Micronucleus Assay: The State of Art, and Future Directions


1
Centre for Radiobiology and Biological Dosimetry, Institute of Nuclear Chemistry and Technology, Dorodna 16, 03-195 Warszawa, Poland
2
Department of Molecular Biology and Translational Research, Institute of Rural Health, Jaczewskiego 2, 20-090 Lublin, Poland
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(4), 1534; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041534
Submission received: 4 February 2020
/
Revised: 13 February 2020
/
Accepted: 14 February 2020
/
Published: 24 February 2020
(This article belongs to the Special Issue Radiation-Induced Damage to DNA)
Abstract
:During almost 40 years of use, the micronucleus assay (MN) has become one of the most popular methods to assess genotoxicity of different chemical and physical factors, including ionizing radiation-induced DNA damage. In this minireview, we focus on the position of MN among the other genotoxicity tests, its usefulness in different applications and visibility by international organizations, such as International Atomic Energy Agency, Organization for Economic Co-operation and Development and International Organization for Standardization. In addition, the mechanism of micronuclei formation is discussed. Finally, foreseen directions of the MN development are pointed, such as automation, buccal cells MN and chromothripsis phenomenon.
1. Introduction
Many chemical, physical or biological factors can cause cell death. However, there is a class of factors that not necessarily kill cells, but only damage their genetic material. Such factors are referred to as genotoxins. Although many effective ways to repair DNA damage were discovered, sometimes they fail, which might lead to damage fixation, if the damaged cell survives, and subsequently to its transfer to next generations, if the damaged cell divides.
In a modern world, humans are exposed to different genotoxic agents present in the polluted environment. Hence, tests are needed to determine the level of exposure and health risk. Although many tests classified as “in vivo biomonitoring” are available, a micronucleus test (MN) is one of the best and the most popular [1,2]. The assay is also widely used to test genotoxicity in vitro [1,3,4]. However, it should be emphasized that the concept of MN covers many different techniques that might be used in certain, specific situations [5,6]. In this review, we present various applications of the MN, discuss associated technical challenges and describe future directions of its development.
2. Why Do We Need Genotoxicity Testing, Biomonitoring, Cytogenetic Tests and Biological Dosimetry?
Many physical and chemical factors affect stability of our genome and can contribute to the development of civilization diseases, such as cancer, cardiovascular disease, chronic obstructive pulmonary disease or neurodegenerative diseases [4,5,7,8,9]. Genotoxicity testing allows for an assessment of their impact on humans and biota [10,11]. Studies related to genome stability are not an easy task, as they are biased by many confounding factors, often giving inconsistent results or of low statistical strength that enable a direct attribution of the disease to examined factors. Nevertheless, in some cases, such studies could be conclusive, e.g., studies on a role of radon-induced DNA damage in development of lung cancer, impact of PM-10 and PM-2.5 dusts on human health, cytogenetic effects of unhealthy nutrition or effects of risky, unhealthy behaviors and addictions, such as smoking cigarettes or chewing betel quid [12,13,14,15,16].
Nearly 20 types of well-described in vitro or in vivo genotoxicity tests are presently used [10,17]. In vitro assays are used to investigate the potential genotoxic effect of new pharmaceuticals and other medical materials, daily use goods, chemical and physical factors and poisons, etc. [18]. In vivo tests, despite the aforementioned applications, allow for investigating the impact of environmental factors on humans or biota, the impact of the harmful working environment on human health or early genetic changes associated with various diseases development. The most often used in vitro assays include testing of induction of nucleotide mutation, e.g., bacterial reverse mutation assay (Ames test), mammalian cell gene mutation assay or mouse lymphoma assay, and testing of more complex genome changes by cytogenetic methods, such as the sister chromatid exchange assay, analysis of chromosomal aberration frequency, cytokinesis-block micronucleus assay (CBMN) and comet assay (single-cell gel electrophoresis). In addition to the aforementioned applications, in vivo tests allow for investigation of the impact of environmental factors, working environment or changes associated with various diseases. The most important in vivo tests include three cytogenetic methods, namely comet assay, chromosomal aberration assay and different kinds of MN, including CBMN, mammalian erythrocyte MN (EMN) or buccal cells MN (BMN) [17].
Cytogenetic methods are commonly used in vivo for genotoxicity testing (on animals) and in vitro for genotoxicity testing (on cell lines) of compounds of drugs or food chemicals [2,4,19]. Cytogenetic methods are important to search for DNA damage resulting from environmental and occupational exposure to chemical pollutants and physical factors, as ionizing radiation (IR) [2,3]. Although DNA damage induced by IR is processed by different DNA damage repair systems [20], its improper repair may lead to cell death or formation of mutagenic lesions of different complexity. Cytogenetic methods deal with complex type mutations, known as structural mutations (aberrations), such as dicentrics, translocations, acentric fragments, rings and chromatid type aberrations, and a special type of chromosomal damage, known as micronucleus (Mn). Knowing the frequency of changes in genetic material, it is possible to estimate the extent of exposure or even to reconstruct an absorbed dose of ionizing radiation [21,22]. The level of cytogenetic damage to the cells is proportional to the radiation dose and can be read. This procedure is called biological dosimetry and it is usually used to reconstruct the dose in the event of uncontrolled exposure during accidents in medicine or industry [21,22,23]. In this regard, the most commonly used technique is the dicentric chromosome assay due to its high specificity for radiation, but CBMN is also often used, less specific, but easier to perform, faster and easy to automatize [21,23]. Methods used in biological dosimetry are used to indicate exposure to different chemical and physical factors, different than IR. Though biological dosimetry is the main field of CBMN application, the method is also used in theater areas of medicine, e.g., to detect and study genetic and civilization diseases. Several examples of its application in biomedical sciences are given in Table 1.
3. How Micronuclei are Formed?
Mn is a small, chromatin containing round-shaped body visible in the cytoplasm of cells [6,30]. Mn is considered to be caused by DNA damage or genomic instability [31]. Mn can occur as a result of natural processes, such as metabolism or aging or can be induced by many environmental factors, hazardous habits and different diseases. The vast majority of factors giving origin to Mn are well recognized and described in literature [6,30,31,32]. The most often mentioned are listed in Table 2.
4. Different Types of Micronucleus Assays
Although all types of MN are based on the analysis of micronuclei frequency, they vary in terms of used cells and protocols. The summary is given in Table 3, followed by a more detailed description.
4.1. Cytokinesis-Block Micronucleus Assay (CBMN)
The most popular version of MN is the cytokinesis-block micronucleus assay (CBMN) [6,30] (Figure 1). Because Mn is visible only after cell division, the cytochalasin B that inhibits actin filaments polymerization and formation of contractile microfilaments is used to stop cytokinesis [42,43]. However, cytochalasin B does not stop karyokinesis, thus binucleated cells are formed with Mn present in their cytoplasm. The influence of cytochalasin B on cell proliferation and induction of Mn was discussed in the past [6,44]. The conclusion reached indicates that in most cases, the usage of cytochalasin B does not induce additional Mn, hence, the use of cytochalasin B is recommended [5,6,30,45,46,47,48]. This is of special importance when human lymphocytes are used, as their cell cycle may vary among individuals [4]. According to the mathematical model described by Fenech, MN with cytochalasin B applied to block cytokinesis is superior over MN without cytochalasin B because there are less false-negative results when MN using cytokinesis block is used [49].
The CBMN is prevalently performed on human peripheral blood lymphocytes to study in vivo formation of Mn for biomonitoring or biological dosimetry, however, can be performed on different lymphocytes of other species, e.g., rodents, fish, dogs, rabbits, monkey and apes or other cells of different origin [50,51,52]. The CBMN is also very often used on blood samples in vitro to study genotoxic effects of chemicals. [6,53,54,55,56,57]. The information about in vitro genotoxicity testing by MN is gathered, revised and systematized in the Organization for Economic Co-operation and Development (OECD) 487 Guideline [4].
The CBMN provides a comprehensive basis for in vitro investigating of the chromosome damaging potential of chemicals, noteworthy, both aneugenic (changes in the chromosome number in the cell caused by e.g., tobacco smoking, pesticides) and clastogenic changes (structural aberration caused e.g., by ionizing radiation; acridine yellow, benzene, ethylene oxide, arsene, phosphine) can be detected. According to the OECD guideline, cells should be are treated with chemical compounds in three different ways: cultured with cytochalasin B, cultured without cytochalasin B and cultured in the presence of exogenous metabolic activation system, usually prepared from the liver of rodents (S9 fraction). It is impossible to enumerate all applications of the CBMN. The most important applications of CBMN have already been described, but several more are mentioned in Table 4.
CBMN disorders characterization-occupational exposure, although in the basic version of the assay only Mn are scored, the assay can be extended by scoring other biomarkers, as nucleoplasmic bridges, nuclear buds, nuclear blebs, necrotic and/or apoptotic cells [32]. This type of the assay, called CBMN cytome assay, gives additional information about DNA damage and its repair, cytostasis and cytotoxicity [72].
4.2. Erythrocyte Micronucleus Assay–the Most Popular In Vivo MN
Erythrocyte micronucleus assay (EMn) was initially performed on immature erythrocytes from bone marrow of young mice and rats [73]. The disadvantage of the assay is that bone marrow examination entails sacrificing rodent life. In addition, potentially confounding factors, such as other nucleated cells (must cells, granulocytes or different types of lymphocytes), are present in the bone marrow [74]. EMn was also performed on cellular material taken from human bone marrow to determine cytogenetic damage after radio- and chemotherapy [75,76,77,78].
Due to the high invasiveness of the method, an alternative approach was developed, based on assessing the frequency of Mn in immature erythrocytes in peripheral blood [1,79]. During maturation, erythrocyte precursor cells lose their nuclei, however, retain Mn formed during the nucleated stage [2]. Immature erythrocytes (also called reticulocytes or polychromatic erythrocytes) can be easily recognized from the mature erythrocytes because they still contain RNA in their cytoplasm [1]. In bone marrow, immature erythrocytes constitute about 50% of all erythrocytes [80,81].
Human erythrocyte precursor cells present in the long bone marrow of control specimens contain nuclei, likely as a result of environmental or occupational exposure or genetic factors (Figure 2). Immature erythrocytes arising from the precursor cells also contain Mn, however, they represent only a few percent of all erythrocytes in peripheral blood [81]. Though frequency of Mn in immature erythrocytes is significantly higher than in mature erythrocytes [1], splenic selection, a process that effectively removes micronucleated erythrocytes from peripheral blood, significantly reduces the frequency of Mn. Splenic selection occurs in rats and humans, also in mice, but to a lesser extent [74,82]. Therefore, in humans, the MN in immature erythrocytes is carried out only on individuals with the spleen removed [13,83]. To increase the assay reliability, the MN in immature erythrocytes in peripheral blood has been automated and is carried out by flow cytometry. With this technique, hundreds of thousands of cells can be analyzed in a reliable time, which allowed overcoming problems with low number of cells available for analysis and Mn splenic selection. Flow cytometry aided scoring of Mn in erythrocytes was validated both in rodents and humans [82,84,85].
4.3. Buccal MN (BMm)–Mature but Underused Assay
Although BMm has been used for about 40 years, it seems that only in recent years it gains more interest. The first publications describing this test appeared in the 1980s [86,87]. However, the first publication of the operational protocol in Nature Protocols falls within the last 10 years, after harmonization of the assay by the international HUMNxl group [88,89]. Mn arise in dividing basal cells of oral epithelium but are observed in differentiated cells in the keratinized layer at the buccal surface [90,91]. In addition to Mn, several other cytogenetic biomarkers, including those related to cell death, can be analyzed, which gives more information of the origin of DNA damage, cytostasis and cytotoxicity, somehow analogous to the CBMN cytome assay mentioned earlier [88,91]. This approach was also called the buccal micronucleus cytome (BMCyt) assay [88,91,92].
