Sex and Gender Aspects for Patient Stratification in Allergy Prevention and Treatment

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
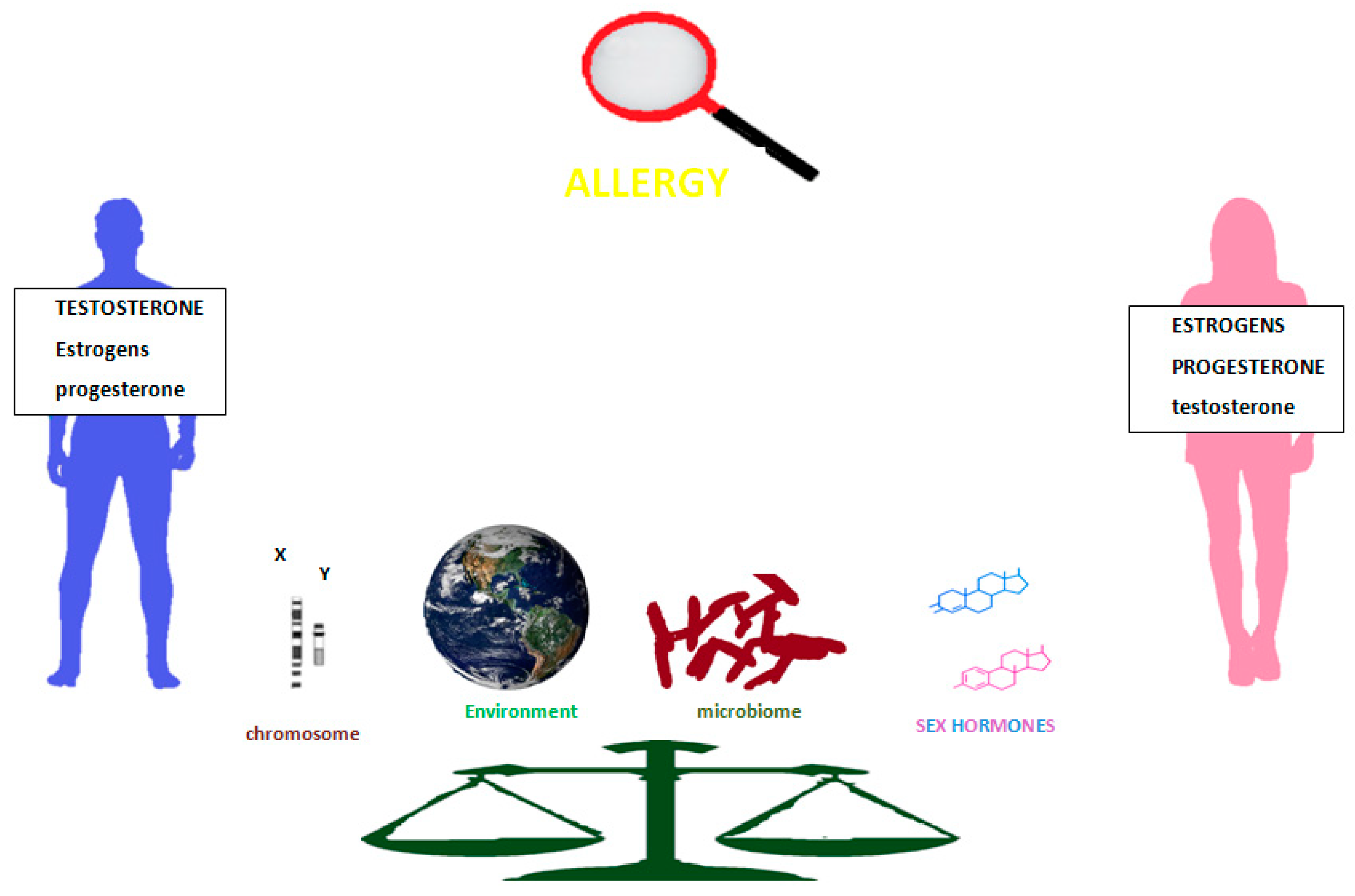
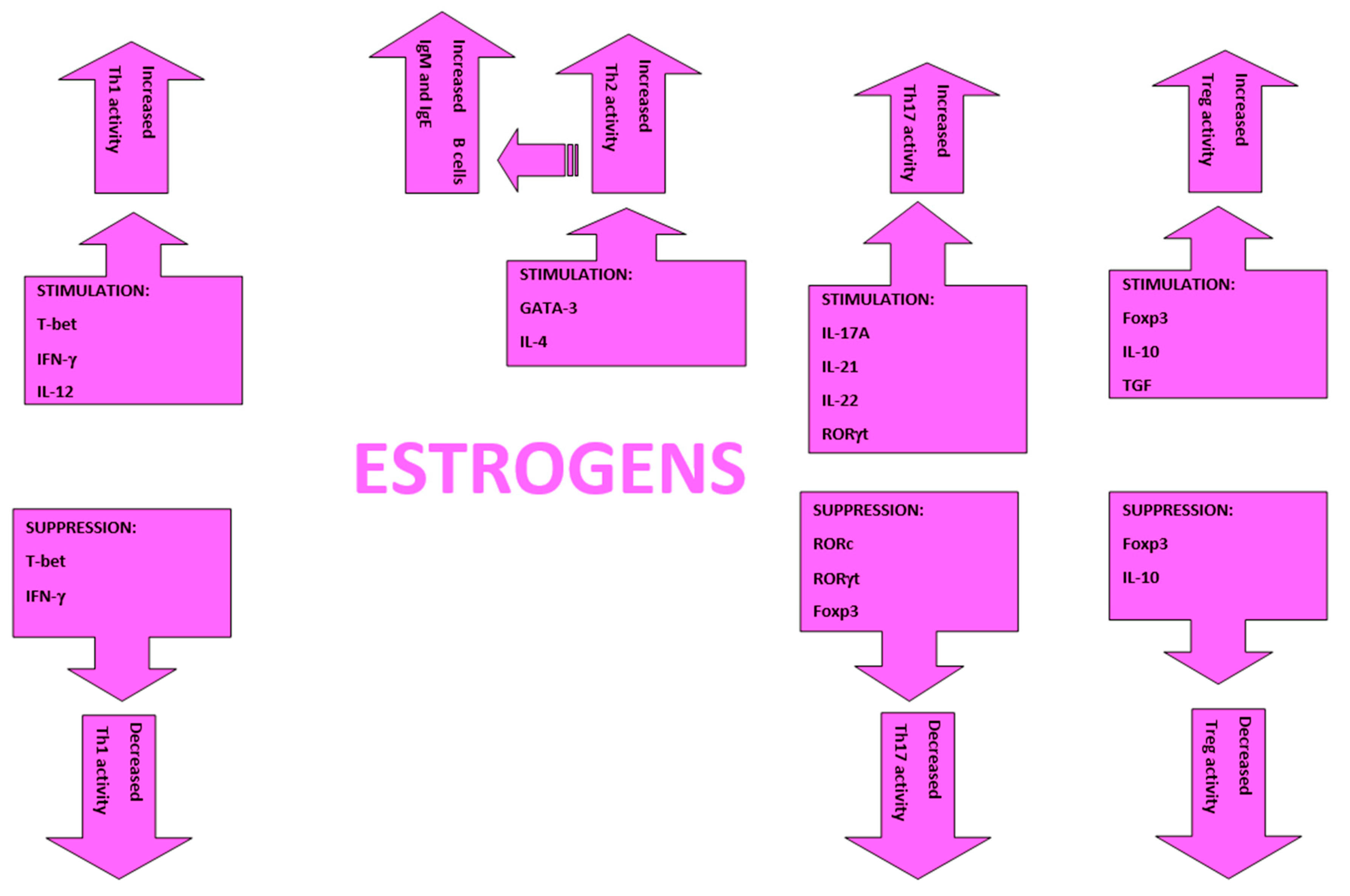
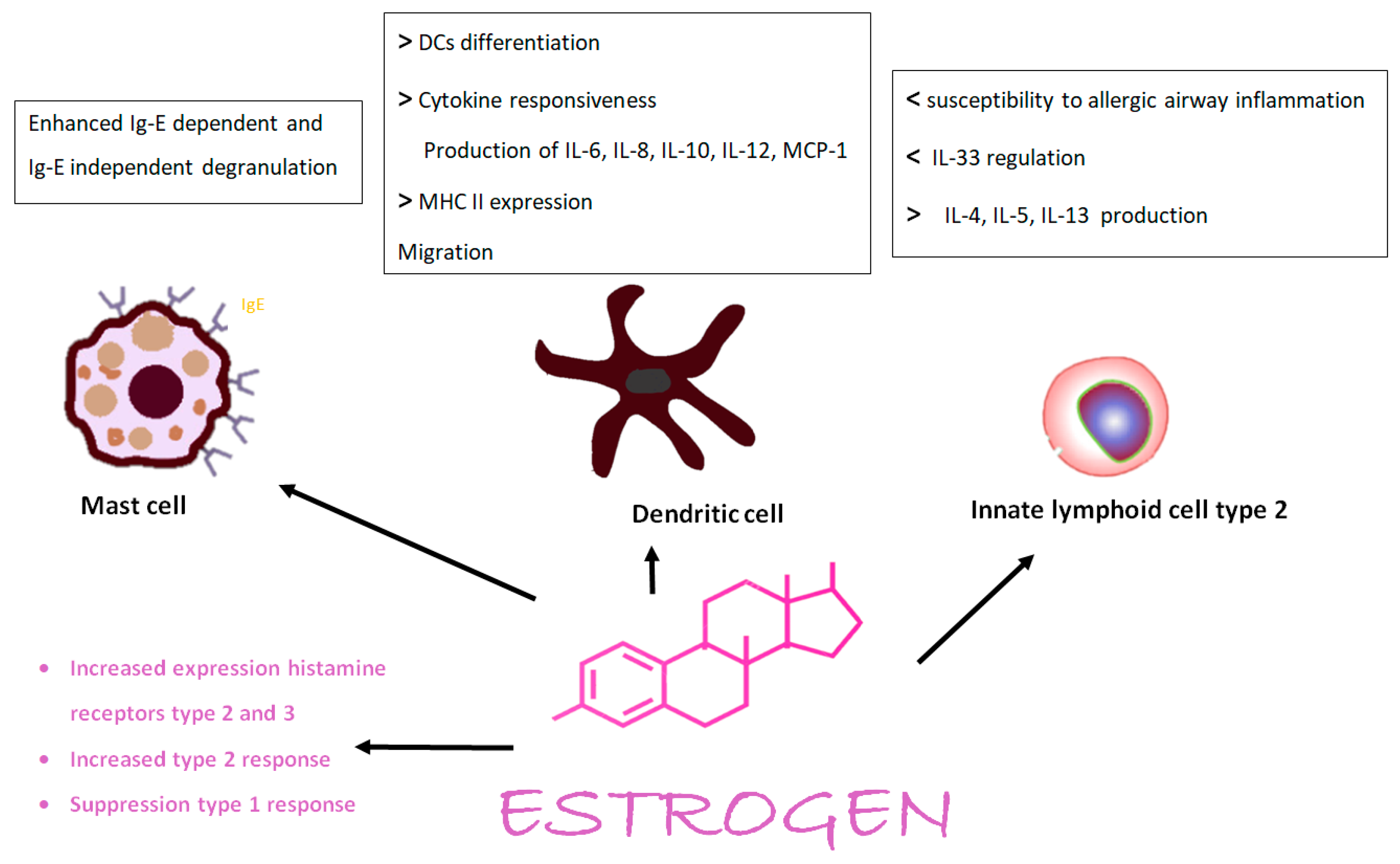
2. Hormones, Chromosomes, Immunity, and Allergy
3. Asthma and Rhinitis
4. Food Allergy
5. Atopic Dermatitis
6. Urticaria
7. Anaphylaxis
8. Drug Allergy
9. Vaccines
10. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Springer, K.W.; Hankivsky, O.; Bates, L.M. Gender and health: Relational, intersectional, and biosocial approaches. Soc. Sci. Med. 2012, 74, 1661–1666. [Google Scholar] [CrossRef]
- Marino, M.; Masella, R.; Bulzomi, P.; Campesi, I.; Malorni, W.; Franconi, F. Nutrition and human health from a sex-gender perspective. Mol. Aspects Med. 2011, 32, 1–70. [Google Scholar] [CrossRef]
- Franconi, F.; Campesi, I.; Colombo, D.; Antonini, P. Sex-Gender Variable: Methodological Recommendations for Increasing Scientific Value of Clinical Studies. Cells 2019, 8, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leffler, J.; Stumbles, P.A.; Strickland, D.H. Immunological Processes Driving IgE Sensitisation and Disease Development in Males and Females. Int. J. Mol. Sci. 2018, 19, 1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agache, I.; Akdis, C.A. Precision medicine and phenotypes, endotypes, genotypes, regiotypes, and theratypes of allergic diseases. J. Clin. Investig. 2019, 130, 1493–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen-Jarolim, E.; Untersmayr, E. Gender-medicine aspects in allergology. Allergy 2008, 63, 610–615. [Google Scholar] [CrossRef] [Green Version]
- Nowak-Wegrzyn, A.; Ellis, A.; Castells, M. Sex and allergicdiseases. Ann. Allergy Asthma Immunol. 2019, 122, 134–135. [Google Scholar] [CrossRef] [PubMed]
- Ciccarelli, F.; De Martinis, M.; Ginaldi, L. Glucocorticoids in Patients With Rheumatic Diseases: Friends Or Enemies of Bone? Curr. Med. Chem. 2015, 22, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Bereshchenko, O.; Bruscoli, S.; Riccardi, C. Glucocorticoids, Sex Hormones, and Immunity. Front. Immunol. 2018, 9, 1332. [Google Scholar] [CrossRef]
- Kim, D.H.; Park, H.J.; Park, H.S.; Lee, J.U.; Ko, C.; Gye, M.C.; Choi, J.M. Estrogen receptor α in T cells suppresses follicular helper T cell responses and prevents autoimmunity. Exp. Mol. Med. 2019, 51, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taneja, V. Sex Hormones Determine Immune Response. Front. Immunol. 2018, 9, 1931. [Google Scholar] [CrossRef] [PubMed]
