Protective Effects of Zerumbone on Colonic Tumorigenesis in Enterotoxigenic Bacteroides fragilis (ETBF)-Colonized AOM/DSS BALB/c Mice

 , , and
, , and
Abstract
:1. Introduction
2. Results
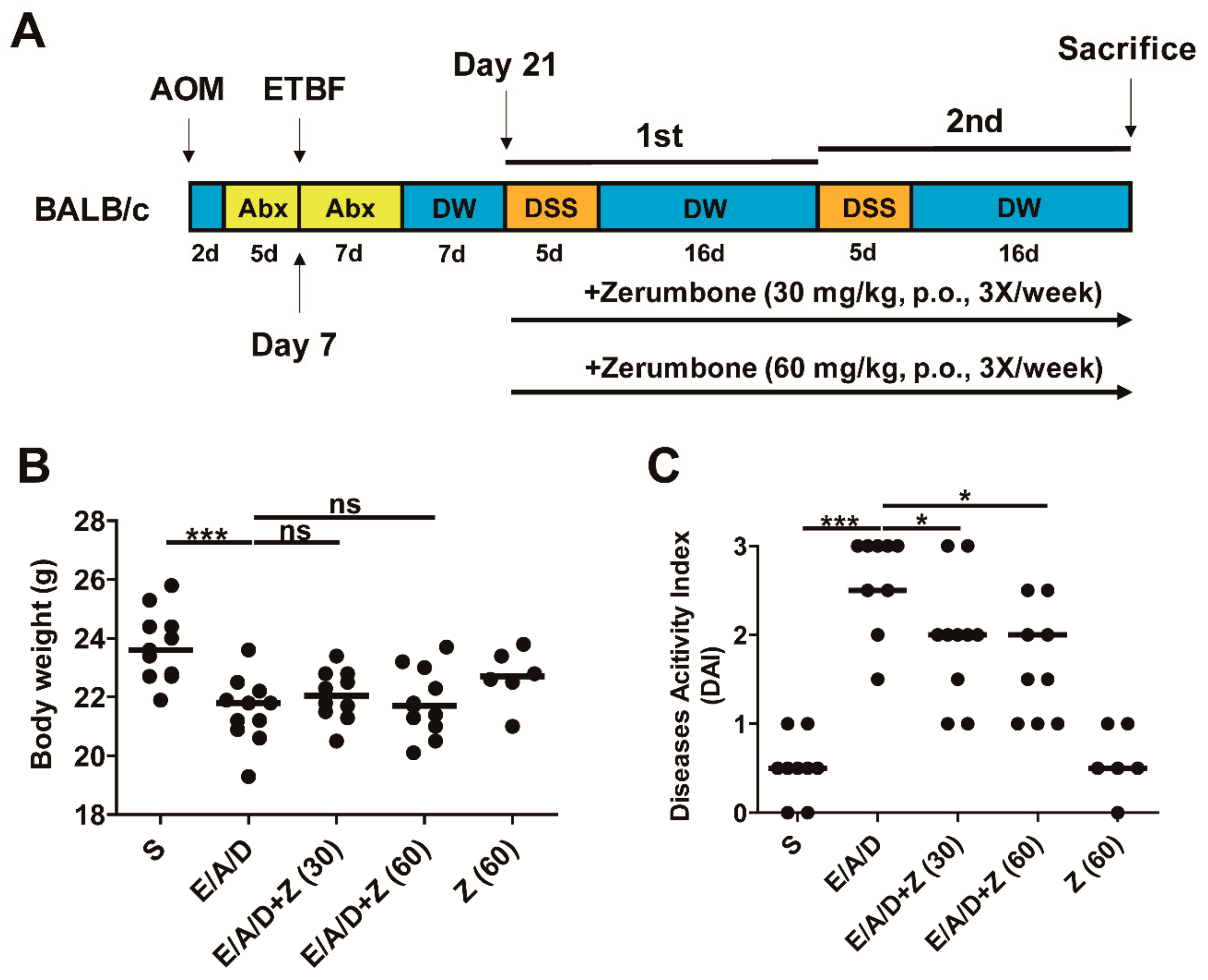
2.1. Experimental Scheme and Effects of Zerumbone on Body Weight and Disease Activity Index in ETBF/AOM/DSS Mice
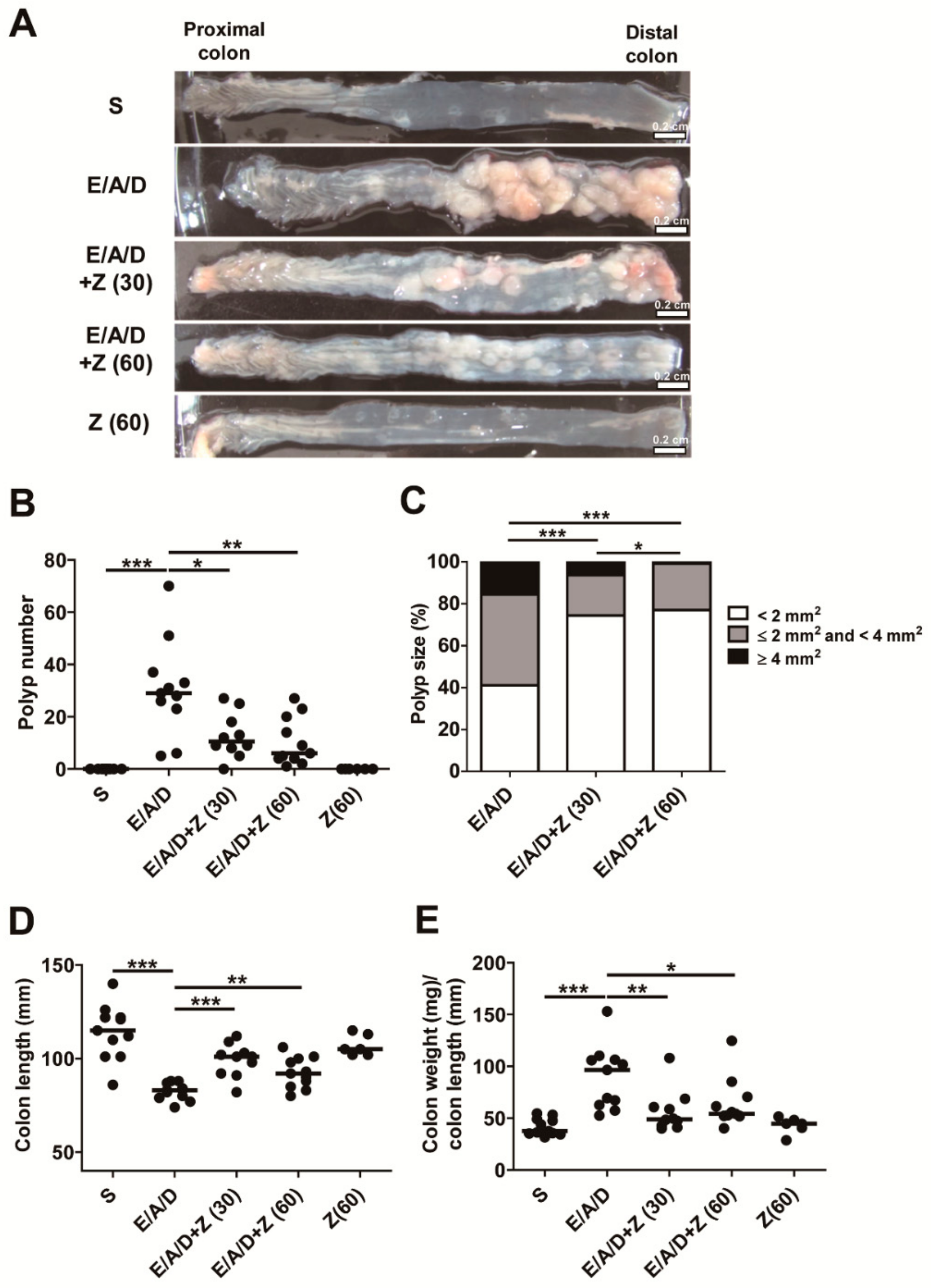
2.2. Zerumbone Reduced ETBF-Mediated Tumorigenesis in AOM/DSS Mice
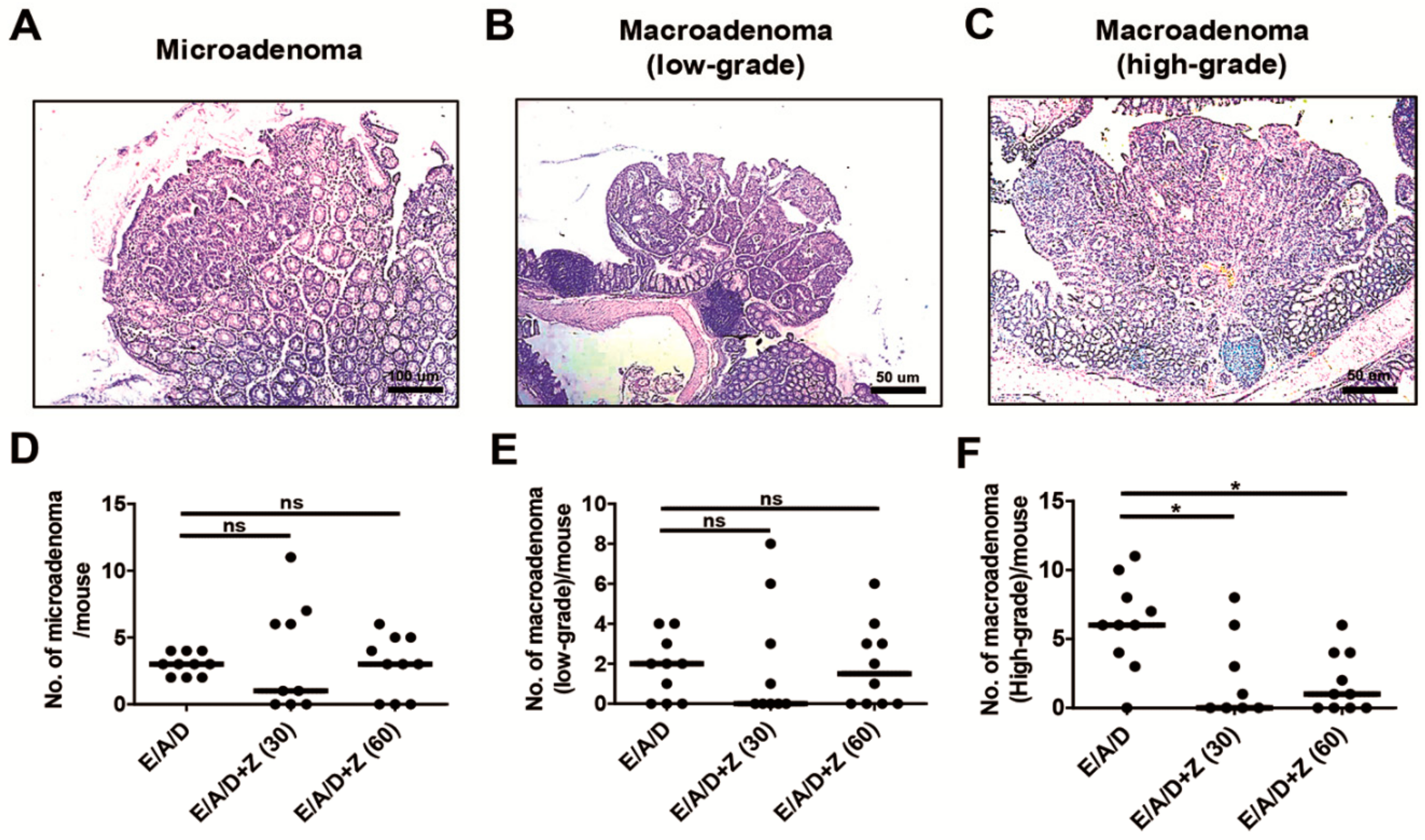
2.3. Zerumbone Reduced the Number of High-Grade Macroadenomas in ETBF-Colonized AOM/DSS Mice
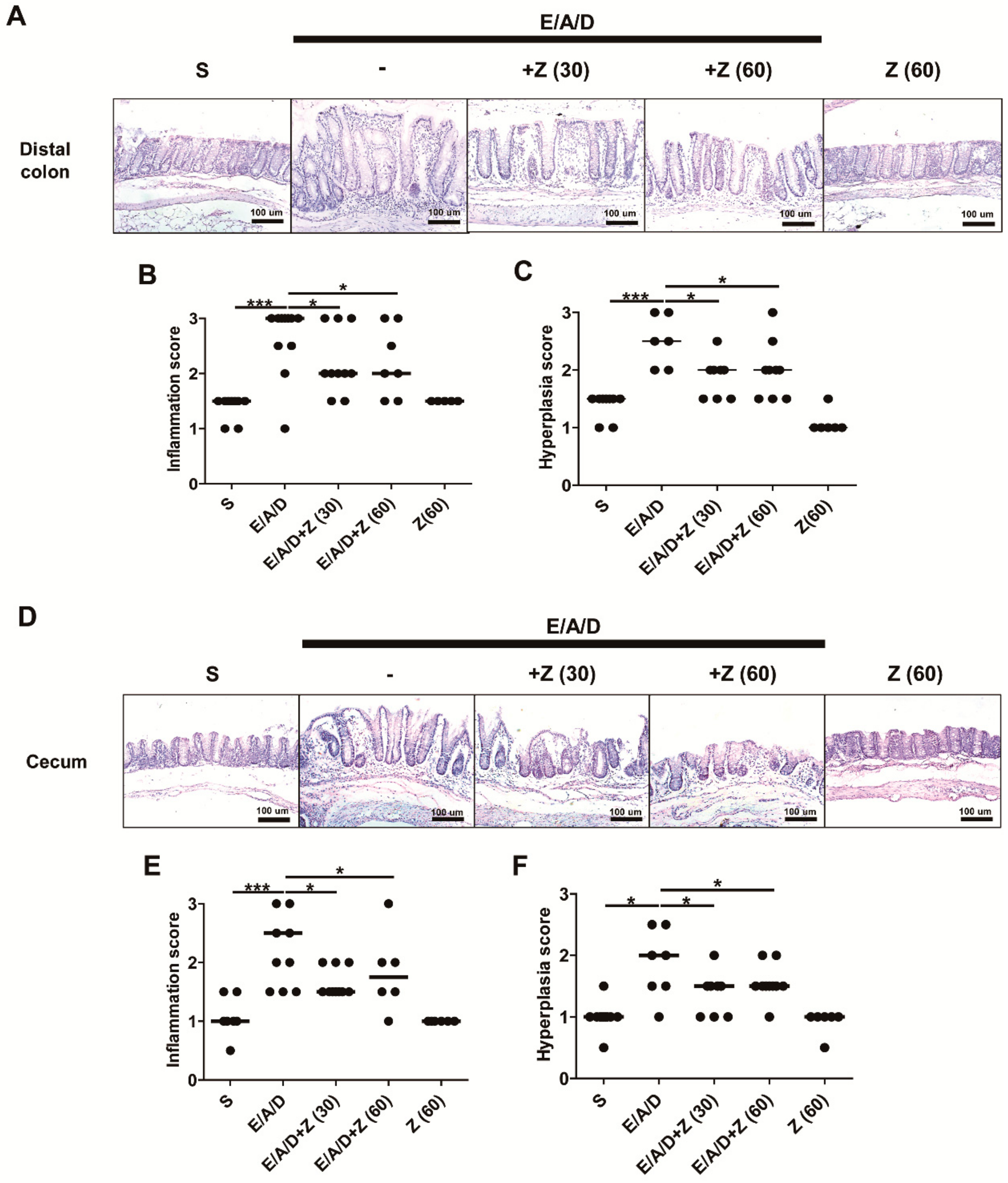
2.4. Zerumbone Decreased Large Intestinal Inflammation and Hyperplasia in ETBF-Colonized AOM/DSS Mice
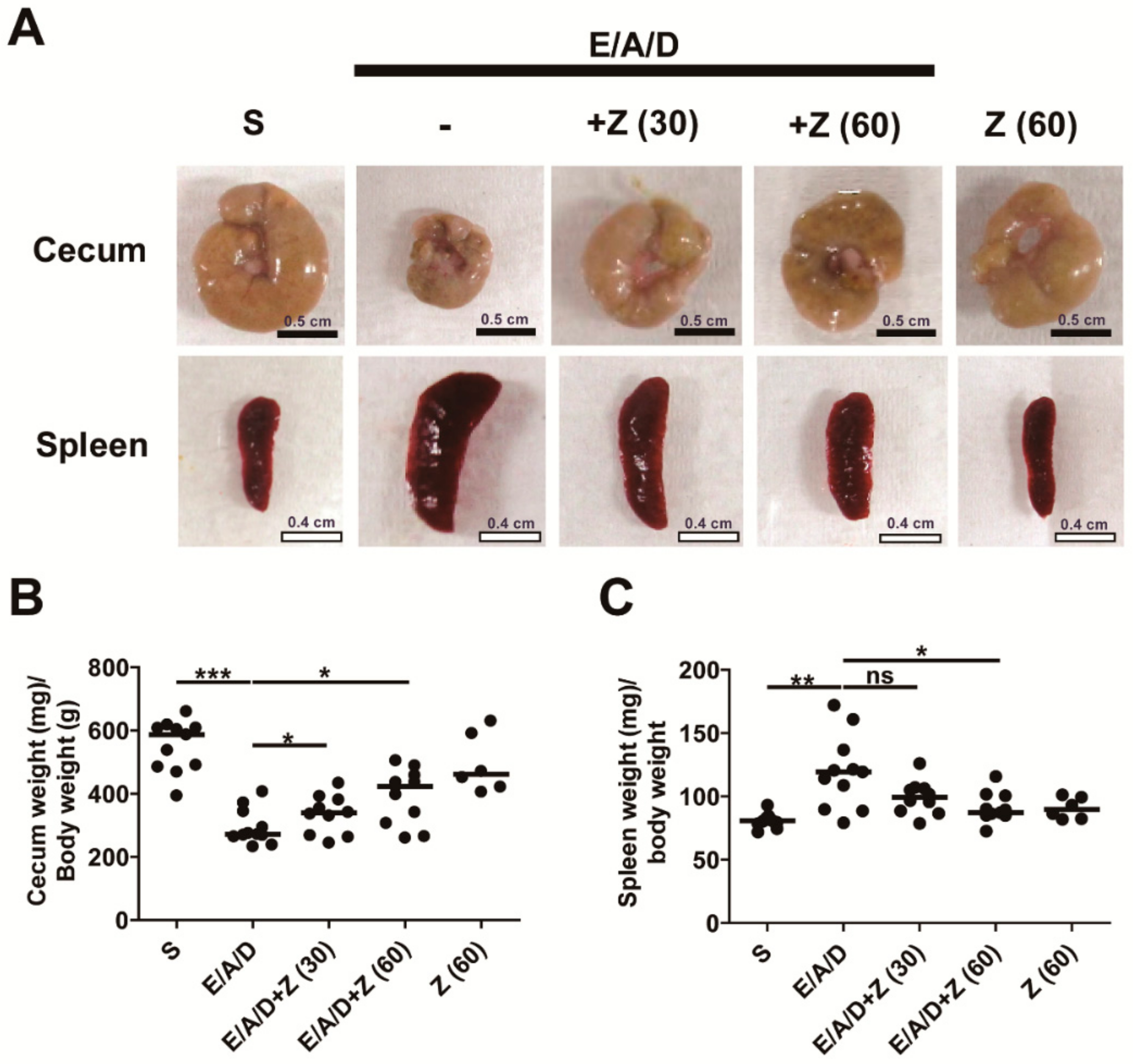
2.5. Zerumbone Prevented Cecum Weight Loss and Spleen Enlargement in ETBF-Colonized AOM/DSS Mice
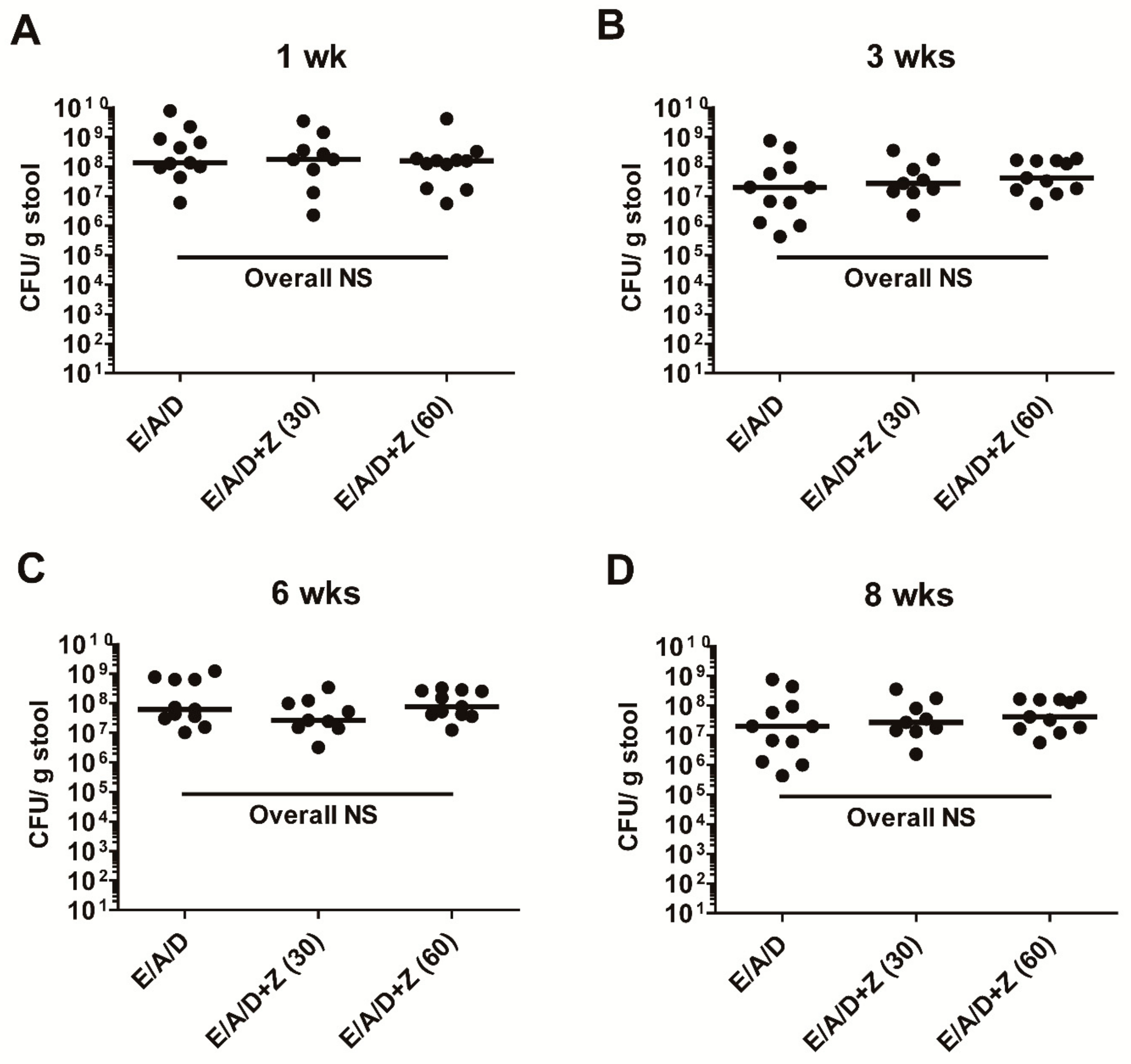
2.6. Zerumbone Does Not Affect ETBF Colonization in AOM/DSS Mice
3. Discussion
4. Materials and Methods
4.1. Bacteria Strains and Bacteria Enumeration in Stool
4.2. ETBF/AOM/DSS Mouse Model
4.3. Tumor Enumeration and Histopathology
4.4. Zerumbone Treatment in ETBF/AOM/DSS Mice
4.5. Evaluation of Inflammation and Hyperplasia
4.6. Statistics
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAI Score | Activity | Stool Consistency | Occult/Gross Bleeding |
|---|---|---|---|
| 0 | Normal | Normal | Normal |