Mn in the buccal cells are formed in the organism, in rapidly dividing buccal epithelial tissue (Figure 3). Although cells from the oral cavity are exposed to genotoxic or cytotoxic factors by inhalation and food intake to a greater extent than peripheral blood lymphocytes, the background frequency of Mn in buccal cells is very low [93,94]. On the other side, patients undergoing head and neck radiotherapy may serve as a positive control, although some ethical issues must be considered, as in their case the collection of material is more problematic due to lesions and inflammation of the oral mucosa [88,95]. The BMm was used to investigate the impact of nutrition, lifestyle factors (as smoking, drinking alcohol or betel chewing), genotoxin and cytotoxin exposure. Interestingly, correlation was found between the frequency of Mn in buccal cells and increased risk of accelerated aging, certain types of cancer and neurodegenerative diseases [92,96].
BMn seems to be advantageous over other types of MN to study how genotoxic factors affect organisms by inhalation. It is the only method capable to show genotoxic effects of moderate concentrations of radon, as those that are met in unventilated rooms, basements or caves [14,97]. Moreover, the sensitivity of BMn allowed also to show genotoxic effects of work in healthcare, where staff was exposed to very low doses of radiation [98]. BMn is gaining popularity and probably will become a standard cytogenetic test, especially since it is minimally invasive, easy to perform, and cell samples are taken from the oral cavity.
4.4. Other Types of MN
Occasionally, the MN is also performed on cells other than lymphocytes, fibroblasts and buccal cells, such as nasal mucosa cells or urine-derived cells [99,100,101]. Both the test objectives and the method of performance remain similar to CBMN or BMn, but these tests have not gained much popularity, so far.
5. Visibility by International Organization
The future application of the assay gains more interest since its recognition by international protection, normalization and scientific organizations, such as International Atomic Energy Agency (IAEA) [21] and biological dosimetry network “Running the European Network of Biological and retrospective Physical dosimetry” (RENEB), both considering MN as a fine biodosimetry method [102] or OECD and International Organization for Standardization (ISO), which issued the standards of how to use the MN in genotoxicology and biological dosimetry, respectively [2,4,103]. Finally, the International Human Micronucleus Project (HUMN), which is the biggest database of results of MN in lymphocytes and buccal cells, was established [96].
The IAEA has always considered biological dosimetry, including MN, as an important component of the radiation protection system. Under its auspices, subsequent editions of textbooks on this issue were developed in 1986, 2001 and most recently in 2011 [22,104,105]. Recent publication: “Cytogenetic Dosimeters: Applications in preparedness for and Response to Radiation Emergencies”, in EPR-Biodosimeters counts over 200 pages and contains 350 references, which makes it the most comprehensive compendium of knowledge about biological dosimetry techniques, so far [22]. In addition, the IAEA prepared a series of reports on accidents involving radiation exposure that happened worldwide, with a thorough analysis of what had happened, why and what conclusions could be drawn for the future. In some of these accidents, the CBMN was used to estimate the radiation dose [106].
The IAEA supports the development and sustainability of biological dosimetry methods in member states counties. This is realized by, among others, so-called coordinated research activities (CRAs), which aim at acquisition and dissemination of new knowledge and technology generated through the use of nuclear science, radiation and isotopic techniques [107]. In 2017, the new CRA titled: “Applications of biological dosimetry methods in radiation oncology, nuclear medicine, diagnostic and interventional radiology” (MEDBIODOSE) was launched [108]. The goal of this CRA is to prove the feasibility of biological dosimetry methods in relation to the effects of medical procedures involving ionizing radiation and their improvement. The CBMN is considered as one of the most valuable assays for this purpose and is being used in 7 out of 39 subprojects, covering such topics as, biological dosimetry for individually tailored patients’ treatment, biological dosimetry for genotoxicity assessment, prediction of radiation toxicity of normal tissue by ex vivo irradiation or by in vivo biomarkers, and use of CBMN in cancer risk prediction [29].
The European biodosimetry network RENEB was created as the result of two projects: MULTIBIODOSE and RENEB focused on the implementation of cytogenetic tests in Europe [55,102]. In the frame of the MULTIBIODOSE project, CBMN was harmonized at partner’s laboratories and the automatic and semi-automatic versions of the assay was successfully validated [55]. It was proven that the semi-automatic version of the assay gives results as good as manual scoring. Its continuation, the RENEB project has proved that CBMN can be used in a mass radiation exposure scenario giving results in a reasonable time [109]. Various types of interlaboratory comparisons were carried out, testing individual methods of biological dosimetry and reaction of laboratories to network activation. The capability of biological dosimetry in Europe was also estimated, calculated as the number of samples possible to analyze in the given time [110]. Since RENEB cooperates with biological dosimetry networks in Canada, South America, Japan and Asia, an international sample exchange is possible (and was tested) [111,112], the first step to the global biological dosimetry network.
CBMN is present also in the area of interest of organizations issuing standards on the use of research methods in various aspects of everyday life and industry, such as OECD or ISO [2,4,103]. Their technical documents and/or standards are the basis for method accreditation, theoretically guaranteeing that the laboratories performing tests according to these documents receive objectively correct results, comparable to other accredited laboratories. As far as MN is concerned, the OECD has issued two Technical Guides: “474 OECD Guideline for the Testing of Chemicals, Mammalian Erythrocyte Micronucleus Test” and “OECD Library Test No. 487 OECD Guidelines for the Testing of Chemicals, In Vitro Mammalian Cell Micronucleus Test” [2,4], both dedicated to laboratories examining the genotoxicity of drugs and chemicals in vivo and in vitro. It has to be emphasized that genotoxicity studies are an important part of human clinical trials of new medicaments [82]. In turn, ISO issued the ISO 17099:2014 standard “Radiological protection—Performance criteria for laboratories using the cytokinesis block micronucleus (CBMN) assay in peripheral blood lymphocytes for biological dosimetry” [103].
The HUMN project (www.humn.org) was founded in 1997 to coordinate worldwide research efforts aimed at using MN to study DNA damage in the human population [31,96]. For 15 years, several goals have been achieved: 130 laboratories that published papers on the MN assay were identified and 42 of them were cooperating in HUMN, CBMN protocol and scoring criteria were standardized and the connection between the level of Mn and age, gender and smoking status was established. In addition, the median control level of 6.5 Mn per 1000 binucleated cells was found for CBMN in a database of 7000 donors. The prospective study with the same data indicated that increased Mn frequency is associated with the cancer risk.
In 2007, the HUMN project coordinating group decided to launch a new project considering BMN and the project was called HUMNXL, as Mn are scored in buccal eXfoLiated cells [96]. The questionnaire was sent to 188 laboratories all over the world, based on issued publications using BMN. Fifty-eight laboratories decided to cooperate in the project and a database of more than 5000 subjects was collected. BMN protocol and scoring criteria were established. Factors as age, gender and smoking, which can change the control level of Mn was considered. The control level of Mn in the buccal cells is as low as 1.1/1000 cells (95 % CI 0.70–1.72), but to reduce the variability of Mn mean estimates, it is recommended to score 4000 cells (instead 2000, as it is usually done) [94,96]. The interlaboratory comparison had been done and proved that participants correctly recognized samples from persons who had undergone RT, based on frequency of Mn, nuclear buds and differentiated binucleated cells [95].
6. Automation of MN
MN is reliable but a laborious and time-consuming method. One of the ways to speed up the results acquisition is its automation. At the same time, automation increases the reliability of the assay, minimalizing a bias associated with different recognition of Mn by different scorers, different levels of experience and fatigue. Moreover, automation increases the statistical certainty of the results, enabling analysis of a larger number of cells, several thousand or even tens of thousands instead of one, two thousand analyzed in the classic version of the assay [53]. Automation is expected to give higher accuracy and allowed to survey effects on the low observable level, not possible with the manual scoring [82]. However, according to Rodrigues [53], the existing automatic micronucleus scoring systems on microscope slides or by flow cytometry experience technical problems and do not fully meet the requirements of rapid biological dosimetry with a large number of samples to be analyzed.
6.1. Automatic/Semiautomatic Scoring by Microscope Aided Systems
Attempts to automate CBMN have been made many times [113,114]. In our subjective opinion, the analysis of microscopic preparations using the image analysis system Metafer (Metasystems, Germany) [115,116] has gained the greatest popularity so far. The system is commercially available (https://metasystems-international.com/). It consists of a motorized microscope, a CCD camera and a computer program that captures pictures of the preparation and searches for double-nucleated cells with or without Mn (Figure 4). The advantage of the program is that it creates a gallery of pictures and allows the researcher to relocate them on a microscope slide to verify the results obtained by the system. The program was used by most participants of the RENEB project, which proved that reconstruction of the radiation dose using automatic or manual counting gave similar results [55]. The program shortens the analysis of preparations by at least 4 times, allowing scoring 2000 cells in 30–40 min instead of 1000 cells in 2 h for manual counting. The software is suitable for Mn analysis for biological dosimetry, but equally well can be used in in vitro tests carried out according to the OECD 487 Guideline.
Microscope aided systems of scoring Mn face several challenges. To perform automated slide-scoring, the cells must be fixed in the mixture of methanol and acetic acid and left for several hours or overnight to dry out [117,118]. This implicates delay in gaining results. Another problem is that to perform a fine automatic analysis of slides, good quality images and the appropriate density of cells are necessary [117,119]. Both too low and too high number of cells increase false Mn positive scores [116,118]. Finally, only the nuclei are stained on the preparation, not cytoplasm [53]. This leads to the situation that only possible recognition of mononucleated, binucleated and polynucleated cells is achieved by mathematical algorithms.
6.2. Flow Cytometry and Imaging Flow Cytometry Aided Mn Scoring
Besides the microscopic analysis of binucleated cells, attempts had been made to use a flow cytometry method to analyze the frequency of Mn in lymphocytes or other nucleated cells [120,121]. However, the method faces two major obstacles. First, the cells must be lysed because micronuclei are only recognized base on their size. Objects smaller than cell nuclei are gated, but it is uncertain whether these are not debris or apoptotic bodies or necrosis residues [120,122]. Secondly, cytochalasin B is not used in this kind of test because the cells are lysed and thus binucleated cells cannot be analyzed. This approach might underestimate the number of Mn [123].
Problems encountered when using automated slide-scoring systems or flow cytometry are solved by the imaging flow cytometry (IFC) method that is claimed to be a combination of flow cytometry with automatic cell image analysis [53]. The system analyzes the cells flowing individually through the lasers beam, and in the meantime, every object in the solution is photographed. Based on the pictures, objects as mononucleated, binucleated, polynucleated cells, Mn, nuclear bridges and nuclear buds, as well as apoptotic bodies can be recognized. The high throughput is one of the system features [53].
6.3. Automatization of EMn
EMn has been automatized by using the automated analysis of images of microscopic preparations or flow cytometry [82,124,125]. The analysis of microscopic slides is described above, thus here we will briefly describe the use of a flow cytometry technique. The method allows analysis of both columns fractionated bone marrow and peripheral blood samples [126,127]. Cells are stained simultaneously with propidium iodide/Hoechst and anti-CD71 antibody (transferrin receptor characteristic for immature erythrocytes) and anti-platelet-specific antibody (anti-CD61), both differentially labeled with fluorochromes. Single, unaggregated cells are gated, excluding those exhibiting anti-platelet associated fluorescence. Then, CD-71 positive cells are recognized and Mn stained with propidium iodide/Hoechst are scored. The method has been transformed into a commercially available kit format (e.g., In vitro MicroFlow, Litron Laboratories, Rochester, NY, USA). The assay has been widely used [128,129]. It has been investigated and validated for different species samples (mice, rats, human) by The Collaborative Study Group of the Japanese Environmental Mutagen Society [130,131]. The assay is important for human population studies because performing it in blood samples makes it not invasive when compared with bone marrow sampling. Automation made the blood erythrocytes examination feasible and it is possible to consider the damage found in human blood, as an index of damage in the bone marrow.
