Chromium-Induced Reactive Oxygen Species Accumulation by Altering the Enzymatic Antioxidant System and Associated Cytotoxic, Genotoxic, Ultrastructural, and Photosynthetic Changes in Plants



Abstract
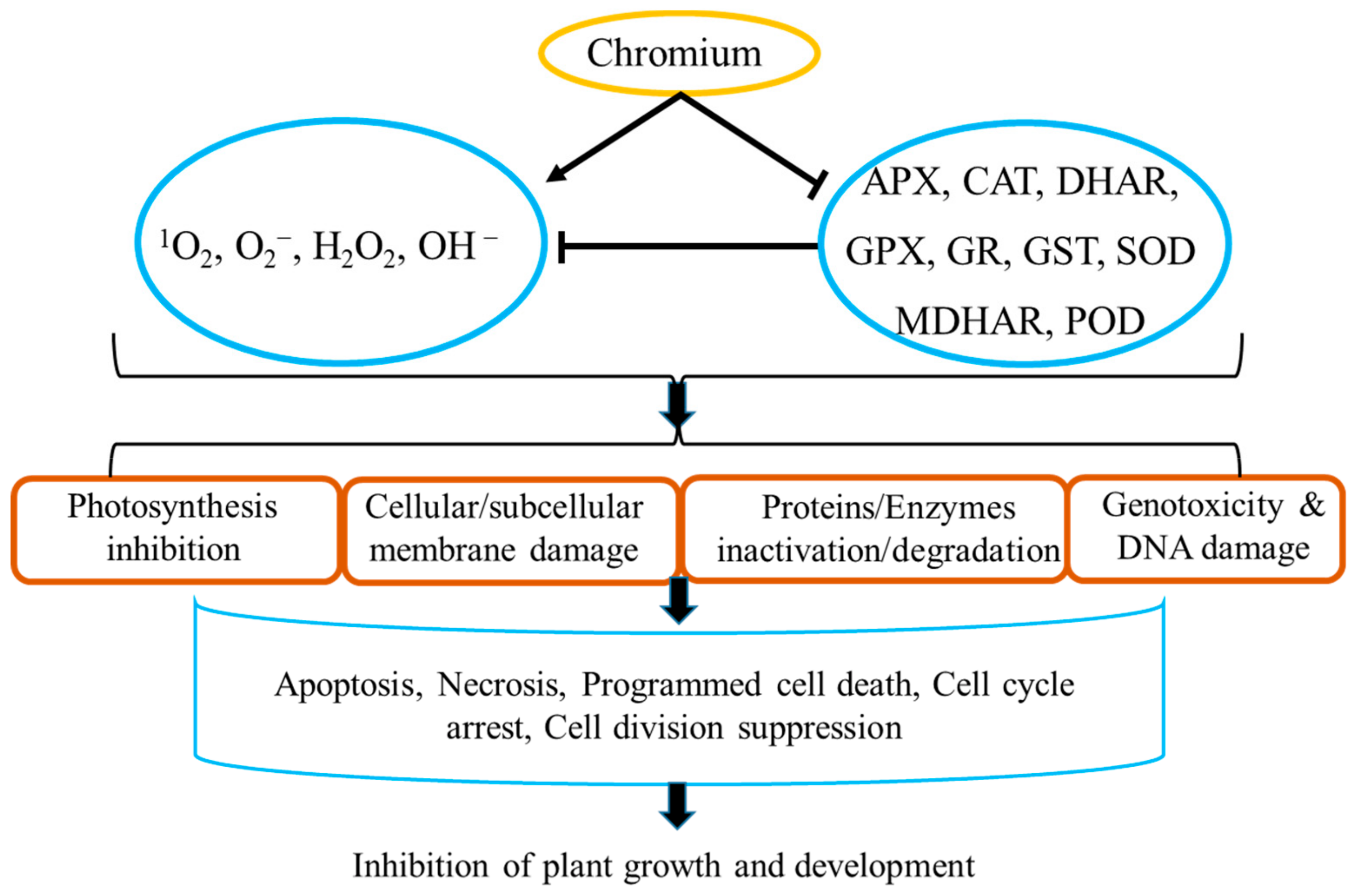
:1. Introduction
2. Chromium-Induced Oxidative Stress in Plants
3. Chromium-Mediated Alteration in the Enzymatic Antioxidant System
4. Chromium-Induced Lipid Peroxidation
5. Chromium-INDUCED DNA DAMAGE and Genotoxicity
6. Chromium-Induced Ultrastructural Changes
6.1. Cr-Induced Necrosis and Cellular Injury
6.2. Electron-Dense Material Deposition in the Subcellular Compartments
7. Chromium-Mediated Changes in Photosynthesis and Photosynthetic Apparatus
8. Strategies to Overcome Cr-Uptake and Phytotoxicity
9. Conclusions
Funding
Conflicts of Interest
References
- Sinha, V.; Pakshirajan, K.; Chaturvedi, R. Chromium tolerance, bioaccumulation and localization in plants: An overview. J. Environ. Manag. 2018, 206, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, N.K.; Dumat, C.; Rashid, M.I. Chromium speciation, bioavailability, uptake, toxicity and detoxification in soil-plant system: A review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, A.; Bibi, I.; Niazi, N.K.; Ok, Y.S.; Murtaza, G.; Shahid, M.; Kunhikrishnan, A.; Li, D.; Mahmood, T. Chromium(VI) sorption efficiency of acid-activated banana peel over organo-montmorillonite in aqueous solutions. Int. J. Phytoremediat. 2017, 19, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Sultana, M.-Y.; Akratos, C.S.; Pavlou, S.; Vayenas, D.V. Chromium removal in constructed wetlands: A review. Int. Biodeterior. Biodegrad. 2014, 96, 181–190. [Google Scholar] [CrossRef]
- Mandiwana, K.L.; Panichev, N.; Kataeva, M.; Siebert, S. The solubility of Cr(III) and Cr(VI) compounds in soil and their availability to plants. J. Hazard. Mater. 2007, 147, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.; Agrawal, M.; Singh, S. Ecological risk assessment of soil contamination by trace elements around coal mining area. J. Soils Sediments 2016, 16, 159–168. [Google Scholar] [CrossRef]
- Ghosh, I.; Ghosh, M.; Mukherjee, A. Remediation of Mine Tailings and Fly Ash Dumpsites: Role of Poaceae Family Members and Aromatic Grasses. In Enhancing Cleanup of Environmental Pollutants; Springer: Cham, Germany, 2017; pp. 117–167. [Google Scholar]
- Wakeel, A.; Ali, I.; Wu, M.; Kkan, A.R.; Jan, M.; Ali, A.; Liu, Y.; Ge, S.; Wu, J.; Gan, Y. Ethylene mediates dichromate-induced oxidative stress and regulation of the enzymatic antioxidant system-related transcriptome in Arabidopsis thaliana. Environ. Exp. Bot. 2019, 161, 166–179. [Google Scholar] [CrossRef]
- Elsawy, E.E.T.; El-Hebeary, M.R.; El Mahallawi, I.S.E. Effect of manganese, silicon and chromium additions on microstructure and wear characteristics of grey cast iron for sugar industries applications. Wear 2017, 390–391, 113–124. [Google Scholar] [CrossRef]
- Florea, C.D.; Carcea, I.; Cimpoesu, R.; Toma, S.L.; Sandu, I.G.; Bejinariu, C. Experimental Analysis of Resistance to Electrocorosion of a High Chromium Cast Iron with Applications in the Vehicle Industry. Rev. Chim. 2017, 68, 2397–2401. [Google Scholar]
- Bharagava, R.N.; Mishra, S. Hexavalent chromium reduction potential of Cellulosimicrobium sp isolated from common effluent treatment plant of tannery industries. Ecotoxicol. Environ. Saf. 2018, 147, 102–109. [Google Scholar] [CrossRef]
- Byrne, P.; Taylor, K.G.; Hudson-Edwards, K.A.; Barrett, J.E.S. Speciation and potential long-term behaviour of chromium in urban sediment particulates. J. Soils Sediments 2017, 17, 2666–2676. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhu, G.; Pan, H.; Shi, P.; Li, J.; Liu, Y.; Tong, H. Surface dust heavy metals in the major cities, China. Environ. Earth Sci. 2017, 76, 757. [Google Scholar]
- Wakeel, A.; Ali, I.; Upreti, S.; Ullah, A.; Liu, B.; Khan, A.R.; Huang, L.; Wu, M.; Gan, Y. Ethylene mediates dichromate induced inhibition of primary root growth by altering AUX1 expression and auxin accumulation in Arabidopsis thaliana. Plant Cell Environ. 2018, 41, 1453–1467. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Wang, H.; Song, J.; Cao, X.; Rogers, H.J.; Francis, D.; Jia, C.; Sun, L.; Hou, M.; Yang, Y.; et al. Cell cycle arrest mediated by Cd-induced DNA damage in Arabidopsis root tips. Ecotoxicol. Environ. Saf. 2017, 145, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Gielen, H.; Vangronsveld, J.; Cuypers, A. Cd-induced Cu deficiency responses in Arabidopsis thaliana: Are phytochelatins involved? Plant Cell Environ. 2017, 40, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Al Mahmud, J.; Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Hossain, M.S.; Fujita, M. Maleic acid assisted improvement of metal chelation and antioxidant metabolism confers chromium tolerance in Brassica juncea L. Ecotoxicol. Environ. Saf. 2017, 144, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Dumat, C.; Khalid, S.; Niazi, N.K.; Antunes, P.M.C. Cadmium Bioavailability, Uptake, Toxicity and Detoxification in Soil-Plant System. In Reviews of Environmental Contamination and Toxicology; DeVoogt, P., Ed.; Springer: Cham, Germany, 2017; Volume 241, pp. 73–137. [Google Scholar]
- Abbas, G.; Murtaza, B.; Bibi, I.; Shahid, M.; Niazi, N.K.; Khan, M.I.; Amjad, M.; Hussain, M.; Natasha. Arsenic Uptake, Toxicity, Detoxification, and Speciation in Plants: Physiological, Biochemical, and Molecular Aspects. Int. J. Environ. Res. Public Health 2018, 15, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Wu, K.; Tang, Z.; Guo, Q.; Guo, X.; Wang, H. Exogenous ethylene enhanced the cadmium resistance and changed the alkaloid biosynthesis in Catharanthus roseus seedlings. Acta Physiol. Plant. 2017, 39, 267. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.N.; Fujita, M. Hydrogen Peroxide Pretreatment Mitigates Cadmium-Induced Oxidative Stress in Brassica napus L.: An Intrinsic Study on Antioxidant Defense and Glyoxalase Systems. Front. Plant Sci. 2017, 8, 115. [Google Scholar] [CrossRef]
- Balasaraswathi, K.; Jayaveni, S.; Sridevi, J.; Sujatha, D.; Aaron, K.P.; Rose, C. Cr-induced cellular injury and necrosis in Glycine max L.: Biochemical mechanism of oxidative damage in chloroplast. Plant Physiol. Biochem. 2017, 118, 653–666. [Google Scholar] [CrossRef]
