Peptidyl Arginine Deiminase 2 (PADI2)-Mediated Arginine Citrullination Modulates Transcription in Cancer


1
Gene Regulation, Stem Cells and Cancer Program, Centre for Genomic Regulation (CRG), The Barcelona Institute of Science and Technology (BIST), Dr. Aiguader 88, 08003 Barcelona, Spain
2
Universitat Pompeu Fabra (UPF), 08003 Barcelona, Spain
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(4), 1351; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041351
Submission received: 17 January 2020
/
Revised: 10 February 2020
/
Accepted: 11 February 2020
/
Published: 17 February 2020
(This article belongs to the Special Issue Peptidylarginine Deiminases and Protein Deimination in Health and Disease)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Protein arginine deimination leading to the non-coded amino acid citrulline remains a key question in the field of post-translational modifications ever since its discovery by Rogers and Simmonds in 1958. Citrullination is catalyzed by a family of enzymes called peptidyl arginine deiminases (PADIs). Initially, increased citrullination was associated with autoimmune diseases, including rheumatoid arthritis and multiple sclerosis, as well as other neurological disorders and multiple types of cancer. During the last decade, research efforts have focused on how citrullination contributes to disease pathogenesis by modulating epigenetic events, pluripotency, immunity and transcriptional regulation. However, our knowledge regarding the functional implications of citrullination remains quite limited, so we still do not completely understand its role in physiological and pathological conditions. Here, we review the recently discovered functions of PADI2-mediated citrullination of the C-terminal domain of RNA polymerase II in transcriptional regulation in breast cancer cells and the proposed mechanisms to reshape the transcription regulatory network that promotes cancer progression.
1. Introduction
Deimination or citrullination as first described in 1958 by Rogers and Simmonds is a post-translational conversion of peptidyl-arginine to non-coded peptidyl-citrulline catalyzed by Ca2+- dependent peptidyl arginine deiminase (PADI) enzymes [1,2,3,4]. Citrullination produces a loss of a positive charge, which increases the mass by 0.984 Da [5], and the acidity of the amino acid side chain, as the iso-electric point (pI) changes from 11.41 for arginine to 5.91 for citrulline [6]. Studies in human embryonic kidney (HEK-293) cells found hundreds of proteins that can undergo citrullination, in which 25% of total citrullinated sites contained arginine-glycine-glycine (RGG)/arginine-glycine (RG) binding motifs [7]. These results highlighted that RNA-binding proteins can undergo citrullination to modulate the RNA processing. Citrullination also increases the hydrophobicity and conformation of proteins, which can potentially affect hydrogen bond formation, protein structure, protein-protein interactions and protein-nucleic acid interactions [7,8,9]. Given the significance of these interactions for essential cellular functions, citrullination may uniquely modulate cellular processes (Figure 1). The transformation in protein complexes can change the half-life, stability, and also expose proteins to degradation. For example, citrullination of filament aggregating protein that binds to keratin fibers in epithelial cells (filaggrin) leads to the partial unfolding of this protein, decreases their affinity for keratins, and, thereby, makes it more prone to degradation by several proteases including caspase 14 [10,11]. Additional evidence that citrullination affects the functional protein-protein interactions derives from observation in macrophage differentiation, where citrullination of plasminogen activator inhibitor-2 inhibits its binding ability to proteasome subunit beta type-1 to then modulate the inflammatory response [12].
Changes in citrullination occur in several pathological conditions such as autoimmune disorders and chronic inflammation-related diseases, including periodontitis, rheumatoid arthritis, atherosclerosis, and diabetes [13,14,15] (see Alghamdi, M. et al. [13] for a detailed review on the intrinsic role of citrullination in autoimmune disorders and Olsen, I. et al., [14] for a detailed review on citrullination as a plausible link to periodontitis, rheumatoid arthritis, atherosclerosis, and Alzheimer’s disease). Altered levels of citrullination have also been shown in neurological disorders and prion diseases, [16,17] and respiratory disorders [18,19]. Citrullination can generate new epitopes, which can produce new autoantigens linked to altered immune responses (see [20] for a review on citrullination and autoimmunity). The citrullination of histones in neutrophils facilitates neutrophil extracellular trap (NET) formation or NETosis, which is implicated in innate immune responses ([21]; see [22] for a detailed review on citrullination and NET). Remarkably, citrullination-directed chromatin decondensation leads to transcriptional firing in neutrophils that facilitates NET formation [23]. Moreover, recent work demonstrated that citrullination in macrophages induced inflammatory processes or the pyroptotic form of cell death, which is required for inflammasome assembly and proinflammatory interleukin-1β (IL-1β) release [24]. Moreover, citrullination contributes to cancer progression. Recent research seeks to elucidate how citrullination of several essential proteins drives multiple disease conditions. In this review, we focus on the role of PADI2-mediated arginine citrullination and their functional impact on transcriptional regulation and cancer progression.
2. Functional Role of PADI Family
Rogers and Taylor [25] discovered a previously unidentified enzymatic activity when generating peptidyl-citrulline in hair follicle extracts in 1977. The responsible enzymes were later identified as peptidyl arginine deiminases (PADI). The PADI family is a group of five calcium-dependent enzymes, PADI1, PADI2, PADI3, PADI4, and PADI6, encoded by the PADI loci in human [26,27] (Figure 2A). Among family members, PADI4 contains a canonical nuclear localization signal (NLS) and hence was long believed to be only PADI that can be localized to the nucleus [3,28]. Recently, work by the Thompson group had shown that PAD2 translocates into the nucleus in response to calcium signaling. They found that binding of calcium to PADI2 switches its binding from annexin 5 to the RanGTPase in the cytoplasm that promotes translocation of PAD2 into the nucleus [29]. Using a calcium-dependent process, a series of conformational changes eventually produce the correct movement of key active residues, including a free cysteine 645 into positions that are competent for catalysis [30]. PADI4 binds five calcium ions per monomer, and calcium binding upregulates enzymatic activity at least by 10, 000-fold without directly participating in catalysis [31]. In PADI2, calcium binding occurs in an ordered fashion at six calcium binding sites and induces a calcium switch (made of calcium binding sites 3, 4, and 5) that controls the overall calcium dependence of the enzyme [32]. Another important component for efficient PADI2 activity is the presence of a reducing environment that maintains the active site cysteine 645 free for catalysis of citrullination [26,27,33].
Human PADI genes are localized together as a cluster in a single topological associated domain (TAD) on chromosome 1 (Figure 2A), but most PADI gene members exhibit tissue-restricted expression. PADI1 is expressed in epidermis and uterus, PADI3 in epidermis and hair follicles, PADI4 in immune cells, brain, uterus and bone marrow, PADI6 in ovary, egg cells, embryo and testicles [34]. PADI2 is more widely expressed, including in the brain, uterus, spleen, breast, pancreas, skin, and skeletal muscles [3,6,34]. Their differential tissue expression patterns already suggest their unique and overlapping substrates in the genome. Human PADIs are homologous both within and between species (44−58% identity between human PADIs) [32]. Despite the high homology among family members, the five PADI enzymes have definite features, which facilitates the regulation of several distinct cellular processes. This feature may explain why altered levels of citrullination are associated with different pathological conditions, including multiple cancers [4,15,35,36,37,38,39,40,41]. Remarkably, PADI2 and PADI4 had distinct as well as overlapping substrates, for example, PADI4 directly citrullinates nuclear factor κB (NF-κB) p65 to promote its nuclear localization and transcriptional activity of key immune genes including IL-1β and tumor necrosis factor α (TNFα) to propagates inflammation in rheumatoid arthritis [42], whereas, mice with TNFα-induced inflammatory arthritis showed that PADI2 contributes to TNFα-induced citrullination, but not required for NETosis. However, the authors had not investigated the PADI2 substrates in this mice model [43]. These studies highlight the relevance of citrullination to propagate inflammatory microenvironment, most likely also involved in cancer cells. Importantly, an elevated citrullination level associated with various cancers and neurodegeneration had been linked to increased release of exosomes and microvesicles (EMVs), which are lipid bilayer-enclosed structures released from cells and participate in cell-to-cell communication [44]. The citrullination connection with EMV release could be coupled with the increased calcium influx. Thus, it is imperative to examine the mechanisms by which citrullination modulates the EMV release during cancer progression. One cellular conditions linked to PADI family members is in response to hypoxia, characterized by insufficient blood supply. In malignant glioma cells hypoxia-induced increase of PAD1, 2, 3, and 4 mRNA levels depends on hypoxia-inducible factor-1α (HIF1α) [45]. Similarly, fibroblast-like synoviocyte hypoxia in humans predominantly induces the expression of PADIs through HIF1α regulation [46]. One unresolved question is how the hypoxic conditions in growing tumors influence PADI activity and citrullination.
3. Impact of PADI Family on Histone Citrullination
PADIs catalyzed histone citrullination also controls transcription regulation and chromatin organization [47,48,49,50,51,52,53,54,55,56]. Remarkably, citrullinated histones found to account for approximately 10% of all histone molecules, emphasizing the essential role of this posttranslational modification in many physiological processes in the nucleus [48]. For example, during preimplantation and early-stage embryo development histone citrullination can lead to chromatin changes [49]. Specifically, PADI1 citrullinates both the histone H3 at 2, 8 and 17 positions (H3Cit2, 8, 17) and histone H4 at arginine 3 (H4Cit3) during 2- and 4-cell stage embryo development, which facilitates early embryo genome transactivation [50]. However, the potential implications of PADI1-directed histone citrullination had not been investigated yet. On the other hand, PADI2 can specificity citrullinates histone H3 arginine 26 citrullination (H3Cit26) that leads to chromatin decondensation and transcriptional activation in human breast cancer cells [51,52]. PADI2 can also catalyze H3Cit2, 8, 17 in mammary epithelial cells to regulate the expression of lactation related genes during diestrus [54]. PADI4 can generate H3Cit2, 8, 17 with higher efficiency than PADI2 and also generate H4Cit3 and H3Cit2, 8, 17 to control chromatin organization and gene expression during key cellular processes [47,54]. Notably, PADI4-mediated citrullination of linker histone H1 regulates stem cell pluripotency during early embryogenesis [55,56].