| 1 | Moderately decreased activity | Slightly loose stool | Slightly hemoccult positive in stool |
| 2 | Severely decreased activity | Severely Loose stool | Hemoccult positive in stool and rectum |
| 3 | None | Diarrhea | Gross bleeding |
References
- Fleming, M.; Ravula, S.; Tatishchev, S.F.; Wang, H.L. Colorectal carcinoma: Pathologic aspects. J. Gastrointest. Oncol. 2012, 3, 153–173. [Google Scholar] [CrossRef]
- Gupta, R.B.; Harpaz, N.; Itzkowitz, S.; Hossain, S.; Matula, S.; Kornbluth, A.; Bodian, C.; Ullman, T. Histologic Inflammation Is a Risk Factor for Progression to Colorectal Neoplasia in Ulcerative Colitis: A cohort study. Gastroenterology 2007, 133, 1099–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mundy, L.M.; Sears, C.L. Detection of Toxin Production by Bacteroides fragilis: Assay Development and Screening of Extraintestinal Clinical Isolates. Clin. Infect. Dis. 1996, 23, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Meisel-Mikolajczyk, F.; Pituch, H.; Rouyan, G.S. Detection of Enterotoxigenic Bacteroides fragilis (ETBF), among Strains Isolated between 1976 and 1995 in Poland. Acta Microbiol. Pol. 1996, 45, 187–192. [Google Scholar] [PubMed]
- Jeon, J.I.; Ko, S.H.; Kim, J.M. Intestinal Epithelial Cells Exposed to Bacteroides fragilis Enterotoxin Regulate NF-kB Activation and Inflammatory Responses through b-Catenin Expression. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Lee, J.Y.; Yoon, Y.M.; Oh, Y.K.; Kang, J.S.; Kim, Y.J.; Kim, K.H. Bacteroides fragilis Enterotoxin Induces Cyclooxygenase-2 and Fluid Secretion in Intestinal Epithelial Cells through NF-kB Activation. Eur. J. Immunol. 2006, 36, 2446–2456. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lim, K.C.; Huang, J.; Saidi, R.F.; Sears, C.L. Bacteroides fragilis Enterotoxin Cleaves the Zonula Adherens Protein, E-cadherin. Proc. Natl. Acad. Sci. USA 1998, 95, 14979–14984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Powell, J.; Mathioudakis, N.; Kane, S.; Fernandez, E.; Sears, C.L. Bacteroides fragilis Enterotoxin Induces Intestinal Epithelial Cell Secretion of Interleukin-8 through Mitogen-Activated Protein Kinases and a Tyrosine Kinase-Regulated Nuclear Factor-kB Pathway. Infect. Immun. 2004, 72, 5832–5839. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Jo, M.; Hong, J.E.; Park, C.O.; Lee, C.G.; Yun, M.; Rhee, K.-J. Zerumbone Suppresses Enterotoxigenic Bacteroides fragilis Infection-Induced Colonic Inflammation through Inhibition of NF-kB. Int. J. Mol. Sci. 2019, 20, 4560. [Google Scholar] [CrossRef] [Green Version]
- Zamani, S.; Hesam Shariati, S.; Zali, M.R.; Asadzadeh Aghdaei, H.; Sarabi Asiabar, A.; Bokaie, S.; Nomanpour, B.; Sechi, L.A.; Feizabadi, M.M. Detection of Enterotoxigenic Bacteroides fragilis in Patients with Ulcerative Colitis. Gut Pathog. 