- Eleftheriou, E.P.; Adamakis, I.-D.S.; Panteris, E.; Fatsiou, M. Chromium-Induced Ultrastructural Changes and Oxidative Stress in Roots of Arabidopsis thaliana. Int. J. Mol. Sci. 2015, 16, 15852–15871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farid, M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Nasir, T.; Abbasi, G.H.; Rehmani, M.I.A.; Ata-Ul-Karim, S.T.; Bukhari, S.A.H.; et al. Phyto-management of chromium contaminated soils through sunflower under exogenously applied 5-aminolevulinic acid. Ecotoxicol. Environ. Saf. 2018, 151, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.; Ali, S.; Akram, N.A.; Rizwan, M.; Abbas, F.; Bukhari, S.A.H.; Saeed, R. Phyto-management of Cr-contaminated soils by sunflower hybrids: Physiological and biochemical response and metal extractability under Cr stress. Environ. Sci. Pollut. Res. 2017, 24, 16845–16859. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.; Ali, S.; Rizwan, M.; Ali, Q.; Abbas, F.; Bukhari, S.A.H.; Saeed, R.; Wu, L. Citric acid assisted phytoextraction of chromium by sunflower; morpho-physiological and biochemical alterations in plants. Ecotoxicol. Environ. Saf. 2017, 145, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Kharbech, O.; Houmani, H.; Chaoui, A.; Corpas, F.J. Alleviation of Cr(VI)-induced oxidative stress in maize (Zea mays L.) seedlings by NO and H2S donors through differential organ-dependent regulation of ROS and NADPH-recycling metabolisms. J. Plant Physiol. 2017, 219, 71–80. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Khan, I.; Tanveer, M.; Shahid, M.; Shakoor, A.; Wang, L. Phyto-Toxicity of Chromium in Maize: Oxidative Damage, Osmolyte Accumulation, Anti-Oxidative Defense and Chromium Uptake. Pedosphere 2017, 27, 262–273. [Google Scholar] [CrossRef]
- Singh, V.P.; Kumar, J.; Singh, M.; Singh, S.; Prasad, S.M.; Dwivedi, R.; Singh, M.P.V.V.B. Role of salicylic acid-seed priming in the regulation of chromium(VI) and UV-B toxicity in maize seedlings. Plant Growth Regul. 2016, 78, 79–91. [Google Scholar] [CrossRef]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Riaz, M.; Shahzad, S.M.; Imran, Q.; Ali, I. Combined ability of chromium(Cr) tolerant plant growth promoting bacteria (PGPB), and salicylic acid (SA) in attenuation of chromium stress in maize plants. Plant Physiol. Biochem. 2016, 108, 456–467. [Google Scholar] [CrossRef]
- Singh, S.; Srivastava, P.K.; Kumar, D.; Tripathi, D.K.; Chauhan, D.K.; Prasad, S.M. Morpho-anatomical and biochemical adapting strategies of maize (Zea mays L.) seedlings against lead and chromium stresses. Biocatal. Agric. Biotechnol. 2015, 4, 286–295. [Google Scholar] [CrossRef]
- Maiti, S.; Ghosh, N.; Mandal, C.; Das, K.; Dey, N.; Adak, M.K. Responses of the maize plant to chromium stress with reference to antioxidation activity. Braz. J. Plant Physiol. 2012, 24, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Handa, N.; Kohli, S.K.; Thukral, A.K.; Arora, S.; Bhardwaj, R. Role of Se(VI) in counteracting oxidative damage in Brassica juncea L. under Cr(VI) stress. Acta Physiol. Plant. 2017, 39, 51. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, X.; Liu, Y.; Wei, J.; Shen, W.; Shen, Z.; Cui, J. Hemin-mediated alleviation of zinc, lead and chromium toxicity is associated with elevated photosynthesis, antioxidative capacity; suppressed metal uptake and oxidative stress in rice seedlings. Plant Growth Regul. 2017, 81, 253–264. [Google Scholar] [CrossRef]
- Ma, J.; Lv, C.; Xu, M.; Chen, G.; Lv, C.; Gao, Z. Photosynthesis performance, antioxidant enzymes, and ultrastructural analyses of rice seedlings under chromium stress. Environ. Sci. Pollut. Res. 2016, 23, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Wu, X.; Qiu, B.; Wu, F.; Jiang, L.; Zhang, G. Physiological and proteomic alterations in rice (Oryza sativa L.) seedlings under hexavalent chromium stress. Planta 2014, 240, 291–308. [Google Scholar] [CrossRef]
- Ngoc-Nam, T.; Huang, T.-L.; Chi, W.-C.; Fu, S.-F.; Chen, C.-C.; Huang, H.-J. Chromium stress response effect on signal transduction and expression of signaling genes in rice. Physiol. Plant. 2014, 150, 205–224. [Google Scholar]
- Bashri, G.; Parihar, P.; Singh, R.; Singh, S.; Singh, V.P.; Prasad, S.M. Physiological and biochemical characterization of two Amaranthus species under Cr(VI) stress differing in Cr(VI) tolerance. Plant Physiol. Biochem. 2016, 108, 12–23. [Google Scholar] [CrossRef]
- Scoccianti, V.; Bucchini, A.E.; Iacobucci, M.; Ruiz, K.B.; Biondi, S. Oxidative stress and antioxidant responses to increasing concentrations of trivalent chromium in the Andean crop species Chenopodium quinoa Willd. Ecotoxicol. Environ. Saf. 2016, 133, 25–35. [Google Scholar] [CrossRef]
- Tripathi, A.; Tripathi, D.K.; Chauhan, D.K.; Kumar, N. Chromium(VI)-induced phytotoxicity in river catchment agriculture: Evidence from physiological, biochemical and anatomical alterations in Cucumis sativus (L.) used as model species. Chem. Ecol. 2016, 32, 12–33. [Google Scholar] [CrossRef]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef]
- Gill, R.A.; Zhang, N.; Ali, B.; Farooq, M.A.; Xu, J.; Gill, M.B.; Mao, B.; Zhou, W. Role of exogenous salicylic acid in regulating physio-morphic and molecular changes under chromium toxicity in black- and yellow-seeded Brassica napus L. Environ. Sci. Pollut. Res. 2016, 23, 20483–20496. [Google Scholar] [CrossRef]
- Qing, X.; Zhao, X.; Hu, C.; Wang, P.; Zhang, Y.; Zhang, X.; Wang, P.; Shi, H.; Jia, F.; Qu, C. Selenium alleviates chromium toxicity by preventing oxidative stress in cabbage (Brassica campestris L. ssp. Pekinensis) leaves. Ecotoxicol. Environ. Saf. 2015, 114, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K. Silicon nanoparticles (SiNp) alleviate chromium(VI) phytotoxicity in Pisum sativum (L.) seedlings. Plant Physiol. Biochem. 2015, 96, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, A.R.; Achary, V.M.M.; Panda, B.B. Chromium(VI)-induced hormesis and genotoxicity are mediated through oxidative stress in root cells of Allium cepa L. Plant Growth Regul. 2013, 71, 157–170. [Google Scholar] [CrossRef]
- Kovacik, J.; Babula, P.; Klejdus, B.; Hedbavny, J. Chromium Uptake and Consequences for Metabolism and Oxidative Stress in Chamomile Plants. J. Agric. Food Chem. 2013, 61, 7864–7873. [Google Scholar] [CrossRef]
- Eleftheriou, E.P.; Adamakis, I.-D.S.; Fatsiou, M.; Panteris, E. Hexavalent chromium disrupts mitosis by stabilizing microtubules in Lens culinaris root tip cells. Physiol. Plant. 2013, 147, 169–180. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Kanwar, M.; Bhardwaj, R.; Yu, J.-Q.; Lam-Son Phan, T. Chromium Stress Mitigation by Polyamine-Brassinosteroid Application Involves Phytohormonal and Physiological Strategies in Raphanus sativus L. PLoS ONE 2012, 7, e33210. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, R.; Panda, S.K. Influence of Chromium Salts on Increased Lipid Peroxidation and Differential Pattern in Antioxidant Metabolism in Pistia stratiotes L. Braz. Arch. Biol. Technol. 2010, 53, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Zhang, M.; Smith, J.A.C.; Harberd, N.P.; Jiang, C. The regulatory roles of ethylene and reactive oxygen species (ROS) in plant salt stress responses. Plant Mol. Biol. 2016, 91, 651–659. [Google Scholar] [CrossRef]
