Survival Motor Neuron Protein Participates in Mouse Germ Cell Development and Spermatogonium Maintenance

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
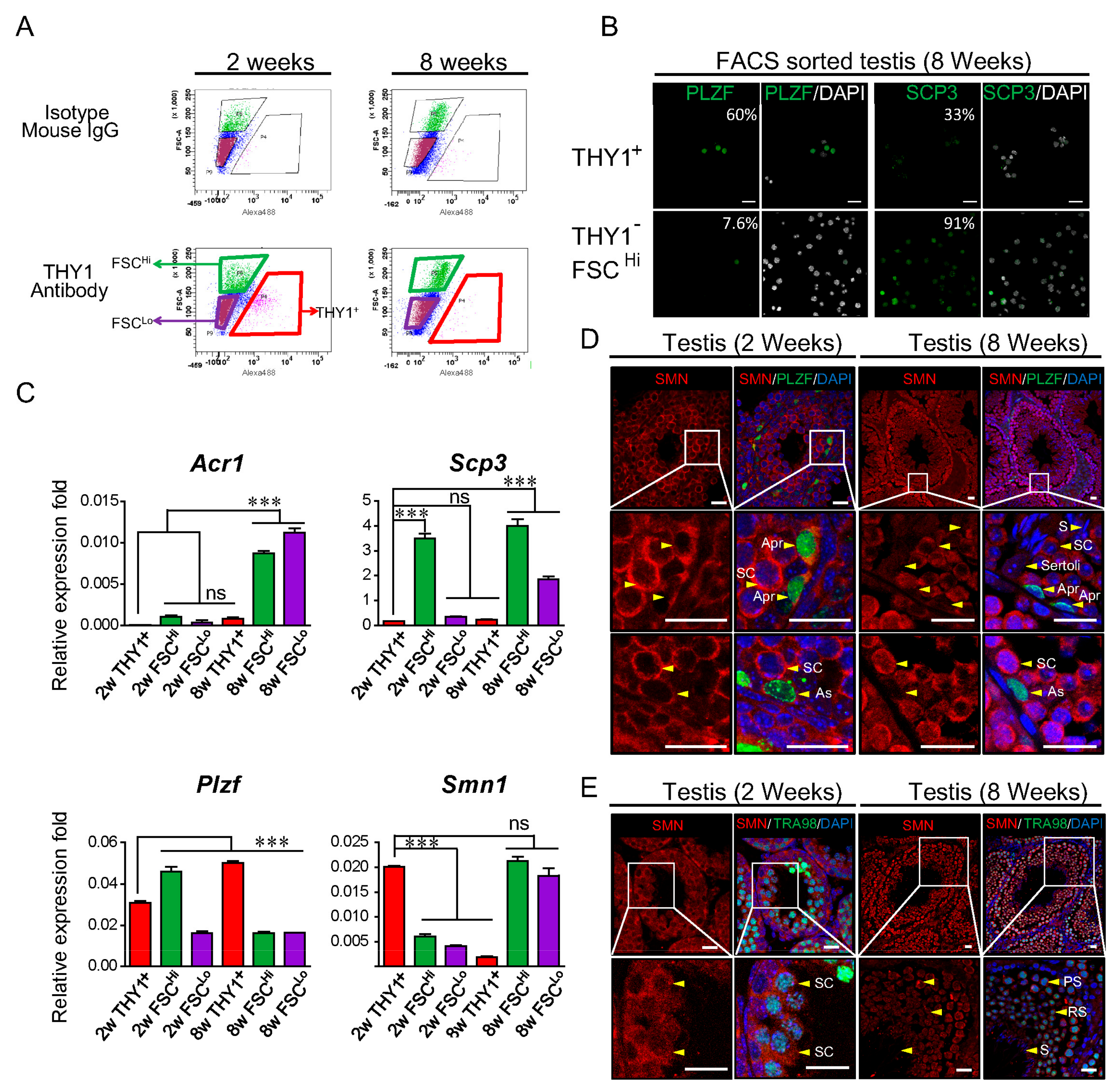
2.1. SMN Expression during Spermatogenesis
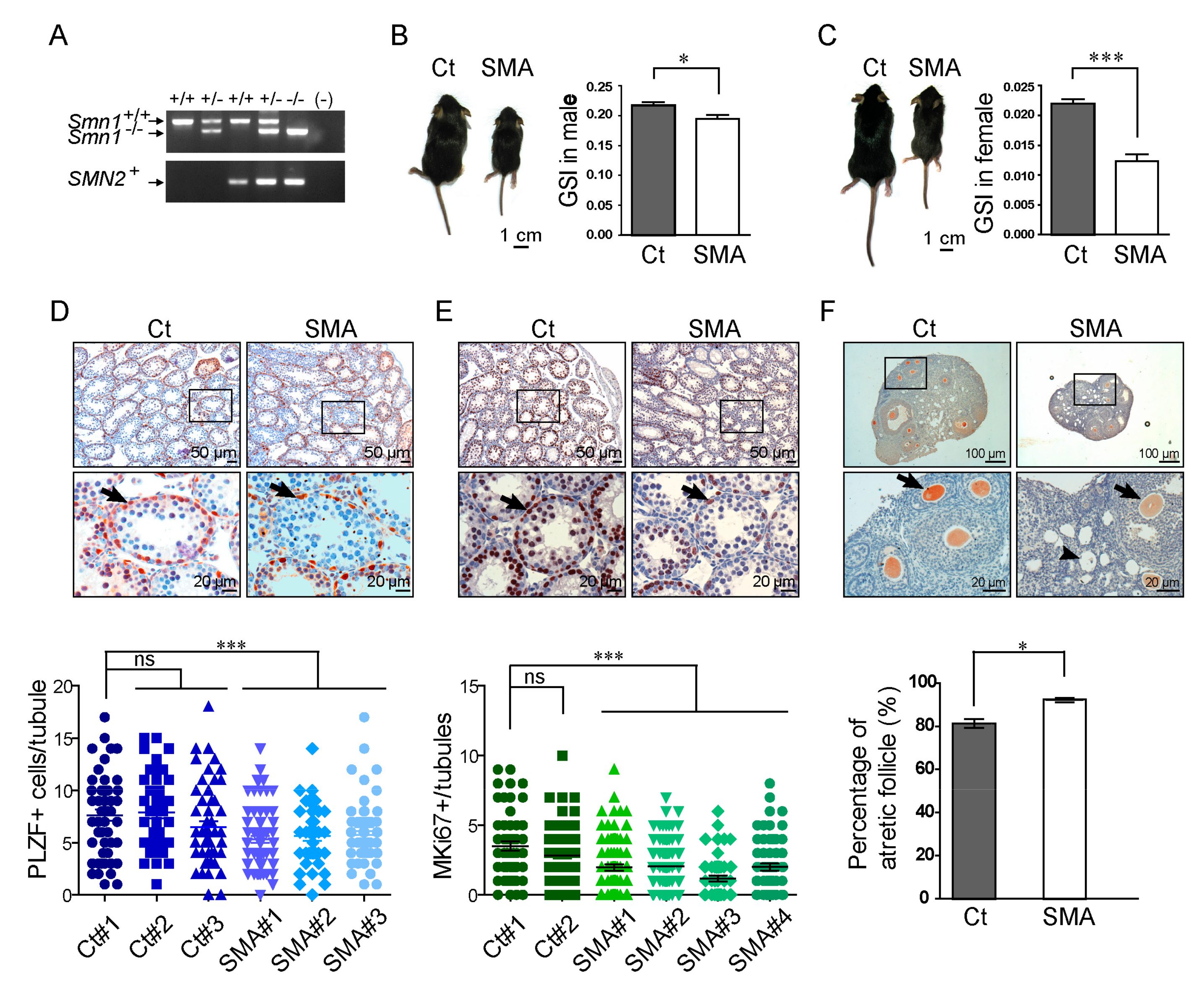
2.2. Developmental Delay in the Gonadal Tissues of SMA-Like Mice
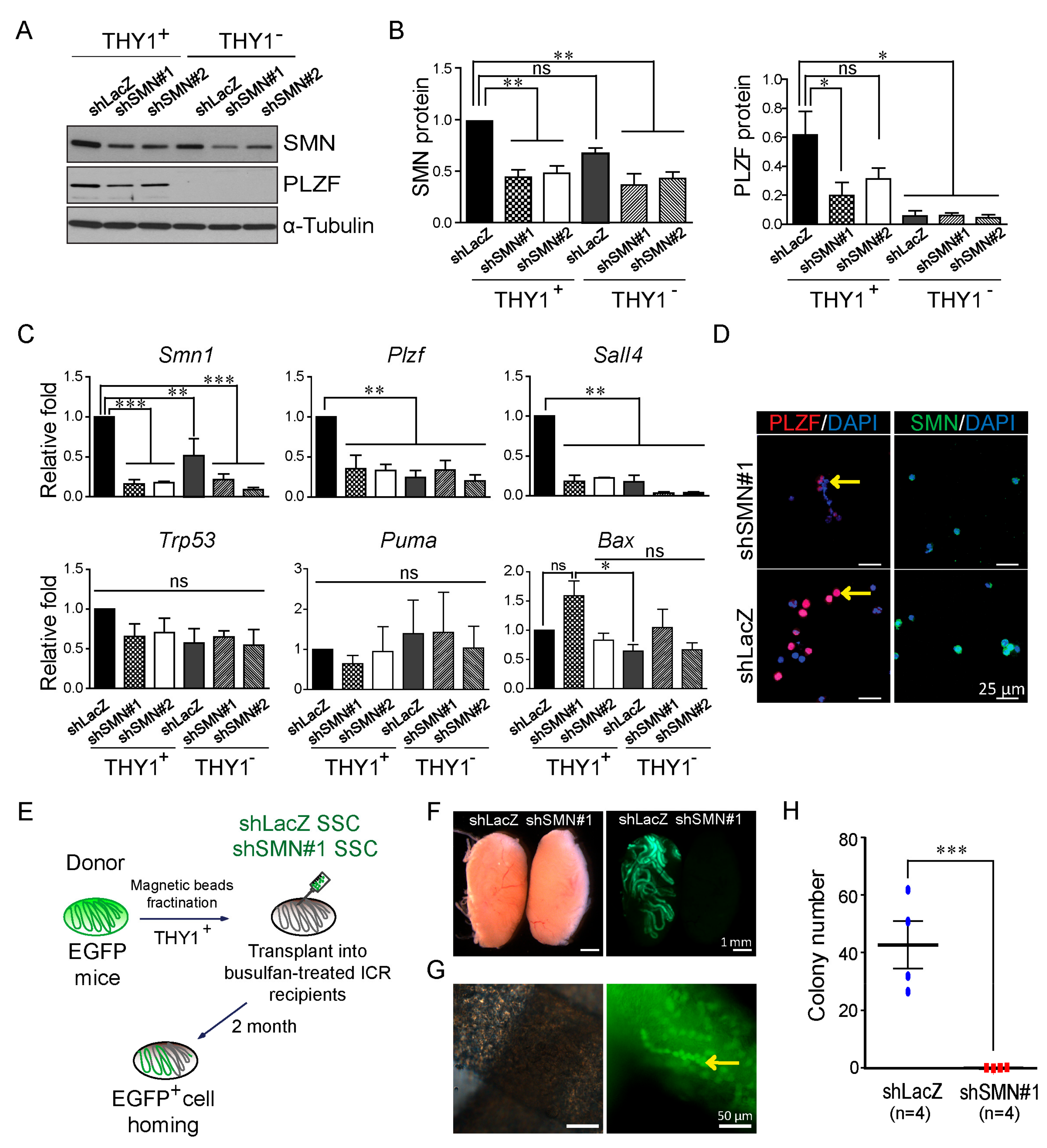
2.3. Decreased Ability of Self-Renewal in SMN-Depleted SSCs
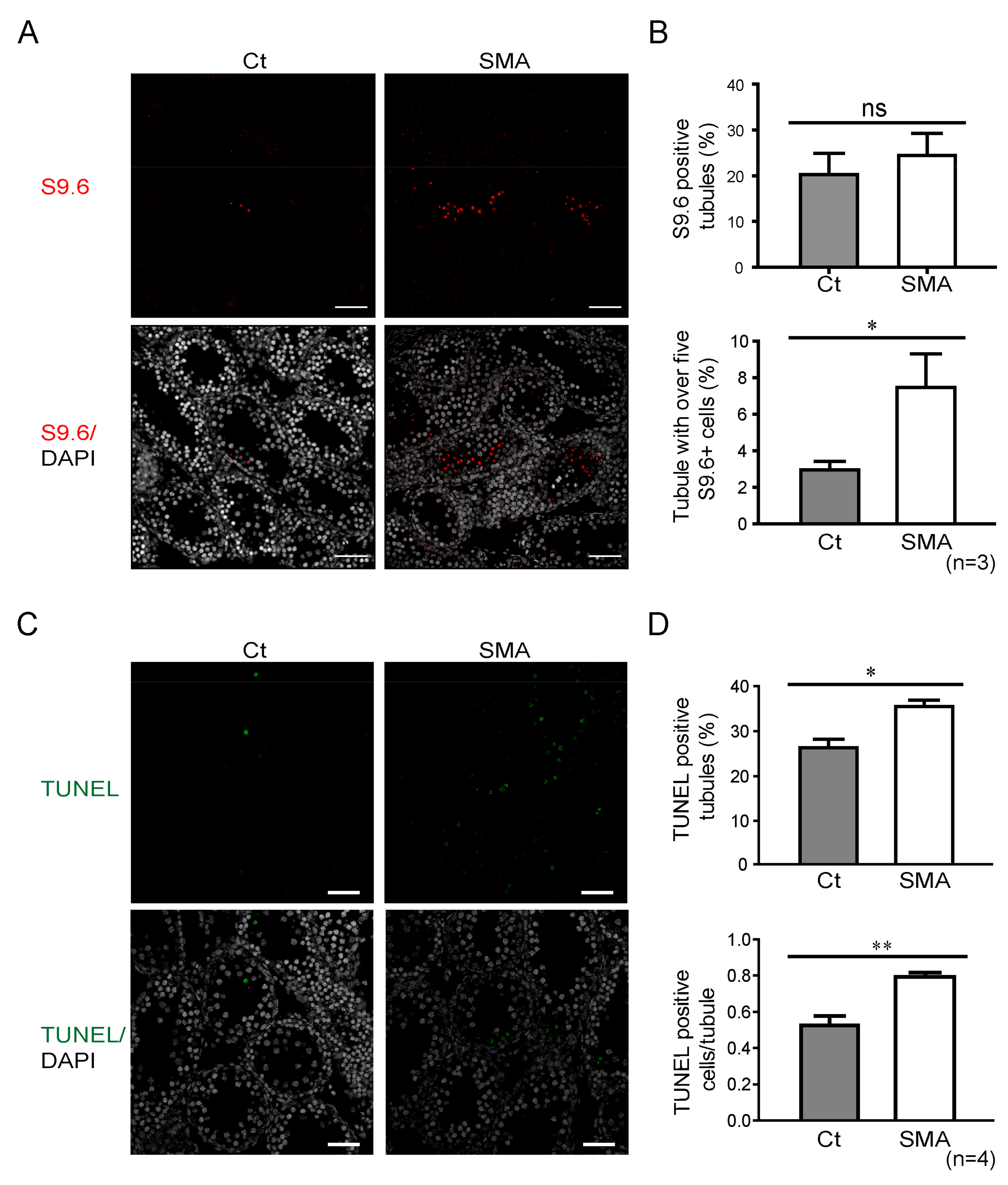
2.4. Increased R-loop Signal Following SMN Depletion
3. Discussion
4. Methods
4.1. Availability of Data and Materials
4.2. Animals and Tissue Sampling
4.3. Fluorescence-Activated Cell Sorting (FACS)
4.4. Immunohistochemistry (IHC)
4.5. Immunofluorescent Staining and Confocal Microscopy
4.6. Western Blotting
4.7. Spermatogonial Stem Cells’ (SSCs) Isolation and shRNA Knockdown
4.8. Germ Cell Transplantation
4.9. Genotyping
4.10. Gene Expression
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grice, S.J.; Liu, J.L. Survival motor neuron protein regulates stem cell division, proliferation, and differentiation in Drosophila. PLoS Genet. 2011, 7, e1002030. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.F.; Xu, J.; Chang, C.C.; Yang, S.H.; Li, H.Y.; Hsieh-Li, H.M.; Tsai, M.H.; Wu, S.C.; Cheng, W.T.K.; Liu, J.L.; et al. SMN is required for the maintenance of embryonic stem cells and neuronal differentiation in mice. Brain Struct. Funct. 2015, 220, 1539–1553. [Google Scholar] [CrossRef]
- Lee, L.; Davies, S.E.; Liu, J.L. The spinal muscular atrophy protein SMN affects Drosophila germline nuclear organization through the U body-P body pathway. Dev. Biol. 2009, 332, 142–155. [Google Scholar] [CrossRef] [Green Version]
- Kaida, D.; Berg, M.G.; Younis, I.; Kasim, M.; Singh, L.N.; Wan, L.; Dreyfuss, G. U1 snRNP protects pre-mRNAs from premature cleavage and polyadenylation. Nature 2010, 468, 664–668. [Google Scholar] [CrossRef]
