 2018, 19, 635–648. [Google Scholar] [CrossRef]
- Chen, H.Y.; Yu, Y.; Chen, X.L.; Zhang, Z.Z.; Gong, C.; Li, J.F.; Wang, A.X. Plastid DNA insertions in plant nuclear genomes: The sites, abundance and ages, and a predicted promoter analysis. Funct. Integr. Genom. 2015, 15, 131–139. [Google Scholar] [CrossRef]
- Michalovova, M.; Vyskot, B.; Kejnovsky, E. Analysis of plastid and mitochondrial DNA insertions in the nucleus (NUPTs and NUMTs) of six plant species: Size, relative age and chromosomal localization. Heredity 2013, 111, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Furihata, H.Y.; Kawabe, A. Analysis of nuclear mitochondrial DNAs and factors affecting patterns of integration in plant species. Genes Genet. Syst. 2017, 92, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.J.; Kim, S. Analysis of nuclear mitochondrial DNA segments of nine plant species: Size, distribution, and insertion loci. Genom. Inform. 2016, 14, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Shahmuradov, I.A.; Akbarova, Y.Y.; Solovyev, V.V.; Aliyev, J.A. Abundance of plastid DNA insertions in nuclear genomes of rice and Arabidopsis. Plant Mol. Biol. 2003, 52, 923–934. [Google Scholar] [CrossRef]
- Kim, H.T.; Lee, J.M. Organellar genome analysis reveals endosymbiotic gene transfers in tomato. PLoS ONE 2018, 13, e0202279. [Google Scholar] [CrossRef] [Green Version]
- Li, S.F.; Li, J.R.; Wang, J.; Dong, R.; Jia, K.L.; Zhu, H.W.; Li, N.; Yuan, J.H.; Deng, C.L.; Gao, W.J. Cytogenetic and genomic organization analyses of chloroplast DNA invasions in the nuclear genome of Asparagus officinalis L. provides signatures of evolutionary complexity and informativity in sex chromosome evolution. BMC Plant Biol. 2019, 19, 361. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F.; Rujan, T.; Richly, E.; Hansen, A.; Cornelsen, S.; Lins, T.; Leister, D.; Stoebe, B.; Hasegawa, M.; Penny, D. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc. Natl. Acad. Sci. USA 2002, 99, 12246–12251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noutsos, C.; Kleine, T.; Armbruster, U.; DalCorso, G.; Leister, D. Nuclear insertions of organellar DNA can create novel patches of functional exon sequences. Trends Genet. 2007, 23, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Knoop, V.; Brennicke, A. A mitochondrial intron sequence in the 5’-flanking region of a plant nuclear lectin gene. Curr. Genet. 1991, 20, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Boogaart, P.V.D.; Samallo, J.; Agsteribbe, E. Similar genes for a mitochondrial ATPase subunit in the nuclear and mitochondrial genomes of Neurospora crassa. Nature 1982, 298, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.V.; Yuhki, N.; Masuda, R.; Modi, W.; O’Brien, S.J. Numt, a recent transfer and tandem amplification of mitochondrial DNA to the nuclear genome of the domestic cat. J. Mol. Evol. 1994, 39, 174–190. [Google Scholar]
- Mourier, T.; Hansen, A.J.; Willerslev, E.; Arctander, P. The Human Genome Project reveals a continuous transfer of large mitochondrial fragments to the nucleus. Mol. Biol. Evol. 2001, 18, 1833–1837. [Google Scholar] [CrossRef] [Green Version]
- Hazkani-Covo, E.; Graur, D. A comparative analysis of numt evolution in human and chimpanzee. Mol. Biol. Evol. 2007, 24, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Hazkani-Covo, E.; Zeller, R.M.; Martin, W.F. Molecular poltergeists: Mitochondrial DNA copies (numts) in sequenced nuclear genomes. PLoS Genet. 2010, 6, e1000834. [Google Scholar] [CrossRef]
- Ding, Y.R.; Li, B.; Zhang, Y.J.; Mao, Q.M.; Chen, B. Complete mitogenome of Anopheles sinensis and mitochondrial insertion segments in the nuclear genomes of 19 mosquito species. PLoS ONE 2018, 13, e0204667. [Google Scholar] [CrossRef]
- Yan, Z.C.; Fang, Q.; Tian, Y.; Wang, F.; Chen, X.X.; Werren, J.H.; Ye, G.Y. Mitochondrial DNA and their nuclear copies in the parasitic wasp Pteromalus puparum: A comparative analysis in Chalcidoidea. Int. J. Biol. Macromol. 2019, 121, 572–579. [Google Scholar] [CrossRef]