- Pourrut, B.; Shahid, M.; Douay, F.; Dumat, C.; Pinelli, E. Molecular mechanisms involved in lead uptake, toxicity and detoxification in higher plants. In Heavy Metal Stress in Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 121–147. [Google Scholar]
- Shahid, M.; Austruy, A.; Echevarria, G.; Arshad, M.; Sanaullah, M.; Aslam, M.; Nadeem, M.; Nasim, W.; Dumat, C. EDTA-enhanced phytoremediation of heavy metals: A review. Soil Sediment Contam. Int. J. 2014, 23, 389–416. [Google Scholar] [CrossRef]
- Panda, S.K. Chromium-mediated oxidative stress and ultrastructural changes in root cells of developing rice seedlings. J. Plant Physiol. 2007, 164, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Chromium toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Park, S.C.; Yun, B.-W.; Kwak, S.-S. Overexpressing sweetpotato peroxidase gene swpa4 affects nitric oxide production by activating the expression of reactive oxygen species- and nitric oxide-related genes in tobacco. Plant Physiol. Biochem. 2017, 120, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Jan, M.; Wakeel, A.; Azizullah, A.; Liu, B.; Islam, F.; Ali, A.; Daud, M.K.; Liu, Y.; Gan, Y. Biochemical responses and ultrastructural changes in ethylene insensitive mutants of Arabidopsis thialiana subjected to bisphenol A exposure. Ecotoxicol. Environ. Saf. 2017, 144, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, J.A.L.; Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Hossain, M.S.; Fujita, M. gamma-aminobutyric acid (GABA) confers chromium stress tolerance in Brassica juncea L. by modulating the antioxidant defense and glyoxalase systems. Ecotoxicology 2017, 26, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Mar Gil-Diaz, M.; Pinilla, P.; Carmen Lobo, M. Impact of Cr and Zn on Growth, Biochemical and Physiological Parameters, and Metal Accumulation by Wheat and Barley Plants. Water Air Soil Pollut. 2017, 228, 419. [Google Scholar] [CrossRef]
- Ahmad, R.; Ali, S.; Hannan, F.; Rizwan, M.; Iqbal, M.; Hassan, Z.; Akram, N.A.; Maqbool, S.; Abbas, F. Promotive role of 5-aminolevulinic acid on chromium-induced morphological, photosynthetic, and oxidative changes in cauliflower (Brassica oleracea botrytis L.). Environ. Sci. Pollut. Res. 2017, 24, 8814–8824. [Google Scholar] [CrossRef]
- Zhu, X.; Hou, G.; Ru, G.; Zhang, L.; Wu, X.; Liu, G. Accumulation and antioxidant properties of chromium stress in Pennisetum alopecuroides. J. Henan Agric. Univ. 2017, 51, 330–334. [Google Scholar]
- Yilmaz, S.H.; Kaplan, M.; Temizgul, R.; Yilmaz, S. Antioxidant enzyme response of sorghum plant upon exposure to Aluminum, Chromium and Lead heavy metals. Turk. J. Biochem. 2017, 42, 503–512. [Google Scholar] [CrossRef]
- Kanwar, M.K.; Poonam, S.P.; Bhardwaj, R. Involvement of Asada-Halliwell Pathway During Phytoremediation of Chromium(VI) in Brassica juncea L. Plants. Int. J. Phytoremediat. 2015, 17, 1237–1243. [Google Scholar] [CrossRef]
- Singh, M.; Kushwaha, B.K.; Singh, S.; Kumar, V.; Singh, V.P.; Prasad, S.M. Sulphur alters chromium(VI) toxicity in Solarium melongena seedlings: Role of sulphur assimilation and sulphur-containing antioxidants. Plant Physiol. Biochem. 2017, 112, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Ashraf, U.; Khan, I.; Tanveer, M.; Saleem, M.F.; Wang, L. Aluminum and Chromium Toxicity in Maize: Implications for Agronomic Attributes, Net Photosynthesis, Physio-Biochemical Oscillations, and Metal Accumulation in Different Plant Parts. Water Air Soil Pollut. 2016, 227, 326. [Google Scholar] [CrossRef]
- Ding, H.; Zhu, X.; Liu, H.; Chen, Y.; Liang, J. Hexavalent chromium(Cr~(6+)) induced oxidative damage to maize (Zea mays L.) root tip cells. J. Yangzhou Univ. 2011, 32, 28–32. [Google Scholar]
- Ding, H.; Wang, G.; Lou, L.; Lv, J. Physiological responses and tolerance of kenaf (Hibiscus cannabinus L.) exposed to chromium. Ecotoxicol. Environ. Saf. 2016, 133, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Huda, A.K.M.N.; Swaraz, A.M.; Abu Reza, M.; Haque, M.A.; Kabir, A.H. Remediation of Chromium Toxicity Through Exogenous Salicylic Acid in Rice (Oryza sativa L.). Water Air Soil Pollut. 2016, 227, 278. [Google Scholar] [CrossRef]
- Cao, F.; Wang, N.; Zhang, M.; Dai, H.; Dawood, M.; Zhang, G.; Wu, F. Comparative study of alleviating effects of GSH, Se and Zn under combined contamination of cadmium and chromium in rice (Oryza sativa). Biometals 2013, 26, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, N.; Abbas, Z.; Iqbal, M.; Rizwan, M.; Jabbar, A.; Farid, M.; Ali, S.; Ibrahim, M.; Abbas, F. Glycinebetaine mediates chromium tolerance in mung bean through lowering of Cr uptake and improved antioxidant system. Arch. Agron. Soil Sci. 2016, 62, 648–662. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, W.; Wang, Q.; Sun, J. The Sub-cellular Chromium(Cr~(6+))Distribution and Tolerance Mechanism to Chromium Stress in Different Tolerant Brassica chinensis L. Cultivars. Acta Bot. Boreali-Occident. Sin. 2016, 36, 954–963. [Google Scholar]
- Shirong, Z.; Shuyi, L.; Xiaodong, D.; Fangbai, L.; Chuanping, L.; Xinrong, L.; Rongping, W. Silicon mediated the detoxification of Cr on pakchoi (Brassica chinensis L.) in Cr-contaminated soil. J. Food Agric. Environ. 2013, 11, 814–819. [Google Scholar]
- Tian, B.; Zhang, Y.; Zhang, L.; Ma, X.; Jin, Z.; Liu, Z.; Liu, D.; Pei, Y. Effects of cadmium or chromium on growth and NADPH oxidase and antioxidant enzyme system of foxtail millet seedlings. J. Agro-Environ. Sci. 2016, 35, 240–246. [Google Scholar]
- UdDin, I.; Bano, A.; Masood, S. Chromium toxicity tolerance of Solanum nigrum L. and Parthenium hysterophorus L. plants with reference to ion pattern, antioxidation activity and root exudation. Ecotoxicol. Environ. Saf. 2015, 113, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, J.; Kumar, P.; Sharma, P.N.; Tewari, R.K. Chromium toxicity induces oxidative stress in turnip. Indian J. Plant Physiol. 2015, 20, 220–226. [Google Scholar] [CrossRef]
- Afshan, S.; Ali, S.; Bharwana, S.A.; Rizwan, M.; Farid, M.; Abbas, F.; Ibrahim, M.; Mehmood, M.A.; Abbasi, G.H. Citric acid enhances the phytoextraction of chromium, plant growth, and photosynthesis by alleviating the oxidative damages in Brassica napus L. Environ. Sci. Pollut. Res. 2015, 22, 11679–11689. [Google Scholar] [CrossRef] [PubMed]
- Daud, M.K.; Mei, L.; Variath, M.T.; Ali, S.; Li, C.; Rafiq, M.T.; Zhu, S.J. Chromium(VI) Uptake and Tolerance Potential in Cotton Cultivars: Effect on Their Root Physiology, Ultramorphology, and Oxidative Metabolism. Biomed Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Islam, M.K.; Khanam, M.S.; Lee, S.Y.; Alam, I.; Huh, M.R. The interaction of arsenic (As) and chromium (Cr) influences growth and antioxidant status in tossa jute (Corchorus olitorius). Plant Omics 2014, 7, 499–509. [Google Scholar]