7. Chromothripsis
Chromothripsis phenomenon—massive changes in the genetic material—hundreds of rearrangements, confined to single or a few chromosomes are important in cancer and congenital diseases development and perhaps in rapid karyotype evolution [39,132,133,134]. A chromothripsis has been found in 2–3% of cancers of all types including 25% of bone cancers and 18% of late-stage neuroblastoma [39,133,134,135,136,137,138,139]. It is suggested that chromothripsis may be the result of one catastrophic event of massive breakage closely followed by rearrangements and repair processes [140,141]. The mechanisms involved and the meaning of phenomena are not fully understood, but the most accepted model of how chromothripsis can be confined to the single chromosome of the chromosome arm is the Mn model [135,142].
Mn has been considered as a result of induction of different kind of DNA damage and a good indicator of chromosomal instability which is always present in pre-cancer cells [30,31,32,33,72]. The replication machinery and DNA repair are impaired in Mn, which in a short time leads to extensive DNA damage [31,142]. Then, if the Mn is included in the cell nucleus again there is a lot of rearranged chromosome material integrated, in most cases derived from a single chromosome and chromothripsis can be initiated. The traditional view of Mn faith is that cells bearing Mn are doomed to death in a close future, after few division latest, as a result of chromosome instability [123], or Mn themselves can be extruded out of the cells [35]. Novel experiments utilizing long-term live-cell imaging show that Mn can be incorporated to the daughter cells during mitosis and stably remains in cytoplasm [142,143,144]. Then, Mn material sometimes is reincorporated to the arising nucleus, which was revealed e.g., by photoactivation of micronuclear chromatin [30,142]. Chromothripsis phenomenon changes traditional thinking about Mn as a passive depiction of genetic material damage processes to the active players in formation of DNA lesions [31].
8. Conclusions
There are obvious reasons to think that Mn is important for future use as one of the most reliable, well established and feasible genotoxicity tests [2,3,4,96]. BMn (described in more detail in Chapter 4) will gain more importance, as a convenient and reliable biomonitoring test, which in the most exposure scenario may be more sensitive than the standard CBMN. The MN is easy to automate, works to fully automatize the method, has been carried out for some time on at least 3 major platforms: automated slide-scoring, flow cytometry and imaging flow cytometry. Certainly, further attempts will be made to determine the relationship between the phenomenon of Mn formation and the development of cancer.
Author Contributions
S.S., I.B. and M.K. wrote the manuscript, revised and prepared the final version. S.S. prepared figures and drawings. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by statutory funding of INCT and IRH.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BMn | Buccal cells micronucleus assay; |
| CBMN | Cytokinesis-block micronucleus assay; |
| CRA | Coordinated Research Activities; |
| DB | Double minutes; |
| EMn | Mammalian erythrocyte micronucleus assay; |
| HUMN | International Human Micronucleus Project; |
| IAEA | International Atomic Energy Agency |
| ISO | International Organization for Standardization; |
| IR | Ionizing radiation; |
| Mn | Micronucleus, micronuclei; |
| MN | Micronucleus assay; |
| OECD | The Organization for Economic Co-operation and Development; |
| RENEB | Running the European Network of Biological and retrospective Physical dosimetry |
| RT | Radiation therapy, radiotherapy; |
References
- Heddle, J.A.; Fenech, M.; Hayashi, M.; MacGregor, J.T. Reflections on the development of micronucleus assays. Mutagenesis 2011, 26, 3–10. [Google Scholar] [CrossRef]
- OECD. Test No. 474: Mammalian Erythrocyte Micronucleus Test, OECD Guideline for the Testing of Chemicals, Section 4, OECD Publishing, Paris. 2016. Available online: https://0-doi-org.brum.beds.ac.uk/10.1787/9789264264762-en (accessed on 20 February 2020).
- Kirsch-Volders, M.; Fenech, M.; Bolognesi, C. Validity of the Lymphocyte Cytokinesis-Block Micronucleus Assay (L-CBMN) as biomarker for human exposure to chemicals with different modes of action: A synthesis of systematic reviews. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 836, 47–52. [Google Scholar] [CrossRef]
- OECD. Test No. 487: In Vitro Mammalian Cell Micronucleus Test, OECD Guideline for the Testing of Chemicals, Section 4, OECD Publishing, Paris. 2016. Available online: https://0-doi-org.brum.beds.ac.uk/10.1787/9789264264861-en (accessed on 20 February 2020).
- Fenech, M. The advantages and disadvantages of the cytokinesis-block micronucleus method. Mutat. Res. 1997, 392, 11–18. [Google Scholar] [CrossRef]
- Fenech, M. The in vitro micronucleus technique. Mutat. Res. 2000, 455, 81–95. [Google Scholar]
- Nersesyan, A.; Fenech, M.; Bolognesi, C.; Mišík, M.; Setayesh, T.; Wultsch, G.; Bonassi, S.; Thomas, P.; Knasmüller, S. Use of the lymphocyte cytokinesis-block micronucleus assay in occupational biomonitoring of genome damage caused by in vivo exposure to chemical genotoxins: Past, present and future. Mutat. Res. 2016, 770, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Maluf, S.W.; Mergener, M.; Dalcanale, L.; Costa, C.C.; Pollo, T.; Kayser, M.; da Silva, L.B.; Pra, D.; Teixeira, P.J. DNA damage in peripheral blood of patients with chronic obstructive pulmonary disease (COPD). Mutat. Res. 2007, 626, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Migliore, L.; Coppedè, F.; Fenech, M.; Thomas, P. Association of micronucleus frequency with neurodegenerative diseases. Mutagenesis 2011, 26, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladeira, C.; Smajdova, L. The use of genotoxicity biomarkers in molecular epidemiology: Applications in environmental, occupational and dietary studies. AIMS Genet. 2017, 4, 166–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkez, H.; Arslan, M.; Ozdemir, O. Genotoxicity testing: Progress and prospects for the next decade. Expert. Opin. Drug Metab. Toxicol. 2017, 13, 1089–1098. [Google Scholar] [CrossRef]
- Bolognesi, C.; Bruzzi, P.; Gismondi, V.; Volpi, S.; Viassolo, V.; Pedemonte, S.; Varesco, L. Clinical Application of Micronucleus Test: A Case-Control Study on the Prediction of Breast Cancer Risk/Susceptibility. PLoS ONE 2014, 9, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.F.; MacGregor, J.T.; Hiatt, R.A.; Hooper, N.K.; Wehr, C.M.; Peters, B.; Goldman, L.R.; Yuan, L.A.; Smith, P.A.; Becker, C.E. Micronucleated erythrocytes as an index of cytogenetic damage in humans: Demographic and dietary factors associated with micronucleated erythrocytes in splenectomized subjects. Cancer Res. 1990, 50, 5049–5054. [Google Scholar] [PubMed]
- Marcon, A.E.; Navoni, J.A.; de Oliveira Galvão, M.F.; Garcia, A.C.F.S.; do Amaral, V.S.; Petta, R.A.; Campos, T.F.D.C.; Panosso, R.; Quinelato, A.L.; de Medeiros, S.R.B. Mutagenic potential assessment associated with human exposure to natural radioactivity. Chemosphere 2017, 167, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Kausar, A.; Giri, S.; Mazumdar, M.; Giri, A.; Roy, P.; Dhar, P. Micronucleus and other nuclear abnormalities among betel quid chewers with or without sadagura, a unique smokeless tobacco preparation, in a population from North-East India. Mutat. Res. 2009, 677, 72–75. [Google Scholar] [CrossRef]
- Cavalcante, D.N.; Sposito, J.C.; Crispim, B.D.; Nascimento, A.V.; Grisolia, A.B. Genotoxic and mutagenic effects of passive smoking and urban air pollutants in buccal mucosa cells of children enrolled in public school. Toxicol. Mech. Methods 2017, 27, 346–351. [Google Scholar] [CrossRef]
- Saks, M.; Upreti, S.; Rajendra, S.V.; Dang, R. Genotoxicity: Mechanisms, Testing Guidelines and Methods. Glob. J. Pharmaceu. Sci. 2017, 555575, 1–6. [Google Scholar] [CrossRef]
- Umbuzeiro, G.A.; Heringa, M.; Zeiger, E. In Vitro Genotoxicity Testing: Significance and Use in Environmental Monitoring. Adv. Biochem. Eng. Biotechnol. 2017, 157, 59–80. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Kishi, M.; Sofuni, T.; Ishidate, M., Jr. Micronucleus tests in mice on 39 food additives and eight miscellaneous chemicals. Food Chem. Toxicol. 1988, 26, 487–500. [Google Scholar] [CrossRef]
- Hall, E.J.; Giaccia, A.J. Radiobiology for the Radiologist, 7th ed.; Wolters Kluver/Lippincott Williams & Wilkins: Philadelfia, PA, USA, 2012; ISBN 978-1-60831-193-4. [Google Scholar]
- Ainsbury, E.A.; Bakhanova, E.; Barquinero, J.F.; Brai, M.; Chumak, V.; Correcher, V.; Darroudi, F.; Fattibene, P.; Gruel, G.; Guclu, I.; et al. Review of retrospective dosimetry techniques for external ionising radiation exposures. Radiat. Prot. Dosimetry 2011, 147, 573–592. [Google Scholar] [CrossRef] [Green Version]
- International Atomic Energy Agency. Cytogenetic Dosimetry: Applications in Preparedness for and Response to Radiation Emergencies; IAEA: Vienna, Austria, 2011. [Google Scholar]
- Sproull, M.T.; Camphausen, K.A.; Koblentz, G.D. Biodosimetry: A Future Tool for Medical Management of Radiological Emergencies. Health Secur. 2017, 15, 599–610. [Google Scholar] [CrossRef]
- Franciesa, F.Z.; Wainwrightc, R.; Pooled, J.; de Leeneere, K.; Coenee, I.; Wiemee, G.; Poirelg, H.A.; Brichardh, B.; Vermeuleni, S.; Vral, A.; et al. Diagnosis of Fanconi Anaemia by ionising radiation- or mitomycin C—Induced micronuclei. DNA Repair. 2018, 61, 17–24. [Google Scholar] [CrossRef]