- Pillai, R.S.; Grimmler, M.; Meister, G.; Will, C.L.; Luhrmann, R.; Fischer, U.; Schumperli, D. Unique Sm core structure of U7 snRNPs: Assembly by a specialized SMN complex and the role of a new component, Lsm11, in histone RNA processing. Genes Dev. 2003, 17, 2321–2333. [Google Scholar] [CrossRef] [Green Version]
- Matera, A.G.; Wang, Z. A day in the life of the spliceosome. Nat. Rev. Mol. Cell Biol. 2014, 15, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Gonsalvez, G.B.; Rajendra, T.K.; Wen, Y.; Praveen, K.; Matera, A.G. Sm proteins specify germ cell fate by facilitating oskar mRNA localization. Development 2010, 137, 2341–2351. [Google Scholar] [CrossRef] [Green Version]
- Gonsalvez, G.B.; Tian, L.; Ospina, J.K.; Boisvert, F.M.; Lamond, A.I.; Matera, A.G. Two distinct arginine methyltransferases are required for biogenesis of Sm-class ribonucleoproteins. J. Cell. Biol. 2007, 178, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, T.; Li, Q.; Liu, C.; Han, F.; Chen, M.; Zhang, L.; Cui, X.; Qin, Y.; Bao, S.; et al. Prmt5 is required for germ cell survival during spermatogenesis in mice. Sci. Rep. 2015, 5, 11031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.Y.; Gish, G.; Braunschweig, U.; Li, Y.; Ni, Z.; Schmitges, F.W.; Zhong, G.; Liu, K.; Li, W.; Moffat, J.; et al. SMN and symmetric arginine dimethylation of RNA polymerase II C-terminal domain control termination. Nature 2016, 529, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Becherel, O.J.; Yeo, A.J.; Stellati, A.; Heng, E.Y.; Luff, J.; Suraweera, A.M.; Woods, R.; Fleming, J.; Carrie, D.; McKinney, K.; et al. Senataxin plays an essential role with DNA damage response proteins in meiotic recombination and gene silencing. PLoS Genet. 2013, 9, e1003435. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Liu, J.G.; Zhao, J.; Brundell, E.; Daneholt, B.; Hoog, C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol. Cell 2000, 5, 73–83. [Google Scholar] [CrossRef]