- Yildiz, M.; Terzi, H.; Bingul, N. Protective role of hydrogen peroxide pretreatment on defense systems and BnMP1 gene expression in Cr(VI)-stressed canola seedlings. Ecotoxicology 2013, 22, 1303–1312. [Google Scholar] [CrossRef]
- Sayantan, D. Shardendu. Amendment in phosphorus levels moderate the chromium toxicity in Raphanus sativus L. as assayed by antioxidant enzymes activities. Ecotoxicol. Environ. Saf. 2013, 95, 161–170. [Google Scholar] [CrossRef]
- Yildiz, M.; Terzi, H.; Cenkci, S.; Yildiz, B. Chromium(VI)-Induced Alterations in 2-D Protein Profiles and Antioxidant Defence Systems of Barley Cultivars. Hacet. J. Biol. Chem. 2012, 40, 257–265. [Google Scholar]
- Yu, X.-Z.; Lin, Y.-J.; Fan, W.-J.; Lu, M.-R. The role of exogenous proline in amelioration of lipid peroxidation in rice seedlings exposed to Cr(VI). Int. Biodeterior. Biodegrad. 2017, 123, 106–112. [Google Scholar] [CrossRef]
- Smaoui-Jardak, M.; Kriaa, W.; Maalej, M.; Zouari, M.; Kamoun, L.; Trabelsi, W.; Ben Abdallah, F.; Elloumi, N. Effect of the phosphogypsum amendment of saline and agricultural soils on growth, productivity and antioxidant enzyme activities of tomato (Solanum lycopersicum L.). Ecotoxicology 2017, 26, 1089–1104. [Google Scholar] [CrossRef]
- Zeng, F.; Qiu, B.; Wu, X.; Niu, S.; Wu, F.; Zhang, G. Glutathione-Mediated Alleviation of Chromium Toxicity in Rice Plants. Biol. Trace Elem. Res. 2012, 148, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.-R.; Zhao, F.-S.; Qiu, B.-Y.; Ouyang, Y.-N.; Wu, F.-B.; Zhang, G.-P. Alleviation of Chromium Toxicity by Silicon Addition in Rice Plants. Agric. Sci. China 2011, 10, 1188–1196. [Google Scholar] [CrossRef]
- Silvina Aran, D.; Alfredo Harguinteguy, C.; Fernandez-Cirelli, A.; Luisa Pignata, M. Phytoextraction of Pb, Cr, Ni, and Zn using the aquatic plant Limnobium laevigatum and its potential use in the treatment of wastewater. Environ. Sci. Pollut. Res. 2017, 24, 18295–18308. [Google Scholar] [CrossRef] [PubMed]
- Balal, R.M.; Shahid, M.A.; Vincent, C.; Zotarelli, L.; Liu, G.; Mattson, N.S.; Rathinasabapathi, B.; Martinez-Nicolas, J.J.; Garcia-Sanchez, F. Kinnow mandarin plants grafted on tetraploid rootstocks are more tolerant to Cr-toxicity than those grafted on its diploids one. Environ. Exp. Bot. 2017, 140, 8–18. [Google Scholar] [CrossRef]
- Ashfaque, F.; Inam, A.; Inam, A.; Iqbal, S.; Sahay, S. Response of silicon on metal accumulation, photosynthetic inhibition and oxidative stress in chromium-induced mustard (Brassica juncea L.). S. Afr. J. Bot. 2017, 111, 153–160. [Google Scholar] [CrossRef]
- Singh, D.; Agnihotri, A.; Seth, C.S. Interactive effects of EDTA and oxalic acid on chromium uptake, translocation and photosynthetic attributes in Indian mustard (Brassica juncea L. var. Varuna). Curr. Sci. 2017, 112, 2034–2042. [Google Scholar] [CrossRef]
- Sinha, V.; Pakshirajan, K.; Manikandan, N.A.; Chaturvedi, R. Kinetics, biochemical and factorial analysis of chromium uptake in a multi-ion system by Tradescantia pallida (Rose) D. R. Hunt. Int. J. Phytoremediat. 2017, 19, 1007–1016. [Google Scholar] [CrossRef]
- De Oliveira, L.M.; Gress, J.; De, J.; Rathinasabapathi, B.; Marchi, G.; Chen, Y.; Ma, L.Q. Sulfate and chromate increased each other’s uptake and translocation in As-hyperaccumulator Pteris vittata. Chemosphere 2016, 147, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.; Singh, S.P.; Singh, A.; Singh, S.; Tripathi, P.; Chandra, A.; Solomon, S. Study on physio-biochemical attributes and metallothionein gene expression affected by chromium(VI) in sugarcane (Saccharum spp. hybrid). J. Environ. Biol. 2016, 37, 375–382. [Google Scholar]
- Terzi, H.; Yildiz, M. Interactive effects of sulfur and chromium on antioxidative defense systems and BnMP1 gene expression in canola (Brassica napus L.) cultivars differing in Cr(VI) tolerance. Ecotoxicology 2015, 24, 1171–1182. [Google Scholar] [CrossRef]
- Terzi, H.; Yildiz, M. Variations in Chromium Tolerance and Accumulation among Canola (Brassica napus L.) Cultivars. Bull. Environ. Contam. Toxicol. 2014, 93, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Shukla, A.K.; Sharma, Y.K. Amelioration of Heavy-Metal Toxicity in Cauliflower by Application of Salicylic Acid. Commun. Soil Sci. Plant Anal. 2015, 46, 1309–1319. [Google Scholar] [CrossRef]
- Prado, C.; Prado, F.E.; Pagano, E.; Rosa, M. Differential Effects of Cr(VI) on the Ultrastructure of Chloroplast and Plasma Membrane of Salvinia minima Growing in Summer and Winter. Relationships with Lipid Peroxidation, Electrolyte Leakage, Photosynthetic Pigments, and Carbohydrates. Water Air Soil Pollut. 2015, 226, 8. [Google Scholar] [CrossRef]
- Sinha, V.; Pakshirajan, K.; Chaturvedi, R. Chromium(VI) Accumulation and Tolerance by Tradescantia pallida: Biochemical and Antioxidant Study. Appl. Biochem. Biotechnol. 2014, 173, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Du, W.; Cai, H.; Jia, Z.; Dong, H. Trivalent chromium pretreatment alleviates the toxicity of oxidative damage in wheat plants exposed to hexavalent chromium. Acta Physiol. Plant. 2014, 36, 787–794. [Google Scholar] [CrossRef]
- Sharmin, S.A.; Alam, I.; Kim, K.-H.; Kim, Y.-G.; Kim, P.J.; Bahk, J.D.; Lee, B.-H. Chromium-induced physiological and proteomic alterations in roots of Miscanthus sinensis. Plant Sci. 2012, 187, 113–126. [Google Scholar] [CrossRef]
- Pesko, M.; Kral’ova, K.; Blasko, J. Phytotoxic Effects of trivalent Chromium on Rapeseed Plants. Fresenius Environ. Bull. 2012, 21, 761–768. [Google Scholar]
- Thompson, C.M.; Fedorov, Y.; Brown, D.D.; Suh, M.; Proctor, D.M.; Kuriakose, L.; Haws, L.C.; Harris, M.A. Assessment of Cr(VI)-Induced Cytotoxicity and Genotoxicity Using High Content Analysis. PLoS ONE 2012, 7, e42720. [Google Scholar] [CrossRef]
- Wakeman, T.P.; Yang, A.; Dalal, N.S.; Boohaker, R.J.; Zeng, Q.; Ding, Q.; Xu, B. DNA mismatch repair protein Mlh1 is required for tetravalent chromium intermediate-induced DNA damage. Oncotarget 2017, 8, 83975–83985. [Google Scholar] [CrossRef] [Green Version]
- Chiu, A.; Shi, X.L.; Lee, W.K.P.; Hill, R.; Wakeman, T.P.; Katz, A.; Xu, B.; Dalal, N.S.; Robertson, J.D.; Chen, C.; et al. Review of Chromium (VI) Apoptosis, Cell-Cycle-Arrest, and Carcinogenesis. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2010, 28, 188–230. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.L.; LaCerte, C.; Wise, S.S.; Holmes, A.; Martino, J.; Wise, J.P., Jr.; Thompson, W.D.; Wise, J.P., Sr. Comparative cytotoxicity and genotoxicity of particulate and soluble hexavalent chromium in human and sperm whale (Physeter macrocephalus) skin cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Z.; Zhao, M.; Zhen, H.; Chen, L.; Shi, P.; Huang, Z. Genotoxicity of Tri- and Hexavalent Chromium Compounds In Vivo and Their Modes of Action on DNA Damage In Vitro. PLoS ONE 2014, 9, e103194. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Hu, J.; Zhang, B.; Li, Y.; Wise, J.P., Sr.; Bassig, B.A.; Zhou, A.; Savitz, D.A.; Xiong, C.; Zhao, J.; et al. A case-control study of maternal exposure to chromium and infant low birth weight in China. Chemosphere 2016, 144, 1484–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasiak, J.; Kowalik, J. A comparison of the in vitro genotoxicity of tri- and hexavalent chromium. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2000, 469, 135–145. [Google Scholar] [CrossRef]