- Schiavo, G.; Strillacci, M.G.; Ribani, A.; Bovo, S.; Román-Ponce, S.I.; Cerolini, S.; Bertolini, F.; Bagnato, A.; Fontanesi, L. Few mitochondrial DNA sequences are inserted into the turkey (Meleagris gallopavo) nuclear genome: Evolutionary analyses and informativity in the domestic lineage. Anim. Genet. 2018, 49, 259–264. [Google Scholar] [CrossRef]
- Huang, C.Y.; Grünheit, N.; Ahmadinejad, N.; Timmis, J.N.; Martin, W.F. Mutational decay and age of chloroplast and mitochondrial genomes transferred recently to angiosperm nuclear chromosomes. Plant Physiol. 2005, 138, 1723–1733. [Google Scholar] [CrossRef] [Green Version]
- The Rice Chromosome 10 Sequencing Consortium. In-depth view of structure, activity, and evolution of rice chromosome 10. Science 2003, 300, 1566–1569. [Google Scholar] [CrossRef]
- Stupar, R.M.; Lilly, J.W.; Town, C.D.; Cheng, Z.; Kaul, S.; Buell, C.R.; Jiang, J. Complex mtDNA constitutes an approximate 620-kb insertion on Arabidopsis thaliana chromosome 2: Implication of potential sequencing errors caused by large-unit repeats. Proc. Natl. Acad. Sci. USA 2001, 98, 5099–5103. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, M.; Ito, Y.; Yamauchi, R.; Obokata, J. The rice nuclear genome continuously integrates, shuffles, and eliminates the chloroplast genome to cause chloroplast–nuclear DNA flux. Plant Cell 2005, 17, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Richly, E.; Lartillot, N.; Philippe, H. NUMTs in sequenced eukaryotic genomes. Mol. Biol. Evol. 2004, 21, 1081–1084. [Google Scholar] [CrossRef] [Green Version]
- Noutsos, C.; Richly, E.; Leister, D. Generation and evolutionary fate of insertions of organelle DNA in the nuclear genomes of flowering plants. Genome Res. 2005, 15, 616–628. [Google Scholar] [CrossRef] [Green Version]
- Richly, E.; Leister, D. NUPTs in sequenced eukaryotes and their genomic organization in relation to NUMTs. Mol. Biol. Evol. 2004, 21, 1972–1980. [Google Scholar] [CrossRef] [Green Version]
- Leister, D. Origin, evolution and genetic effects of nuclear insertions of organelle DNA. Trends Genet. 2005, 21, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Portugez, S.; Martin, W.F.; Hazkani-Covo, E. Mosaic mitochondrial-plastid insertions into the nuclear genome show evidence of both non-homologous end joining and homologous recombination. BMC Evol. Biol. 2018, 18, 162. [Google Scholar] [CrossRef] [Green Version]
- Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.; Navrátilová, A.; Kobližková, A.; Kejnovsky, E.; Hribova, E.; Hobza, R.; Widmer, A.; Dolezel, J.; Macas, J. Plant centromeric retrotransposons: A structural and cytogenetic perspective. Mob. DNA 2011, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Zhang, X.; Li, B.; Liu, T.; Chen, J.; Bai, Z.; Wang, M.; Shi, J.; Walling, J.G.; Wing, R.A.; et al. Comparison of Oryza sativa and Oryza brachyantha genomes reveals selection-driven gene escape from the centromeric regions. Plant Cell 2018, 30, 1729–1744. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Timmis, J.N. Cytoplasmic organelle DNA preferentially inserts into open chromatin. Genome Biol. Evol. 2013, 5, 1060–1064. [Google Scholar] [CrossRef] [Green Version]
- Pecinka, A.; Dinh, H.Q.; Baubec, T.; Rosa, M.; Lettner, N.; Scheid, O.M. Epigenetic regulation of repetitive elements is attenuated by prolonged heat stress in Arabidopsis. Plant Cell 2010, 22, 3118–3129. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.S.; Hu, B.; Becker, C.; Doğan, E.S.; Berendzen, K.W.; Weigel, D.; Liu, C. Altered chromatin compaction and histone methylation drive non-additive gene expression in an interspecific Arabidopsis hybrid. Genome Biol. 2017, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, A.E.; Timmis, J.N. Instability of plastid DNA in the nuclear genome. PLoS Genet. 2009, 5, e1000323. [Google Scholar] [CrossRef] [Green Version]