- Claes, K.; Depuydt, J.; Taylor, A.M.; Last, J.I.; Baert, A.; Schietecatte, P.; Vandersickel, V.; Poppe, B.; de Leeneer, K.; D’Hooghe, M.; et al. Variant ataxia telangiectasia: Clinical and molecular findings and evaluation of radiosensitive phenotypes in a patient and relatives. Neuromolecular Med. 2013, 15, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Torres-Bugarín, O.; Macriz Romero, N.; Ramos Ibarra, M.L.; Flores-García, A.; Valdez Aburto, P.; Zavala-Cerna, M.G. Genotoxic Effect in Autoimmune Diseases Evaluated by the Micronucleus Test Assay: Our Experience and Literature Review. Biomed. Res. Int. 2015, 194031, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Padjas, A.; Kedzierawski, P.; Florek, A.; Kukolowicz, P.; Kuszewski, T.; Góźdz, S.; Lankoff, A.; Wojcik, A.; Lisowska, H. Comparative analysis of three functional predictive assays in lymphocytes of patients with breast and gynaecological cancer treated by radiotherapy. J. Contemp. Brachytherapy. 2012, 4, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Encheva, E.; Deleva, S.; Hristova, R.; Hadjidekova, V.; Hadjieva, T. Investigating micronucleus assay applicability for prediction of normal tissue intrinsic radiosensitivity in gynecological cancer patients. Rep. Pract. Oncol. Radiother. 2011, 17, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Vinnikov, V.; Belyakov, O. Clinical applications of biomarkers of radiation exposure: Limitations and possible solutions through coordinated research. Radiat. Prot. Dosimetry 2019, 1–6. [Google Scholar] [CrossRef]
- Fenech, M.; Morley, A. Measurement of micronuclei in lymphocytes. Mutat. Res. 1985, 147, 29–36. [Google Scholar] [CrossRef]
- Terradas, M.; Martín, M.; Genescà, A. Impaired nuclear functions in micronuclei results in genome instability and chromothripsis. Arch. Toxicol. 2016, 90, 2657–2667. [Google Scholar] [CrossRef]
- Kirsch-Volders, M.; Bonassi, S.; Knasmueller, S.; Holland, N.; Bolognesi, C.; Fenech, M.F. Commentary: Critical questions, misconceptions and a road map for improving the use of the lymphocyte cytokinesis-block micronucleus assay for in vivo biomonitoring of human exposure to genotoxic chemicals-a HUMN project perspective. Mutat. Res. Rev. Mutat. Res. 2014, 759, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Fenech, M.; Kirsch-Volders, M.; Natarajan, A.T.; Surralles, J.; Crott, J.W.; Parry, J.; Norppa, L.H.; Eastmond, D.A.; Tucker, J.D.; Thomas, P. Molecular mechanisms of micronucleus, nucleoplasmic bridge and nuclear bud formation in mammalian and human cells. Mutagenesis 2011, 26, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Gisselsson, D.; Jonson, T.; Petersén, A.; Strömbeck, B.; dal Cin, P.; Höglund, M.; Mitelman, F.; Mertens, F.; Mandahl, N. Telomere dysfunction triggers extensive DNA fragmentation and evolution of complex chromosome abnormalities in human malignant tumors. Proc. Natl. Acad. Sci. USA 2001, 98, 12683–12688. [Google Scholar] [CrossRef] [Green Version]
- Gisselsson, D.; Björk, J.; Höglund, M.; Mertens, F.; dal Cin, P.; Akerman, M.; Mandahl, N. Abnormal nuclear shape in solid tumors reflects mitotic instability. Am. J. Pathol. 2001, 158, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Barker, P.E. Double minutes in human tumor cells. Cancer Genet. Cytogenet. 1982, 5, 81–94. [Google Scholar] [CrossRef]
- Masters, J.; Keeley, B.; Gay, H.; Attardi, G. Variable content of double minute chromosomes is not correlated with degree of phenotype instability in methotrexate-resistant human cell lines. Mol. Cell. Biol. 1982, 2, 498–507. [Google Scholar] [CrossRef] [Green Version]
- Baskin, F.; Rosenberg, R.N.; Dev, V. Correlation of double-minute chromosomes with unstable multidrug cross-resistance in uptake mutants of neuroblastoma cells. Proc. Natl. Acad. Sci. USA 1981, 78, 3654–3658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, P.J.; Greenman, C.D.; Fu, B.; Yang, F.; Bignell, G.R.; Mudie, L.J.; Pleasance, E.D.; Lau, K.W.; Beare, D.; Stebbings, L.A.; et al. Massive Genomic Rearrangement Acquired in a Single Catastrophic Event during Cancer Development. Cell 2011, 144, 27–40. [Google Scholar] [CrossRef]
- Kanda, T.; Sullivan, K.F.; Wahl, G.M. Histone–GFP fusion protein enables sensitive analysis of chromosome dynamics in living mammalian cells. Curr. Biol. 1988, 8, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Kanda, T.; Wahl, G.M. The dynamics of acentric chromosomes in cancer cells revealed by GFP-based chromosome labeling strategies. J. Cell Biochem. Suppl. 2000, 35, 107–114. [Google Scholar] [CrossRef]
- Scherlach, K.; Boettger, D.; Remme, N.; Hertweck, C. The chemistry and biology of cytochalasans. Natural. Product. Reports 2010, 27, 869–886. [Google Scholar] [CrossRef]
- Theodoropoulos, P.A.; Gravanis, A.; Tsapara, A.; Margioris, A.N.; Papadogiorgaki, E.; Galanopoulos, V.; Stournaras, C. Cytochalasin B may shorten actin filaments by a mechanism independent of barbed end capping. Biochem. Pharmacol. 1994, 47, 1875–1881. [Google Scholar] [CrossRef]
- Kirsch-Volders, M.; Sofuni, T.; Aardema, M.; Albertini, S.; Eastmond, D.; Fenech, M.; Ishidate, M.; Lorge, E.; Norppa, H.; Surralles, J.; et al. Report from the In Vitro Micronucleus Assay Working Group. Environ. Mol. Mutagen. 2000, 35, 167–172. [Google Scholar] [CrossRef]
- Fenech, M.; Morley, A.A. Cytokinesis-block micronucleus method in human lymphocytes: Effect of in vivo ageing and low-dose x-irradiation. Mutat. Res. 1986, 161, 193–198. [Google Scholar] [CrossRef]
- Wakata, A.; Sasaki, M.S. Measurement of micronuclei by cytokinesis-block method in cultured Chinese hamster cells: Comparison with types and rates of chromosome aberrations. Mutat. Res. 1987, 190, 51–57. [Google Scholar] [CrossRef]
- Prosser, J.S.; Moquet, J.E.; Lloyd, D.C.; Edwards, A.A. Radiation induction of micronuclei in human lymphocytes. Mutat. Res. 1988, 199, 37–45. [Google Scholar] [CrossRef]
- Lindholm, C.; Norrpa, H.; Hayashi, M.; Sorsa, M. Induction of micronuclei and anaphase aberrations by cytochalasin-B in human lymphocyte cultures. Mutat. Res. 1991, 260, 369–375. [Google Scholar] [CrossRef]
- Fenech, M. A mathematical model of the in vitro micronucleus assay predicts false negative results if micronuclei are not specifically scored in binucleated cells or in cells that have completed one nuclear division. Mutagenesis 2000, 15, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.R.; Kim, T.H.; Ryu, S.Y.; Lee, H.J.; Oh, H.; Jo, S.K.; Oh, K.S.; Park, I.C.; Kim, J.C.; Kang, C.M.; et al. Measurement of micronuclei by cytokinesis-block method in human, cattle, goat, pig, rabbit, chicken and fish peripheral blood lymphocytes irradiated in vitro with gamma radiation. In Vivo 2003, 17, 433–438. [Google Scholar]
- Catena, C.; Conti, D.; Villani, P.; Nastasi, R.; Archilei, R.; Righi, E. Micronuclei and 3AB index in human and canine lymphocytes after in vitro X-irradiation. Mutat. Res. 1994, 312, 1–8. [Google Scholar] [CrossRef]
- Resendes, A.S.; dos Santos, D.S.; França, F.M.; Petesse, M.L.; Badaró-Pedroso, C.; Ferreira, C.M. Acute toxic and genotoxic effects of formalin in Danio rerio (zebrafish). Ecotoxicology 2018, 27, 1379–1386. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Beaton-Green, L.A.; Wilkins, R.C.; Fenech, M.F. The potential for complete automated scoring of the cytokinesis block micronucleus cytome assay using imaging flow cytometry. Mutat. Res. Gen. Tox. En. 2018, 836, 53–64. [Google Scholar] [CrossRef]
- Wang, Q.; Rodrigues, M.A.; Repin, M.; Pampou, S.; Beaton-Green, L.A.; Perrier, J.; Garty, G.; Brenner, D.J.; Turner, H.C.; Wilkins, R.C. Automated Triage Radiation Biodosimetry: Integrating Imaging Flow Cytometry with High-Throughput Robotics to Perform the Cytokinesis-Block Micronucleus Assay. Radiat. Res. 2019, 191, 342–351. [Google Scholar] [CrossRef]
- Thierens, H.; Vral, A.; Vandevoorde, C.; Vandersickel, V.; de Gelder, V.; Romm, H.; Oestreicher, U.; Rothkamm, K.; Barnard, S.; Ainsbury, E.; et al. Is a semi-automated approach indicated in the application of the automated micronucleus assay for triage purposes? Radiat. Prot. Dosimetry 2014, 159, 87–94. [Google Scholar] [CrossRef]
- Kirsch-Volders, M. Towards a validation of the micronucleus test. Mutat. Res. 1997, 392, 1–4. [Google Scholar] [CrossRef]
- Parry, J.M.; Sors, A. The detection and assessment of the aneugenic potential of environmental chemicals: The European Community aneuploidy project. Mutat. Res. 1993, 287, 3–15. [Google Scholar] [CrossRef]
- Rodrigues, A.S.; Oliveira, N.G.; Gil, O.M.; Léonard, A.; Rueff, J. Use of cytogenetic indicators in radiobiology. Radiat. Prot. Dosim. 2005, 115, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.J.; Sharpe, Z.; Alemara, S.; Mackenzie, S.; Liu, G.; Abdallah, B.; Horne, S.; Regan, S.; Heng, H.H. Micronuclei and Genome Chaos: Changing the System Inheritance. Genes 2019, 10, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.; Jeggo, P.A.; West, C.; Gomolka, M.; Quintens, R.; Badie, C.; Laurent, O.; Aerts, A.; Anastasov, N.; Azimzadeh, O.; et al. Ionizing radiation biomarkers for epidemiological studies—An update. Mutat. Res. 2017, 771, 59–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baert, A.; Depuydt, J.; van Maerken, T.; Poppe, B.; Malfait, F.; Storm, K.; van den Ende, J.; Van Damme, T.; De Nobele, S.; Perletti, G.; et al. Increased chromosomal radiosensitivity in asymptomatic carriers of a heterozygous BRCA1 mutation. Breast Cancer Res. 2016, 18, 52. [Google Scholar] [CrossRef] [Green Version]
- Vral, A.; Thierens, H.; de Ridder, L. Micronucleus induction by 60Co γ-rays and fast neutrons in ataxia telangiectasia lymphocytes. Int. J. Radiat. Biol. 1996, 70, 171–176. [Google Scholar] [CrossRef]
- Okunieff, P.; Chen, Y.; Maguire, D.J.; Huser, A.K. Molecular markers of radiation related normal tissue toxicity. Cancer Metastasis Rev. 2008, 27, 363–374. [Google Scholar] [CrossRef]
- Bentzen, S.M.; Parliament, M.; Deasy, J.O.; Dicker, A.; Curran, W.J.; Williams, J.P.; Rosenstein, B.S. Biomarkers and surrogate endpoints for normal-tissue effects of radiation therapy: The importance of dose-volume effects. Int. J. Radiat. Oncol. Biol. Phys. 2010, 76, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Bonassi, S.; Znaor, A.; Ceppi, M.; Lando, C.; Chang, W.P.; Holland, N.; Kirsch-Volders, M.; Zeiger, E.; Ban, S.; Barale, R.; et al. An increased micronucleus frequency in peripheral blood lymphocytes predicts the risk of cancer in humans. Carcinogenesis 2007, 28, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Bolognesi, C.; Bonassi, S.; Knasmueller, S.; Fenech, M.; Bruzzone, M.; Lando, C.; Ceppi, M. Clinical application of micronucleus test in exfoliated buccal cells: A systematic review and metanalysis. Mutat. Res. 2015, 766, 20–31. [Google Scholar] [CrossRef]