- Meng, X.; Fang, Z.; Wei, H.; Yang, Z.; Huang, Z. Genotoxicity investigation of tri-and hexavalent chromium using a yeast SUP4-o mutation assay. Carcinog. Teratog. Mutagenesis 2017, 29, 65–69. [Google Scholar]
- Salnikow, K.; Zhitkovich, A. Genetic and epigenetic mechanisms in metal carcinogenesis and cocarcinogenesis: Nickel, arsenic, and chromium. Chem. Res. Toxicol. 2008, 21, 28–44. [Google Scholar] [CrossRef] [Green Version]
- Alimba, C.G.; Dhillon, V.; Bakare, A.A.; Fenech, M. Genotoxicity and cytotoxicity of chromium, copper, manganese and lead, and their mixture in WIL2-NS human B lymphoblastoid cells is enhanced by folate depletion. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2016, 798, 35–47. [Google Scholar] [CrossRef]
- Kumari, V.; Yadav, A.; Haq, I.; Kumar, S.; Bharagava, R.N.; Singh, S.K.; Raj, A. Genotoxicity evaluation of tannery effluent treated with newly isolated hexavalent chromium reducing Bacillus cereuss. J. Environ. Manag. 2016, 183, 204–211. [Google Scholar] [CrossRef]
- Bordeerat, N.K. Evaluation of genotoxicity in Welders exposed to hexavalent chromium. Environ. Mol. Mutagenesis 2017, 58, S65–S66. [Google Scholar]
- Taju, G.; Majeed, S.A.; Nambi, K.S.N.; Hameed, A.S.S. Application of fish cell lines for evaluating the chromium induced cytotoxicity, genotoxicity and oxidative stress. Chemosphere 2017, 184, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Zhong, C.; Xiao, F. Cr(VI) induces premature senescence through ROS-mediated p53 pathway in L-02 hepatocytes. Sci. Rep. 2016, 6, 34578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim Linh, N.; Hoang Anh, N.; Richter, O.; Minh Thinh, P.; Van Phuoc, N. Ecophysiological responses of young mangrove species Rhizophora apiculata (Blume) to different chromium contaminated environments. Sci. Total Environ. 2017, 574, 369–380. [Google Scholar]
- Suraweera, D.D.; Groom, T.; Taylor, P.W.J.; Jayasinghe, C.S.; Nicolas, M.E. Dynamics of flower, achene and trichome development governs the accumulation of pyrethrins in pyrethrum (Tanacetum cinerariifolium) under irrigated and dryland conditions. Ind. Crop. Prod. 2017, 109, 123–133. [Google Scholar] [CrossRef]
- Vadde, B.V.L.; Challa, K.R.; Nath, U. The TCP4 transcription factor regulates trichome cell differentiation by directly activating GLABROUS INFLORESCENCE STEMS in Arabidopsis thaliana. Plant J. 2018, 93, 259–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speranza, A.; Ferri, P.; Battistelli, M.; Falcieri, E.; Crinelli, R.; Scoccianti, V. Both trivalent and hexavalent chromium strongly alter in vitro germination and ultrastructure of kiwifruit pollen. Chemosphere 2007, 66, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Mangabeira, P.A.; Ferreira, A.S.; de Almeida, A.-A.F.; Fernandes, V.F.; Lucena, E.; Souza, V.L.; dos Santos Junior, A.J.; Oliveira, A.H.; Grenier-Loustalot, M.F.; Barbier, F.; et al. Compartmentalization and ultrastructural alterations induced by chromium in aquatic macrophytes. Biometals 2011, 24, 1017–1026. [Google Scholar] [CrossRef]
- Gill, R.A.; Ali, B.; Yang, S.; Tong, C.; Islam, F.; Gill, M.B.; Mwamba, T.M.; Ali, S.; Mao, B.; Liu, S.; et al. Reduced Glutathione Mediates Phone-Ultrastrcture, Kinome and Transportome in Chromium-Induced Brassica napus L. Front. Plant Sci. 2017, 8, 2037. [Google Scholar] [CrossRef] [Green Version]
- Andosch, A.; Hoeftberger, M.; Luetz, C.; Luetz-Meindl, U. Subcellular Sequestration and Impact of Heavy Metals on the Ultrastructure and Physiology of the Multicellular Freshwater Alga Desmidium swartzii. Int. J. Mol. Sci. 2015, 16, 10389–10410, Erratum in 2015, 16, 20239.. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, S.A.H.; Zheng, W.; Xie, L.; Zhang, G.; Shang, S.; Wu, F. Cr-induced changes in leaf protein profile, ultrastructure and photosynthetic traits in the two contrasting tobacco genotypes. Plant Growth Regul. 2016, 79, 147–156. [Google Scholar] [CrossRef]
- Bukhari, S.A.H.; Wang, R.; Wang, W.; Ahmed, I.M.; Zheng, W.; Cao, F. Genotype-dependent effect of exogenous 24-epibrassinolide on chromium-induced changes in ultrastructure and physicochemical traits in tobacco seedlings. Environ. Sci. Pollut. Res. 2016, 23, 18229–18238. [Google Scholar] [CrossRef]
- El Fels, L.; Hafidi, M.; Silvestre, J.; Kallerhoff, J.; Merlina, G.; Pinelli, E. Efficiency of co-composting process to remove genotoxicity from sewage sludge contaminated with hexavalent chromium. Ecol. Eng. 2015, 82, 355–360. [Google Scholar]
- Fargasova, A. Plants as models for chromium and nickel risk assessment. Ecotoxicology 2012, 21, 1476–1483. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Chauhan, L.K.S.; Pande, P.N.; Gupta, S.K. Cytogenetic effects of leachates from tannery solid waste on the somatic cells of Vicia faba. Environ. Toxicol. 2004, 19, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.; Azevedo, R.; Fernandes, P.; Santos, C. Cr(VI) Induces DNA Damage, Cell Cycle Arrest and Polyploidization: A How Cytometric and Comet Assay Study in Pisum sativum. Chem. Res. Toxicol. 2011, 24, 1040–1047. [Google Scholar] [CrossRef]
- Hemachandra, C.K.; Pathiratne, A. Assessing Toxicity of Copper, Cadmium and Chromium Levels Relevant to Discharge Limits of Industrial Effluents into Inland Surface Waters Using Common Onion, Allium cepa Bioassay. Bull. Environ. Contam. Toxicol. 2015, 94, 199–203. [Google Scholar] [CrossRef]
- Qian, X.-W. Mutagenic effects of chromium trioxide on root tip cells of Vicia faba. J. Zhejiang Univ. Sci. 2004, 5, 1570–1576. [Google Scholar] [CrossRef]
- Truta, E.; Mihai, C.; Gherghel, D.; Vochita, G. Assessment of the Cytogenetic Damage Induced by Chromium Short-Term Exposure in Root Tip Meristems of Barley Seedlings. Water Air Soil Pollut. 2014, 225, 1933. [Google Scholar] [CrossRef]
- Miadokova, E.; Duhova, V.; Vlckova, V.; Sladkova, L.; Sucha, V.; Vlcek, D. Genetic risk assessment of acid waste water containing heavy metals. Gen. Physiol. Biophys. 1999, 18, 92–98. [Google Scholar]
- Labra, M.; Grassi, F.; Imazio, S.; Di Fabio, T.; Citterio, S.; Sgorbati, S.; Agradi, E. Genetic and DNA-methylation changes induced by potassium dichromate in Brassica napus L. Chemosphere 2004, 54, 1049–1058. [Google Scholar] [CrossRef]
