Functional Analysis of Long Non-Coding RNAs Reveal Their Novel Roles in Biocontrol of Bacteria-Induced Tomato Resistance to Meloidogyne incognita

 and
and
Abstract
:1. Introduction
2. Results
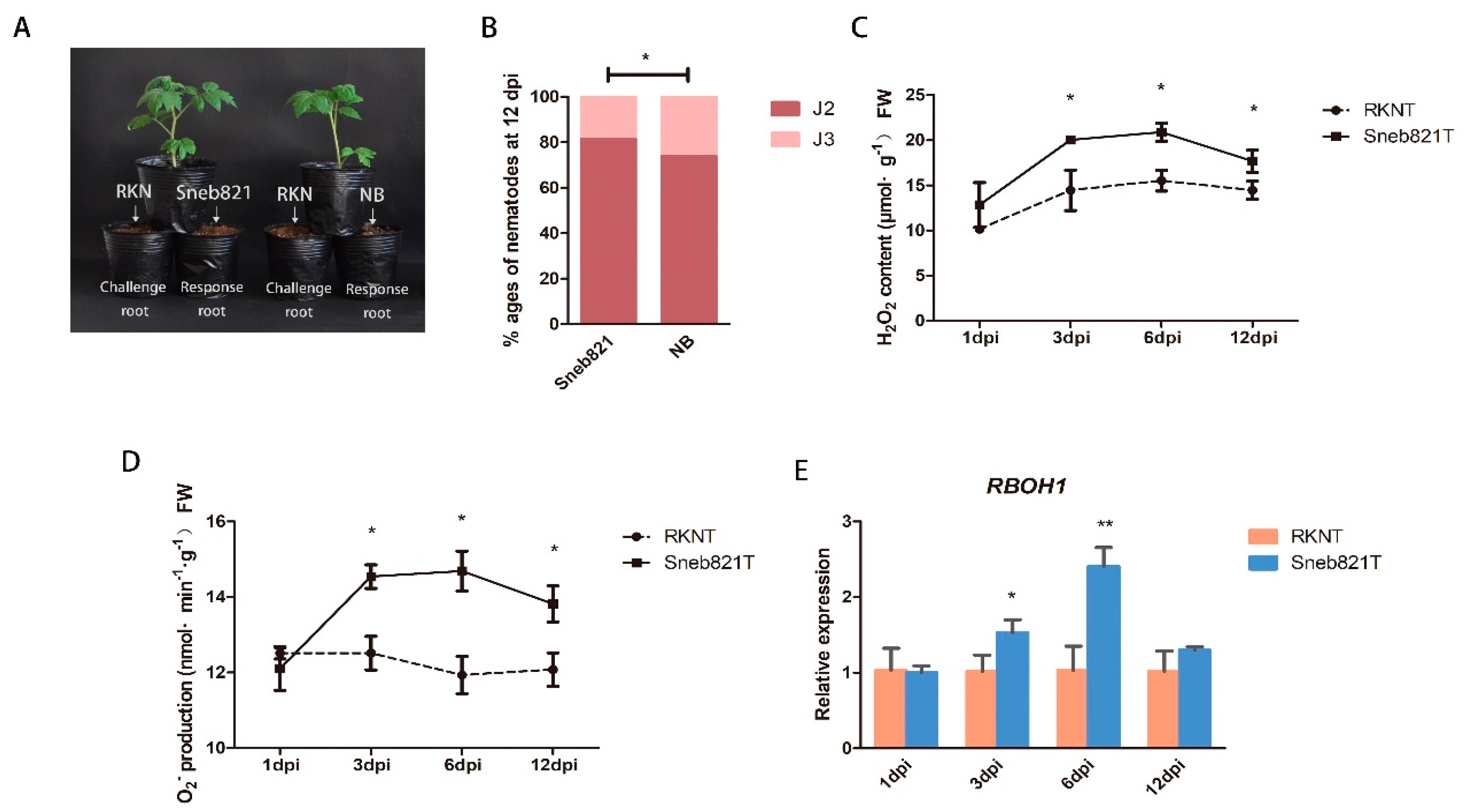
2.1. Inducement Mechanisms of Sneb821
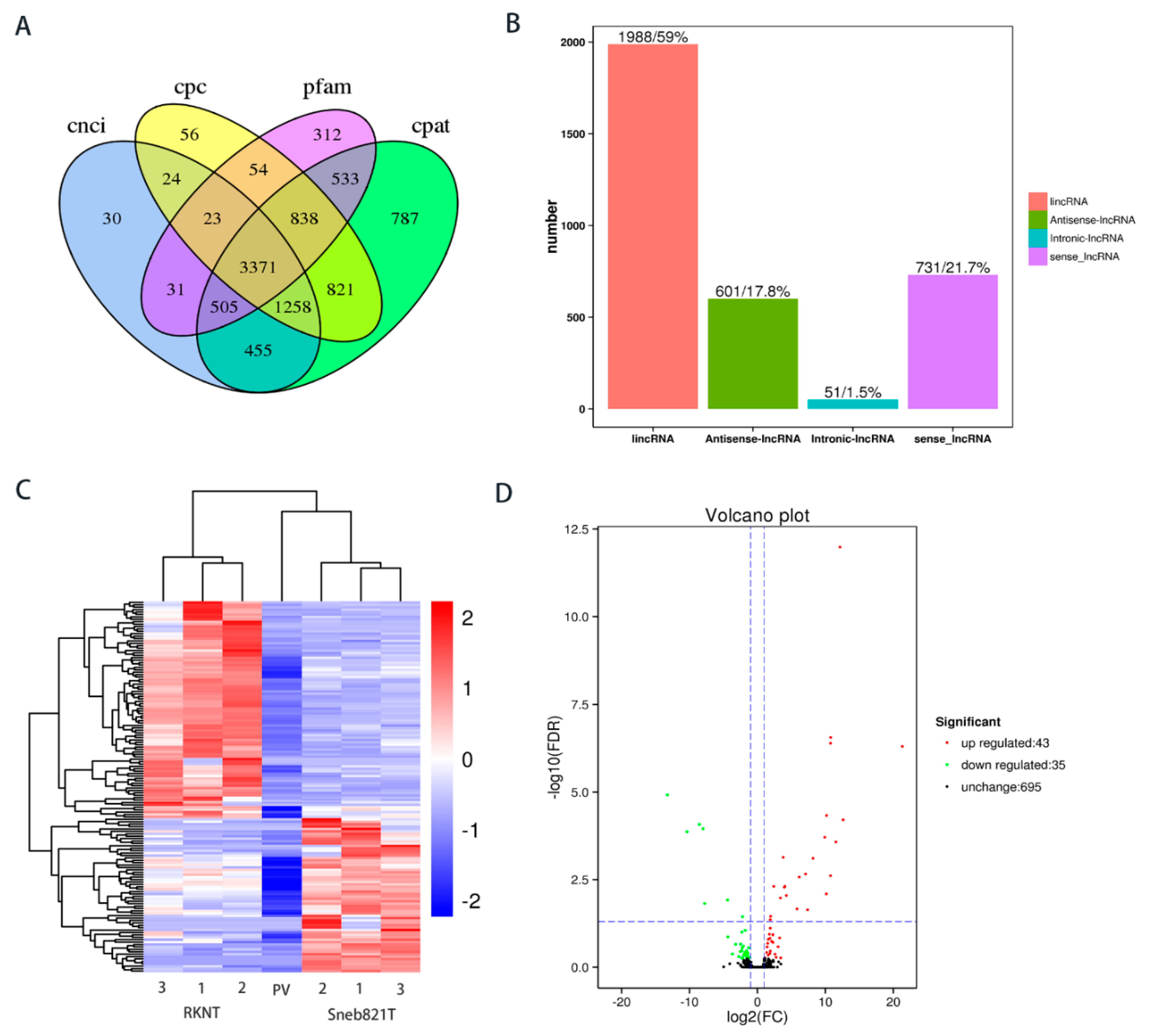
2.2. Sequencing and Characteristics of Transcripts
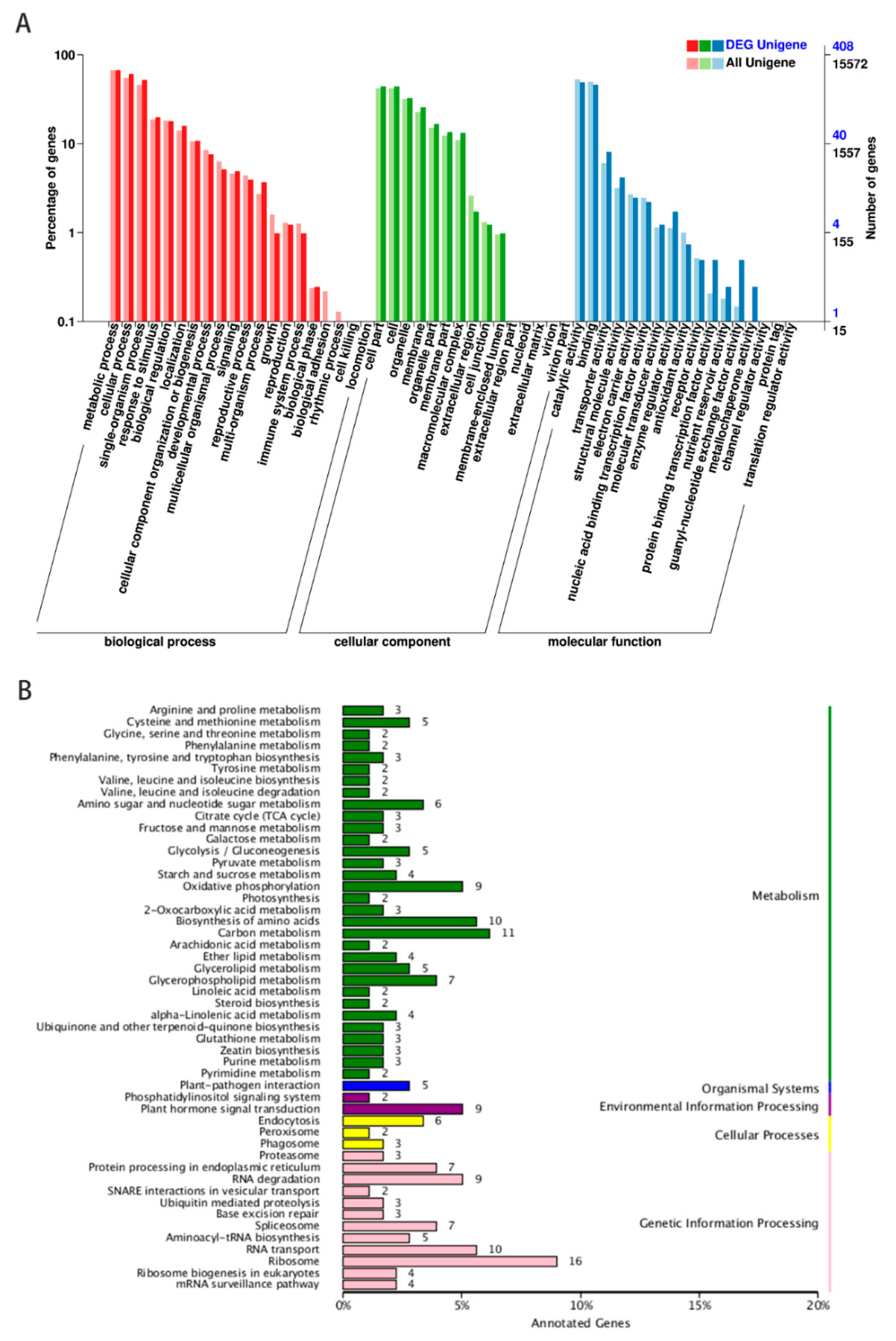
2.3. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis
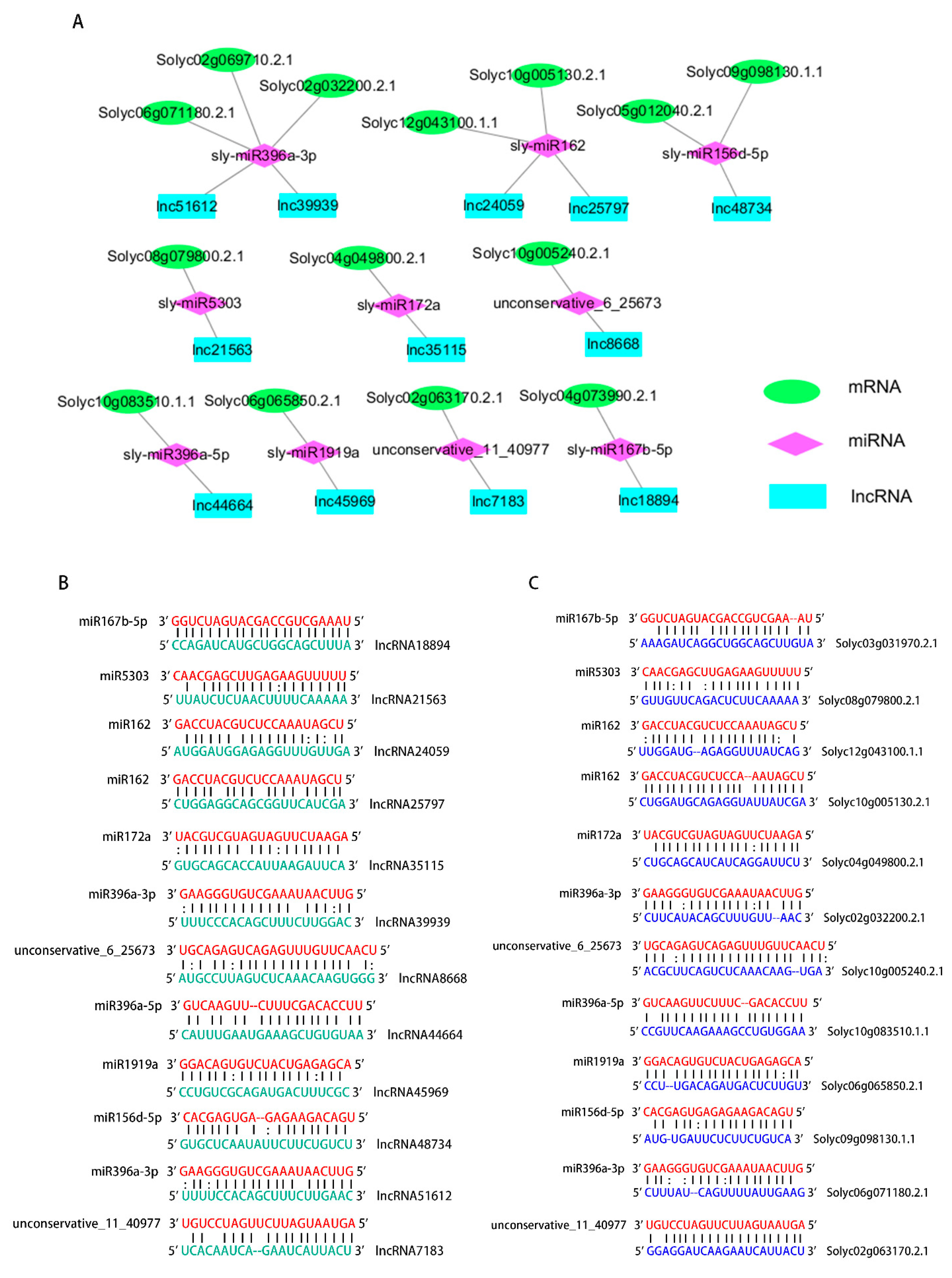
2.4. Construction of the CeRNA Network and Predicted Interaction between lncRNAs, miRNAs, and mRNAs
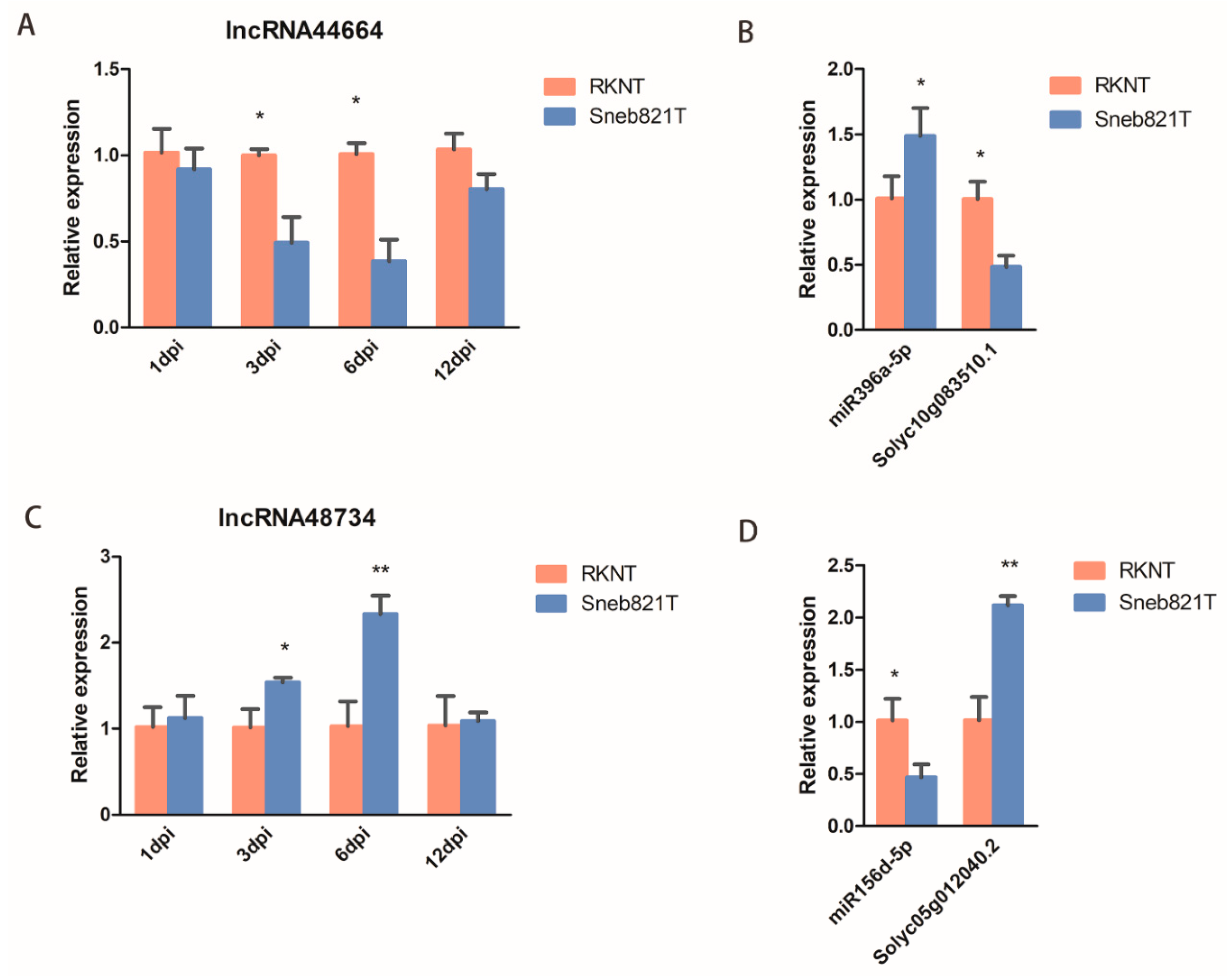
2.5. Verification of Differentially Expressed lncRNAs
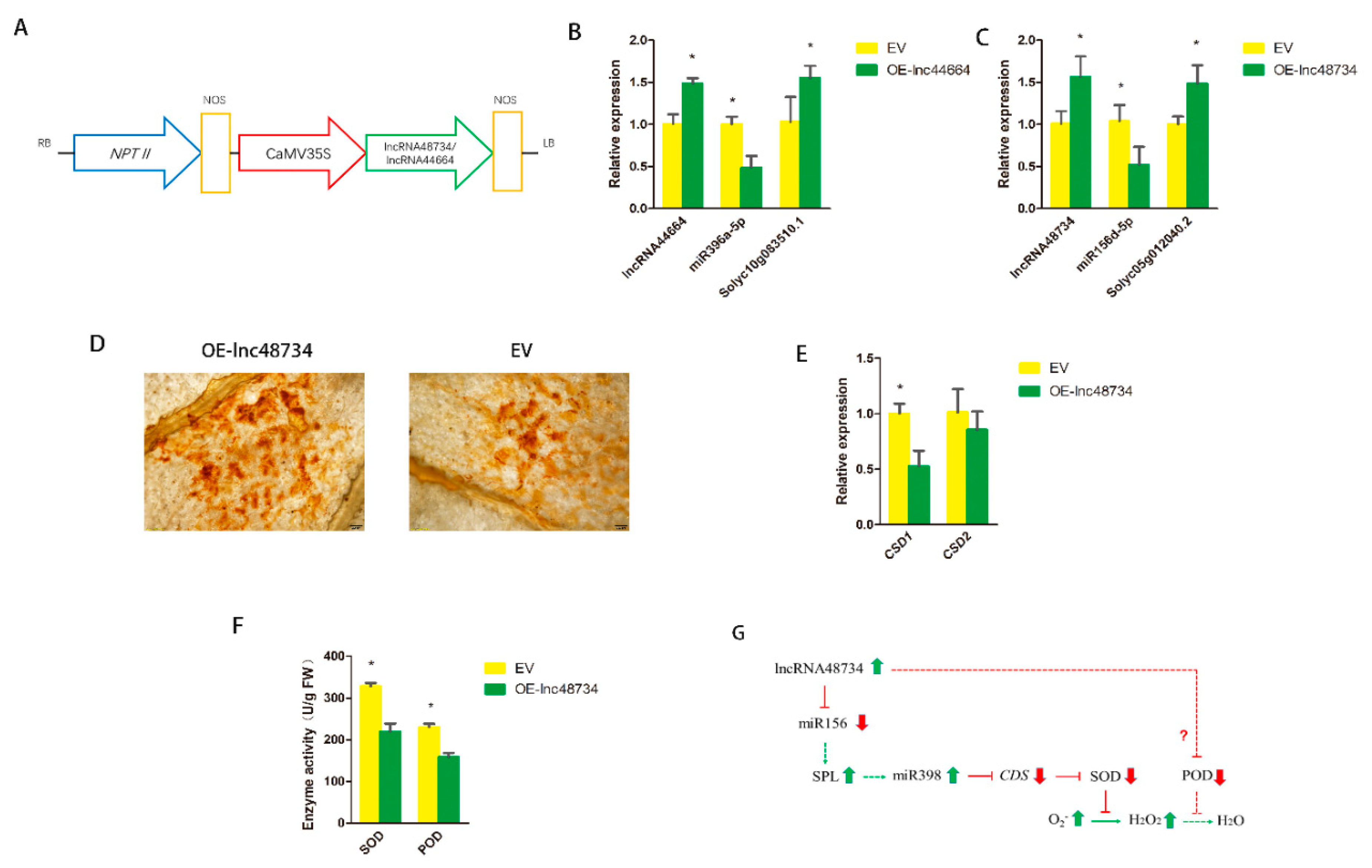
2.6. Function Verification of lncRNAs
3. Discussion
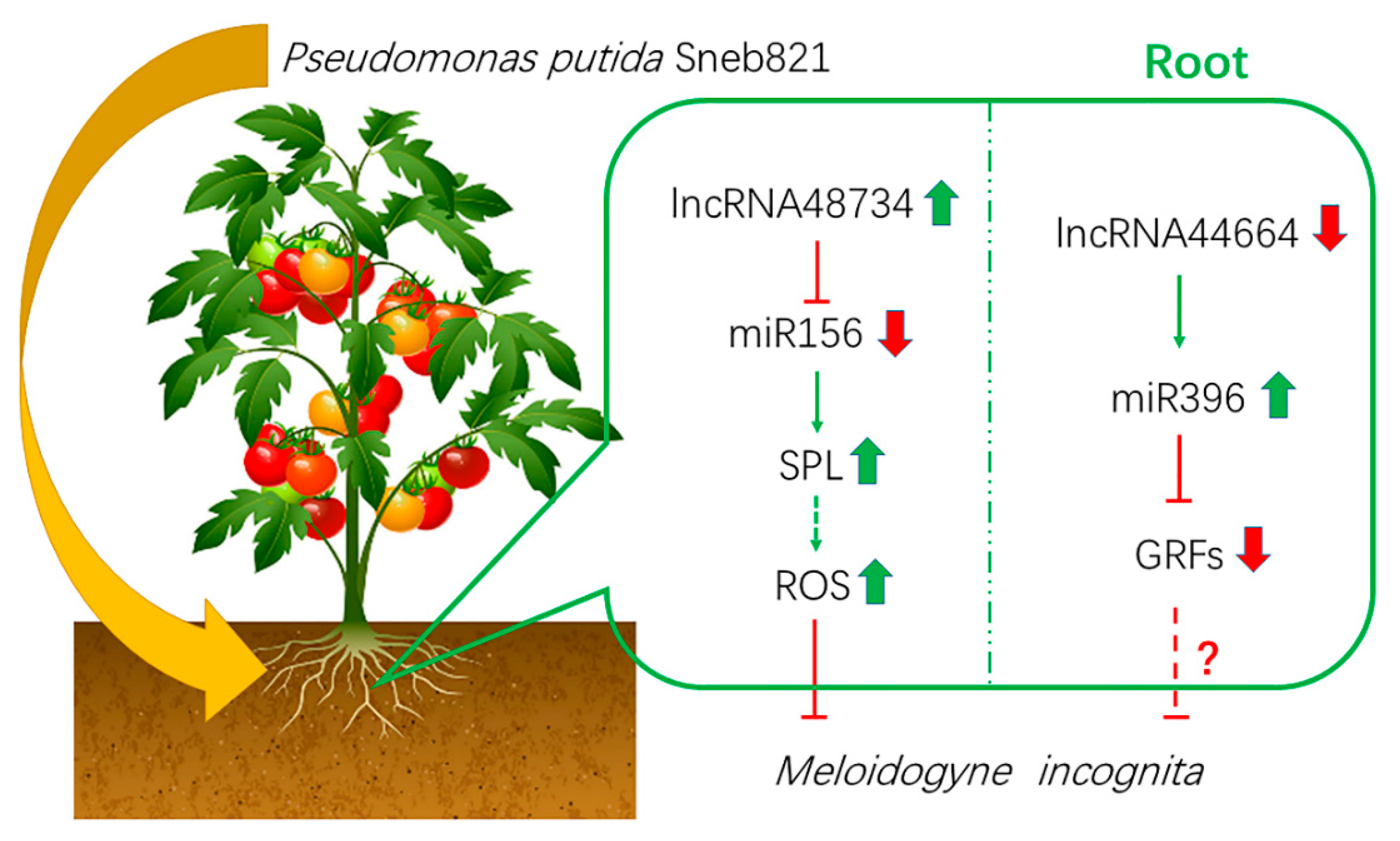
3.1. ROS Participate in Sneb821-induced Tomato Resistance to M. incognita
3.2. Certain lncRNAs Participate in Sneb821-induced Tomato Resistance to M. incognita
3.3. Inducement Resistance Function in lncRNAs through the ceRNA Network
3.4. lncRNA48734 and lncRNA44664 in Sneb821-induced Tomato Resistance to M. incognita
4. Materials and Methods
4.1. Plant Materials, Biocontrol Bacteria and Nematode Inoculum
4.2. RNA Extraction and Library Construction
4.3. lncRNA Analysis
4.4. RT-PCR Validation
4.5. Quantitative Real-Time PCR Validation
4.6. Prediction of lncRNA Targets and miRNA eTMs from mRNA
4.7. ceRNA Network Analysis
4.8. GO and KEGG Pathway Analysis
4.9. Transient Overexpression and Agrobacteria Infiltration
4.10. ROS Level Determination and DAB Staining
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| lncRNA | Long non-coding RNA |
| siRNA | Small interfering RNA |
| miRNA | MicroRNA |
| tasiRNA | Trans-acting siRNA |
| RKN | Root-knot nematode |
| GRF | Growth-regulating factors |
| SPL | Squamosal promoter-binding protein-like |
| ORF | Open reading frame |
| JA | Jasmonate acid |
| TOE1 | Target of early activation tagged 1 |
| FT | Flowering locus T |