- McClelland, S.E. Role of chromosomal instability in cancer progression. Endocr. Relat. Cancer 2017, 24, T23–T31. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Rondón, N.; Villegas, V.E.; Rondón-Lagos, M. The role of chromosomal instability in cancer and therapeutic responses. Cancers 2017, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wild, C.P.; Scalbert, A.; Herceg, Z. Measuring the exposome: A powerful basis for evaluating environmental exposures and cancer risk. Environ. Mol. Mutagen. 2013, 54, 480–499. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, S.M.; Smith, M.T. Epidemiology, Environment and disease risks. Science 2010, 330, 460–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, V.A.; Nersesyan, A.K.; Atefie, K.; Hoelzl, C.; Ferk, F.; Bichler, J.; Valic, E.; Schaffer, A.; Schulte-Hermann, R.; Fenech, M.; et al. Inhalativeexposure to vanadium pentoxide causes DNA damage in workers: Results of amultiple end point study. Environ. Health Perspect. 2008, 116, 1689–1693. [Google Scholar]
- Fenech, M. Cytokinesis-block micronucleus cytome assay. Nat Protoc. 2007, 2, 1084–1104. [Google Scholar] [CrossRef] [Green Version]
- Von Ledebur, M.; Schmid, W. The micronucleus test: Methodological aspects. Mutat. Res. 1973, 19, 109–117. [Google Scholar] [CrossRef]
- MacGregor, J.T.; Wehr, C.M.; Gould, D.H. Clastogen-induced micronuclei in peripheral blood erythrocytes: The basis of an improved micronucleus test. Environ. Mutagen. 1980, 2, 509–514. [Google Scholar] [CrossRef]
- Abe, T.; Isemura, T.; Kikuchi, Y. Micronuclei in human bone-marrow cells: Evaluation of the micronucleus test using human leukemia patients treated with antileukemic agents. Mutat. Res. 1984, 130, 113–120. [Google Scholar] [CrossRef]
- Goetz, P.; Šrám, R.J.; Dohnalová, J. Relationship between experimental results in mammals and man: Cytogenetic analysis of bone marrow injury induced by a single dose of cyclophosphamide. Mutat. Res. 1975, 31, 247–254. [Google Scholar] [CrossRef]
- Udroiu, I. Feasibility of conducting the micronucleus test in circulating erythrocytes from different mammalian species: An anatomical perspective. Environ. Mol. Mutagen. 2006, 47, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Polard, T.; Jean, S.; Merlina, G.; Laplanche, C.; Pinelli, E.; Gauthier, L. Giemsa versus acridine orange staining in the fish micronucleus assay and validation for use in water quality monitoring. Ecotoxicol. Environ. Saf. 2011, 74, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.M.; Taketomi, M.; Nakajima, M. Effect of aging on spontaneous micronucleus frequencies in peripheral blood of nine mouse strains: The results of the 7th collaborative study organized by CSGMT/MMS. Mutat. Res. 1995, 338, 51–57. [Google Scholar] [CrossRef]
- Dobrzyńska, M.; Gajowik, A.; Radzikowska, J.; Lankoff, A.; Dušinská, M.; Kruszewski, M. Genotoxicity of silver and titanium dioxide nanoparticles in bone marrow cells of rats in vivo. Toxicology 2014, 315, 86–91. [Google Scholar] [CrossRef]
- Cao, Y.; Xu, Q.; Jin, Z.D.; Zhou, Z.; Nie, J.H.; Tong, J. Induction of adaptive response: Pre-exposure of mice to 900 MHz radiofrequency fields reduces hematopoietic damage caused by subsequent exposure to ionising radiation. Int. J. Radiat. Biol. 2011, 87, 720–728. [Google Scholar] [CrossRef]
- Dertinger, S.D.; Torous, D.K.; Hayashi, M.; MacGregor, J.T. Flow cytometric scoring of micronucleated erythrocytes: An efficient platform for assessing in vivo cytogenetic damage. Mutagenesis 2011, 26, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, R.; MacGregor, J.T.; Everson, R.B. Assessment of cytogenetic damage by quantitation of micronuclei in human peripheral blood erythrocytes. Cancer Res. 1986, 46, 3717–3721. [Google Scholar]
- Abramsson-Zetterberg, L.; Zetterberg, G.; Bergqvist, M.; Grawé, J. Human cytogenetic biomonitoring using flow-cytometric analysis of micronuclei in transferrin-positive immature peripheral blood reticulocytes. Environ. Mol. Mutagen. 2000, 36, 22–31. [Google Scholar] [CrossRef]
- Hayashi, M.; MacGregor, J.T.; Gatehouse, D.G.; Blakey, D.H.; Dertinger, S.D.; Abramsson-Zetterberg, L.; Krishna, G.; Morita, T.; Russo, A.; Asano, N.; et al. In Vivo Micronucleus Assay Working Group, IWGT. In vivo erythrocyte micronucleus assay III. Validation and regulatory acceptance of automated scoring and the use of rat peripheral blood reticulocytes, with discussion of non-hematopoietic target cells and a single dose-level limit test. Mutat. Res. 2007, 627, 10–30. [Google Scholar] [CrossRef] [PubMed]
- Stich, H.F.; San, R.H.; Rosin, M.P. Adaptation of the DNA-repair and micronucleus tests to human cell suspensions and exfoliated cells. Ann. N. Y. Acad. Sci. 1983, 407, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Stich, H.F.; Rosin, M.P. Quantitating the synergistic effect of smoking and alcohol consumption with the micronucleus test on human buccal mucosa cells. Int. J. Cancer. 1983, 31, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Holland, N.; Bolognesi, C.; Kirsch-Volders, M.; Bonassi, S.; Zeiger, E.; Knasmueller, S.; Fenech, M. Buccal micronucleus cytome assay. Nat. Protoc. 2009, 4, 825–837. [Google Scholar] [CrossRef]
- Thomas, P.; Fenech, M. Buccal micronucleus cytome assay. Methods Mol. Biol. 2011, 682, 235–248. [Google Scholar] [CrossRef]
- Holland, N.; Bolognesi, C.; Kirsch-Volders, M.; Bonassi, S.; Zeiger, E.; Knasmueller, S.; Fenech, M. The micronucleus assay in human buccal cells as a tool for biomonitoring DNA damage: The HUMN project perspective on current status and knowledge gaps. Mutat. Res. 2008, 659, 93–108. [Google Scholar] [CrossRef]
- Bolognesi, C.; Knasmueller, S.; Nersesyan, A.; Thomas, P.; Fenech, M. The HUMNxl scoring criteria for different cell types and nuclear anomalies in the buccal micronucleus cytome assay—An update and expanded photogallery. Mutat. Res. 2013, 753, 100–113. [Google Scholar] [CrossRef]
- Bolognesi, C.; Fenech, M. Micronucleus Cytome Assays in Human Lymphocytes and Buccal Cells. Methods Mol. Biol. 2019, 2031, 147–163. [Google Scholar] [CrossRef]
- De Oliveira, F.M.; Carmona, A.M.; Ladeira, C. Is mobile phone radiation genotoxic? An analysis of micronucleus frequency in exfoliated buccal cells. Mutat. Res. 2017, 822, 41–46. [Google Scholar] [CrossRef]
- Ceppi, M.; Biasotti, B.; Fenech, M.; Bonassi, S. Human population studies with the exfoliated buccal micronucleus assay: Statistical and epidemiological issues. Mutat Res. 2010, 705, 11–19. [Google Scholar] [CrossRef]
- Bolognesi, C.; Knasmueller, S.; Nersesyan, A.; Roggieri, P.; Ceppi, M.; Bruzzone, M.; Blaszczyk, E.; Mielzynska-Svach, D.; Milic, M.; Bonassi, S.; et al. Inter-laboratory consistency and variability in the buccal micronucleus cytome assay depends on biomarker scored and laboratory experience: Results from the HUMNxl international inter-laboratory scoring exercise. Mutagenesis 2017, 32, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenech, M.; Holland, N.; Zeiger, E.; Chang, W.P.; Burgaz, S.; Thomas, P.; Bolognesi, C.; Knasmueller, S.; Kirsch-Volders, M.; Bonassi, S. The HUMN and HUMNxL international collaboration projects on human micronucleus assays in lymphocytes and buccal cells—Past, present and future. Mutagenesis 2011, 26, 239–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linhares, D.P.S.; Garcia, P.V.; Silva, C.; Barroso, J.; Kazachkova, N.; Pereira, R.; Lima, M.; Camarinho, R.; Ferreira, T.; Dos Santos Rodrigues, A. DNA damage in oral epithelial cells of individuals chronically exposed to indoor radon (222Rn) in a hydrothermal area. Environ. Geochem. Health 2018, 40, 1713–1724. [Google Scholar] [CrossRef] [PubMed]
- Aguiar Torres, L.; Dos Santos Rodrigues, A.; Linhares, D.; Camarinho, R.; Nunes Páscoa Soares Rego, Z.M.; Ventura Garcia, P. Buccal epithelial cell micronuclei: Sensitive, non-invasive biomarkers of occupational exposure to low doses of ionizing radiation. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 838, 54–58. [Google Scholar] [CrossRef]
- Nersesyan, A.; Kundi, M.; Fenech, M.; Bolognesi, C.; Misik, M.; Wultsch, G.; Hartmann, M.; Knasmueller, S. Micronucleus assay with urine derived cells (UDC): A review of its application in human studies investigating genotoxin exposure and bladder cancer risk. Mutat. Res. Rev. Mutat. Res. 2014, 762, 37–51. [Google Scholar] [CrossRef]
- Kesimci, E.; Çoşkun, E.; Uğur, G.; Müderris, T.; İzdeş, S.; Karahalil, B. Can Sevoflurane Induce Micronuclei Formation in Nasal Epithelial Cells of Adult Patients? Turk. J. Anaesthesiol. Reanim. 2017, 45, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Bonassi, S.; Milić, M.; Neri, M. Frequency of micronuclei and other biomarkers of DNA damage in populations exposed to dusts, asbestos and other fibers. A systematic review. Mutat. Res. 2016, 770, 106–118. [Google Scholar] [CrossRef]
- Kulka, U.; Abend, M.; Ainsbury, E.; Badie, C.; Barquinero, J.F.; Barrios, L.; Beinke, C.; Bortolin, E.; Cucu, A.; De Amicis, A.; et al. RENEB—Running the European Network of biological dosimetry and physical retrospective dosimetry. Int. J. Radiat. Biol. 2017, 93, 2–14. [Google Scholar] [CrossRef] [Green Version]
- ISO 17099:2014. Radiological Protection—Performance Criteria for Laboratories Using the Cytokinesis Block Micronucleus (CBMN) Assay in Peripheral Blood Lymphocytes for Biological Dosimetry. 2014. Available online: https://www.iso.org/standard/59141.html (accessed on 20 February 2020).
- International Atomic Energy Agency. Biological Dosimetry: Chromosomal Aberration Analysis for Dose Assessment, Technical Reports Series No. 260; IAEA: Vienna, Austria, 1986. [Google Scholar]
- International Atomic Energy Agency. Cytogenetic Analysis for Radiation Dose Assessment, Technical Reports Series No. 405; IAEA: Vienna, Austria, 2001. [Google Scholar]
- International Atomic Energy Agency. The Radiological Accident in Istanbul; IAEA: Vienna, Austria, 2000. [Google Scholar]
- International Atomic Energy Agency Factsheet. IAEA Coordinated Research Activities, IAEA Office of Public Information and Communication, August 2019. Available online: https://www.iaea.org/sites/default/files/19/09/iaea-coordinated-research-activities.pdf (accessed on 20 February 2020).