Citrullination of histone H3 can promote oncogenesis by repressing the expression of tumor suppressor miRNAs, which ultimately increases oncogene mRNAs in somatolactotrope cells [57] (see Section 5). Histone citrullination modifies gene regulation by modulating chromatin condensation at several levels. For example, PADI4-mediated citrullination of histone H3 at arginine 8 (H3Cit8) leads to the eviction of heterochromatin protein 1 α (HP1α) from the chromatin at regulatory regions of human endogenous retroviruses (HERVs) and cytokines, which generates a chromatin accessible state and promotes their gene transcription [58]. These findings provided the first evidence that HP1α and citrullination are regulators of immune genes and HERVs, and that reactivation of cytokine genes and HERVs in multiple sclerosis patients arises from perturbed chromatin-mediated repression. Notably, citrullination of histone H1 promote its dissociation from DNA to create an open chromatin state that is essential for stem cell pluripotency during early embryogenesis [55,56]. Moreover, heterochromatin protein 1γ (HP1γ) citrullination within the chromodomain decreases its interaction with chromatin, which is necessary to facilitate the opening of chromatin in pluripotent stem cells [59]. However, the levels of histone H1 and HP1γ citrullination accompanied by compacted chromatin state during differentiation highlight the functional fine-tuning of citrullination to modulate the chromatin organization and gene expression.
PADI4-mediated histones citrullination also plays a critical role in response to DNA damage. Citrullinated histones interact with p53 through distinct domains and also associate with the p21 promoter to regulate gene expression of several p53 target genes [60]. Elevated levels of histone citrullination in hematopoietic cancer cell lines can induce apoptosis by increasing the expression of p53 and p21 [61]. Expression of the p53 target gene OKL38 gene that encodes an oxidative stress response protein, is induced by DNA damage and regulated by H3Cit2, 8, 17 [62]. The tumor suppressor protein, inhibitor of growth 4 (ING4), undergoes citrullination and regulates gene expression. citrullination of ING4 at the nuclear localization sequence prevents p53 binding to ING4, represses p53 acetylation, and consequently inhibits downstream p21 expression [63]. Further, citrullination of histone chaperone protein, nucleophosmin can occur in p53 dependent manner, which promotes its translocation from the nucleoli to the nucleoplasm and controls the p53-driven growth [64]. DNA damage via the p53 pathway also induces H4Cit3 and of Lamin C around fragmented nuclei in apoptotic cells. Since H4Cit3 negatively correlates with p53 protein expression and with tumor size in non-small cell lung cancer tissues, it is proposed to play a crucial role in carcinogenesis [65]. Taken together, these findings suggest citrullination in the p53 signaling pathway and DNA damage contribute to altered gene regulation in cancer.
4. PADI2 Shows Specific Substrate Preferences
PADI2 is the most widely expressed member of the PADI family. Compiling the data for all PADI gene family members from RNA-sequencing experiments across 106 human tissues (Gene expression Atlas, https://www.ebi.ac.uk/gxa/genes/ensg00000117115, Figure 2B), including NIH Roadmap Epigenomics Mapping Consortium, Encyclopedia of DNA Elements (ENCODE), Genotype-Tissue Expression (GTEx) Project, and Illumina body map project [66,67,68,69,70], shows that PADI2 is highly and widely expressed. Phylogenetic analysis of the PADI family showed that PADI2 is the most conserved family member and likely the founder member of the family. Interestingly, PADI1 and PADI3 enzymes, as well as PADI4 and PADI6 enzymes are more closely related to each other than to PADI2 [8].
One unique feature of PADI2 is that arginine catalysis relies on a substrate-assisted mechanism, so sequential ordered calcium binding is required for proper positioning of a catalytic cysteine-645 residue in relation to the substrate pocket [32,71,72]. This mechanism confers selectivity for certain substrates leading to specific citrullination of H3Cit26 [50,51,52]. Interestingly, H3Cit26 preferentially associates with SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A containing DEAD/H box 1 (SMARCAD1), which regulates naive pluripotency by suppressing heterochromatin formation [73]. This study also supports our previous finding that histone citrullination weakens the binding of H3K9me3 (lysine 9 trimethylation of histone H3) to the heterochromatin proteins, which consequently leads to gene activation of key transcription units [58,73]. Combined, this evidence supports the notion that PADI2 exhibits a unique ability to citrullinates specific proteins that regulate gene expression. Moreover, PADI2 specifically citrullinates glial fibrillary acidic protein (GFAP), an astrocyte-specific marker protein found in Alzheimer’s disease (AD) patients [74]. This work demonstrates the clinical relevance of one specific PADI2 substrate in AD patients. However, the mechanistic insights of this GFAP citrullination remain unaddressed.
Specifically, PADI2 is the most prevalent expressed PADI member and is preferentially expressed in oligodendrocytes (OLs) and other glial cells [75] that link PADI2 specific actions to myelination. The most prominent functions of OLs is forming the myelin sheath, which is essential for proper neuronal communication and central nervous system function [76]. Notably, PADI2 levels increased during OLs differentiation, and PADI2 acts as a chromatin modifier to promotes oligodendrocyte precursor cell differentiation [77]. Therefore, PADI2 contributes to efficient myelination and motor and cognitive functions.
PADI2 is highly expressed in peripheral blood mononuclear cells of healthy individuals and was also found to affect the differentiation of T helper (Th) cells, which play a critical role in immune defense and are associated with autoimmune and allergic diseases [78]. Further, PADI2 specifically citrullinates arginine330 of GATA3 and arginine4 of RORtγ, key transcription factors required for Th cell functions. These are PADI2 specific substrates that are not citrullinated by PADI4, which further highlights the unique features of PADI2. Interestingly, GATA3 citrullination weakens its DNA binding, whereas RORtγ citrullination strengthens its DNA binding ability, so PADI2-directed citrullination attenuates the Th2 cells differentiation and potentiates Th17 cells differentiation [9]. Taken together, these results highlight the functional implication of PADI2 in immune regulation and Th cell driven diseases.
5. The Role of PADI2 in Cancer
Citrullination is also associated with multiple cancers (see [41] for a review on citrullination in cancer and [79] for a review on citrullination in gastrointestinal cancers). Given the fact that PADI2 and 4 are the most expressed in humans, therefore most of the studies had focused on understanding their functions and other members of the PADI family have not been studied in the context of their implications in the specific type of cancer progression. More detailed function of PADI4 in cancer had been described previously [41]. Among PADI family members, PADI2 is highly expressed in primary breast tumors and luminal breast cancer cell lines [80,81]. PADI2 regulates transcription in human mammary epithelial cells [82,83], promotes epithelial-to-mesenchymal transition in mammary tumor cells [84], and facilitates ductal invasion in primary mouse mammary organoids [84]. Meta-analysis from large cohorts of breast cancer patients and related healthy individuals showed that among PADI gene family members, only PADI2 is overexpressed in breast cancer and other cancers, as its overexpression correlates with poor prognosis [85]. All these observations clearly highlight the functional association of PADI2 in breast cancer pathogenesis.
In addition to its role in inflammation and in breast cancers, PADI2 is also implicated in other cancer types, including skin cancer. PADI2 overexpressing transgenic mice spontaneously develop neoplastic skin lesions, of which a subset can progress to invasive squamous cell carcinoma (SCC). These lesions display an increased level of invasiveness and epithelial-to-mesenchymal transition, along with a high level of inflammation [86,87]. These studies suggest that PADI2 facilitates skin tumor progression by enhancing inflammation within the tumor microenvironment. In this context, further mechanistic and clinical studies will reveal how using PADI2 based screening assays will improve cancer treatment strategies.
In castration-resistance prostate cancer (CRPC) patients, PADI2 is overexpressed and modulates androgen receptor (AR) signaling despite androgen depletion therapy (88). PADI2 gene expression is required for cell-cycle progression of prostate cancer cells and for proliferation of CRPC prostate cancer cells both in vitro or in vivo. PADI2-directed citrullination stabilizes the AR and mediates its nuclear translocation, which reduces AR degradation and facilitates its binding to target genes [88]. Taken together, PADI2 promotes prostate cancer progression. Further, PADI2 overexpression can occur in multiple cancers, including breast ductal carcinoma, ovarian serous papillary adenocarcinoma, cervical squamous cell carcinoma, liver hepatocellular carcinoma, lung cancer, and also papillary thyroid carcinoma [89]. Finally, PADI2 may mediate an essential function in gastric cancers by promoting cell proliferation and invasion [79,88].