- Buaas, F.W.; Kirsh, A.L.; Sharma, M.; McLean, D.J.; Morris, J.L.; Griswold, M.D.; de Rooij, D.G.; Braun, R.E. Plzf is required in adult male germ cells for stem cell self-renewal. Nat. Genet. 2004, 36, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Costoya, J.A.; Hobbs, R.M.; Barna, M.; Cattoretti, G.; Manova, K.; Sukhwani, M.; Orwig, K.E.; Wolgemuth, D.J.; Pandolfi, P.P. Essential role of Plzf in maintenance of spermatogonial stem cells. Nat. Genet. 2004, 36, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Fayomi, A.P.; Orwig, K.E. Spermatogonial stem cells and spermatogenesis in mice, monkeys and men. Stem Cell Res. 2018, 29, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Oakberg, E.F. Spermatogonial stem-cell renewal in the mouse. Anat. Rec. 1971, 169, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Bebee, T.W.; Dominguez, C.E.; Chandler, D.S. Mouse models of SMA: Tools for disease characterization and therapeutic development. Hum. Genet. 2012, 131, 1277–1293. [Google Scholar] [CrossRef]
- Hsieh-Li, H.M.; Chang, J.G.; Jong, Y.J.; Wu, M.H.; Wang, N.M.; Tsai, C.H.; Li, H. A mouse model for spinal muscular atrophy. Nat. Genet. 2000, 24, 66–70. [Google Scholar] [CrossRef]
- Hermann, B.P.; Sukhwani, M.; Winkler, F.; Pascarella, J.N.; Peters, K.A.; Sheng, Y.; Valli, H.; Rodriguez, M.; Ezzelarab, M.; Dargo, G.; et al. Spermatogonial stem cell transplantation into rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell 2012, 11, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Brinster, R.L.; Avarbock, M.R. Germline transmission of donor haplotype