- Fuseini, H.; Cephus, J.Y.; Wu, P.; Davis, J.B.; Contreras, D.C.; Gandhi, V.D.; Rathmell, J.C.; Newcomb, D.C. ERα Signaling Increased IL-17A Production in Th17 Cells by Upregulating IL-23R Expression, Mitochondrial Respiration, and Proliferation. Front. Immunol. 2019, 10, 2740. [Google Scholar] [CrossRef] [Green Version]
- Asaba, J.; Bandyopadhyay, M.; Kindy, M.; Dasgupta, S. Estrogen receptor signal in regulation of B cell activation during diverse immune responses. Int. J. Biochem. Cell Biol. 2015, 68, 42–47. [Google Scholar] [CrossRef]
- Mo, R.; Chen, J.; Grolleau-Julius, A.; Murphy, H.S.; Richardson, B.C.; Yung, R.L. Estrogen regulates CCR gene expression and function in T lymphocytes. J. Immunol. 2005, 174, 6023–6029. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, T.N. Sex disparities in the immune response. Cell Immunol. 2015, 294, 61–62. [Google Scholar] [CrossRef]
- Lotter, H.; Altfeld, M. Sex differences in immunity. Semin. Immunopathol. 2019, 41, 133–135. [Google Scholar] [CrossRef] [Green Version]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Allergy and aging: An old/new emerging health issue. Aging Dis. 2017, 8, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Afify, S.M.; Pali-Schöll, I. Adverse reactions to food: The female dominance—A secondary publication and update. World Allergy Organ. J. 2017, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, A.; Luckey, D.; Taneja, V. The gut microbiome in autoimmunity: Sex matters. Clin. Immunol. 2015, 159, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Wallis, A.; Butt, H.; Ball, M.; Lewis, D.P.; Bruck, D. Support for the microgenderome invites enquiry into sex differences. Gut Microbes 2017, 8, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.; Denison, M.S.; Tachibana, H.; Yamada, K. Effects of estrogenic compounds on immunoglobulin production by mouse splenocytes. Biol. Pharm. Bull. 2002, 25, 1263–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridolo, E.; Incorvaia, C.; Martignago, I.; Caminati, M.; Canonica, G.W.; Senna, G. Sex in Respiratory and Skin Allergies. Clin. Rev. Allergy Immunol. 2019, 56, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Ginaldi, L.; De Martinis, M.; Saitta, S.; Sirufo, M.M.; Mannucci, C.; Casciaro, M.; Ciccarelli, F.; Gangemi, S. Interleukin-33 serum levels in postmenopausal women with osteoporosis. Sci. Rep. 2019, 9, 3786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace, S.; Sautebin, L.; Werz, O. Sex-biased eicosanoid biology: Impact for sex differences in inflammation and consequences for pharmacotherapy. Biochem. Pharmacol. 