2017, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Boleij, A.; Hechenbleikner, E.M.; Goodwin, A.C.; Badani, R.; Stein, E.M.; Lazarev, M.G.; Ellis, B.; Carroll, K.C.; Albesiano, E.; Wick, E.C.; et al. The Bacteroides fragilis Toxin Gene Is Prevalent in the Colon Mucosa of Colorectal Cancer Patients. Clin. Infect. Dis. 2015, 60, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Rhee, K.-J.; Wu, S.; Wu, X.; Huso, D.L.; Karim, B.; Franco, A.A.; Rabizadeh, S.; Golub, J.E.; Mathews, L.E.; Shin, J.; et al. Induction of Persistent Colitis by a Human Commensal, Enterotoxigenic Bacteroides fragilis, in Wild-Type C57BL/6 Mice. Infect. Immun. 2009, 77, 1708–1718. [Google Scholar] [CrossRef] [Green Version]
- Housseau, F.; Wu, S.; Wick, E.C.; Fan, H.; Wu, X.; Llosa, N.J.; Smith, K.N.; Tam, A.; Ganguly, S.; Wanyiri, J.W.; et al. Redundant Innate and Adaptive Sources of IL17 Production Drive Colon Tumorigenesis. Cancer Res. 2016, 76, 2115–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, E.C.; Rabizadeh, S.; Albesiano, E.; Wu, X.; Wu, S.; Chan, J.; Rhee, K.-J.; Ortega, G.; Huso, D.L.; Pardoll, D.; et al. Stat3 Activation in Murine Colitis Induced by Enterotoxigenic Bacteroides fragilis. Inflamm. Bowel Dis. 2014, 20, 821–834. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Rhee, K.-J.; Albesiano, E.; Rabizadeh, S.; Wu, X.; Yen, H.R.; Huso, D.L.; Brancati, F.L.; Wick, E.; McAllister, F.; et al. A Human Colonic Commensal Promotes Colon Tumorigenesis via Activation of T Helper Type 17 T cell Responses. Nat. Med. 2009, 15, 1016–1022. [Google Scholar] [CrossRef]