In contrast to these cancers, PADI2 levels can be downregulated in colorectal cancer (CRC) patients. This was first time reported by Cantariño et al. [90] as they found low levels of PADI2 expression in 98 CRC patients compared to adjacent mucosa from 50 healthy controls. Importantly, this downregulated of PADI2 expression in CRC patients correlated with poor prognosis [90]. Likewise, PADI2 is required to inhibit cellular proliferation of colon cancer cells by arresting the G1 phase of the cell cycle [91]. This result likely arose through PADI2-mediated citrullination which plays a critical role in Wnt (wingless)/ β-catenin signaling pathway in CRC patients [92]. In fact, the anti-parasitic nitazoxanide used to treat CRC patients, efficiently inhibits the Wnt signaling pathway by increasing the stability of the PADI2 protein and enhancing the citrullination of β-catenin, which inhibits the Wnt pathway and thereby decrease the cellular proliferation of colon cancer cells.
Notably, PADI2 is also found to be highly expressed in bone marrow mesenchymal stem cells (BMMSCs) [93]. These BMMSCs are key components for both the normal and transformed bone marrow niche. The stable alteration in the bone marrow niche leads to multiple myeloma, an incurable malignancy of clonal plasma cells, which accounts for approximately 10% of hematological malignancies [94]. PADI2 can increase interleukin-6 (IL-6) levels by modulating occupancy of the histone H3Cit26 mark at the regulatory region of IL-6 by malignant plasma cells in response to the chemotherapeutic agent, bortezomib which suggests PADI2 is involved in the acquisition of drug resistance [93,95]. Importantly, monitoring of PADI2 enzymatic activity is proposed in the early stage of multiple myeloma patients to improve the efficacy of their therapeutic strategies [95].
6. PADI2 Is a Novel Regulator of Transcription
The regulated transcription of genes determines cell identity and malignant transformation and citrullination influences gene expression [30,70]. However, how PADI2 influences gene expression in cancer cells is not fully understood. Recently, we have identified a novel PADI2 specific substrate central to the basal transcriptional machinery. We found that PADI2, but not the other PADI family members, citrullinates arginine1810 (Cit1810) in repeat 31 of the carboxyl-terminal domain (CTD) of the largest subunit of RNA polymerase II (RNAP2) [85]. Strikingly, cit1810 is crucial for RNAP2 to overcome the pausing barrier close to the transcription start site, which enable the efficient transcription of highly expressed genes needed for cell cycle progression, metabolism, and cell proliferation [85]. So, we developed a specific antibody that recognizes Cit1810 and discovered this modification occurs in active RNAP2 phosphorylated at serine residues of the conserved CTD repeats [85].
However, in vitro citrullination of R1810 by PADI2 is blocked by phosphorylation of the adjacent residues, suggesting that repeat 31 is not phosphorylated in vivo [85]. In fact, cit1810 enhances the association of active RNAP2 with the positive transcription elongation factor b (P-TEFb) kinase complex, comprises of cyclin dependent kinase 9 (CDK9) and Cyclin T1 (CCNT1). Thus, cit1810 enables paused RNAP2 to overcome pausing and facilitates gene transcription needed for cellular growth [96]. Taken together, we proposed that PADI2-mediated cit1810 of RNP2 enhances the interaction of the CTD with P-TEFb complex and its associated factors including SEC (super elongating complex), which facilitates the productive elongation (Figure 3). The citrullination of R1810 is required for RNAP2 pause release or for high turnover at promoters of highly expressed genes that maintain cellular proliferation [97,98,99], which may help drive tumor progression and metastasis. This observation is consistent with the recent findings that inhibiting of PADI2 activity suppresses the mammary gland tumor invasion in mice [83] and reduces the mammary cancer progression in dogs and cats [100]. Taken together, this work highlights a mechanism by which PADI2 overexpression may contribute to tumorigenesis.
Since no enzymes that erase citrullination have been described, alternative mechanisms may exit to replace the citrullinated RNAP2 before termination of the transcription cycle. Importantly, as observed for other dynamically regulated arginine residues [7,101], R1810 at RNAP2-CTD can also undergo asymmetrical and symmetrical dimethylation by coactivator associated arginine methyltransferase 1 (CARM1) and protein arginine methyltransferase 5 (PRMT5), respectively [102,103]. Notably, the asymmetrical dimethylation (me2a) of R1810 occurs mainly in hypo-phosphorylated RNAP2 or prior to the initiation of transcription. R1810 is involved in regulating the expression of the small nuclear RNAs (snRNAs) and the nucleolar RNA (snoRNA) in human cells [102]. However, PRMT5 catalyzed symmetrical dimethylation (me2s) of R1810 promotes the recruiting of the survival of motor neuron protein and its interaction with senataxin, a helicase that enhances transcriptional termination [103]. We found that PADI2 cannot use me2a R1810 as a substrate and that citrullination precludes PRMT5 from catalyzing me2s R1810. Moreover, depletion of PRMT5 and/or CARM1 does not influence PADI2 mediated citrullination of R1810. Such depletion also does not interfere with its transcriptional effect nor with the interaction of citrullinated RNAP2 and the P-TEFb complex. These results indicate that R1810 methylation and citrullination are independent regulatory pathways [85].
Future studies should elucidate how a relative minor change of a single arginine to citrulline in the middle of CTD alters cell fate. The loss of the positive charge of the arginine residue could affect the structure by directly changing electrostatic interactions with relevant proteins like P-TEFb. However, arginine-mediated interactions of the intriniscally disordered protein domain of RNAP2-CTD with RNAs and its associated proteins may participate in conjunction with serine residues phosphorylation within consensus CTD repeats to induce changes in macromolecular condesates within the cell nucleus [104,105,106]. In that respect, two independent macromolecular condensates have been postulated, one related to transcription initiation or mediator complex condensate and another with transcription elongation and splicing factors [107]. Recently elongating form of RNAP2 was found to be preferentially incoporated into condesates that are formed by the splicing factors [107,108]. In fact, inhibition of PADI2 activity or mutation of R1810 produces large changes in RNA splicing (unpublished observation). Based on these results, we posit that cit1810 at RNAP2 could participate in modulating these key interactions, which can consequently influence transcriptional output and RNA processing through liquid-liquid phase separation (LLPS, Figure 3). Further work will resolve these possibilities.
PADI2 plays also an essential role in mediating the activation of estrogen receptor (ER) target genes by citrullination of histone H3 arginine 26 (H3Cit26) on chromatin. In this case, PADI2 facilitates transcriptional activation by modulating the chromatin architecture around the estrogen receptor elements of estradiol-induced genes [51]. Further, two minutes of estradiol exposure increased H3Cit26 at the ERα binding sites in chromatin, suggesting that PADI2 mediated H3Cit26 facilitates ERα access to DNA [52]. Given that progestin activated progesterone receptor modulates gene expression and genome organization of breast cancer cells by targeting kinases and chromatin remodelers to chromatin [109,110,111], we are investigating whether PADI2 directed citrullination can modulate progestin gene regulation and genome organization in breast cancer cells.
Many elongation factors and kinases participate in controlling the RNAP2 transcription pause release, a mechanism that regulates the expression of many genes involved in cancer progression and metastasis, like cyclin dependent kinase 9 (CDK9), v-myc myelocytomatosis viral oncogene homolog (MYC), Jumonji domain containing 6 (JMJD6), arginine demethylase and lysine hydroxylase and BRD4 [112,113,114,115,116]. We found that the absence of PADI2-mediated cit1810 of RNAP2 reduced cell proliferation of breast cancer cells, by modulating cell cycle progression. PADI2 modifies the transcriptional machinery and promotes the oncogenic gene regulation. PADI2 depletion drastically affects the transcription elongation of key genes, including MYC gene, which eventually inhibits cell proliferation [85]. Given the tight link between transcription elongation and RNA splicing [117], it will intriguing to explore how citrullination regulates RNA processing during tumor progression.
7. Conclusions and Perspectives
Protein arginine citrullination affects multiple cellular processes by adding a new functional group that modulates key interactions with other proteins or with RNA. Despite the relevance of citrullination in multiple pathophysiological conditions, our knowledge of the underlying molecular mechanisms remains quite limited, so we must overcome this barrier to develop novel therapeutic avenues. PADI2 is the most widely expressed isoform of the PADI family and is highly prevalent across multiple tumor tissues, indicating its potential functional relevance in tumorigenesis. In this review, we have summarized PADI2 specific functions on transcription regulation not only by citrullination of arginine 26 on histone H3 but also by targeting the master regulator of transcription, namely RNAP2. Noteworthy, cit1810 of RNAP2 controls global transcription by modulating the recruitment of P-TEFb and associated protein complexes to the transcription machinery. We highlight the utility to elucidate how PADI2-mediated citrullination contributes to cancer metastasis within the specific tumor microenvironment. We propose PADI2 is a novel therapeutic target, not only for autoimmune and inflammatory diseases, but also for several cancer types.
Author Contributions
Conceptualization, P.S. and M.B.; Writing-original draft preparation, P.S.; Writing-Review & Editing, P.S., M.B.; Supervision, P.S., M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Spanish MEC (SAF2016–75006), the Catalan Government (2017–2019 SGR 747_MBeato), and the European Research Council Synergy Grant “4DGenome” (609989). Also, we acknowledge the support of the Spanish Ministry of Economy and Competitiveness, “Centro de Excelencia Severo Ochoa” and the CERCA Programme/Generalitat de Catalunya”.
Acknowledgments
We thank all the members of the chromatin and gene expression lab for useful suggestions, special thanks to François Le Dily for revising the manuscript and his constructive advice. We thank Life Science Editors for editing assistance.
Conflicts of Interest
The authors declare no competing financial interests.