2017, 145, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Marriott, I.; Bost, K.L.; Huet-Hudson, Y.M. Sexual dimorphism in expression of receptors for bacterial lipopolysaccharides in murine macrophages: A possible mechanism for gender-based differences in endotoxic shock susceptibility. J. Reprod. Immunol. 2006, 71, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.J.; Dragon, J.A.; Moussawi, M.; Huber, S.A. Sex-specific signaling through Toll-Like Receptors 2 and 4 contributes to survival outcome of Coxsackievirus B3 infection in C57Bl/6 mice. Biol. Sex Differ. 2012, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Torcia, M.G.; Nencioni, L.; Clemente, A.M.; Civitelli, L.; Celestino, I.; Limongi, D.; Fadigati, G.; Perissi, E.; Cozzolino, F.; Garaci, E.; et al. Sex differences in the response to viral infections: TLR8 and TLR9 ligand stimulation induce higher IL10 production in males. PLoS ONE 2012, 7, e39853. [Google Scholar] [CrossRef]
- Smith, L.J. Leukotrienes and sex: Strange bedfellows? J. Clin. Investig. 2017, 127, 2895–2896. [Google Scholar] [CrossRef] [Green Version]
- Massimini, M.; Palmieri, C.; De Maria, R.; Romanucci, M.; Malatesta, D.; De Martinis, M.; Maniscalco, L.; Ciccarelli, A.; Ginaldi, L.; Buracco, P.; et al. 17-AAG and Apoptosis, Autophagy and Mitophagy in Canine Osteosarcoma Cell Lines. Vet. Pathol. 2017, 54, 405–412. [Google Scholar] [CrossRef]
- Pace, S.; Pergola, C.; Dehm, F.; Rossi, A.; Gerstmeier, J.; Troisi, F.; Pein, H.; Schaible, A.M.; Weinigel, C.; Rummler, S.; et al. Androgen-mediated sex bias impairs efficiency of leukotriene biosynthesis inhibitors in males. J. Clin. Investig. 2017, 127, 3167–3176. [Google Scholar] [CrossRef] [Green Version]
- Quintero, O.L.; Amador-Patarroyo, M.J.; Montoya-Ortiz, G.; Rojas-Villarraga, A.; Anaya, J.M. Autoimmune disease and gender: Plausible mechanisms for the female predominance of autoimmunity. J. Autoimmun. 2012, 38, J109–J119. [Google Scholar] [CrossRef] [PubMed]
- Abramowitz, L.K.; Olivier-Van Stichelen, S.; Hanover, J.A. Chromosome imbalance as a driver of sex disparity in disease. J. Genom. 2014, 2, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, R.; McReynolds, S.; Leroith, T.; Heid, B.; Liang, Z.; Ahmed, S.A. Sex differences in the expression of lupus-associated miRNAs in splenocytes from lupus-prone NZB/WF1 mice. Biol. Sex Differ. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azcoitia, I.; Barreto, G.E.; Garcia-Segura, L.M. Molecular mechanisms and cellular events involved in the neuroprotective actions of estradiol. Analysis of sex differences. Front. Neuroendocrinol. 2019, 55, 100787. [Google Scholar] [CrossRef]