- Hwang, S.; Lee, C.G.; Jo, M.; Park, C.O.; Gwon, S.-Y.; Hwang, S.; Yi, H.C.; Lee, S.-Y.; Eom, Y.B.; Karim, B.; et al. Enterotoxigenic Bacteroides fragilis Infection Exacerbates Tumorigenesis in AOM/DSS Mouse Model. Int. J. Med. Sci. 2020, 17, 145–152. [Google Scholar] [CrossRef] [Green Version]
- DeStefano Shields, C.E.; Van Meerbeke, S.W.; Housseau, F.; Wang, H.; Huso, D.L.; Casero, R.A., Jr.; O’Hagan, H.M.; Sears, C.L. Reduction of Murine Colon Tumorigenesis Driven by Enterotoxigenic Bacteroides fragilis Using Cefoxitin Treatment. J. Infect. Dis. 2016, 214, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.H. Health Benefits of Fruit and Vegetables are from Additive and Synergistic Combinations of Phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef]
- Moreira da Silva, T.; Pinheiro, C.D.; Puccinelli Orlandi, P.; Pinheiro, C.C.; Soares Pontes, G. Zerumbone from Zingiber zerumbet (L.) smith: A Potential Prophylactic and Therapeutic Agent against the Cariogenic Bacterium Streptococcus mutans. BMC Complement. Altern. Med. 2018, 18, 301. [Google Scholar] [CrossRef] [Green Version]
- Santosh Kumar, S.C.; Srinivas, P.; Negi, P.S.; Bettadaiah, B.K. Antibacterial and Antimutagenic Activities of Novel Zerumbone Analogues. Food Chem. 2013, 141, 1097–1103. [Google Scholar] [CrossRef]