| PGPR | Plant growth-promoting rhizobacteria |
| ROS | Reactive oxygen species |
| ceRNA | Competing endogenous RNA |
| TCS | Two-component system |
| J2 | Second-stage juvenile |
| J3 | Third-stage juvenile |
| dpi | Days post-inoculation |
| GUS | β-Glucuronidase |
| NB | Nutrient broth |
| qRT-PCR | Quantitative real-time PCR |
| GO | Gene ontology |
| KEGG | Kyoto encyclopedia of genes and genomes |
| CaMV | Cauliflower mosaic virus |
| SOD | Superoxide |
| POD | Peroxidase |
| DAB | Diaminobenzidine |
| eTMs | Endogenous target mimics |
| CSD | Cu/Zn-superoxide dismutase |
| HR | Hypersensitive response |
References
- Castel, S.E.; Martienssen, R.A. RNA interference in the nucleus: Roles for small RNAs in transcription, epigenetics and beyond. Nat. Rev. Genet. 2013, 14, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Mohammad Ali, F.; Claes, W. Regulatory roles of natural antisense transcripts. Nat. Rev. Mol. Cell Biol. 2009, 10, 637–643. [Google Scholar]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Eun-Deok, K.; Sibum, S. Long noncoding RNA: Unveiling hidden layer of gene regulatory networks. Trends Plant Sci. 2012, 17, 16–21. [Google Scholar]
- Cuperus, J.T.; Noah, F.; Carrington, J.C. Evolution and functional diversification of MIRNA genes. Plant Cell 2011, 23, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Simon, S.A.; Meyers, B.C. Small RNA-mediated epigenetic modifications in plants. Curr. Opin. Plant Biol. 2011, 14, 148–155. [Google Scholar] [CrossRef]
- Eddy, S.R. Non-coding RNA genes and the modern RNA world. Nat. Rev. Genet. 2001, 2, 919–929. [Google Scholar] [CrossRef]
- Mercer, T.R.; Mattick, J.S. Structure and function of long noncoding RNAs in epigenetic regulation. Nat. Struct. Mol. Biol. 2013, 20, 300–307. [Google Scholar] [CrossRef]
- Cui, J.; Jiang, N.; Meng, J.; Yang, G.; Liu, W.; Zhou, X.; Ma, N.; Hou, X.; Luan, Y. LncRNA33732-respiratory burst oxidase module associated with WRKY1 in tomato-Phytophthora infestans interactions. Plant J. 2018, 97, 933–946. [Google Scholar] [CrossRef]