- Applications of Biological Dosimetry Methods in Radiation Oncology, Nuclear Medicine, and Diagnostic and Interventional Radiology (MEDBIODOSE). Available online: https://www.iaea.org/projects/crp/e35010 (accessed on 20 February 2020).
- Depuydt, J.; Baeyens, A.; Barnard, S.; Beinke, C.; Benedek, A.; Beukes, P.; Buraczewska, I.; Darroudi, F.; De Sanctis, S.; Dominguez, I.; et al. RENEB intercomparison exercises analyzing micronuclei (Cytokinesis-block Micronucleus Assay). Int. J. Radiat. Biol. 2017, 93, 36–47. [Google Scholar] [CrossRef]
- Brzozowska, B.; Ainsbury, E.; Baert, A.; Beaton-Green, L.; Barrios, L.; Barquinero, J.F.; Bassinet, C.; Beinke, C.; Benedek, A.; Beukes, P.; et al. RENEB accident simulation exercise. Int J Radiat Biol. 2017, 93, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Ainsbury, E.A.; Al-Hafidh, J.; Bajinskis, A.; Barnard, S.; Barquinero, J.F.; Beinke, C.; de Gelder, V.; Gregoire, E.; Jaworska, A.; Lindholm, C.; et al. Inter- and intra-laboratory comparison of a multibiodosimetric approach to triage in a simulated, large scale radiation emergency. Int. J. Radiat. Biol. 2014, 90, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kulka, U.; Ainsbury, L.; Atkinson, M.; Barnard, S.; Smith, R.; Barquinero, J.F.; Barrios, L.; Bassinet, C.; Beinke, C.; Cucu, A.; et al. Realising the European network of biodosimetry: RENEB-status quo. Radiat Prot Dosim. 2015, 164, 42–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decordier, I.; Papine, A.; Vande Loock, K.; Plas, G.; Soussaline, F.; Kirsch-Volders, M. Automated image analysis of micronuclei by IMSTAR for biomonitoring. Mutagenesis 2011, 26, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frieauff, W.; Martus, H.J.; Suter, W.; Elhajouji, A. Automatic analysis of the micronucleus test in primary human lymphocytes using image analysis. Mutagenesis 2013, 28, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Schunck, C.; Johannes, T.; Varga, D.; Lörch, T.; Plesch, A. New developments in automated cytogenetic imaging: Unattended scoring of dicentric chromosomes, micronuclei, single cell gel electrophoresis, and fluorescence signals. Cytogenet Genome Res. 2004, 104, 383–389. [Google Scholar] [CrossRef]
- Rossnerova, A.; Spatova, M.; Schunck, C.; Sram, R.J. Automated scoring of lymphocyte micronuclei by the MetaSystems Metafer image cytometry system and its application in studies of human mutagen sensitivity and biodosimetry of genotoxin exposure. Mutagenesis 2011, 26, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Seager, A.L.; Shah, U.K.; Brüsehafer, K.; Wills, J.; Manshian, B.; Chapman, K.E.; Thomas, A.D.; Scott, A.D.; Doherty, A.T.; Doak, S.H.; et al. Recommendations, evaluation and validation of a semi-automated, fluorescent-based scoring protocol for micronucleus testing in human cells. Mutagenesis 2014, 29, 155–164. [Google Scholar] [CrossRef]
- Decordier, I.; Papine, A.; Plas, G.; Roesems, S.; Vande Loock, K.; Moreno-Palomo, J.; Cemeli, E.; Anderson, D.; Fucic, A.; Marcos, R.; et al. Automated image analysis of cytokinesis-blocked micronuclei: An adapted protocol and a validated scoring procedure for biomonitoring. Mutagenesis 2009, 24, 85–93. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Smolewski, P.; Holden, E.; Luther, E.; Henriksen, M.; François, M.; Leifert, W.; Fenech, M. Laser scanning cytometry for automation of the micronucleus assay. Mutagenesis 2011, 26, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Nüsse, M.; Marx, K. Flow cytometric analysis of micronuclei in cell cultures and human lymphocytes: Advantages and disadvantages. Mutat. Res. 1997, 392, 109–115. [Google Scholar] [CrossRef]
- Schreiber, G.A.; Beisker, W.; Bauchinger, M.; Nüsse, M. Multiparametric flow cytometric analysis of radiation-induced micronuclei in mammalian cell cultures. Cytometry 1992, 13, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Avlasevich, S.L.; Bryce, S.M.; Cairns, S.E.; Dertinger, S.D. In vitro micronucleus scoring by flow cytometry: Differential staining of micronuclei versus apoptotic and necrotic chromatin enhances assay reliability. Environ. Mol. Mutagen. 2006, 47, 56–66. [Google Scholar] [CrossRef]
- Hintzsche, H.; Hemmann, U.; Poth, A.; Utesch, D.; Lott, J.; Stopper, H.; Working Group “In vitro micronucleus test”; Gesellschaft für Umwelt-Mutationsforschung (GUM, German-speaking section of the European Environmental Mutagenesis and Genomics Society EEMGS). Fate of micronuclei and micronucleated cells. Mutat. Res. 2017, 771, 85–98. [Google Scholar] [CrossRef]
- Romagna, F.; Staniforth, C.D. The automated bone marrow micronucleus test. Mutat. Res. 1989, 213, 91–104. [Google Scholar] [CrossRef]
- Frieauff, W.; Romagna, F. Technical aspects of automatic micronucleus analysis in rodent bone marrow assays. Cell. Biol. Toxicol. 1994, 10, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Dertinger, S.D.; Torous, D.K.; Tometsko, K.R. Simple and reliable enumeration of micronucleated reticulocytes with a single-laser flow cytometer. Mutat. Res. 1996, 371, 283–292. [Google Scholar] [CrossRef]
- Dertinger, S.D.; Miller, R.K.; Brewer, K.; Smudzin, T.; Torous, D.K.; Roberts, D.J.; Avlasevich, S.L.; Bryce, S.M.; Sugunan, S.; Chen, Y. Automated human blood micronucleated reticulocyte measurements for rapid assessment of chromosomal damage. Mutat. Res. 2007, 626, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Zeiger, E.; Recio, L.; Fennell, T.R.; Haseman, J.K.; Snyder, R.W.; Friedman, m. Investigation of the low-dose response in the in vivo induction of micronuclei and adducts by acrylamide. Toxicol. Sci. 2009, 107, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Cammerer, Z.; Schumacher, M.M.; Kirsch-Volders, M.; Suter, W.; Elhajouji, A. Flow cytometry peripheral blood micronucleus test in vivo: Determination of potential thresholds for aneuploidy induced by spindle poisons. Environ. Mol. Mutagen. 2010, 51, 278–284. [Google Scholar] [CrossRef]
- Wakata, A.; Miyamae, Y.; Sato, S.; Suzuki, T.; Morita, T.; Asano, N.; Awogi, T.; Kondo, K.; Hayashi, M. Evaluation of the rat micronucleus test with bone marrow and peripheral blood: Summary of the 9th collaborative study by CSGMT/JEMS. MMS. Collaborative Study Group for the Micronucleus Test. Environmental Mutagen Society of Japan. Mammalian Mutagenicity Study Group. Environ. Mol. Mutagen. 1998, 32, 84–100. [Google Scholar]
- Hamada, S.; Sutou, S.; Morita, T.; Wakata, A.; Asanami, S.; Hosoya, S.; Ozawa, S.; Kondo, K.; Nakajima, M.; Shimada, H.; et al. Evaluation of the rodent micronucleus assay by a 28-day treatment protocol: Summary of the 13th Collaborative Study by the Collaborative Study Group for the Micronucleus Test (CSGMT)/Environmental Mutagen Society of Japan (JEMS)-Mammalian Mutagenicity Study Group (MMS). Environ Mol. Mutagen. 2001, 37, 93–110. [Google Scholar] [PubMed]
- Maher, C.A.; Wilson, R.K. Chromothripsis and Human Disease: Piecing Together the Shattering Process. Cell 2012, 148, 29–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloosterman, W.P.; Cuppen, E. Chromothripsis in congenital disorders and cancer: Similarities and differences. Curr Opin Cell Biol. 2013, 25, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, M.L.; Zhang, C.Z.; Pellman, D. Chromothripsis: A New Mechanism for Rapid Karyotype Evolution. Annu. Rev. Genet. 2015, 49, 183–211. [Google Scholar] [CrossRef] [PubMed]
- Forment, J.V.; Kaidi, A.; Jackson, S.P. Chromothripsis and cancer: Causes and consequences of chromosome shattering. Nature Reviews Cancer. 2012, 12, 663–670. [Google Scholar] [CrossRef]
- Molenaar, J.J.; Koster, J.; Zwijnenburg, D.A.; van Sluis, P.; Valentijn, L.J.; van der Ploeg, I.; Hamdi, M.; van Nes, J.; Westerman, B.A.; van Arkel, J.; et al. Sequencing of neuroblastoma identifies chromothripsis and defects in neuritogenesis genes. Nature 2012, 483, 589–593. [Google Scholar] [CrossRef]
- Cai, H.; Kumar, N.; Bagheri, H.C.; von Mering, C.; Robinson, M.D.; Baudis, M. Chromothripsis-like patterns are recurring but heterogeneously distributed features in a survey of 22,347 cancer genome screens. BMC Genom. 2014, 15, 82. [Google Scholar] [CrossRef] [Green Version]
- Waddell, N.; Pajic, M.; Patch, A.M.; Chang, D.K.; Kassahn, K.S.; Bailey, P.; Johns, A.L.; Miller, D.; Nones, K.; Quek, K.; et al. Whole genomes redefine the mutational landscape of pancreatic cancer. Nature 2015, 518, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Patch, A.M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; Bailey, P.J.; et al. Whole-genome characterization of chemoresistant ovarian cancer. Nature 2015, 527, 398. [Google Scholar] [CrossRef]
- Meyerson, M.; Pellman, D. Cancer genomes evolve by pulverizing single chromosomes. Cell 2011, 144, 9–10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Z.; Leibowitz, M.L.; Pellman, D. Chromothripsis and beyond: Rapid genome evolution from complex chromosomal rearrangements. Genes Dev. 2013, 27, 2513–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crasta, K.; Ganem, N.J.; Dagher, R.; Lantermann, A.B.; Ivanova, E.V.; Pan, Y.; Nezi, L.; Protopopov, A.; Chowdhury, D.; Pellman, D. DNA breaks and chromosome pulverization from errors in mitosis. Nature 2012, 482, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Spektor, A.; Cornils, H.; Francis, J.M.; Jackson, E.K.; Liu, S.; Meyerson, M.; Pellman, D. Chromothripsis from DNA damage in micronuclei. Nature 2015, 522, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Hou, H.; Yi, Q.; Zhang, Y.; Chen, D.; Jiang, E.; Xia, Y.; Fenech, M.; Shi, Q. The fate of micronucleated cells post X-irradiation detected by live cell imaging. DNA Repair (Amst). 2011, 10, 629–638. [Google Scholar] [CrossRef] [PubMed]
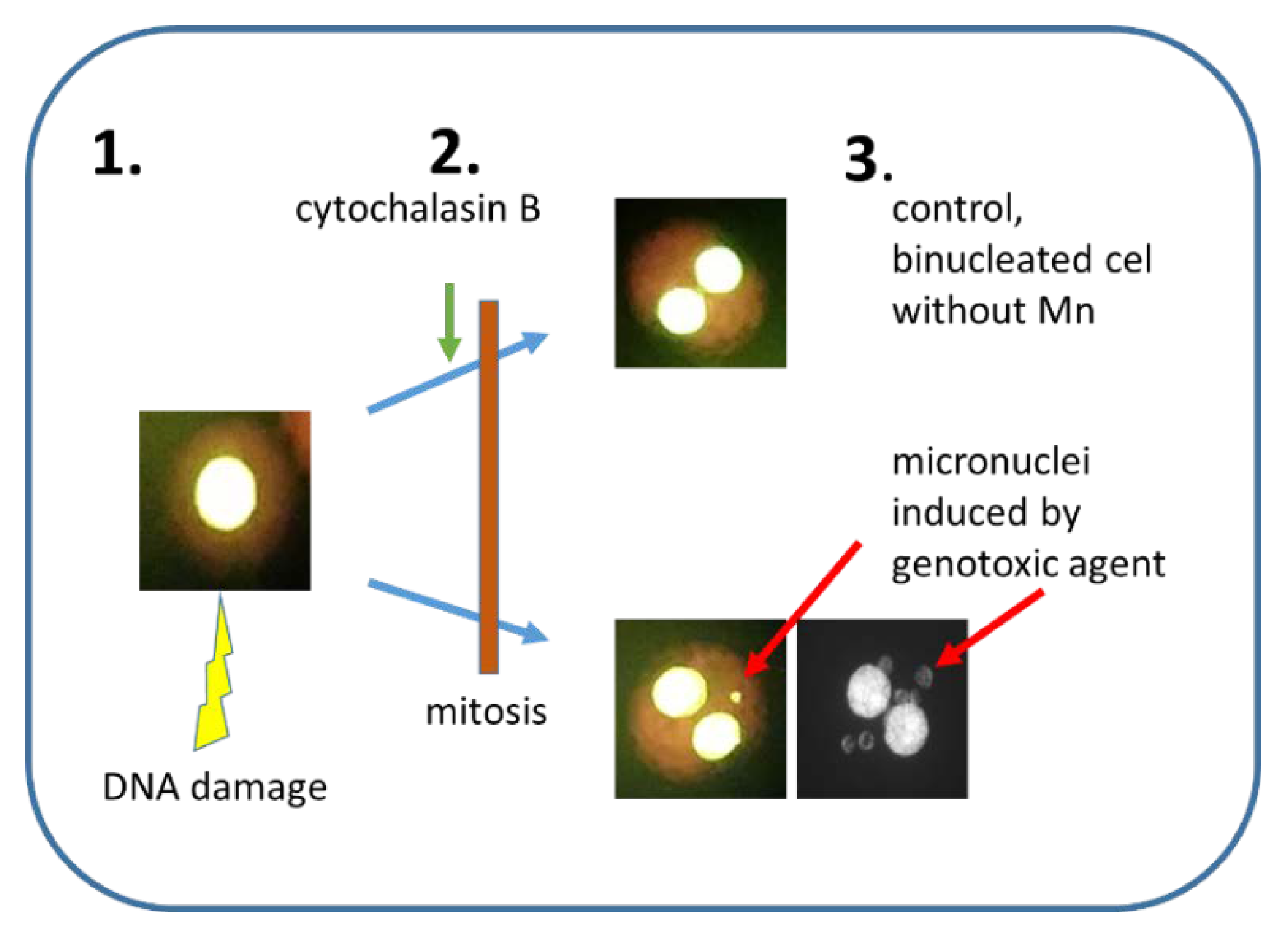
Figure 1.