- Labra, M.; Di Fabio, T.; Grassi, F.; Regondi, S.M.G.; Bracale, M.; Vannini, C.; Agradi, E. AFLP analysis as biomarker of exposure to organic and inorganic genotoxic substances in plants. Chemosphere 2003, 52, 1183–1188. [Google Scholar] [CrossRef] [Green Version]
- Gill, R.A.; Hu, X.Q.; Ali, B.; Yang, C.; Shou, J.Y.; Wu, Y.Y.; Zhou, W.J. Genotypic variation of the responses to chromium toxicity in four oilseed rape cultivars. Biol. Plant. 2014, 58, 539–550. [Google Scholar] [CrossRef]
- Qiu, B.; Zeng, F.; Cai, S.; Wu, X.; Haider, S.I.; Wu, F.; Zhang, G. Alleviation of chromium toxicity in rice seedlings by applying exogenous glutathione. J. Plant Physiol. 2013, 170, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriou, E.P.; Adamakis, I.-D.S.; Melissa, P. Effects of hexavalent chromium on microtubule organization, ER distribution and callose deposition in root tip cells of Allium cepa L. Protoplasma 2012, 249, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Palomino, W.; Javier Pena-Salamanea, E.; Andres Torres-Rodriguez, G. Ultraestructural changes induced by chromium(VI) in water hyacinth (Eichhornia crassipes) leaves. Limnetica 2015, 34, 85–94. [Google Scholar]
- Maleci, L.; Buffa, G.; Wahsha, M.; Bini, C. Morphological changes induced by heavy metals in dandelion (Taraxacum officinale Web.) growing on mine soils. J. Soils Sediments 2014, 14, 731–743. [Google Scholar] [CrossRef]
- Ali, S.; Farooq, M.A.; Yasmeen, T.; Hussain, S.; Arif, M.S.; Abbas, F.; Bharwana, S.A.; Zhang, G. The influence of silicon on barley growth, photosynthesis and ultra-structure under chromium stress. Ecotoxicol. Environ. Saf. 2013, 89, 66–72. [Google Scholar] [CrossRef]
- Mangabeira, P.A.; de Almeida, A.A.F.; Souza, V.L.; dos Santos Junior, A.J.; Silva, D.C.; de Jesus, R.M. Ultrastructural damage in tomato plants exposed to chromium III. In Proceedings of the 16th International Conference on Heavy Metals in the Environment, Rome, Italy, 23–27 September 2012; Pirrone, N., Ed.; EDP Sciences: Ilhéus, Bahia, Brazil, 2013; Volume 1. [Google Scholar]
- Hui, C.; Guo-Xin, S.H.I.; Qin-Song, X.U.; Wang, H.-X.; Yang, H.-Y.; Qiu-Hong, P.A.N. Toxic Effects of Cr~(6+) on Chlorophyll Fluorescence Parameters, Antioxidant Systems and Ultrastructure of Potamogeton crispus. Bull. Bot. Res. 2009, 29, 559–564. [Google Scholar]
- Sruthi, P.; Shackira, A.M.; Puthur, J.T. Heavy metal detoxification mechanisms in halophytes: An overview. Wetl. Ecol. Manag. 2017, 25, 129–148. [Google Scholar] [CrossRef]
- Yang, C.; Li, W.; Cao, J.; Meng, F.; Yu, Y.; Huang, J.; Jiang, L.; Liu, M.; Zhang, Z.; Chen, X.; et al. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice. Plant J. 2017, 89, 338–353. [Google Scholar] [CrossRef] [Green Version]
- Farias, D.R.; Hurd, C.L.; Eriksen, R.S.; Simioni, C.; Schmidt, E.; Bouzon, Z.L.; Macleod, C.K. In situ assessment of Ulva australis as a monitoring and management tool for metal pollution. J. Appl. Phycol. 2017, 29, 2489–2502. [Google Scholar] [CrossRef]
- Farias, D.R.; Schmidt, E.; Simioni, C.; Bouzon, Z.L.; Hurd, C.L.; Eriksen, R.S.; Macleod, C.K. Photosynthetic and ultrastructural responses of Ulva australis to Zn stress. Micron 2017, 103, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zou, J.; Wang, M.; Jiang, W. Hexavalent chromium uptake and its effects on mineral uptake, antioxidant defence system and photosynthesis in Amaranthus viridis L. Bioresour. Technol. 2008, 99, 2628–2636. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Khan, I.; Saleem, M.F.; Wang, L.C. Chromium toxicity induced alterations in growth, photosynthesis, gas exchange attributes and yield formation in maize. Pak. J. Agric. Sci. 2016, 53, 751–757. [Google Scholar]
- Pandey, V.; Dixit, V.; Shyam, R. Chromium effect on ROS generation and detoxification in pea (Pisum sativum) leaf chloroplasts. Protoplasma 2009, 236, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Dixit, V.; Pandey, V.; Shyam, R. Chromium ions inactivate electron transport and enhance superoxide generation in vivo in pea (Pisum sativum L. cv. Azad) root mitochondria. Plant Cell Environ. 2002, 25, 687–693. [Google Scholar] [CrossRef]
- Yewalkar, S.N.; Dhumal, K.N.; Sainis, J.K. Effect of chromate on photosynthesis in Cr(VI)-resistant Chlorella. Photosynthetica 2013, 51, 565–573. [Google Scholar] [CrossRef]
- Cho, D.-W.; Song, H.; Schwartz, F.W.; Kim, B.; Jeon, B.-H. The role of magnetite nanoparticles in the reduction of nitrate in groundwater by zero-valent iron. Chemosphere 2015, 125, 41–49. [Google Scholar] [CrossRef]
- El-Esawi, M.; Arthaut, L.-D.; Jourdan, N.; d’Harlingue, A.; Link, J.; Martino, C.F.; Ahmad, M. Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef]
- Choudhary, K.K.; Agrawal, S.B. Assessment of Fatty Acid Profile and Seed Mineral Nutrients of Two Soybean (Glycine max L.) Cultivars under Elevated Ultraviolet-B: Role of ROS, Pigments and Antioxidants. Photochem. Photobiol. 2016, 92, 134–143. [Google Scholar] [CrossRef]
- Dey, S.; Paul, A.K. Evaluation of chromate reductase activity in the cell-free culture filtrate of Arthrobacter sp. SUK 1201 isolated from chromite mine overburden. Chemosphere 2016, 156, 69–75. [Google Scholar] [CrossRef]
- Gautam, M.; Singh, A.K.; Johri, R.M. Effect of chromium toxicity on growth, chlorophyll and some macronutrients of Solanum lycopersicum and Solanum melongena. Indian J. Agric. Sci. 2014, 84, 1115–1123. [Google Scholar]
- Hammud, H.H.; Abbas, I.; Al-Khalili, D. Kinetics and thermodynamics of chromate and phosphate uptake by polypyrrole: Batch and column studies. J. Incl. Phenom. Macrocycl. Chem. 2015, 82, 395–405. [Google Scholar] [CrossRef]
- Sheetal, K.R.; Singh, S.D.; Anand, A.; Prasad, S. Heavy metal accumulation and effects on growth, biomass and physiological processes in mustard. Indian J. Plant Physiol. 2016, 21, 219–223. [Google Scholar] [CrossRef]
- Yasin, M.; Faisal, M. Comparative Analysis of Tannery-Effluent Contaminated Soil and Mixed Culture Bacterial Inoculation on Helianthus annuus L. Growth. J. Chem. Soc. Pak. 2012, 34, 1573–1577. [Google Scholar]
- Li, S.; Huang, H.; Li, Z.; Li, Z.; He, Z.; Liang, H. Chromium removal capability and photosynthetic characteristics of Cyperus alternifolius and Coix lacryma-jobi L. in vertical flow constructed wetland treated with hexavalent chromium bearing domestic sewage. Water Sci. Technol. 2017, 76, 2203–2212. [Google Scholar] [CrossRef]
- Yasin, M.; Faisal, M. Assessing the Phytotoxicity of Tannery Waste-Contaminated Soil on Zea mays (Lin) Growth. Pol. J. Environ. Stud. 2013, 22, 1871–1876. [Google Scholar]
- Revathi, S.; Subhashree, V. Comparative Study of Hexavalent Chromium Induced Biochemical Changes With and Without EDTA in Sesbania grandiflora L. Pers. J. Agric. Sci. Technol. 2016, 18, 1289–1301. [Google Scholar]