- Jiang, N.; Cui, J.; Shi, Y.; Yang, G.; Zhou, X.; Hou, X.; Meng, J.; Luan, Y. Tomato lncRNA23468 functions as a competing endogenous RNA to modulate NBS-LRR genes by decoying miR482b in the tomato-Phytophthora infestans interaction. Hortic. Res. 2019, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Trudgill, D.L.; Blok, V.C. Apomictic, polyphagous root-knot nematodes: Exceptionally successful and damaging biotrophic root pathogens. Annu. Rev. Phytopathol. 2001, 39, 53. [Google Scholar] [CrossRef] [PubMed]
- Adam, M.; Westphal, A.; Hallmann, J.; Heuer, H. Specific microbial attachment to root knot nematodes in suppressive soil. Appl. Environ. Microbiol. 2014, 80, 2679–2686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Zhao, H.; Zhao, D.; Zhu, X.; Wang, Y.; Duan, Y.; Xuan, Y.; Chen, L. Isolation and identification of bacteria from rhizosphere soil and their effect on plant growth promotion and root-knot nematode disease. Biol. Control 2018, 119, 12–19. [Google Scholar] [CrossRef]
- Zhao, W.; Li, Z.; Fan, J.; Hu, C.; Yang, R.; Qi, X.; Chen, H.; Zhao, F.; Wang, S. Identification of jasmonic acid-associated microRNAs and characterization of the regulatory roles of the miR319/TCP4 module under root-knot nematode stress in tomato. J. Exp. Bot. 2015, 66, 4653–4667. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.; Rocha, M.; Magliano, M.; Ratpopoulo, A.; Revel, B.; Marteu, N.; Magnone, V.; Lebrigand, K.; Cabrera, J.; Barcala, M. Characterization of microRNAs from Arabidopsis galls highlights a role for miR159 in the plant response to the root-knot nematode Meloidogyne incognita. New Phytol. 2017, 216, 882–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, J.; Barcala, M.; García, A.; Rio-Machín, A.; Medina, C.; Jaubert-Possamai, S.; Favery, B.; Maizel, A.; Ruiz-Ferrer, V.; Fenoll, C. Differentially expressed small RNAs in Arabidopsis galls formed by Meloidogyne javanica: A functional role for miR390 and its TAS3-derived tasiRNAs. New Phytol. 2016, 209, 1625–1640. [Google Scholar] [CrossRef] [Green Version]