- Li, S.F.; Zhang, G.J.; Yuan, J.H.; Deng, C.L.; Gao, W.J. Repetitive sequences and epigenetic modification: Inseparable partners play important roles in the evolution of plant sex chromosomes. Planta 2016, 243, 1083–1095. [Google Scholar] [CrossRef]
- Galindo-González, L.; Sarmiento, F.; Quimbaya, M.A. Shaping plant adaptability, genome structure and gene expression through transposable element epigenetic control: Focus on methylation. Agronomy 2018, 8, 180. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Furihata, H.Y.; To, T.K.; Kakutani, T.; Kawabe, A. Genome defense against integrated organellar DNA fragments from plastids into plant nuclear genomes through DNA methylation. Sci. Rep. 2019, 9, 2060. [Google Scholar] [CrossRef] [Green Version]
- Holliday, R.; Grigg, G. DNA methylation and mutation. Mutat. Res. 1993, 285, 61–67. [Google Scholar] [CrossRef]
- Paszkowski, J.; Whitham, S.A. Gene silencing and DNA methylation processes. Curr. Opin. Plant Biol. 2001, 4, 123–129. [Google Scholar] [CrossRef]
- Bender, J. Chromatin-based silencing mechanisms. Curr. Opin. Plant Biol. 2004, 7, 521–526. [Google Scholar] [CrossRef]
- Wang, D.; Lloyd, A.H.; Timmis, J.N. Environmental stress increases the entry the cytoplasmic organellar DNA into the nucleus in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 2444–2448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Lloyd, A.H.; Timmis, J.N. Nuclear genome diversity in somatic cells is accelerated by environmental stress. Plant Signal. Behav. 2012, 7, 595–597. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.R.; Crosby, K.; Lee, R.W. Correlation between nuclear plastid DNA abundance and plastid number supports the limited transfer window hypothesis. Genome Biol. Evol. 2011, 3, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Thorsness, P.E.; Fox, T.D. Escape of DNA from mitochondria to the nucleus in Saccharomyces cerevisiae. Nature 1990, 346, 376–379. [Google Scholar] [CrossRef]
- Ruf, S.; Hasse, C.; Stegemann, S.; Bock, R. Plastid Transmission, Gene Transfer and the Impact of the Environment; ISCGGE: Maynooth, Ireland, 2010. [Google Scholar]
- Sheppard, A.E.; Ayliffe, M.A.; Blatch, L.; Day, A.; Delaney, S.K.; Khairul-Fahmy, N.; Li, Y.; Madesis, P.; Pryor, A.J.; Timmis, J.N. Transfer of plastid DNA to the nucleus is elevated during male gametogenesis in tobacco. Plant Physiol. 2008, 148, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, H.L. The hows and whys of cytoplasmic inheritance in seed plants. Am. J. Bot. 1996, 83, 383–404. [Google Scholar] [CrossRef]
- Wang, D.; Gu, J.B.; David, R.; Wang, Z.; Yang, S.T.; Searle, I.R.; Zhu, J.K.; Timmis, J.N. Experimental reconstruction of double-stranded break repair-mediated plastid DNA insertion into the tobacco nucleus. Plant J. 2018, 93, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, W.M.; George, M.; Wilson, A.C. Rapid evolution of animal mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1979, 76, 1967–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, W.M.; Prager, E.M.; Wang, A.; Wilson, A.C. Mitochondrial DNA sequences of primates: Tempo and mode of evolution. J. Mol. Evol. 1982, 18, 225–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag-Liautard, C.; Coffey, N.; Houle, D.; Lynch, M.; Charlesworth, B.; Keightley, P.D. Direct estimation of the mitochondrial DNA mutation rate in Drosophila melanogaster. PLoS Biol. 2008, 6, e204. [Google Scholar] [CrossRef] [Green Version]
- Perna, N.T.; Kocher, T.D. Mitochondrial DNA: Molecular fossils in the nucleus. Curr. Biol. 1996, 6, 128–129. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.X.; Hewitt, G.M. Nuclear integrations: Challenges for mitochondrial DNA markers. Trends Ecol. Evol. 1996, 11, 247–251. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [Green Version]