- Sulaiman, M.R.; Perimal, E.K.; Akhtar, M.N.; Mohamad, A.S.; Khalid, M.H.; Tasrip, N.A.; Mokhtar, F.; Zakaria, Z.A.; Lajis, N.H.; Israf, D.A. Anti-Inflammatory Effect of Zerumbone on Acute and Chronic Inflammation Models in Mice. Fitoterapia 2010, 81, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Yodkeeree, S.; Sung, B.; Limtrakul, P.; Aggarwal, B.B. Zerumbone Enhances TRAIL-Induced Apoptosis through the Induction of Death Receptors in Human Colon Cancer Cells: Evidence for an Essential Role of Reactive Oxygen Species. Cancer Res. 2009, 69, 6581–6589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitayama, T.; Yamamoto, K.; Utsumi, R.; Takatani, M.; Hill, R.K.; Kawai, Y.; Sawada, S.; Okamoto, T. Chemistry of Zerumbone. 2. Regulation of Ring Bond Cleavage and Unique Antibacterial Activities of Zerumbone Derivatives. Biosci. Biotechnol. 2001, 65, 2193–2199. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.S.; Cheon, S.Y.; Roh, S.S.; Lee, M.; An, H.J. Chemopreventive Effect of Aster glehni on Inflammation-Induced Colorectal Carcinogenesis in Mice. Nutrients 2018, 10, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washington, M.K.; Powell, A.E.; Sullivan, R.; Sundberg, J.P.; Wright, N.; Coffey, R.J.; Dove, W.F. Pathology of rodent models of intestinal cancer: Progress Report and Recommendations. Gastroenterology 2013, 144, 705–717. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.; Karin, E.; Terzic, J.; Mucida, D.; Yu, G.Y.; Vallabhapurapu, S.; Scheller, J.; Rose-John, S.; Cheroutre, H.; Eckmann, L.; et al. IL-6 and Stat3 Are Required for Survival of Intestinal Epithelial cells and Development of Colitis-Associated