Abbreviations
| AR | Androgen receptor |
| CCNT1 | Cyclin T1 |
| CDK9 | Cyclin-dependent kinase 9 |
| Cit1810 | Citrullination of arginine1810 at RNA polymerase II |
| H3Cit26 | Citrullination of arginine 26 at histone H3 |
| H3Cit2, 8, 17 | Citrullination of arginine 26 at histone H3 |
| H4Cit3 | Citrullination of arginine 3 at histone H4 |
| CRC | Colorectal Cancer |
| CTD | Carboxyl-terminal domain |
| EMVs | Exosomes and microvesicles |
| ER | Estrogen receptor |
| GTFs | General transcription factors |
| ING4 | Inhibitor of growth 4 |
| IL-6 | Interleukin-6 |
| LLPS | Liquid-liquid phase separation |
| NET | Neutrophil extracellular trap |
| OLs | Oligodendrocytes |
| PADI | Peptidyl arginine deiminase |
| P-TEFb | Positive transcription elongation factor b |
| RNAP2 | RNA polymerase II |
| SEC | Super elongation complex |
References
- Rogers, G.E.; Simmonds, D.H. Content of citrulline and other amino-acids in a protein of hair follicles. Nature 1958, 182, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Van Venrooij, W.J.; Pruijn, G.J. Citrullination: A small change for a protein with great consequences for rheumatoid arthritis. Arthritis Res. 2000, 2, 249–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhrmann, J.; Clancy, K.W.; Thompson, P.R. Chemical biology of protein arginine modifications in epigenetic regulation. Chem. Rev. 2015, 115, 5413–5461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Thompson, P.R. Protein Arginine Deiminases (PADs): Biochemistry and chemical biology of protein citrullination. Acc. Chem. Res. 2019, 52, 818–832. [Google Scholar] [CrossRef]
- Witalison, E.E.; Thompson, P.R.; Hofseth, L.J. Protein Arginine Deiminases and Associated Citrullination: Physiological Functions and Diseases Associated with Dysregulation. Curr. Drug Targets 2015, 16, 700–710. [Google Scholar] [CrossRef]
- Orgován, G.; Noszál, B. The complete microspeciation of arginine and citrulline. J. Pharm. Biomed. Anal. 2011, 54, 965–971. [Google Scholar] [CrossRef]
- Tanikawa, C.; Ueda, K.; Suzuki, A.; Iida, A.; Nakamura, R.; Atsuta, N.; Tohnai, G.; Sobue, G.; Saichi, N.; Momozawa, Y.; et al. Citrullination of RGG motifs in FET proteins by PAD4 regulates protein aggregation and ALS susceptibility. Cell Rep. 2018, 22, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Vossenaar, E.R.; Zendman, A.J.; van Venrooij, W.J.; Pruijn, G.J. PAD, a growing family of citrullinating enzymes: Genes, features and involvement in disease. Bioessays 2003, 25, 1106–1118. [Google Scholar] [CrossRef]
- Sun, B.; Chang, H.H.; Salinger, A.; Tomita, B.; Bawadekar, M.; Holmes, C.L.; Shelef, M.A.; Weerapana, E.; Thompson, P.R.; Ho, I.C. Reciprocal regulation of Th2 and Th17 cells by PAD2-mediated citrullination. JCI Insight 2019, 4, 129687. [Google Scholar] [CrossRef]
- Senshu, T.; Kan, S.; Ogawa, H.; Manabe, M.; Asaga, H. Preferential deimination of keratin K1 and filaggrin during the terminal differentiation of human epidermis. Biochem. Biophys. Res. Commun. 1996, 225, 712–719. [Google Scholar] [CrossRef]
- Méchin, M.C.; Takahara, H.; Simon, M. Deimination and peptidylarginine deiminases in skin physiology and diseases. Int. J. Mol. Sci. 2020, 21, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, N.S.; Yu, H.C.; Tung, C.H.; Huang, K.Y.; Huang, H.B.; Lu, M.C. Increased peptidylarginine deiminases expression during the macrophage differentiation and participated inflammatory responses. Arthritis Res. 2019, 21, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghamdi, M.; Alasmari, D.; Assiri, A.; Mattar, E.; Aljaddawi, A.A.; Alattas, S.G.; Redwan, E.M. An overview of the intrinsic role of citrullination in autoimmune disorders. J. Immunol. Res. 2019, 2019, 7592851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, I.; Singhrao, S.K.; Potempa, J. Citrullination as a plausible link to periodontitis, rheumatoid arthritis, atherosclerosis and Alzheimer’s disease. J. Oral Microbiol. 2018, 10, 1487742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rondas, D.; Crèvecoeur, I.; D’Hertog, W.; Ferreira, G.B.; Staes, A.; Garg, A.D.; Eizirik, D.L.; Agostinis, P.; Gevaert, K.; Overbergh, L.; et al. Citrullinated glucose-regulated protein 78 is an autoantigen in type 1 diabetes. Diabetes 2015, 64, 573–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarus, R.C.; Buonora, J.E.; Flora, M.N.; Freedy, J.G.; Holstein, G.R.; Martinelli, G.P.; Jacobowitz, D.M.; Mueller, G.P. Protein citrullination: A proposed mechanism for pathology in traumatic brain injury. Front. Neurol. 2015, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Jang, B.; Jeon, Y.C.; Shin, H.Y.; Lee, Y.J.; Kim, H.; Kondo, Y.; Ishigami, A.; Kim, Y.S.; Choi, E.K. Myelin basic protein citrullination, a hallmark of central nervous system demyelination, assessed by novel monoclonal antibodies in prion diseases. Mol. Neurobiol. 2018, 55, 3172–3184. [Google Scholar] [CrossRef]
- Samara, K.D.; Trachalaki, A.; Tsitoura, E.; Koutsopoulos, A.V.; Lagoudaki, E.D.; Lasithiotaki, I.; Margaritopoulos, G.; Pantelidis, P.; Bibaki, E.; Siafakas, N.M.; et al. Upregulation of citrullination pathway: From autoimmune to idiopathic lung fibrosis. Respir. Res. 2017, 18, 218. [Google Scholar] [CrossRef] [Green Version]
- Muraro, S.P.; De Souza, G.F.; Gallo, S.W.; Da Silva, B.K.; De Oliveira, S.D.; Vinolo, M.A.R.; Saraiva, E.M.; Porto, B.N. Respiratory Syncytial Virus induces the classical ROS-dependent NETosis through PAD-4 and necroptosis pathways activation. Sci. Rep. 2018, 8, 14166. [Google Scholar] [CrossRef]
- Valesini, G.; Gerardi, M.C.; Iannuccelli, C.; Pacucci, V.A.; Pendolino, M.; Shoenfeld, Y. Citrullination and autoimmunity. Autoimmune. Rev. 2015, 14, 490–497. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, C.L.; Shim, D.; Kernien, J.; Johnson, C.J.; Nett, J.E.; Shelef, M.A. Insights into Neutrophil Extracellular Traps through Systematic Evaluation of Citrullination and Peptidylarginine Deiminases. J. Immunol. Res. 2019, 2019, 2160192. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Palaniyar, N. Transcriptional firing helps to drive NETosis. Sci. Rep. 2017, 7, 41749. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Schwerdtner, L.; Sams, K.; Mondal, S.; Ahmad, F.; Schmidt, R.E.; Coonrod, S.A.; Thompson, P.R.; Lerch, M.M.; Bossaller, L. Cutting Edge: Protein Arginine Deiminase 2 and 4 Regulate NLRP3 Inflammasome-Dependent IL-1β Maturation and ASC Speck Formation in Macrophages. J. Immunol. 2019, 203, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.E.; Taylor, L.D. The enzymic derivation of citrulline residues from arginine residues in situ during the biosynthesis of hair proteins that are cross-linked by isopeptide bonds. Adv. Exp. Med. Biol. 1977, 86A, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, M.; Sugawara, K. Properties of peptidylarginine deiminase from the epidermis of newborn rats. J. Biochem. 1981, 89, 257–263. [Google Scholar] [CrossRef]
- Chavanas, S.; Méchin, M.C.; Takahara, H.; Kawada, A.; Nachat, R.; Serre, G.; Simon, M. Comparative analysis of the mouse and human peptidylarginine deiminase gene clusters reveals highly conserved non-coding segments and a new human gene, PADI6. Gene 2004, 330, 19–27. [Google Scholar] [CrossRef]
- Nakashima, K.; Hagiwara, T.; Yamada, M. Nuclear localization of peptidyl arginine deiminase V and histone deimination in granulocytes. J. Biochem. 2002, 277, 49562–49568. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Nagar, M.; Maurais, A.J.; Slade, D.J.; Parelkar, S.S.; Coonrod, S.A.; Weerapana, E.; Thompson, P.R. Calcium regulates the nuclear localization of protein arginine deiminase 2. Biochemistry 2019, 58, 3042–3056. [Google Scholar] [CrossRef]