- Dã-Az-Manzano, F.E.; Cabrera, J.; Ripoll, J.J.; Del, O.I.; Andrãs, M.F.; Silva, A.C.; Barcala, M.; Sã, N.M.; Ruã-Z-Ferrer, V.; De, A.-E.J. A role for the gene regulatory module microRNA172/TARGET OF EARLY ACTIVATION TAGGED 1/FLOWERING LOCUS T (miRNA172/TOE1/FT) in the feeding sites induced by Meloidogyne javanica in Arabidopsis thaliana. New Phytol. 2018, 217, 813–827. [Google Scholar] [CrossRef] [Green Version]
- Hua-Jun, W.; Zhi-Min, W.; Meng, W.; Xiu-Jie, W. Widespread long noncoding RNAs as endogenous target mimics for microRNAs in plants. Plant Physiol. 2013, 161, 1875–1884. [Google Scholar]
- Rabhi, N.E.H.; Silini, A.; Cherif-Silini, H.; Yahiaoui, B.; Sanchez, L. Pseudomonas knackmussii MLR6, a rhizospheric strain isolated from halophyte, enhances salt tolerance in Arabidopsis thaliana. J. Appl. Microbiol. 2018, 125, 1836–1851. [Google Scholar] [CrossRef]
- Chambonnier, G.; Roux, L.; Redelberger, D.; Fadel, F.; Filloux, A.; Sivaneson, M.; De, B.S.; Bordi, C. The hybrid histidine kinase lads forms a multicomponent signal transduction system with the GacS/GacA two-component system in Pseudomonas aeruginosa. PLoS Genet. 2016, 12, e1006032. [Google Scholar] [CrossRef] [Green Version]
- Katiyaragarwal, S.; Morgan, R.; Dahlbeck, D.; Borsani, O.; Jr, V.A.; Zhu, J.K.; Staskawicz, B.J.; Jin, H. A pathogen-inducible endogenous siRNA in plant immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 18002–18007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Functional interactions among microRNAs and long noncoding RNAs. Semin. Cell Dev. Biol. 2014, 34, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuai, P.; Liang, D.; Tang, S.; Zhang, Z.; Ye, C.Y.; Su, Y.; Xia, X.; Yin, W. Genome-wide identification and functional prediction of novel and drought-responsive lincRNAs in Populus trichocarpa. J. Exp. Bot. 2014, 65, 4975–4983. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Wang, D.; He, R.; Liu, S.; Zhu, J.A. Mutations in MIR396e and MIR396f increase grain size and modulate shoot architecture in rice. Plant Biotechnol. J. 2019, 18, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of plant microRNA targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H.; Hayashi, M.; Fukazawa, M.; Kobayashi, Y.; Shikanai, T. SQUAMOSA promoter binding protein–Like7 is a central regulator for copper homeostasis in Arabidopsis. Plant Cell 2009, 21, 347–361. [Google Scholar] [CrossRef] [Green Version]
- Axtell, M.J.; Snyder, J.A.; Bartel, D.P. Common functions for diverse small RNAs of land plants. Plant Cell 2007, 19, 1750–1769. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2000, 7, 405–410. [Google Scholar] [CrossRef]
- Yamashita, A.; Shichino, Y.; Yamamoto, M. The long non-coding RNA world in yeasts. Biochim Biophys Acta 2016, 1859, 147–154. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, X.; Wang, C.; Xu, Z.; Wang, Y.; Liu, X.; Kang, Z.; Ji, W. Long non-coding genes implicated in response to stripe rust pathogen stress in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2013, 40, 6245–6253. [Google Scholar] [CrossRef]
- Cui, J.; Luan, Y.; Jiang, N.; Bao, H.; Meng, J. Comparative transcriptome analysis between resistant and susceptible tomato allows the identification of lncRNA16397 conferring resistance to Phytophthora infestans by co-expressing glutaredoxin. Plant J. 2017, 89, 577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Zhou, Y.-F.; Feng, Y.Z.; He, H.; Lian, J.P.; Yang, Y.W.; Lei, M.Q.; Zhang, Y.C.; Chen, Y.Q. Transcriptional landscape of pathogen-responsive lncRNAs in rice unveils the role of ALEX1 in jasmonate pathway and disease resistance. Plant Biotechnol. J. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yuwen, Y.; Lamei, J.; Xitie, L.; Tingli, L.; Tianzi, C.; Yinghua, J.; Wengui, Y.; Baolong, Z. Re-analysis of long non-coding RNAs and prediction of circRNAs reveal their novel roles in susceptible tomato following TYLCV infection. Bmc Plant Biol. 2018, 18, 