A principle of cytokinesis-block micronucleus assay. 1. Nucleus with damaged DNA. 2. Inhibition of cytokinesis by the addition of cytochalasin B. 3. The Mn frequency is scored in binucleated cells only. Upper part—control binucleated cells without Mn, lower part—two binucleated cells with 1 or 6 Mn visible in the cytoplasm.
Figure 1.
A principle of cytokinesis-block micronucleus assay. 1. Nucleus with damaged DNA. 2. Inhibition of cytokinesis by the addition of cytochalasin B. 3. The Mn frequency is scored in binucleated cells only. Upper part—control binucleated cells without Mn, lower part—two binucleated cells with 1 or 6 Mn visible in the cytoplasm.

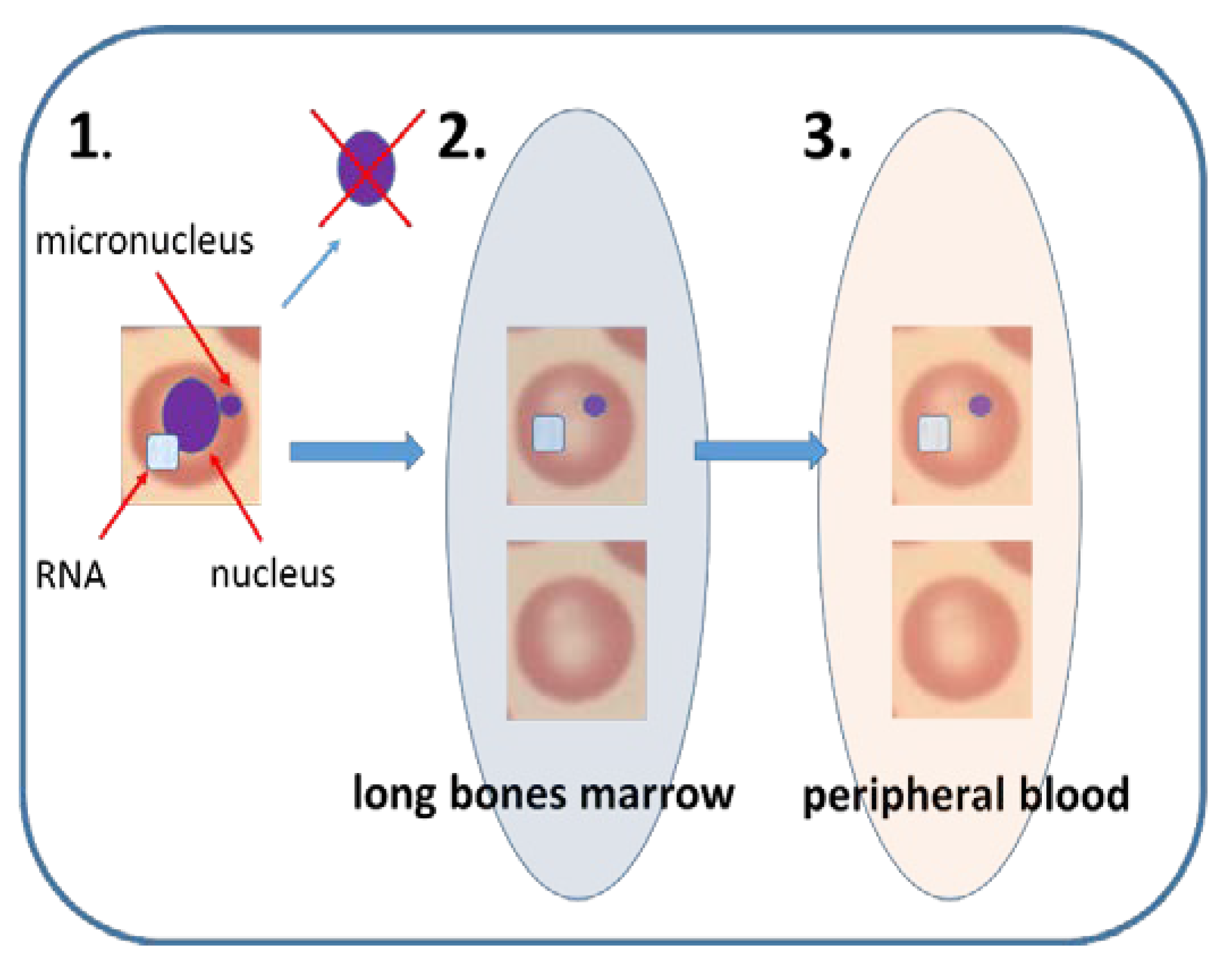
Figure 2.
Mammalian erythrocyte micronucleus assay. (1). Immature erythrocyte in bone marrow contains nucleus and RNA in its cytoplasm. When DNA damage is induced in vivo, micronucleus can arise in the nucleated erythrocyte. When nucleus is excluded during erythrocyte maturation the micronucleus stays in the cytoplasm. (2). In bone marrow, immature erythrocytes consist of around 50% of all erythrocytes. Occasionally these immature erythrocytes may contain Mn. (3). Sometimes the immature erythrocytes are released to peripheral blood, where they constitute less than 5% of all erythrocytes. The immature erythrocytes in blood can be recognized due to their specific surface receptors or RNA content. Flow cytometry technique makes the EMn feasible in peripheral blood of rodents and humans.
Figure 2.
Mammalian erythrocyte micronucleus assay. (1). Immature erythrocyte in bone marrow contains nucleus and RNA in its cytoplasm. When DNA damage is induced in vivo, micronucleus can arise in the nucleated erythrocyte. When nucleus is excluded during erythrocyte maturation the micronucleus stays in the cytoplasm. (2). In bone marrow, immature erythrocytes consist of around 50% of all erythrocytes. Occasionally these immature erythrocytes may contain Mn. (3). Sometimes the immature erythrocytes are released to peripheral blood, where they constitute less than 5% of all erythrocytes. The immature erythrocytes in blood can be recognized due to their specific surface receptors or RNA content. Flow cytometry technique makes the EMn feasible in peripheral blood of rodents and humans.

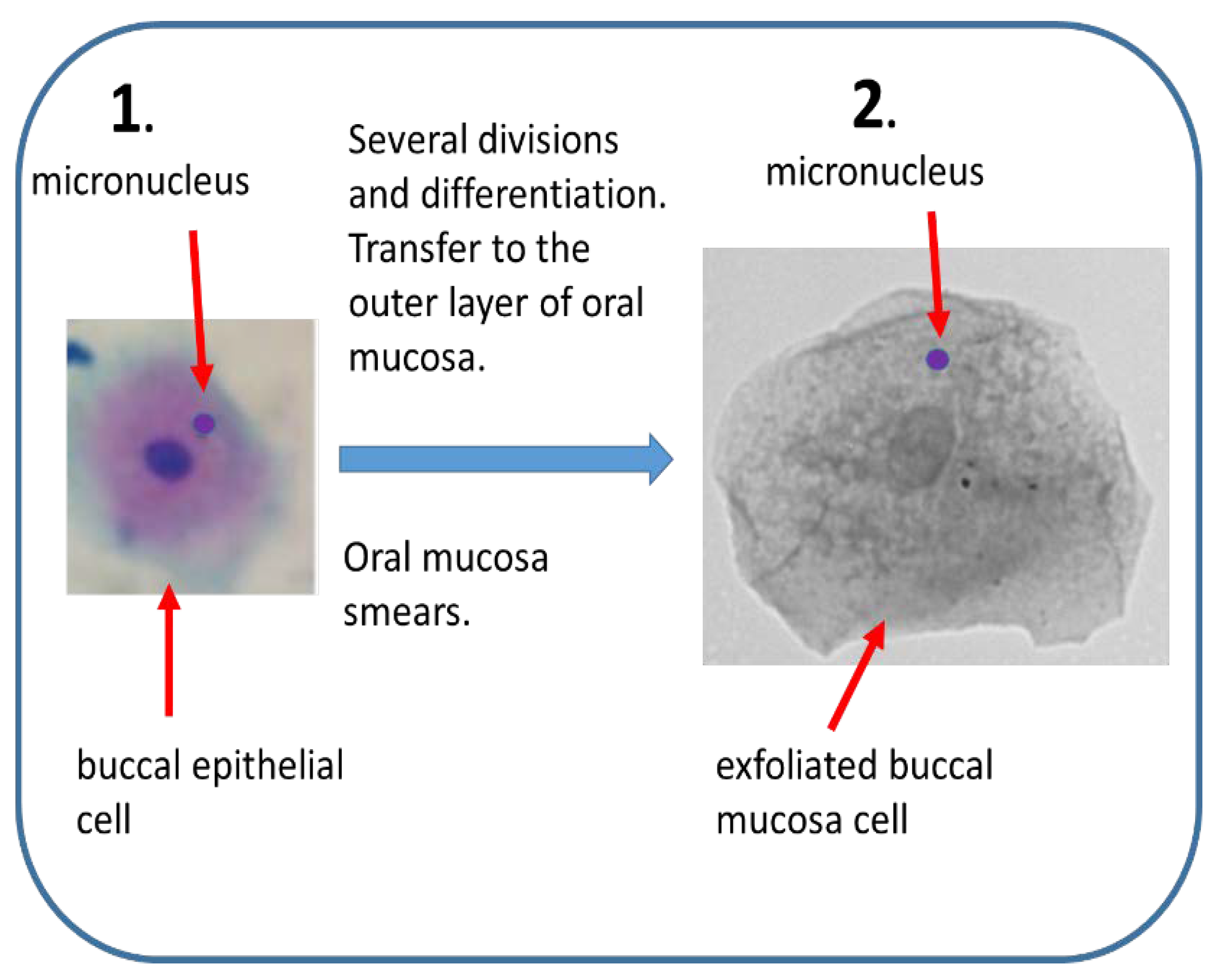
Figure 3.