- Bicker, K.L.; Thompson, P.R. The protein arginine deiminases: Structure, function, inhibition, and disease. Biopolymers 2013, 99, 155–163. [Google Scholar] [CrossRef]
- Arita, K.; Hashimoto, H.; Shimizu, T.; Nakashima, K.; Yamada, M.; Sato, M. Structural basis for Ca (2+)-induced activation of human PAD4. Nat. Struct. Mol. Biol. 2004, 11, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Slade, D.J.; Fang, P.; Dreyton, C.J.; Zhang, Y.; Fuhrmann, J.; Rempel, D.; Bax, B.D.; Coonrod, S.A.; Lewis, H.D.; Guo, M.; et al. Protein Arginine Deiminase 2 Binds Calcium in an Ordered Fashion: Implications for Inhibitor Design. ACS Chem. Biol. 2015, 10, 1043–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, G.E.; Harding, H.W.; Llewellyn-Smith, I.J. The origin of citrulline-containing proteins in the hair follicle and the chemical nature of trichohyalin, an intracellular precursor. Biochim. Biophys. Acta 1977, 495, 159–175. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y. Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis. Biochim. Biophys. Acta 2013, 182910, 1126–1135. [Google Scholar] [CrossRef] [Green Version]
- Lewis, H.D.; Nacht, M. iPAD or PADi-’tablets’ with therapeutic disease potential? Curr. Opin. Chem. Biol. 2016, 33, 169–178. [Google Scholar] [CrossRef]
- Durrant, L.G.; Metheringham, R.L.; Brentville, V.A. Autophagy, citrullination and cancer. Autophagy 2016, 12, 1055–1056. [Google Scholar] [CrossRef] [Green Version]
- Darrah, E.; Andrade, F. Rheumatoid arthritis and citrullination. Curr. Opin. Rheumatol. 2018, 29, 72–78. [Google Scholar] [CrossRef]
- Tu, R.; Grover, H.M.; Kotra, L.P. Peptidyl Arginine Deiminases and Neurodegenerative Diseases. Curr. Med. Chem. 2016, 23, 104–114. [Google Scholar] [CrossRef]
- McGinty, J.W.; Chow, I.T.; Greenbaum, C.; Odegard, J.; Kwok, W.W.; James, E.A. Recognition of posttranslationally modified GAD65 epitopes in subjects with type 1 diabetes. Diabetes 2014, 63, 3033–3040. [Google Scholar] [CrossRef] [Green Version]
- Mohanan, S.; Cherrington, B.D.; Horibata, S.; McElwee, J.L.; Thompson, P.R.; Coonrod, S.A. Potential role of peptidylarginine deiminase enzymes and protein citrullination in cancer pathogenesis. Bio. Res. Int. 2012, 2012, 895343. [Google Scholar] [CrossRef] [Green Version]
- Yuzhalin, A.E. Citrullination in Cancer. Cancer Res. 2019, 79, 1274–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Dwivedi, N.; Bechtel, T.J.; Paulsen, J.L.; Muth, A.; Bawadekar, M.; Li, G.; Thompson, P.R.; Shelef, M.A.; Schiffer, C.A.; et al. Citrullination of NF-κB p65 promotes its nuclear localization and TLR-induced expression of IL-1β and TNFα. Sci. Immunol. 2017, 2, eaal3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bawadekar, M.; Shim, D.; Johnson, C.J.; Warner, T.F.; Rebernick, R.; Damgaard, D.; Nielsen, C.H.; Pruijn, G.J.M.; Nett, J.E.; Shelef, M.A. Peptidylarginine deiminase 2 is required for tumor necrosis factor alpha-induced citrullination and arthritis, but not neutrophil extracellular trap formation. J. Autoimmun. 2017, 80, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Gallagher, M.; Kholia, S.; Kosgodage, U.S.; Hristova, M.; Hardy, J.; Inal, J.M. Peptidylarginine deiminases-Roles in cancer and neurodegeneration and possible avenues for therapeutic intervention via modulation of exosome and microvesicle (EMV) release? Int. J. Mol. Sci. 2017, 18, 1196. [Google Scholar] [CrossRef] [PubMed]
- Sase, T.; Arito, M.; Onodera, H.; Omoteyama, K.; Kurokawa, M.S.; Kagami, Y.; Ishigami, A.; Tanaka, Y.; Kato, T. Hypoxia-induced production of peptidylarginine deiminases and citrullinated proteins in malignant glioma cells. Biochem. Biophys. Res. Commun. 2017, 482, 50–56. [Google Scholar] [CrossRef]
- Yu, R.; Li, C.; Sun, L.; Jian, L.; Ma, Z.; Zhao, J.; Liu, X. Hypoxia induces production of citrullinated proteins in human fibroblast-like synoviocytes through regulating HIF1α. Scand. J. Immunol. 2018, 87, e12654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wysocka, J.; Sayegh, J.; Lee, Y.H.; Perlin, J.R.; Leonelli, L.; Sonbuchner, L.S.; McDonald, C.H.; Cook, R.G.; Dou, Y.; et al. Human PAD4 Regulates Histone Arginine Methylation Levels via Demethylimination. Science 2004, 306, 279–283. [Google Scholar] [CrossRef]
- Hagiwara, T.; Nakashima, K.; Hirano, H.; Senshu, T.; Yamada, M. Deimination of arginine residues in nucleophosmin/B23 and histones in HL-60 granulocytes. Biochem. Biophys. Res. Commun. 2002, 290, 979–983. [Google Scholar] [CrossRef]
- Kan, R.; Jin, M.; Subramanian, V.; Causey, C.P.; Thompson, P.R.; Coonrod, S.A. Potential role for PADI-mediated histone citrullination in preimplantation development. BMC Dev. Biol. 2012, 12, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, X.; Zhang, M.; Li, T.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; Zhang, X. Peptidylarginine deiminase 1 catalyzed histone citrullination is essential for early embryo development. Sci. Rep. 2016, 6, 38727. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Bolt, M.; Guertin, M.J.; Chen, W.; Zhang, S.; Cherrington, B.D.; Slade, D.J.; Dreyton, C.J.; Subramanian, V.; Bicker, K.L.; et al. Peptidylarginine deiminase 2-catalyzed histone H3 arginine 26 citrullination facilitates estrogen receptor α target gene activation. Proc. Natl. Acad. Sci. USA 2012, 109, 13331–13336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guertin, M.J.; Zhang, X.; Anguish, L.; Kim, S.; Varticovski, L.; Lis, J.T.; Hager, G.L.; Coonrod, S.A. Targeted H3R26 deimination specifically facilitates estrogen receptor binding by modifying nucleosome structure. PLoS Genet. 2014, 10, e1004613. [Google Scholar] [CrossRef] [PubMed]
- Cherrington, B.D.; Morency, E.; Struble, A.M.; Coonrod, S.A.; Wakshlag, J.J. Potential role for peptidylarginine deiminase 2 (PAD2) in citrullination of canine mammary epithelial in citrullination of caninine mammary epithelial cell histones. PLoS ONE 2010, 5, e11768. [Google Scholar] [CrossRef] [Green Version]
- Damgaard, D.; Bawadekar, M.; Senolt, L.; Stensballe, A.; Shelef, M.A.; Nielsen, C.H. Relative efficiencies of peptidylarginine deiminase 2 and 4 in generating target sites for anti-citrullinated protein antibodies in fibrinogen, alpha-enolase and histone H3. PLoS ONE 2018, 13, e0203214. [Google Scholar] [CrossRef] [PubMed]
- Christophorou, M.A.; Castelo-Branco, G.; Halley-Stott, R.P.; Oliveira, C.S.; Loos, R.; Radzisheuskaya, A.; Mowen, K.A.; Bertone, P.; Silva, J.C.; Zernicka-Goetz, M.; et al. Citrullination regulates pluripotency and histone H1 binding to chromatin. Nature 2014, 507, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slade, D.J.; Subramanian, V.; Thompson, P.R. Pluripotency: Citrullination unravels stem cells. Nat. Chem. Biol. 2014, 10, 327–328. [Google Scholar] [CrossRef] [Green Version]
- De Vore, S.B.; Young, C.H.; Li, G.; Sundararajan, A.; Ramaraj, T.; Mudge, J.; Schilkey, F.; Muth, A.; Thompson, P.R.; Cherrington, B.D. Histone Citrullination Represses MicroRNA Expression, Resulting in Increased Oncogene mRNAs in Somatolactotrope Cells. Mol. Cell Biol. 2018, 38, e00084-18. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Azebi, S.; England, P.; Christensen, T.; Møller-Larsen, A.; Petersen, T.; Batsché, E.; Muchardt, C. Citrullination of histone H3 interferes with HP1-mediated transcriptional repression. PLoS Genet. 2012, 9, e1002934. [Google Scholar] [CrossRef] [Green Version]