104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.C.; Liao, J.Y.; Li, Z.Y.; Yu, Y.; Zhang, J.P.; Li, Q.F.; Qu, L.H.; Shu, W.S.; Chen, Y.Q. Genome-wide screening and functional analysis identify a large number of long noncoding RNAs involved in the sexual reproduction of rice. Genome Biol. 2014, 15, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewezi, T.; Howe, P.; Maier, T.R.; Baum, T.J. Arabidopsis small RNAs and their targets during cyst nematode parasitism. Mol. Plant Microbe Interact. 2008, 21, 1622–1634. [Google Scholar] [CrossRef] [Green Version]
- Hewezi, T.; Maier, T.R.; Nettleton, D.; Baum, T.J. The Arabidopsis microRNA396-GRF1/GRF3 regulatory module acts as a developmental regulator in the reprogramming of root cells during cyst nematode infection. Plant Physiol. 2012, 159, 321–335. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Yi, L.; Ji, W. Long non-coding RNA expression profiling of mouse testis during postnatal development. PLoS ONE 2013, 8, e75750. [Google Scholar]
- Lei, K.; Yong, Z.; Zhi-Qiang, Y.; Xiao-Qiao, L.; Shu-Qi, Z.; Liping, W.; Ge, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, 345–349. [Google Scholar]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔCT) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xinbin, D.; Patrick Xuechun, Z. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, 155. [Google Scholar]
- Hua-Jun, W.; Ying-Ke, M.; Tong, C.; Meng, W.; Xiu-Jie, W. PsRobot: A web-based plant small RNA meta-analysis toolbox. Nucleic Acids Res. 2012, 40, 22–28. [Google Scholar]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Xu, P.; Meng, J.; Li, J.; Jiang, N.; Luan, Y. Transcriptome signatures of tomato leaf induced by Phytophthora infestans and functional identification of transcription factor Sp WRKY3. Theor. Appl. Genet. 2017, 131, 1–14. [Google Scholar]
- Iberkleid, I.; Yeheskel, A.; Belausov, E.; Chinnapandi, B.; Fitoussi, N.; Miyara, S.B. Structural characterization and localization analysis of the root-knot nematode Meloidogyne javanica fatty acid and retinol binding protein (Mj-FAR-1). Eur. J. Plant Pathol. 2015, 143, 133–149. [Google Scholar] [CrossRef]