Cancer. Cancer Cell 2009, 15, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Fatima, A.; Abdul, A.B.; Abdullah, R.; Karjiban, R.A.; Lee, V.S. Binding Mode Analysis of Zerumbone to Key Signal Proteins in the Tumor Necrosis Factor Pathway. Int. J. Mol. Sci. 2015, 16, 2747–2766. [Google Scholar] [CrossRef]
- Takada, Y.; Murakami, A.; Aggarwal, B.B. Zerumbone Abolishes NF-kB and IkBa Kinase Activation Leading to Suppression of Antiapoptotic and Metastatic Gene Expression, Upregulation of Apoptosis, and Downregulation of Invasion. Oncogene 2005, 24, 6957–6969. [Google Scholar] [CrossRef] [Green Version]
- Girisa, S.; Shabnam, B.; Monisha, J.; Fan, L.; Halim, C.E.; Arfuso, F.; Ahn, K.S.; Sethi, G.; Kunnumakkara, A.B. Potential of Zerumbone as an Anti-Cancer Agent. Molecules 2019, 24, 734. [Google Scholar] [CrossRef] [Green Version]
- Deorukhkar, A.; Ahuja, N.; Mercado, A.L.; Diagaradjane, P.; Raju, U.; Patel, N.; Mohindra, P.; Diep, N.; Guha, S.; Krishnan, S. Zerumbone Increases Oxidative Stress in a Thiol-Dependent ROS-Independent Manner to Increase DNA Damage and Sensitize Colorectal Cancer Cells to Radiation. Cancer Med. 2015, 4, 278–292. [Google Scholar] [CrossRef]
- Kim, M.; Miyamoto, S.; Yasui, Y.; Oyama, T.; Murakami, A.; Tanaka, T. Zerumbone, a Tropical Ginger Sesquiterpene, Inhibits Colon and Lung Carcinogenesis in Mice. Int. J. Cancer 2009, 124, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Dermani, F.K.; Amini, R.; Saidijam, M.; Pourjafar, M.; Saki, S.; Najafi, R. Zerumbone Inhibits Epithelial-Mesenchymal Transition and Cancer Stem Cells Properties by Inhibiting the b-catenin Pathway through miR-200c. J. Cell. Physiol. 