- Wiese, M.; Bannister, A.J.; Basu, S.; Boucher, W.; Wohlfahrt, K.; Christophorou, M.A.; Nielsen, M.L.; Klenerman, D.; Laune, E.D.; Kouzarides, T. Citrullination of HP1γ chromodomain affects association association with chromatin. Epigenet. Chromatin 2019, 12, 21. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Wang, D.; Yao, H.; Doret, P.; Hao, G.; Shen, Q.; Qiu, H.; Zhang, X.; Wang, Y.; Chen, G.; et al. Coordination of PAD4 and HDAC2 in the regulation of p53-target gene expression. Oncogene 2010, 29, 3153–3162. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.; Liao, Y.F.; Chang, W.H.; Liu, C.C.; Hsieh, M.C.; Hsu, P.C.; Tsay, G.J.; Hung, H.C. Overexpression of peptidylarginine deiminase IV features in apoptosis of haematopoietic cells. Apoptosis 2006, 11, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Li, P.; Venters, B.J.; Zheng, S.; Thompson, P.R.; Pugh, B.F.; Wang, Y. Histone Arg modifications and p53 regulate the expression of OKL38, a mediator of apoptosis. J. Biol. Chem. 2008, 283, 20060–20068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Fast, W. Citrullination of inhibitor of growth 4 (ING4) by peptidylarginine deminase 4 (PAD4) disrupts the interaction between ING4 and p53. J. Biol. Chem. 2011, 286, 17069–17078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanikawa, C.; Ueda, K.; Nakagawa, H.; Yoshida, N.; Nakamura, Y.; Matsuda, K. Regulation of protein Citrullination through p53/PADI4 network in DNA damage response. Cancer Res. 2009, 69, 8761–8769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanikawa, C.; Espinosa, M.; Suzuki, A.; Masuda, K.; Yamamoto, K.; Tsuchiya, E.; Ueda, K.; Daigo, Y.; Nakamura, Y.; Matsuda, K. Regulation of histone modification and chromatin structure by the p53-PADI4 pathway. Nat. Commun. 2012, 3, 676. [Google Scholar] [CrossRef] [PubMed]
- Roadmap, E.C.; Anshul, K.; Wouter, M.; Jason, E.; Misha, B.; Angela, Y.; Alireza, H.-M.; Pouya, K.; Zhang, Z.Z.; Wang, J.R.; et al. Integrative analysis of 111 reference human epigenomes. Nature 2015, 518, 317–330. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Lin, Y.; Nery, J.R.; Urich, M.A.; Breschi, A.; Davis, C.A.; Dobin, A.; Zaleski, C.; Beer, M.A.; Chapman, W.C.; et al. Comparison of the transcriptional landscapes between human and mouse tissus. Proc. Natl. Acad. Sci. USA 2014, 111, 17224–17229. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Qing, T.; Zhu, J.; Wen, Z.; Yu, Y.; Fukumura, R.; Zheng, Y.; Gondo, Y.; Shi, L. A Comprehensive Mouse Transcriptomic BodyMap across 17 Tissues by RNA-seq. Sci. Rep. 2017, 7, 4200. [Google Scholar] [CrossRef] [Green Version]
- Ardlie, K.G.; Deluca, D.S.; Segrè, A.V.; Sullivan, T.J.; Young, T.R.; Gelfand, E.T.; Trowbridge, C.A.; Maller, J.B.; Tukiainen, T.; Lek, M.; et al. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef] [Green Version]
- Asmann, Y.W.; Necela, B.M.; Kalari, K.R.; Hossain, A.; Baker, T.R.; Carr, J.M.; Davis, C.; Getz, J.E.; Hostetter, G.; Li, X.; et al. Detection of redundant fusion transcripts as biomarker or disease-specific therapeutic targets in breast cancer. Cancer Res. 2012, 72, 1921–1928. [Google Scholar] [CrossRef] [Green Version]
- Darrah, E.; Rosen, A.; Giles, J.T.; Andrade, F. Peptidyl arginine deiminase 2, 3 and 4 have distinct specificities against cellular substrates: Novel insights into autoantigen selection in rheumatoid arthritis. Ann. Rheum. Dis. 2012, 71, 92–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyton, C.J.; Knuckley, B.; Jones, J.E.; Lewallen, D.M.; Thompson, P.R. Mechanistic studies of protein arginine deiminase 2: Evidence for a substrate assisted mechanism. Biochemistry 2014, 53, 4426–4433. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Lu, J.; Sridhar, B.; Cao, X.; Yu, P.; Zhao, T.; Chen, C.C.; McDee, D.; Sloofman, L.; Wang, Y.; et al. SMARCAD1 Contributes to the Regulation of Naive Pluripotency by Interacting with Histone Citrullination. Cell Rep. 2017, 18, 3117–3128. [Google Scholar] [CrossRef]
- Ishigami, A.; Masutomi, H.; Handa, S.; Nakamura, M.; Nakaya, S.; Uchida, Y.; Saito, Y.; Murayama, S.; Jang, B.; Jeon, Y.C.; et al. Mass spectrometric identification of citrullination sites and immunohistochemical detection of citrullinated glial fibrillary acidic protein in Alzheimer’s disease brains. J. Neurosci. Res. 2015, 93, 1664–1674. [Google Scholar] [CrossRef]
- Zeisel, A.; Muñoz-Manchado, A.B.; Codeluppi, S.; Lönnerberg, P.; La Manno, G.; Juréus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef]
- Bergles, D.E.; Richardson, W.D. Oligodendrocyte Development and Plasticity. Cold Spring Harb. Perspect. Biol. 2015, 8, a020453. [Google Scholar] [CrossRef]
- Falcão, A.M.; Meijer, M.; Scaglione, A.; Rinwa, P.; Agirre, E.; Liang, J.; Larsen, S.C.; Heskol, A.; Frawley, R.; Klingener, M.; et al. PAD2-mediated citrullination contributes to efficient oligodendrocyte differentiation and myelination. Cell Rep. 2019, 27, 1090–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.H.; Liu, G.Y.; Dwivedi, N.; Sun, B.; Okamoto, Y.; Kinslow, J.D.; Deane, K.D.; Demoruelle, M.K.; Norris, J.M. A molecular signature of preclinical rheumatoid arthritis triggered by dysregulated PTPN22. JCI Insights 2016, 1, e90045. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Yu, Y. Progression on Citrullination of Proteins in Gastrointestinal Cancers. Front. Oncol. 2019, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Blick, T.; Hugo, H.; Widodo, E.; Waltham, M.; Pinto, C.; Mani, S.A.; Weinberg, R.A.; Neve, R.M.; Lenburg, M.E.; Thompson, E.W. Epithelial Mesenchymal Transition Traits in Human Breast Cancer Cell Lines Parallel the CD44hi/CD24lo/- Stem Cell Phenotype in Human Breast Cancer. J. Mammary Gland Biol. Neoplasia 2010, 15, 235–252. [Google Scholar] [CrossRef]
- Mackay, A.; Tamber, N.; Fenwick, K.; Iravani, M.; Grigoriadis, A.; Dexter, T.; Lord, C.J.; Reis-Filho, J.S.; Ashworth, A. High-resolution integrated analysis of genetic and expression profiles of breast cancer cell lines. Breast Cancer Res. Treat. 2009, 118, 481–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherrington, B.D.; Zhang, X.; McElwee, J.L.; Morency, E.; Anguish, L.J.; Coonrod, S.A. Potential role of PAD2 in gene regulation in breast cancer cells. PLoS ONE 2012, 7, e41242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horibata, S.; Rogers, K.E.; Sadegh, D.; Anguish, L.J.; McElwee, J.L.; Shah, P.; Thompson, P.R.; Coonrod, S.A. Role of peptidylarginine deiminase 2 (PAD2) in mammary carcinoma cell migration. BMC Cancer 2017, 17, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanan, S.; Griffith, O.L.; Breuer, H.C.; Anguish, L.J.; Cherrington, B.D.; Palmer, A.M.; Howe, L.R.; Subramanian, V.; Causey, C.P. Identification of PADI2 as a potential breast cancer biomarker and therapeutic target. BMC Cancer 2012, 12, 500. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Lioutas, A.; Fernandez-Fuentes, N.; Quilez, J.; Carbonell-Caballero, J.; Wright, R.H.G.; Di Vona, C.; Le Dily, F.; Schüller, R.; Eick, D.; et al. Arginine Citrullination at the C-Terminal Domain Controls RNA Polymerase II Transcription. Mol. Cell 2019, 73, 84–96. [Google Scholar] [CrossRef] [Green Version]
- McElwee, J.L.; Mohanan, S.; Horibata, S.; Sams, K.L.; Anguish, L.J.; McLean, D.; Cvitaš, I.; Wakshlag, J.J.; Coonrod, S.A. PAD2 overexpression in transgenic mice promotes spontaneous skin neoplasia. Cancer Res. 2014, 74, 6306–6363. [Google Scholar] [CrossRef] [Green Version]
- Mohanan, S.; Horibata, S.; Anguish, L.J.; Mukai, C.; Sams, K.; McElwee, J.L.; McLean, D.; Yan, A.; Coonrod, S.A. PAD2 overexpression in transgenic mice augments malignancy and tumor-associated inflammation in chemically initiated skin tumors. Cell Tissue Res. 2017, 370, 275–283. [Google Scholar] [CrossRef]