- Dias, M.C.; Moutinho-Pereira, J.; Correia, C.; Monteiro, C.; Araujo, M.; Brueggemann, W.; Santos, C. Physiological mechanisms to cope with Cr(VI) toxicity in lettuce: Can lettuce be used in Cr phytoremediation? Environ. Sci. Pollut. Res. 2016, 23, 15627–15637. [Google Scholar] [CrossRef]
- Mathur, S.; Kalaji, H.M.; Jajoo, A. Investigation of deleterious effects of chromium phytotoxicity and photosynthesis in wheat plant. Photosynthetica 2016, 54, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Xiao, J.; Feng, X.; Gan, L.; Tang, Y. Effects of Chromium stress on physiological and ecophysiological characteristics of male and female plants of Humulus scandens. Acta Pratacult. Sin. 2016, 25, 131–139. [Google Scholar]
- Reale, L.; Ferranti, F.; Mantilacci, S.; Corboli, M.; Aversa, S.; Landucci, F.; Baldisserotto, C.; Ferroni, L.; Pancaldi, S.; Venanzoni, R. Cyto-histological and morpho-physiological responses of common duckweed (Lemna minor L.) to chromium. Chemosphere 2016, 145, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Sheetal, K.R.; Singh, S.; Prasad, S.; Mina, U.; Datta, S.P.; Anand, A. Physiological and biochemical responses of different vegetables to chromium. Biochem. Cell. Arch. 2015, 15, 421–425. [Google Scholar]
- Da Conceicao Gomes, M.A.; Hauser-Davis, R.A.; Suzuki, M.S.; Vitoria, A.P. Plant chromium uptake and transport, physiological effects and recent advances in molecular investigations. Ecotoxicol. Environ. Saf. 2017, 140, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Vernay, P.; Gauthier-Moussard, C.; Hitmi, A. Interaction of bioaccumulation of heavy metal chromium with water relation, mineral nutrition and photosynthesis in developed leaves of Lolium perenne L. Chemosphere 2007, 68, 1563–1575. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wu, F.; Huang, R.; Zang, G. A chromium-tolerant plant growing in Cr-contaminated land. Int. J. Phytoremediation 2007, 9, 167–179. [Google Scholar] [CrossRef]
- Mallick, S.; Sinam, G.; Mishra, R.K.; Sinha, S. Interactive effects of Cr and Fe treatments on plants growth, nutrition and oxidative status in Zea mays L. Ecotoxicol. Environ. Saf. 2010, 73, 987–995. [Google Scholar] [CrossRef]
- Xu, F.; Deng, J. Effects of Salicylic Acid on Germination of Soybean Seed under Chromium Stress. Soybean Sci. 2012, 31, 852–854. [Google Scholar]


{kind=link}
| Plant Species | Common Name | ROS Types | Cr(VI) Concentration | References |
|---|---|---|---|---|
| Arabidopsis thaliana | Arabidopsis | O2−, H2O2 | 100–400 µM | [8,23] |
| Helianthus annuus | Sunflower | O2−, OH−, H2O2 | 20 mg/L & 20 mg/Kg | [24,25,26] |
| Zea mays | Maize | O2−, H2O2, OH− | 100–300 µM & 100–300 mg/Kg | [27,28,29,30,31,32] |
| Brassica juncea | Indian mustard | 1O2, O2−, H2O2, OH− | 300 µM | [17,33] |
| Glycine max | Soybean | H2O2 | 400 mg/kg & 500 mg/kg Cr(III) | [22] |
| Oryza sativa | Rice | O2−, H2O2 | 80–200 µM | [34,35,36,37] |
| Amaranthus viridis & Amaranthus cruentus | Green & Blood amaranth | O2−, H2O2 | 50 µM | [38] |
| Chenopodium quinoa | Quinoa | H2O2 | 5 mM Cr(III) | [39] |
| Cucumis sativus | Cucumber | O2−, H2O2 | 200 µM | [40] |
| Brassica napus | oilseed rape | O2−, H2O2, OH− | 400 μM | [41,42] |
| Brassica campestris | Cabbage | O2− | 1 mg/L | [43] |
| Pisum sativum | Pea | O2−, H2O2 | 100 μM | [44] |
| Allium cepa | Onion | O2−, H2O2, OH− | 200 µM | [45] |
| Matricaria chamomilla | Chamomile | H2O2 | 120 µM Cr(III) | [46] |
| Lens culinaris | Lentil | H2O | 250 µM | [47] |
| Raphanus sativus | Radish | O2−, H2O2 | 1.2 mM | [48] |
| Pistia Stratiotes | Lettuce | H2O2 | 10 mM | [49] |
| Plant Species | Common Name | Enzymes | Cr(VI) | References |
|---|---|---|---|---|
| Helianthus annuus | Sunflower | CAT, SOD, POD, APX | 20 mg/kg | [25,26] |
| Triticum aestivum Hordeum vulgare | Wheat & Barley | CAT, APX | 22 mg/kg | [59] |
| Brassica oleracea | Cauliflower | CAT, SOD, POD | 200 μM | [60] |
| Pennisetum alopecuroides | Fountain Grass | CAT, SOD, POD | 1500 mg/kg | [61] |
| Sorghum bicolor | Sorghum | CAT, SOD, APX, GR, GST | 64 ppm | [62] |
| Brassica juncea | Indian Mustard | GR, GPX, CAT, SOD, POD, APX, MDHAR, DHAR | 300–500 μM | [17,63] |
| Solanum melongena | Eggplant | APX, GST, GR | 25 µM | [64] |
| Amaranthus viridis & Amaranthus cruentus | Green & Blood Amaranth | CAT, SOD, POD, GST | 50 μM | [38] |
| Zea mays | Maize | APX, CAT, SOD, POD | 100–250 μM | [65,66] |
| Hibiscus cannabinus | Kenaf | CAT, SOD, POD | 1.5 Mm Cr(III) | [67] |
| Oryza sativa | Rice | APX, CAT, SOD, POD, GR | 20–100 μM | [68,69] |
| Vigna radiate | Mung Bean | CAT, SOD, POD | 500 μM | [70] |
| Brassica chinensis | Pakchoi | CAT, SOD, POD | 100 μM & 200 mg/kg | [71,72] |
| Setaria italic | Foxtail Millet | CAT, SOD, POD, APX | 1000 μM | [73] |
| Solanum nigrum & Parthenium hysterophorus | Black Nightshade & Santa-maria | SOD, POD | 500 μM Cr(III) | [74] |
| Brassica rapa | Turnip | SOD, APX | 250 µM | [75] |
| Brassica napus | Rapeseed | CAT, SOD, POD, APX | 500 μM | [76] |
| Brassica campestris | Cabbage | SOD, POD | 1 mg/L | [43] |
| Gossypium hirsutum | Cotton | CAT, SOD, POD, APX | 100 μM | [77] |
| Corchorus olitorius | Tossa Jute | CAT, SOD, POD, APX, GR | 400 mg/kg | [78] |
| Brassica napus | Canola | CAT, SOD, POD, APX | 50 μM | [79] |
| Raphanus sativus | Radish | CAT, SOD, POD | 8 mM | [80] |
| Hordeum vulgare | Barley | CAT, SOD, POD, APX | 225 μM | [81] |
| Plant Species | Common Name | LPO | Cr(VI) | References |
|---|---|---|---|---|
| Arabidopsis thaliana | Arabidopsis | MDA | 400 µM | [8] |
| Zea mays | Maize | MDA | 100–300 µM | [27,28,31,32,65] |
| Triticum aestivum Hordeum vulgare | Wheat & Barley | MDA | 22 mg/kg | [59] |
| Solanum lycopersicum | Tomatoes | MDA | 24.66 mg/k | [83] |
| Oryza sativa | Rice | MDA, TBARS | 20–200 µM & 20 mg/L | [35,36,69,82,84,85] |
| Limnobium laevigatum | Floating Plant | MDA | 70 µg/L Cr(III) | [86] |
| Citrus reticulata Blanco | Kinnow | MDA | 750 µM | [87] |
| Sorghum bicolor | Sorghum | MDA | 64 ppm | [62] |
| Helianthus annuus | Sunflower | MDA | 20 mg/kg | [25] |
| Brassica juncea | Indian Mustard | MDA | 100–500 μM & 100 mg/Kg | [17,63,88,89] |
| Solanum melongena | Eggplant | MDA | 25 µM | [64] |
| Tradescantia pallida | Rose | MDA | 20 mg/L | [90] |
| Amaranthus viridis & Amaranthus cruentus | Green & Blood Amaranth | MDA | 50 μM | [38] |
| Pteris vittata | Chinese Brake | TBARS | 5 mM | [91] |
| Chenopodium quinoa | Quinoa | MDA | 5 mM Cr(III) | [39] |
| Saccharum spp. Hybrid | Sugarcane | MDA | 50 ppm | [92] |
| Cucumis sativus | Cucumber | MDA | 200 µM | [40] |
| Pisum sativum | Pea | MDA | 100 μM | [44] |
| Brassica rapa | Turnip | MDA | 250 µM Cr(III) | [75] |
| Brassica napus | Canola | MDA | 50–100 μM | [79,93,94] |
| Brassica oleracea | Cauliflower | MDA | 250 μM | [95] |
| Salvinia minima | Floating Fern | MDA | 20 mg/L | [96] |
| Tradescantia pallida | Wandering Jew | TBARS | 20 mg/L | [97] |
| Gossypium hirsutum | Cotton | MDA | 100 μM | [77] |