- Kong, Q.; Wu, W.; Fan, Y.; Yan, Y.; Huang, Y. Distinct different expression of Th17 and Th9 cells in coxsackie virus B3-induced mice viral myocarditis. Virol. J. 2011, 8, 267. [Google Scholar]
- Lee, B.-H. A mitochondrial complex I defect Impairs cold-regulated nuclear gene expression. Plant Cell Online 2002, 14, 1235–1251. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Reads | Mapped Reads | Mapped Ratio |
|---|---|---|---|
| RKNT 1 | 123068008 | 108742693 | 88.36% |
| RKNT 2 | 118020220 | 105048103 | 89.01% |
| RKNT 3 | 129145372 | 114727340 | 88.84% |
| Sneb821T1 | 115966444 | 104203875 | 89.86% |
| Sneb821T 2 | 126689802 | 114918861 | 90.71% |
| Sneb821T 3 | 109378368 | 99030869 | 90.54% |
| lncRNA | mRNA | GO Terms | KEGG Pathway |
|---|---|---|---|
| lncRNA18894 | Solyc04g073990.2.1 | transferase activity | - |
| lncRNA21563 | Solyc08g079800.2.1 | - | plant hormone signal transduction |
| lncRNA24059 | Solyc12g043100.1.1 | - | endocytosis |
| lncRNA25797 | Solyc10g005130.2.1 | DNA binding | - |
| lncRNA35115 | Solyc04g049800.2.1 | transcription cofactor activity | - |
| lncRNA39939 | Solyc02g069710.2.1 | - | ribosome |
| Solyc02g032200.2.1 | - | plant-pathogen interaction | |
| lncRNA8668 | Solyc10g005240.2.1 | - | biosynthesis of amino acids |
| lncRNA44664 | Solyc10g083510.1.1 | cell proliferation | - |
| lncRNA45969 | Solyc06g065850.2.1 | - | biosynthesis of amino acids |
| lncRNA48734 | Solyc05g012040.2.1 | - | oxidative phosphorylation |
| Solyc09g098130.1.1 | - | plant-pathogen interaction | |
| lncRNA51612 | Solyc06g071180.2.1 | - | spliceosome |
| lncRNA7183 | Solyc02g063170.2.1 | cell death | - |
| lncRNA | miRNA | mRNA | Annotation |
|---|---|---|---|
| lncRNA18894 | miR167b-5p | Solyc04g073990.2.1 | Annexin |
| lncRNA21563 | miR5303 | Solyc08g079800.2.1 | Growth-regulating factor 12 |
| lncRNA24059 | miR162 | Solyc12g043100.1.1 | Pentatricopeptide repeat-containing protein |