- Wang, L.; Song, G.; Zhang, X.; Feng, T.; Pan, J.; Chen, W.; Yang, M.; Bai, X.; Pang, Y.; Yu, J.; et al. PADI2-Mediated Citrullination Promotes Prostate Cancer Progression. Cancer Res. 2017, 77, 5755–5768. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zheng, Y.; Xu, B.; Ma, F.; Li, C.; Zhang, X.; Wang, Y.; Chang, X. Investigating the expression, effect and tumorigenic pathway of PADI2 in tumors. OncoTargets Ther. 2017, 10, 1475–1485. [Google Scholar] [CrossRef] [Green Version]
- Cantariño, N.; Musulén, E.; Valero, V.; Peinado, M.A.; Perucho, M.; Moreno, V.; Forcales, S.V.; Douet, J.; Buschbeck, M. Downregulation of the Deiminase PADI2 Is an Early Event in Colorectal Carcinogenesis and Indicates Poor Prognosis. Mol. Cancer Res. 2016, 9, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Funayama, R.; Taniguchi, H.; Mizuma, M.; Fujishima, F.; Kobayashi, M.; Ohnuma, S.; Unno, M.; Nakayama, K. Protein-arginine deiminase 2 suppresses proliferation of colon cancer cells through protein citrullination. Cancer Sci. 2017, 108, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Olsen, J.R.; Yuan, X.; Cheng, P.F.; Levesque, M.P.; Brokstad, K.A.; Hoffman, P.S.; Oyan, A.M.; Zhang, W.; Kalland, K.H.; et al. Small molecule promotes β-catenin citrullination and inhibits Wnt signaling in cancer. Nat. Chem. Biol. 2018, 14, 94–101. [Google Scholar] [CrossRef] [PubMed]
- McNee, G.; Eales, K.L.; Wei, W.; Williams, D.S.; Barkhuizen, A.; Bartlett, D.B.; Essex, S.; Anandram, S.; Filer, A.; Moss, P.A.; et al. Citrullination of histone H3 drives IL-6 production by bone marrow mesenchymal stem cells in MGUS and multiple myeloma. Leukemia 2017, 31, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Mogollón, P.; Díaz-Tejedor, A.; Algarín, E.M.; Paíno, T.; Garayoa, M.; Ocio, E.M. Biological Background of Resistance to Current Standards of Care in Multiple Myeloma. Cells 2019, 8, 1432. [Google Scholar] [CrossRef] [Green Version]
- Tanday, S. Targeting PADI2 could stop the progression of myeloma. Lancet Oncol. 2016, 8, e325. [Google Scholar] [CrossRef]
- Corden, J.L. An Arginine Nexus in the RNA Polymerase II CTD. Mol. Cell 2019, 73, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Krebs, A.R.; Imanci, D.; Hoerner, L.; Gaidatzis, D.; Burger, L.; Schübeler, D. Genome-wide single molecule footprinting reveals high RNA polymerase II turnover at paused promoters. Mol. Cell 2017, 67, 411–422. [Google Scholar] [CrossRef] [Green Version]
- Day, D.S.; Zhang, B.; Stevens, S.M.; Ferrari, F.; Larschan, E.N.; Park, P.J.; Pu, W.T. Comprehensive analysis of promoter proximal RNA polymerase II pausing across mammalian cell types. Genome Biol. 2016, 17, 120. [Google Scholar] [CrossRef] [Green Version]
- Zeitlinger, J.; Stark, A.; Kellis, M.; Hong, J.W.; Nechaev, S.; Adelman, K.; Levine, M.; Young, R.A. RNA polymerase stalling at developmental control genes in the Drosophila melanogaster embryo. Nat. Genet. 2007, 39, 1512–1516. [Google Scholar] [CrossRef]
- Ledet, M.M.; Anderson, R.; Harman, R.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; Van de Walle, G.R. BB-Cl-Amidine as a novel therapeutic for canine and feline mammary cancer via activation of the endoplasmic reticulum stress pathway. BMC Cancer 2018, 18, 412. [Google Scholar] [CrossRef]
- Cuthbert, G.L.; Daujat, S.; Snowden, A.W.; Erdjument Bromage, H.; Hagiwara, T.; Yamada, M.; Schneider, R.; Gregory, P.D.; Tempst, P.; Bannister, A.J.; et al. Histone deimination antagonizes arginine methylation. Cell 2004, 118, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, R.J.; Rojas, L.A.; Beck, D.B.; Bonasio, R.; Schüller, R.; Drury, W.J.; Eick, D.; Reinberg, D. The C-terminal domain of RNA polymerase II is modified by site-specific methylation. Science 2011, 332, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.Y.; Gish, G.; Braunschweig, U.; Li, Y.; Ni, Z.; Schmitges, F.W.; Zhong, G.; Liu, K.; Li, W.; Moffat, J.; et al. SMN and symmetric arginine dimethylation of RNA polymerase II C-terminal domain control termination. Nature 2016, 529, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Burke, K.A.; Janke, A.M.; Rhine, C.L.; Fawzi, N.L. Residue-by-residue view of in vitro FUS granules that bind the C-terminal domain of RNA Polymerase II. Mol. Cell 2015, 60, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hnisz, D.; Shrinivas, K.; Young, R.A.; Chakraborty, A.K.; Sharp, P.A. A phase separation model for transcriptional control. Cell 2017, 169, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harlen, K.M.; Churchman, L.S. The code and beyond: Transcription regulation by the RNA polymerase II carboxy-terminal domain. Nat. Rev. Mol. Cell Biol. 2017, 18, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.E.; Manteiga, J.C.; Henninger, J.E.; Sabari, B.R.; Dall’Agnese, A.; Hannett, N.M.; Spille, J.H.; Afeyan, L.K.; Zamudio, A.V.; Shrinivas, K.; et al. Pol II phosphorylation regulates a switch between transcriptional and splicing condensates. Nature 2019, 572, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Portz, B.; Shorter, J. Switching Condensates: The CTD Code Goes Liquid. Trends Biochem. Sci. 2020, 45, 1–3. [Google Scholar] [CrossRef]
- Le Dily, F.; Baù, D.; Pohl, A.; Vicent, G.P.; Serra, F.; Soronellas, D.; Castellano, G.; Wright, R.H.; Ballare, C.; Filion, G.; et al. Distinct structural transitions of chromatin topological domains correlate with coordinated hormone-induced gene regulation. Genes Dev. 2014, 28, 2151–2162. [Google Scholar] [CrossRef]
- Le Dily, F.; Vidal, E.; Cuartero, Y.; Quilez, J.; Nacht, A.S.; Vicent, G.P.; Carbonell-Caballero, J.; Sharma, P.; Villanueva-Cañas, J.L.; Ferrari, R.; et al. Hormone-control regions mediate steroid receptor-dependent genome organization. Genome Res. 2019, 29, 29–39. [Google Scholar] [CrossRef]
- Wright, R.H.; Lioutas, A.; Le Dily, F.; Soronellas, D.; Pohl, A.; Bonet, J.; Nacht, A.S.; Samino, S.; Font-Mateu, J.; Vicent, G.P.; et al. ADP-ribose-derived nuclear ATP synthesis by NUDIX5 is required for chromatin remodeling. Science 2016, 352, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, L.; Leng, Y.; Dai, Y.; Orlowski, R.Z.; Grant, S. Positive transcription elongation factor b (P-TEFb) is a therapeutic target I human multiple myeloma. Oncotarget 2017, 8, 59476–59491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bywater, M.J.; Pearson, R.B.; McArthur, G.A.; Hannan, R.D. Dysregulation of the basal RNA polymerase transcription apparatus in cancer. Nat. Rev. Cancer 2013, 13, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.E.; Liau, B.B.; Wallac, L.C.; Morton, A.R.; Xie, Q.; Dixit, D.; Factor, D.C.; Kim, L.J.Y.; Morrow, J.J.; Wu, Q.; et al. Transcription elongation factors represent in vivo cancer dependencies in glioblastoma. Nature 2017, 547. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Yik, J.H.; Lew, Q.J.; Chao, S.H. Brd4 and HEXIM1: Multiple roles in P-TEFb regulation and cancer. BioMed. Res. Int. 2014, 2014, 232870. [Google Scholar] [CrossRef]
- Zanconato, F.; Battilana, G.; Forcato, M.; Filippi, L.; Azzolin, L.; Manfrin, A.; Quaranta, E.; Di Biagio, D.; Sigismondo, G.; Guzzardo, V.; et al. Transcriptional addiction in cancer cells is mediated by YAP/TAZ through BRD4. Nat. Med. 2018, 10, 1599–1610. [Google Scholar] [CrossRef]
- Naftelberg, S.; Schor, I.E.; Ast, G.; Kornblihtt, A.R. Regulation of alternative splicing through coupling with transcription and chromatin structure. Annu. Rev. Biochem. 2015, 84, 165–198. [Google Scholar] [CrossRef]
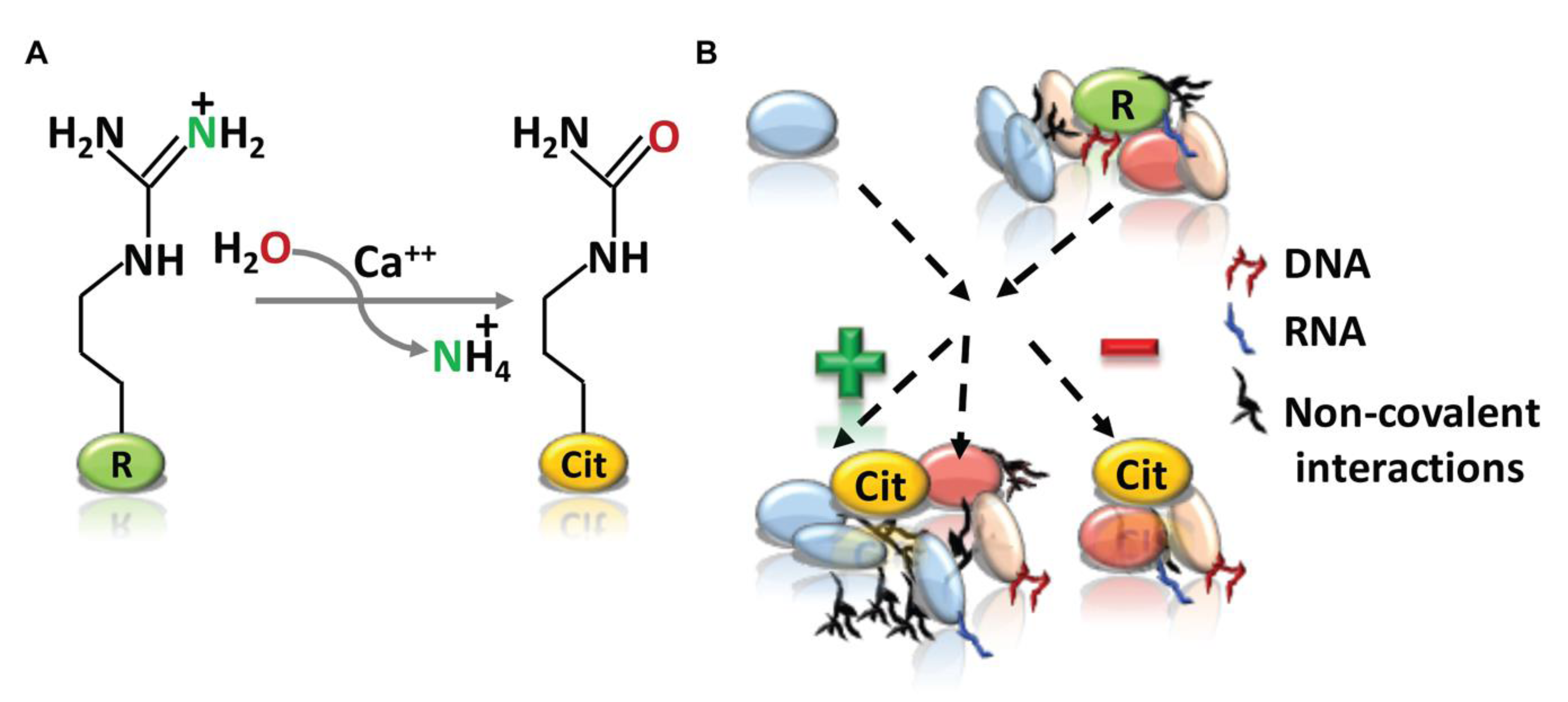
Figure 1.