- Irelli, A.; Sirufo, M.M.; Scipioni, T.; De Pietro, F.; Pancotti, A.; Ginaldi, L.; De Martinis, M. mTOR links tumor immunity and bone metabolism: What clinical implications? Int. J. Mol. Sci. 2019, 20, 5841. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, K.; Do, D.C.; Zhang, Y.; Hu, X.; Chen, J.; Gao, P. Environmental Exposures and Asthma Development: Autophagy, Mitophagy, and Cellular Senescence. Front. Immunol. 2019, 10, 2787. [Google Scholar] [CrossRef]
- Xia, F.; Deng, C.; Jiang, Y.; Qu, Y.; Deng, J.; Cai, Z.; Ding, Y.; Guo, Z.; Wang, J. IL4 (interleukin 4) induces autophagy in B cells leading to exacerbated asthma. Autophagy 2018, 14, 450–464. [Google Scholar] [CrossRef]
- Yung, J.A.; Fuseini, H.; Newcomb, D.C. Hormones, sex, and asthma. Ann. Allergy Asthma Immunol. 2018, 120, 488–494. [Google Scholar] [CrossRef] [Green Version]
- Stark, J.M.; Tibbitt, C.A.; Coquet, J.M. The Metabolic Requirements of Th2 Cell Differentiation. Front. Immunol. 2019, 10, 2318. [Google Scholar] [CrossRef]
- Rostamzadeh, D.; Yousefi, M.; Haghshenas, M.R.; Ahmadi, M.; Dolati, S.; Babaloo, Z. mTOR Signaling pathway as a master regulator of memory CD8+ T-cells, Th17, and NK cells development and their functional properties. J. Cell Physiol. 2019, 234, 12353–12368. [Google Scholar] [CrossRef]
- Conciatori, F.; Bazzichetto, C.; Falcone, I.; Pilotto, S.; Bria, E.; Cognetti, F.; Milella, M.; Ciuffreda, L. Role of mTOR Signaling in Tumor Microenvironment: An Overview. Int. J. Mol. Sci. 2018, 19, 2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, S.; Kurebayashi, Y.; Koyasu, S. Role of PI3K/Akt and mTOR complexes in Th17 cell differentiation. Ann. N. Y. Acad Sci. 2013, 1280, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Malik, N.; Sansom, O.J.; Michie, A.M. The role of mTOR-mediated signals during haemopoiesis and lineage commitment. Biochem. Soc. Trans. 2018, 46, 1313–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, N.; Dunn, K.M.; Cassels, J.; Hay, J.; Estell, C.; Sansom, O.J.; Michie, A.M. mTORC1 activity is essential for erythropoiesis and B cell lineage commitment. Sci. Rep. 2019, 9, 16917. [Google Scholar] [CrossRef]
- Powell, J.D.; Pollizzi, K.N.; Heikamp, E.B.; Horton, M.R. Regulation of immune responses by mTOR. Annu. Rev. Immunol. 2012, 30, 39–68. [Google Scholar] [CrossRef] [Green Version]
- Chiu, H.; Jackson, L.V.; Oh, K.I.; Mai, A.; Ronai, Z.A.; Ruggero, D.; Fruman, D.A. The mTORC1/4E-BP/eIF4E Axis Promotes Antibody Class Switching in B Lymphocytes. J. Immunol. 2019, 202, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Zein, J.G.; Denson, J.L.; Wechsler, M.E. Asthma over the Adult Life Course: Gender and Hormonal Influences. Clin. Chest Med. 2019, 40, 149–161. [Google Scholar] [CrossRef]