2018, 233, 9538–9547. [Google Scholar] [CrossRef] [PubMed]
- Jantan, I.; Haque, M.A.; Ilangkovan, M.; Arshad, L. Zerumbone from Zingiber zerumbet Inhibits Innate and Adaptive Immune Responses in Balb/C Mice. Int. Immunopharmacol. 2019, 73, 552–559. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.; Jo, M.; Hong, J.E.; Park, C.O.; Lee, C.G.; Rhee, K.-J. Protective Effects of Zerumbone on Colonic Tumorigenesis in Enterotoxigenic Bacteroides fragilis (ETBF)-Colonized AOM/DSS BALB/c Mice. Int. J. Mol. Sci. 2020, 21, 857. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030857
Hwang S, Jo M, Hong JE, Park CO, Lee CG, Rhee K-J. Protective Effects of Zerumbone on Colonic Tumorigenesis in Enterotoxigenic Bacteroides fragilis (ETBF)-Colonized AOM/DSS BALB/c Mice. International Journal of Molecular Sciences. 2020; 21(3):857. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030857
Chicago/Turabian StyleHwang, Soonjae, Minjeong Jo, Ju Eun Hong, Chan Oh Park, Chang Gun Lee, and Ki-Jong Rhee. 2020. "Protective Effects of Zerumbone on Colonic Tumorigenesis in Enterotoxigenic Bacteroides fragilis (ETBF)-Colonized AOM/DSS BALB/c Mice" International Journal of Molecular Sciences 21, no. 3: 857. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030857