| Triticum aestivum | Wheat | TBARS | 200 μM | [98] |
| Allium cepa | Onion | MDA | 200 μM | [45] |
| Raphanus sativus | Radish | MDA | 125 m | [80] |
| Miscanthus sinensis | Chinese Reed | MDA | 1000 μM | [99] |
| Brassica napus | Rapeseed | TBARS | 480 μM Cr(III) | [100] |
| Plant Species | Common Name | Genotoxicity | Cr- Type | References |
|---|---|---|---|---|
| Glycine max | Soybean | DNA damage | Cr(VI)/(III) | [22] |
| Vicia faba | Faba Bean | Micronucleus, Chromosomal fragmentation & bridging, Increase in % tail DNA, tail moment and Tail length | Tannery solid waste & Cr(VI) | [124,125,126,127] |
| Allium cepa | Onion | DNA damage, Chromosomal Aberrations, Micronuclei, Chromosomal fragmentation & bridging | Tannery solid waste, Tannery effluent & Cr(VI) | [45,125,127,128,129] |
| Hordeum vulgare | Barley | Chromosomal aberrations | Cr(VI) | [130] |
| Vicia sativa | Vetch | Chromosomal aberration, Chromosomal fragmentation & bridging | Wastes, Cr(VI)/(III) | [125,127,131] |
| Raphanus sativus | Radish | Chromosomal aberration | Cr(VI)/(III) | [125] |
| Zea mays | Maize | Chromosomal aberration | Cr(VI)/(III) | [125] |
| Brassica napus | Oilseed Rape | Methylation changes, mutation | Cr(VI) | [127,132] |
| Arabidopsis thaliana | Arabidopsis | DNA mutation | Cr(VI) | [127,133] |
| Plant Species | Common Names | PT | Effect | Cr-Type | References |
|---|---|---|---|---|---|
| Glycine max | Soybean | L | Loss of Epi-C-wax increased TRICH-number | Cr(VI)/(III) | [22] |
| Brassica napus | Oilseed rape | L & R | Alteration in CW, MITO, CM, THY, PG, SG, GB, ER, Irregular nucleus, THY disappeared, Increased SG number/size. | Cr(VI) | [41,42,120,134] |
| Triticum aestivum Hordeum vulgare | Wheat & Barley | L | Damaged CHLP, THY; Increased PG, Swollen MITO; altered I-cristae | Cr(VI) | [59] |
| Nicotiana tabacum | Tobacco | L & R | CW/CM not distinguishable, Disarranged CHLP structure, Undeveloped nucleus, damaged NM, Swelled/distorted THY, Damaged CHLP, MITO, Altered THY-O, Increased PG, Large SG | Cr(VI) | [122,123] |
| Oryza sativa | Rice | L | Swollen CHLP, grana/stroma/lamellae, Reduced grana/CHLP, Increased SG, Matrix zone expanded. | Cr(VI) | [35,135] |
| Arabidopsis thaliana | Arabidopsis | R | T-nuclei, GB disintegrated, spherical MITO, plastids; T-stroma; damaged MIOT, plastids; increased SG, amorphous material deposition in CW, ML, vacuoles, collapsed vacuoles, cytoplasm contained opaque lipid, | Cr(VI) | [23,47,136] |
| Eichhornia crassipes | Water Hyacinth | L | Damaged THY, MITO, CHLP (structure/distribution), grana | Cr(VI) | [137] |
| Salvinia minima | Floating Fern | L | Damaged CHLP, grana, THY, increased number/size of SG; large PG | Cr(VI) | [96] |
| Taraxacum officinale | Dandelion | C | Altered MITO with no/reduced I-cristae | Cr(VI) | [138] |
| Hordeum vulgare | Barley | L & R | Swollen CHLP, increased PG, Disintegrated/disappeared THY, MITO, Increased SG size/number, Increased vacuolar size, Cr-presence in CW, Vacuoles, Nucleus disruption/disappearance | Cr(VI) | [139] |
| Solanum lycopersicum | Tomatoes | P | Abnormal shaped reduced grana/CHLP; altered THY, MITO; reduced cristae numbers | Cr(III) | [140] |
| Potamogeton crispus | Curled Pondweed | L | Swollen CHLP, CHLP- envelop breakage, decreasing cristae, MITO vacuolization | Cr(VI) | [141] |
| Plant Species | Common Name | Alteration in Photosynthetic Parameters | Cr(VI) | References |
|---|---|---|---|---|
| Arabidopsis thaliana & Brassica juncea | Arabidopsis & Indian Mustard | Reduced chl a, b, and t Reduced chl a, Reduced Chl t, Carotenoids, and net photo, b, and t, Gas exchange | 400 µM 100–300 µM & 100 mg/Kg | [8] & [58,88,89,157] |
| Helianthus annuus | Sunflower | Reduced chl a, b, t, gas exchange, and carotenoid levels | Tannery effluent & 20 mg/kg | [26,158] |
| Citrus reticulate | Kinnow Mandarin | Decreased chl t, photosynthetic activity, Trmmol, Cond, and water use efficiency | 0.75 mM | [87] |
| Cyperus alternifolius & Coix lacryma-jobi | Umbrella Palm & Adlay Millet | Inhibition in photosynthetic capacities | 40 mg/L | [159] |
| Solanum melongena | Eggplant | Reduced pigments, photo, photochemistry of PSII | 25 μM | [64] |
| Oryza sativa | Rice | Reduced Chl a, b, and carotenoids, Reduced Fv/Fm | 80–200 µM | [34,35] |
| Zea mays | Maize | Reduced carotenoids, chl a, b, and t, Photo, Trmmol, Ci, Water use efficiency and intrinsic, Alteration in Fv/Fm, Fv/F0, Fm/F0, and qP | Tannery effluent & 150–250 μM | [29,147,160] |
| Amaranthus viridis & Amaranthus cruentus | Green & Blood Amaranth | Inhibition photochemistry of PSII | 50 μM | [38] |
| Nicotiana tabacum | Tobacco | Reduced Chl a, b, carotenoids, photo, gas exchange, Fv/Fm fluorescence | 50 μM | [122] |
| Sesbania grandiflora | Hummingbird Tree | Reduced Chl t | 1.92 mM/Kg | [161] |
| Lactuca sativa | Lettuce | Decreased levels Chl a, ΦPSII, qp, NPQ, PN and RuBisCO activity | 200 mg/L | [162] |
| Triticum aestivum | Wheat | Decline active reaction centers of PSII, ETR, and PSII heterogeneity | 300 μM | [163] |
| Humulus scandens | Asian Hop | Decreased chl f parameters, chl t, and PSII reaction | 300 mg/kg Cr(III) | [164] |
| Cucumis sativus | Cucumber | Decline in Fm, Fv, Fv/Fm, Fm/F0, and Fv/F0 | 200 µM | [40] |
| Lemna minor | Duckweed | Decreased in Fv/Fm, chl b | 6 mg/L | [165] |
| Pisum sativum | Pea | Decreased pigments and Fv/Fm, Fv/F0 and qP, and NPQ increased | 100 μM | [44] |
| Raphanus sativus, Solanum lycopersicum & Spinacia oleracea | Radish, Tomato & Spinach | Reduced photosynthetic activity and Chl t | 100 mg/kg | [166] |
| Brassica napus | Rapeseed | Reduced chl t, and carotenoid | 500 μM | [76] |
| Solanum lycopersicum & Solanum melongena | Tomato & Eggplant | Reduced pigments | 7.5 ppm | [155] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wakeel, A.; Xu, M.; Gan, Y. Chromium-Induced Reactive Oxygen Species Accumulation by Altering the Enzymatic Antioxidant System and Associated Cytotoxic, Genotoxic, Ultrastructural, and Photosynthetic Changes in Plants. Int. J. Mol. Sci. 2020, 21, 728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030728
Wakeel A, Xu M, Gan Y. Chromium-Induced Reactive Oxygen Species Accumulation by Altering the Enzymatic Antioxidant System and Associated Cytotoxic, Genotoxic, Ultrastructural, and Photosynthetic Changes in Plants. International Journal of Molecular Sciences. 2020; 21(3):728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030728
Chicago/Turabian StyleWakeel, Abdul, Ming Xu, and Yinbo Gan. 2020. "Chromium-Induced Reactive Oxygen Species Accumulation by Altering the Enzymatic Antioxidant System and Associated Cytotoxic, Genotoxic, Ultrastructural, and Photosynthetic Changes in Plants" International Journal of Molecular Sciences 21, no. 3: 728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030728