- Koper, I.; Hufnagl, K.; Ehmann, R. Gender aspects and influence of hormones on bronchial asthma—Secondary publication and update. World Allergy Organ. J. 2017, 10, 46. [Google Scholar] [CrossRef]
- Hohmann, C.; Keller, T.; Gehring, U.; Wijga, A.; Standl, M.; Kull, I.; Bergstrom, A.; Lehmann, I.; von Berg, A.; Heinrich, J.; et al. Sex-specific incidence of asthma, rhinitis and respiratory multimorbidity before and after puberty onset: Individual participant meta-analysis of five birth cohorts collaborating in MeDALL. BMJ Open Respir. Res. 2019, 6, e000460. [Google Scholar] [CrossRef]
- Keller, T.; Hohmann, C.; Standl, M.; Wijga, A.H.; Gehring, U.; Melén, E.; Almqvist, C.; Lau, S.; Eller, E.; Wahn, U.; et al. The sex-shift in single disease and multimorbid asthma and rhinitis during puberty—A study by MeDALL. Allergy 2018, 73, 602–614. [Google Scholar] [CrossRef] [Green Version]
- Fröhlich, M.; Pinart, M.; Keller, T.; Reich, A.; Cabieses, B.; Hohmann, C.; Postma, D.S.; Bousquet, J.; Antó, J.M.; Keil, T.; et al. Is there a sex-shift in prevalence of allergic rhinitis and comorbid asthma from childhood to adulthood? A meta-analysis. Clin. Transl. Allergy 2017, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Pignataro, F.S.; Bonini, M.; Forgione, A.; Melandri, S.; Usmani, O.S. Asthma and gender: The female lung. Pharmacol. Res. 2017, 119, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zein, J. Novel Insights on Sex-Related Differences in Asthma. Curr. Allergy Asthma Rep. 2019, 19, 44. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Zheng, L.D.; Smith, C.; Luo, J.; Robinson, A.; Almeida, F.A.; Wang, Z.; Olumi, A.F.; Liu, D.; Cheng, Z. Estradiol signaling mediates gender difference in visceral adiposity via autophagy. Cell Death Dis. 2018, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Zein, J.G.; Yaqoob, Z.; Al-Kindi, S.G.; DeBoer, M.; Teague, W.G.; Gaston, B.M.; Erzurum, S.C. Polycystic ovary syndrome is associated with a higher risk for asthma. Am. J. Respir. Crit. Care Med. 2017, 195, A1307. [Google Scholar]
- E-Lacerda, R.R.; Anhê, G.F.; Page, C.P.; Riffo-Vasquez, Y. Sex differences in the influence of obesity on a murine model of allergic lung inflammation. Clin. Exp. Allergy 2019, 50, 256–266. [Google Scholar] [CrossRef]