- Middleton, C.P.; Senerchia, N.; Stein, N.; Akhunov, E.D.; Keller, B.; Wicker, T.; Kilian, B. Sequencing of chloroplast genomes from wheat, barley, rye and their relatives provides a detailed insight into the evolution of the Triticeae tribe. PLoS ONE 2014, 9, e85761. [Google Scholar] [CrossRef] [Green Version]
- Ossowski, S.; Schneeberger, K.; Lucas-Lledó, J.I.; Warthmann, N.; Clark, R.M.; Shaw, R.G.; Weigel, D.; Lynch, M. The rate and molecular spectrum of spontaneous mutations in Arabidopsis thaliana. Science 2010, 327, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Unseld, M.; Marienfeld, J.R.; Brandt, P.; Brennicke, A. The mitochondrial genome of Arabidopsis thaliana contains 57 genes in 366,924 nucleotides. Nat. Genet. 1997, 15, 57–61. [Google Scholar] [CrossRef]
- Lin, X.; Kaul, S.; Rounsley, S.; Shea, T.P.; Benito, M.I.; Town, C.D.; Fujii, C.Y.; Mason, T.; Bowman, C.L.; Barnstead, M.; et al. Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana. Nature 1999, 402, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Hazkani-Covo, E.; Sorek, R.; Graur, D. Evolutionary dynamics of large Numts in the human genome: rarity of independent insertions and abundance of post-insertion duplications. J. Mol. Evol. 2003, 56, 169–174. [Google Scholar] [CrossRef] [Green Version]
- VanBuren, R.; Ming, R. Organelle DNA accumulation in the recently evolved papaya sex chromosomes. Mol. Genet. Genom. 2013, 288, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.W.; Callis, J. Recent stable insertion of mitochondrial-DNA into an Arabidopsis polyubiquitin gene by nonhomologous recombination. Plant Cell 1993, 5, 97–107. [Google Scholar]
- Ullrich, H.; Lättig, K.; Brennicke, A.; Knoop, V. Mitochondrial DNA variations and nuclear RFLPs reflect different genetic similarities among 23 Arabidopsis thaliana ecotypes. Plant Mol. Biol. 1997, 33, 37–45. [Google Scholar] [CrossRef]
- Kuo, H.F.; Olsen, K.M.; Richards, E.J. Natural variation in a subtelomeric region of Arabidopsis: Implications for the genomic dynamics of a chromosome end. Genetics 2005, 173, 401–417. [Google Scholar] [CrossRef] [Green Version]
- Lough, A.N.; Roark, L.M.; Kato, A.; Ream, T.S.; Lamb, J.C.; Birchler, J.A.; Newton, K.J. Mitochondrial DNA transfer to the nucleus generates extensive insertion site variation in maize. Genetics 2008, 178, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Ricchetti, M.; Fairhead, C.; Dujon, B. Mitochondrial DNA repairs double-strand breaks in yeast chromosomes. Nature 1999, 402, 96–100. [Google Scholar] [CrossRef]
- Blanchard, J.L.; Schmidt, G.W. Pervasive migration of organellar DNA to the nucleus in plants. J. Mol. Evol. 1995, 41, 397–406. [Google Scholar] [CrossRef]
- Maiti, A.K.; Jorissen, M.; Bouvagnet, P. Isolation, in silico characterization and chromosomal localization of a group of cDNAs from ciliated epithelial cells after in vitro ciliogenesis. Genome Biol. 2001, 2. [Google Scholar] [CrossRef]
- Ott, R.W.; Chua, N.H. Enhancer sequences from Arabidopsis thaliana obtained by library transformation of Nicotiana tabacum. Mol. Genet. Genom. 1990, 223, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Kejnovsky, E.; Kubat, Z.; Hobza, R.; Lengerova, M.; Sato, S.; Tabata, S.; Fukui, K.; Matsunaga, S.; Vyskot, B. Accumulation of chloroplast DNA sequences on the Y chromosome of Silene latifolia. Genetica 2006, 128, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Steflova, P.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Strong accumulation of chloroplast DNA in the Y chromosomes of Rumex acetosa and Silene latifolia. Cytogenet. Genome Res. 2014, 142, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Bellot, S.; Fuchs, J.; Houben, A.; Renner, S.S. Analysis of transposable elements and organellar DNA in male and female genomes of a species with a huge Y chromosome reveals distinct Y centromeres. Plant J. 2016, 88, 387–396. [Google Scholar] [CrossRef]