| lncRNA25797 | miR162 | Solyc10g005130.2.1 | Dicer double-stranded RNA-binding fold |
| lncRNA35115 | miR172a | Solyc04g049800.2.1 | AP2-like ethylene-responsive transcription factor |
| lncRNA39939 | miR396a-3p | Solyc02g069710.2.1 | Sodium/calcium exchanger protein |
| Solyc02g032200.2.1 | Tir-lrr, resistance protein fragment | ||
| lncRNA8668 | unconservative_6_25673 | Solyc10g005240.2.1 | Transcription factor Myb |
| lncRNA44664 | miR396a-5p | Solyc10g083510.1.1 | Growth-regulating factor 2 |
| lncRNA45969 | miR1919a | Solyc06g065850.2.1 | Flotillin domain protein |
| lncRNA48734 | miR156d-5p | Solyc05g012040.2.1 | Squamosa promoter binding protein 3 |
| Solyc09g098130.1.1 | Cc-nbs-lrr, resistance protein | ||
| lncRNA51612 lncRNA7183 | miR396a-3p unconservative_11_40977 | Solyc06g071180.2.1 Solyc02g063170.2.1 | Dynein light chain 1 cytoplasmic Homology |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Zhao, D.; Fan, H.; Zhu, X.; Wang, Y.; Liu, X.; Duan, Y.; Xuan, Y.; Chen, L. Functional Analysis of Long Non-Coding RNAs Reveal Their Novel Roles in Biocontrol of Bacteria-Induced Tomato Resistance to Meloidogyne incognita. Int. J. Mol. Sci. 2020, 21, 911. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030911
Yang F, Zhao D, Fan H, Zhu X, Wang Y, Liu X, Duan Y, Xuan Y, Chen L. Functional Analysis of Long Non-Coding RNAs Reveal Their Novel Roles in Biocontrol of Bacteria-Induced Tomato Resistance to Meloidogyne incognita. International Journal of Molecular Sciences. 2020; 21(3):911. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030911
Chicago/Turabian StyleYang, Fan, Dan Zhao, Haiyan Fan, Xiaofeng Zhu, Yuanyuan Wang, Xiaoyu Liu, Yuxi Duan, Yuanhu Xuan, and Lijie Chen. 2020. "Functional Analysis of Long Non-Coding RNAs Reveal Their Novel Roles in Biocontrol of Bacteria-Induced Tomato Resistance to Meloidogyne incognita" International Journal of Molecular Sciences 21, no. 3: 911. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030911