Arginine citrullination and its cellular functions. (A) The peptidyl arginine deiminases (PADI) enzyme catalyze peptidyl-arginine (positively charged) to peptidyl-citrulline (neutral in charge) and release of ammonia. (B) Arginine (green) to citrulline (orange) conversion could consequently affects the functional protein-protein and protein-nucleic acid interactions by affecting the protein folding, as well as intermolecular and intramolecular interactions. “+” and “−” signs representing the positive and negative interactions respectively.
Figure 1.
Arginine citrullination and its cellular functions. (A) The peptidyl arginine deiminases (PADI) enzyme catalyze peptidyl-arginine (positively charged) to peptidyl-citrulline (neutral in charge) and release of ammonia. (B) Arginine (green) to citrulline (orange) conversion could consequently affects the functional protein-protein and protein-nucleic acid interactions by affecting the protein folding, as well as intermolecular and intramolecular interactions. “+” and “−” signs representing the positive and negative interactions respectively.

Figure 2.
Human PADI gene family. (A). Five human PADIs genes (PADI1, PADI2, PADI3, PADI4 and PADI6) are present in the p36.13 of chromosome 1. Among the family members, PADI2 gene is the only one in reverse direction. PADI loci encompasses the same topological associated domain (TAD) in T47D breast cancer cells. (B). PADI2 is widely expressed among family members. Heatmap representing the basal expression level of all five human PADI members across 106 human tissues compiled from the gene expression atlas (https://www.ebi.ac.uk/gxa/home). Basal gene expression represented as TPM, transcripts per million from RNA-sequencing.
Figure 2.
Human PADI gene family. (A). Five human PADIs genes (PADI1, PADI2, PADI3, PADI4 and PADI6) are present in the p36.13 of chromosome 1. Among the family members, PADI2 gene is the only one in reverse direction. PADI loci encompasses the same topological associated domain (TAD) in T47D breast cancer cells. (B). PADI2 is widely expressed among family members. Heatmap representing the basal expression level of all five human PADI members across 106 human tissues compiled from the gene expression atlas (https://www.ebi.ac.uk/gxa/home). Basal gene expression represented as TPM, transcripts per million from RNA-sequencing.

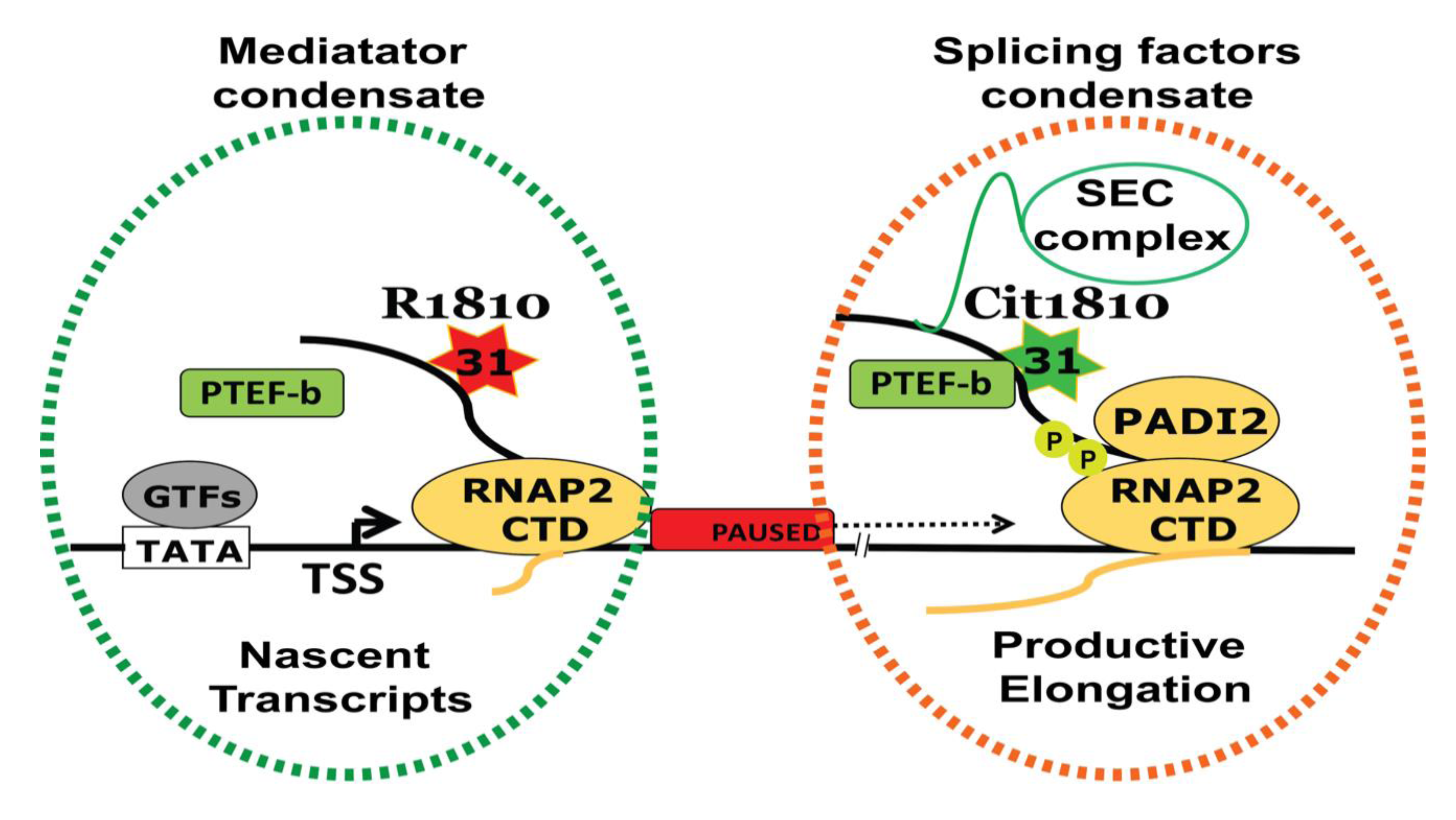
Figure 3.
PADI2-mediated cit1810 RNAP2 is a new player in transcription regulation. PADI2 catalyzed R1810 to Cit1810 at RNAP2-CTD facilitates the association with the P-TEFb complex (comprises CDK9-CCNT1). This interaction overcomes RNAP2 pausing and increases in transcription and cell proliferation. An increased association between the P-TEFb complex and RNAP2-CTD most likely attracts associated complexes including Super Elongating Complex (SEC) to facilitate active RNAP2 encompassing splicing factor condensate (orange dotted) derive transcription regulation. However, R1810 along with nascent transcripts forming mediator complexes condensate (green dotted). TSS = transcription start site; GTFs = general transcription factors.
Figure 3.
PADI2-mediated cit1810 RNAP2 is a new player in transcription regulation. PADI2 catalyzed R1810 to Cit1810 at RNAP2-CTD facilitates the association with the P-TEFb complex (comprises CDK9-CCNT1). This interaction overcomes RNAP2 pausing and increases in transcription and cell proliferation. An increased association between the P-TEFb complex and RNAP2-CTD most likely attracts associated complexes including Super Elongating Complex (SEC) to facilitate active RNAP2 encompassing splicing factor condensate (orange dotted) derive transcription regulation. However, R1810 along with nascent transcripts forming mediator complexes condensate (green dotted). TSS = transcription start site; GTFs = general transcription factors.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Beato, M.; Sharma, P. Peptidyl Arginine Deiminase 2 (PADI2)-Mediated Arginine Citrullination Modulates Transcription in Cancer. Int. J. Mol. Sci. 2020, 21, 1351. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041351
AMA Style
Beato M, Sharma P. Peptidyl Arginine Deiminase 2 (PADI2)-Mediated Arginine Citrullination Modulates Transcription in Cancer. International Journal of Molecular Sciences. 2020; 21(4):1351. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041351
Chicago/Turabian StyleBeato, Miguel, and Priyanka Sharma. 2020. "Peptidyl Arginine Deiminase 2 (PADI2)-Mediated Arginine Citrullination Modulates Transcription in Cancer" International Journal of Molecular Sciences 21, no. 4: 1351. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041351
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.