- Laffont, S.; Seillet, C.; Guéry, J.C. Estrogen Receptor-Dependent Regulation of Dendritic Cell Development and Function. Front. Immunol. 2017, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. IL-33/IL-31 axis in osteoporosis. Int. J. Mol. Sci. 2020, 21, 1239. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.; Newcomb, D.C. Sex Bias in Asthma Prevalence and Pathogenesis. Front. Immunol. 2018, 9, 2997. [Google Scholar] [CrossRef] [Green Version]
- Fuseini, H.; Newcomb, D.C. Mechanisms Driving Gender Differences in Asthma. Curr. Allergy Asthma Rep. 2017, 17, 19. [Google Scholar] [CrossRef] [Green Version]
- Takeda, M.; Tanabe, M.; Ito, W.; Ueki, S.; Konnno, Y.; Chihara, M.; Itoga, M.; Kobayashi, Y.; Moritoki, Y.; Kayaba, H.; et al. Gender difference in allergic airway remodelling and immunoglobulin production in mouse model of asthma. Respirology 2013, 18, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Salomonsson, M.; Malinovschi, A.; Kalm-Stephens, P.; Dahlin, J.S.; Janson, C.; Alving, K.; Hallgren, J. Circulating mast cell progenitors correlate with reduced lung function in allergic asthma. Clin. Exp. Allergy 2019, 49, 874–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jing, Y.; Qiao, J.; Luan, B.; Wang, X.; Wang, L.; Song, Z. Activation of the mTOR signaling pathway is required for asthma onset. Sci. Rep. 2017, 7, 4532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Xia, L.; Li, F.; Zhou, L.; Weng, Q.; Li, Z.; Wu, Y.; Mao, Y.; Zhang, C.; Wu, Y.; et al. mTOR complexes differentially orchestrates eosinophil development in allergy. Sci. Rep. 2018, 8, 6883. [Google Scholar] [CrossRef]
- Shin, S.; Lee, J.H.; Lee, H.J.; Chang, S.Y.; Chung, S.H. Rapamycin attenuates Th2-driven experimental allergic conjunctivitis. Clin. Immunol. 2018, 190, 1–10. [Google Scholar] [CrossRef]
- Fang, L.; Wang, X.; Sun, Q.; Papakonstantinou, E.; S’ng, C.; Tamm, M.; Stolz, D.; Roth, M. IgE Downregulates PTEN through MicroRNA-21–5p and Stimulates Airway Smooth Muscle Cell Remodeling. Int. J. Mol. Sci. 2019, 20, 875. [Google Scholar] [CrossRef] [Green Version]
- Rakhmanova, V.; Jin, M.; Shin, J. Inhibition of Mast Cell Function and Proliferation by mTOR Activator MHY1485. Immune Netw. 2018, 18, e18. [Google Scholar] [CrossRef]
- Pali-Scholl, I.; Jensen-Jarolim, E. Gender aspects in food allergy. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 249–255. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Food allergy insights: A changing landscape. Arch. Immunol. Ther. Exp. AITE 2020, in press. [Google Scholar]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Food allergies and ageing. Int. J. Mol. Sci. 2019, 20, 5580. [Google Scholar] [CrossRef] [Green Version]