following spermatogonial transplantation. Proc. Natl. Acad Sci. USA 1994, 91, 11303–11307. [Google Scholar] [CrossRef] [Green Version]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis following male germ-cell transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, M.; Avarbock, M.R.; Brinster, R.L. Pattern and kinetics of mouse donor spermatogonial stem cell colonization in recipient testes. Biol. Reprod. 1999, 60, 1429–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jangi, M.; Fleet, C.; Cullen, P.; Gupta, S.V.; Mekhoubad, S.; Chiao, E.; Allaire, N.; Bennett, C.F.; Rigo, F.; Krainer, A.R.; et al. SMN deficiency in severe models of spinal muscular atrophy causes widespread intron retention and DNA damage. Proc. Natl. Acad Sci. USA 2017, 114, E2347–E2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotti, F.; Imlach, W.L.; Saieva, L.; Beck, E.S.; Hao le, T.; Li, D.K.; Jiao, W.; Mentis, G.Z.; Beattie, C.E.; McCabe, B.D.; et al. An SMN-dependent U12 splicing event essential for motor circuit function. Cell 2012, 151, 440–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skourti-Stathaki, K.; Kamieniarz-Gdula, K.; Proudfoot, N.J. R-loops induce repressive chromatin marks over mammalian gene terminators. Nature 2014, 516, 436–439. [Google Scholar] [CrossRef]
- Chernikova, S.B.; Brown, J.M. R-loops and genomic instability in Bre1 (RNF20/40)-deficient cells. Cell Cycle 2012, 11, 2980–2984. [Google Scholar] [CrossRef] [Green Version]
- Ottesen, E.W.; Howell, M.D.; Singh, N.N.; Seo, J.; Whitley, E.M.; Singh, R.N. Severe impairment of male reproductive organ development in a low SMN expressing mouse model of spinal muscular atrophy. Sci. Rep. 2016, 6, 20193. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Chen, X.; Wang, Z.; Wang, D. Is transcription in sperm stationary or dynamic? J. Reprod. Dev. 2017, 63, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Barna, M.; Merghoub, T.; Costoya, J.A.; Ruggero, D.; Branford, M.; Bergia, A.; Samori, B.; Pandolfi, P.P. Plzf mediates transcriptional repression of HoxD gene expression through chromatin remodeling. Dev. Cell 2002, 3, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Yeyati, P.L.; Shaknovich, R.; Boterashvili, S.; Li, J.; Ball, H.J.; Waxman, S.; Nason-Burchenal, K.; Dmitrovsky, E.; Zelent, A.; Licht, J.D. Leukemia translocation protein PLZF inhibits cell growth and expression of cyclin A. Oncogene 1999, 18, 925–934. [Google Scholar] [CrossRef] [Green Version]
- McConnell, M.J.; Chevallier, N.; Berkofsky-Fessler, W.; Giltnane, J.M.; Malani, R.B.; Staudt, L.M.; Licht, J.D. Growth suppression by acute promyelocytic leukemia-associated protein PLZF is mediated by repression of c-myc expression. Mol. Cell Biol. 2003, 23, 9375–9388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.Y.; English, M.A.; Ball, H.J.; Yeyati, P.L.; Waxman, S.; Licht, J.D. Sequence-specific DNA binding and transcriptional regulation by the promyelocytic leukemia zinc finger protein. J. Biol. Chem. 1997, 272, 22447–22455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovelace, D.L.; Gao, Z.; Mutoji, K.; Song, Y.C.; Ruan, J.; Hermann, B.P. The regulatory repertoire of PLZF and SALL4 in undifferentiated spermatogonia. Development 2016, 143, 1893–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, B.T.; Gassei, K.; Orwig, K.E. Spermatogonial stem cell regulation and spermatogenesis. Philos. Trans. R Soc. Lond. B Biol. Sci. 2010, 365, 1663–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.L.; Tanaka, S.S.; Kumagai, M.; Fujimoto, Y.; Terabayashi, T.; Matsui, Y.; Nishinakamura, R. Sall4 is essential for mouse primordial germ cell specification by suppressing somatic cell program genes. Stem Cells 2015, 33, 289–300. [Google Scholar] [CrossRef]