- Ming, R.; Bendahmane, A.; Renner, S.S. Sex chromosomes in land plants. Annu. Rev. Plant Biol. 2011, 62, 485–514. [Google Scholar] [CrossRef] [Green Version]
- Bergero, R.; Charlesworth, D. The evolution of restricted recombination in sex chromosomes. Trends Ecol. Evol. 2009, 24, 94–102. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Kejnovsky, E.; Marquioni, V.; Poltronieri, J.; Molina, W.F.; Diniz, D.; Bertollo, L.A.C. The key role of repeated DNAs in sex chromosome evolution in two fish species with ZW sex chromosome system. Mol. Cytogenet. 2012, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Woischnik, M.; Moraes, C.T. Pattern of organization of human mitochondrial pseudogenes in the nuclear genome. Genome Res. 2002, 12, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Nugent, J.M.; Palmer, J.D. RNA-mediated transfer of the gene coxII from the mitochondrion to the nucleus during flowering plant evolution. Cell 1991, 66, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Shafer, K.S.; Hanekamp, T.; White, K.H.; Thorsness, P.E. Mechanisms of mitochondrial DNA escape to the nucleus in the yeast Saccharomyces cerevisiae. Curr. Genet. 1999, 36, 183–194. [Google Scholar] [CrossRef]
- Adams, K.L.; Daley, D.O.; Qiu, Y.L.; Whelan, J.; Palmer, J.D. Repeated, recent and diverse transfers of a mitochondrial gene to the nucleus in flowering plants. Nature 2000, 408, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, L.; Brennicke, A.; Schuster, W. The mitochondrial gene encoding ribosomal protein S12 has been translocated to the nuclear genome in Oenothera. Nucleic Acids Res. 1992, 20, 5641–5646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Gabriel, A. Patching broken chromosomes with extranuclear cellular DNA. Mol. Cell 1999, 4, 873–881. [Google Scholar] [CrossRef]
- Shin, D.S.; Chahwan, C.; Huffman, J.L.; Tainer, J.A. Structure and function of the double-strand break repair machinery. DNA Repair 2004, 3, 863–873. [Google Scholar] [CrossRef]
- Van Gent, D.C.; Hoeijmakers, J.H.J.; Kanaar, R. Chromosomal stability and the DNA double-stranded break connection. Nat. Rev. Genet. 2001, 2, 196–206. [Google Scholar] [CrossRef]
- Puchta, H. The repair of double-strand breaks in plants: Mechanisms and consequences for genome evolution. J. Exp. Bot. 2005, 56, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hazkani-Covo, E.; Covo, S. Numt-mediated double-strand break repair mitigates deletions during primate genome evolution. PLoS Genet. 2008, 4, e1000237. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, J.L.; Schmidt, G.W. Mitochondrial DNA migration events in yeast and humans: Integration by a common end-joining mechanism and alternative perspectives on nucleotide substitution patterns. Mol. Biol. Evol. 1996, 13, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Ricchetti, M.; Tekaia, F.; Dujon, B. Continued colonization of the human genome by mitochondrial DNA. PLoS Biol. 2004, 2, e273. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.-J.; Dong, R.; Lan, L.-N.; Li, S.-F.; Gao, W.-J.; Niu, H.-X. Nuclear Integrants of Organellar DNA Contribute to Genome Structure and Evolution in Plants. Int. J. Mol. Sci. 2020, 21, 707. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030707
Zhang G-J, Dong R, Lan L-N, Li S-F, Gao W-J, Niu H-X. Nuclear Integrants of Organellar DNA Contribute to Genome Structure and Evolution in Plants. International Journal of Molecular Sciences. 2020; 21(3):707. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030707
Chicago/Turabian StyleZhang, Guo-Jun, Ran Dong, Li-Na Lan, Shu-Fen Li, Wu-Jun Gao, and Hong-Xing Niu. 2020. "Nuclear Integrants of Organellar DNA Contribute to Genome Structure and Evolution in Plants" International Journal of Molecular Sciences 21, no. 3: 707. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030707