- Deschildre, A.; Lejeune, S.; Cap, M.; Flammarion, S.; Jouannic, L.; Amat, F.; Just, J. Food allergy phenotypes: The key to personalized therapy. Clin. Exp. Allergy 2017, 47, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. New perspectives in food allergy. Int. J. Mol. Sci. 2020, 21, 1474. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.L.; Allen, K.J.; Dharmage, S.C.; Lodge, C.J.; Koplin, J.J.; Ponsonby, A.L.; Wake, M.; Lowe, A.J.; Tang, M.L.K.; Matheson, M.C.; et al. Health Nuts study. Differential factors associated with challenge-proven food allergy phenotypes in a population cohort of infants: A latent class analysis. Clin. Exp. Allergy 2015, 45, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Just, J.; Elegbede, C.F.; Deschildre, A.; Bousquet, J.; Moneret-Vautrin, D.A.; Crepet, A.; Mirabel Study Group. Three peanut-allergic/sensitized phenotypes with gender difference. Clin. Exp. Allergy 2016, 46, 1596–1604. [Google Scholar] [CrossRef]
- Acker, W.W.; Plasek, J.M.; Blumenthal, K.G.; Lai, K.H.; Topaz, M.; Seger, D.L.; Goss, F.R.; Slight, S.P.; Bates, D.W.; Zhou, L. Prevalence of food allergies and intolerances documented in electronic health records. J. Allergy Clin. Immunol. 2017, 140, 1587–1591.e1. [Google Scholar] [CrossRef] [Green Version]
- Sirufo, M.M.; De Martinis, M.; Ginaldi, L. Omalizumab an effective and safe alternative therapy in severe refractory atopic dermatitis: A case report. Medicine 2018, 97, e10897. [Google Scholar] [CrossRef]
- Kanda, N.; Hoashi, T.; Saeki, H. The Roles of Sex Hormones in the Course of Atopic Dermatitis. Int. J. Mol. Sci. 2019, 20, 4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuramoto, T.; Yokoe, M.; Tanaka, D.; Yuri, A.; Nishitani, A.; Higuchi, Y.; Yoshimi, K.; Tanaka, M.; Kuwamura, M.; Hiai, H.; et al. Atopic dermatitis-like skin lesions with IgE hyperproduction and pruritus in KFRS4/Kyo rats. J. Dermatol. Sci. 2015, 80, 116–123. [Google Scholar] [CrossRef] [Green Version]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. A “Stadium” Urticaria, Cold Urticaria Is Still a Mostly Unknown Disease, with a Wide Spectrum of Severity Degrees and Few Therapeutic Certainties: Is Omalizumab One of These? Reflections from a Clinical Case Report. Iran. Red Cresc. Med. J. 2019, 21, e84250. [Google Scholar] [CrossRef]
- Sirufo, M.M.; Ginaldi, L.; De Martinis, M. Asthma, urticaria and omalizumab in children: Reflections from a clinical case report. Front. Pediatr. 2019, 7, 213. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Ginaldi, L. Solar urticaria, a disease with many dark sides: Is omalizumab the right therapeutic response? Reflections from a clinical case report. Open Med. Wars 2019, 14, 403–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassano, N.; Colombo, D.; Bellia, G.; Zagni, E.; Vena, G.A. Gender-related differences in chronic urticaria. G. Ital. Dermatol. Venereol. 2016, 151, 544–552. [Google Scholar] [PubMed]