- Santos-Pereira, J.M.; Aguilera, A. R loops: New modulators of genome dynamics and function. Nat. Rev. Genet. 2015, 16, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Nussbacher, J.K.; Tabet, R.; Yeo, G.W.; Lagier-Tourenne, C. Disruption of RNA Metabolism in Neurological Diseases and Emerging Therapeutic Interventions. Neuron 2019, 102, 294–320. [Google Scholar] [CrossRef]
- Parker, G.C.; Li, X.; Anguelov, R.A.; Toth, G.; Cristescu, A.; Acsadi, G. Survival motor neuron protein regulates apoptosis in an in vitro model of spinal muscular atrophy. Neurotox Res. 2008, 13, 39–48. [Google Scholar] [CrossRef]
- Zhang, Z.; Lotti, F.; Dittmar, K.; Younis, I.; Wan, L.; Kasim, M.; Dreyfuss, G. SMN deficiency causes tissue-specific perturbations in the repertoire of snRNAs and widespread defects in splicing. Cell 2008, 133, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Fuller, H.R.; Gillingwater, T.H.; Wishart, T.M. Commonality amid diversity: Multi-study proteomic identification of conserved disease mechanisms in spinal muscular atrophy. Neuromuscul Disord 2016, 26, 560–569. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, G.K.; Szunyogova, E.; Shorrock, H.K.; Gillingwater, T.H.; Parson, S.H. Developmental and degenerative cardiac defects in the Taiwanese mouse model of severe spinal muscular atrophy. J. Anat. 2018, 232, 965–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.F.; Chen, W.S.; Chen, Y.H.; Kao, T.H.; Tseng, Y.T.; Lee, C.Y.; Chiu, Y.C.; Lee, P.L.; Lin, Q.J.; Ching, Y.H.; et al. DNMT3L promotes quiescence in postnatal spermatogonial progenitor cells. Development 2014, 141, 2402–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Growth factors essential for self-renewal and expansion of mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16489–16494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, F.S.; Cheng, C.C.; Peng, S.Y.; Huang, H.Y.; Lian, W.S.; Jan, M.L.; Fang, Y.T.; Cheng, E.C.; Lee, K.H.; Cheng, W.T.; et al. Isolation of therapeutically functional mouse bone marrow mesenchymal stem cells within 3 h by an effective single-step plastic-adherent method. Cell Prolif. 2010, 43, 235–248. [Google Scholar] [CrossRef]
- Ogawa, T.; Arechaga, J.M.; Avarbock, M.R.; Brinster, R.L. Transplantation of testis germinal cells into mouse seminiferous tubules. Int. J. Dev. Biol. 1997, 41, 111–122. [Google Scholar]
- Sato, T.; Katagiri, K.; Kubota, Y.; Ogawa, T. In vitro sperm production from mouse spermatogonial stem cell lines using an organ culture method. Nat. Protoc. 2013, 8, 2098–2104. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-F.; Xu, J.; Lin, T.-Y.; Hsu, J.; Hsieh-Li, H.-M.; Hwu, Y.-M.; Liu, J.-L.; Lu, C.-H.; Sung, L.-Y. Survival Motor Neuron Protein Participates in Mouse Germ Cell Development and Spermatogonium Maintenance. Int. J. Mol. Sci. 2020, 21, 794. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030794
Chang W-F, Xu J, Lin T-Y, Hsu J, Hsieh-Li H-M, Hwu Y-M, Liu J-L, Lu C-H, Sung L-Y. Survival Motor Neuron Protein Participates in Mouse Germ Cell Development and Spermatogonium Maintenance. International Journal of Molecular Sciences. 2020; 21(3):794. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030794
Chicago/Turabian StyleChang, Wei-Fang, Jie Xu, Tzu-Ying Lin, Jing Hsu, Hsiu-Mei Hsieh-Li, Yuh-Ming Hwu, Ji-Long Liu, Chung-Hao Lu, and Li-Ying Sung. 2020. "Survival Motor Neuron Protein Participates in Mouse Germ Cell Development and Spermatogonium Maintenance" International Journal of Molecular Sciences 21, no. 3: 794. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030794