Buccal micronucleus assay. (1) Mn are induced in vivo by genotoxic agents in rapidly dividing buccal epithelial tissue. Epithelial cells differentiate and move towards outer layer of oral mucosa. (2) Mn frequency can be estimated in smears of exfoliated buccal cells.
Figure 3.
Buccal micronucleus assay. (1) Mn are induced in vivo by genotoxic agents in rapidly dividing buccal epithelial tissue. Epithelial cells differentiate and move towards outer layer of oral mucosa. (2) Mn frequency can be estimated in smears of exfoliated buccal cells.

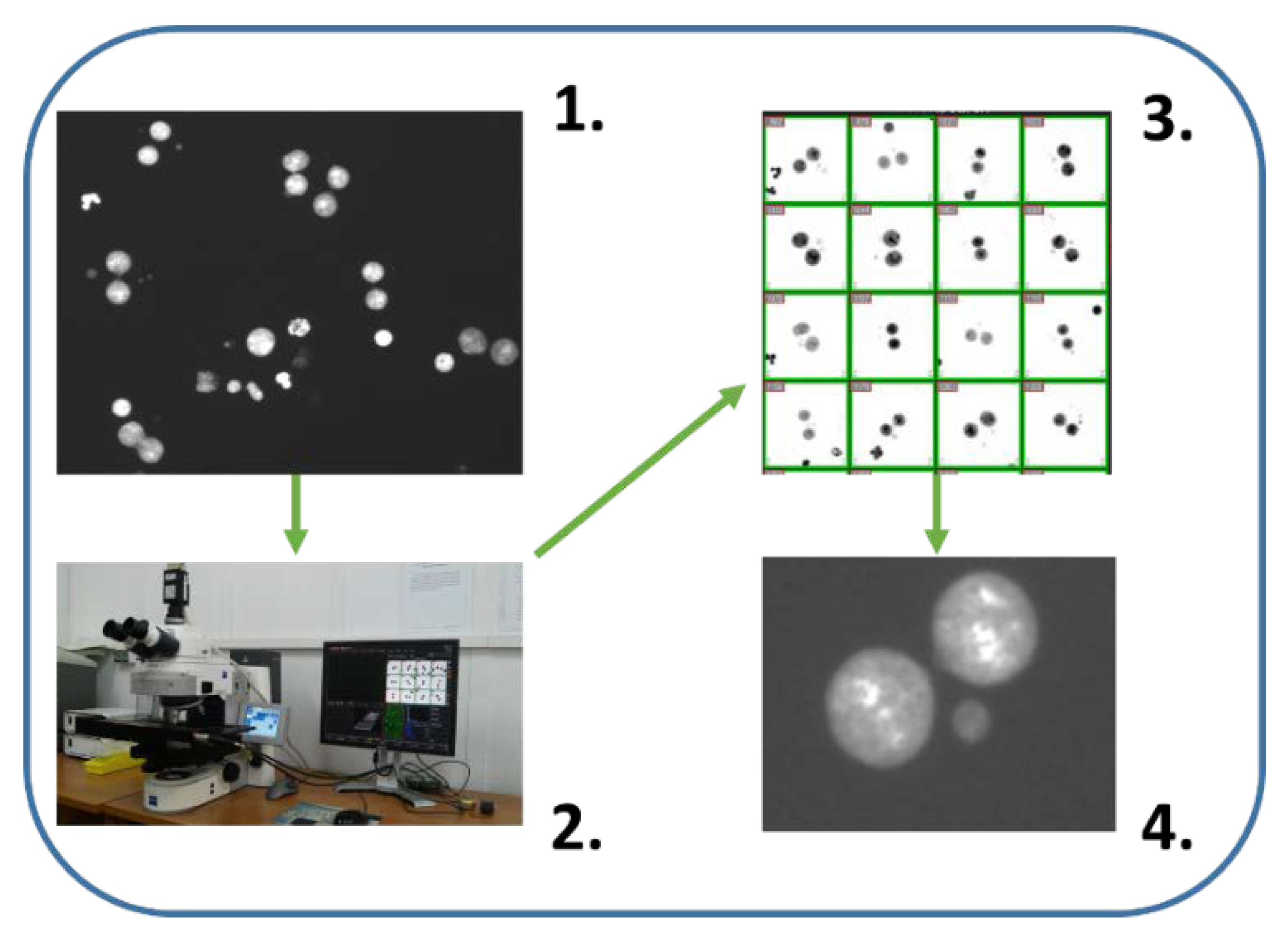
Figure 4.
Automatic/semiautomatic scoring by microscope aided systems (Metasystems). (1) Nuclei of cytokinesis-block cells are visible. System recognizes binucleated cells when both nuclei are the same size and they are in proximity to each other. (2) Motorized microscope with image analysis system safe galleries of binucleated cells and recognize Mn in an automatic manner. (3) Gallery of binucleated cells. Mathematical algorithms to recognize binucleated cells implemented in Metasystems has been used many times with good results [55,115,116]. (4) Each cell from the gallery can be localized and e.g., false positive Mn can be verified in high magnification. Such semiautomatic scoring gives better results than automatic scoring.
Figure 4.
Automatic/semiautomatic scoring by microscope aided systems (Metasystems). (1) Nuclei of cytokinesis-block cells are visible. System recognizes binucleated cells when both nuclei are the same size and they are in proximity to each other. (2) Motorized microscope with image analysis system safe galleries of binucleated cells and recognize Mn in an automatic manner. (3) Gallery of binucleated cells. Mathematical algorithms to recognize binucleated cells implemented in Metasystems has been used many times with good results [55,115,116]. (4) Each cell from the gallery can be localized and e.g., false positive Mn can be verified in high magnification. Such semiautomatic scoring gives better results than automatic scoring.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Examples of the applications of cytokinesis-block micronucleus assay to detect and study genetic and civilization diseases.
Table 1.
Examples of the applications of cytokinesis-block micronucleus assay to detect and study genetic and civilization diseases.
| Diagnose genetic disorders like Fanconi anemia, ataxia telangiectasia or various autoimmune diseases [24,25,26]. |
| Evaluation of individual susceptibility to the effects of exogenous or endogenous genotoxic agents [3,12]. |
| Assessment of the risk of developing cancer and other chronic diseases [12]. |
| Prediction assay for a radiation side effect (normal tissue reaction) of patients with different cancers subjected to RT and post RT follow up [27,28,29]. |
| Quantification of in vitro genotoxicity of different schemes of RT [29]. |
Table 2.
Possible origin of micronuclei.
| Acentric chromosome fragments |
| Acentric chromosome fragments result from unrepaired DNA strand breaks or misrepaired DNA strand breaks, DNA strand cross-links or adducts leading to chromosome or chromatid type aberrations, e.g., polycentric chromosomes usually accompanied with acentric fragments; [22,31,32,33]. |
| Malsegregation of chromosomes |
| A whole chromosome lagging behind during mitosis or numerical chromosome aberrations, as a result of centromere dysfunction; kinetochore dysfunction, spindle dysfunction. Aging of women, when some chromosomes X are excluded from the nucleus [22,31,32,33]. |
| Dicentric chromosome breakage |
| Polycentric chromosomes, when spread between opposite cells, may break in many pieces, giving rise to Mn or/and broken unprotected chromatid ends [31,32,33,34]. Unprotected chromatid ends are susceptible to different reorganization processes and can start breakage–fusion–bridge cycles [34], that give rise to Mn formation and are considered as one of the mechanisms of chromosome instability of cancer cells [34]. |
| Chromosome instability |
| Chromosome instability is a condition in which cells gain changes in their genome at a high rate. Chromosome instability is often displayed by pre-neoplastic and cancerous cells, which usually show a high frequency of Mn [31,35]. It is accepted that Mn is a good indicator of chromosome instability [31]. |
| Aggregation of double minutes (DB) |
| DB are small acentric and telomere-free extrachromosomal bodies composed of circular DNA [31,36,37]. They have been observed in many kinds of tumors including breast, lung, ovary, colon and neuroblastoma [36,37,38]. DB are the manifestation of genomic instability and recently they have been linked to chromothripsis phenomenon [39]. They carry multiple copies of amplified genes, usually oncogenes or genes involved in drug resistance. Many copies of DB can be found in a single cell often stuck to the chromosomes [40]. DB, when detached from chromosomes, can aggregate and form Mn [41]. |
Table 3.
Types of micronucleus assays.
| Type of Test | Cells Used | Purpose | Short Characteristics |
|---|---|---|---|
| Cytokinesis-block micro-nucleus assay (CBMN) | human, rodents, rabbit, fish, dogs, primates, etc. lymphocytes or cell lines. |
| Mn is scored in binucleated cells, where cytokinesis is stopped by addition of cytochalasin B. The most popular in vitro and in vivo MN. |
| Mammalian Erythrocyte MN | human, rodents, rabbit, fish, dogs, primates, immature erythrocytes. |
| Test performed usually on young rodents, but biomonitoring of the human population based on peripheral blood is possible. When performed in peripheral blood erythrocytes splenic selection must be considered. |
| Buccal MN | human epithelial buccal cells. |
| Incoming, little invasive in vivo test. Suitable for biomonitoring. |
| MN in other cell types | nasal mucosa cells, urine-derived cells. |
| Not very popular, although there are new publications. |
Table 4.
Application of CBMN.
| basic research on DNA damage and repair [58,59]; |
| radiosensitivity studies of various groups, whether healthy or with genetic disorders [24,60,61,62]; |
| attempts to link the radiosensitivity with the radiation reaction of normal tissues in persons undergoing radiotherapy [63,64]; |
| predictive tests of neoplastic disease [60,65,66]; |
| characterizations of cytogenetic damage during chemo- and radiotherapy [67,68]; |
| biomonitoring of the environment or occupational exposure [32,69,70,71]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sommer, S.; Buraczewska, I.; Kruszewski, M. Micronucleus Assay: The State of Art, and Future Directions. Int. J. Mol. Sci. 2020, 21, 1534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041534
AMA Style
Sommer S, Buraczewska I, Kruszewski M. Micronucleus Assay: The State of Art, and Future Directions. International Journal of Molecular Sciences. 2020; 21(4):1534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041534
Chicago/Turabian StyleSommer, Sylwester, Iwona Buraczewska, and Marcin Kruszewski. 2020. "Micronucleus Assay: The State of Art, and Future Directions" International Journal of Molecular Sciences 21, no. 4: 1534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041534
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.