- Hox, V.; Desai, A.; Bandara, G.; Gilfillan, A.M.; Metcalfe, D.D.; Olivera, A. Estrogen increases the severity of anaphylaxis in female mice through enhanced endothelial nitric oxide synthase expression and nitric oxide production. J. Allergy Clin. Immunol. 2015, 135, 729–736.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hox, V.; O’Connell, M.P.; Lyons, J.J.; Sackstein, P.; Dimaggio, T.; Jones, N.; Nelson, C.; Boehm, M.; Holland, S.M.; Freeman, A.F.; et al. Diminution of signal transducer and activator of transcription 3 signaling inhibits vascular permeability and anaphylaxis. J. Allergy Clin. Immunol. 2016, 138, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, J.A. Progestogen Sensitization: A Unique Female Presentation of Anaphylaxis. Curr. Allergy Asthma Rep. 2020, 20, 4. [Google Scholar] [CrossRef]
- Lieberman, P. Catamenial anaphylaxis. J. Allergy Clin. Immunol. Pract. 2014, 2, 358–359. [Google Scholar] [CrossRef]
- Turner, P.J.; Jerschow, E.; Umasunthar, T.; Lin, R.; Campbell, D.E.; Boyle, R.J. Fatal Anaphylaxis: Mortality Rate and Risk Factors. J. Allergy Clin. Immunol. Pract. 2017, 5, 1169–1178. [Google Scholar] [CrossRef]
- Norton, A.E.; Konvinse, K.; Phillips, E.J.; Broyles, A.D. Antibiotic allergy in pediatrics. Pediatrics 2018, 141, e20172497. [Google Scholar] [CrossRef] [Green Version]
- Dhopeshwarkar, N.; Sheikh, A.; Doan, R.; Topaz, M.; Bates, D.W.; Blumenthal, K.G.; Zhou, L. Drug-Induced Anaphylaxis Documented in Electronic Health Records. J. Allergy Clin. Immunol. Pract. 2019, 7, 103–111. [Google Scholar] [CrossRef]
- Salvati, L.; Vitiello, G.; Parronchi, P. Gender differences in anaphylaxis. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 417–424. [Google Scholar] [CrossRef]
- Gabrielli, S.; Clarke, A.E.; Eisman, H.; Morris, J.; Joseph, L.; La Vieille, S.; Small, P.; Lim, R.; Enarson, P.; Zelcer, M.; et al. Disparities in rate, triggers, and management in pediatric and adult cases of suspected drug-induced anaphylaxis in Canada. Immun. Inflamm. Dis. 2018, 6, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerji, A.; Rudders, S.; Clark, S.; Wei, W.; Long, A.A.; Camargo, C.A., Jr. Retrospective study of drug-induced anaphylaxis treated in the emergency department or hospital: Patient characteristics, management, and 1-Year follow-up. J. Allergy Clin. Immunol. 2014, 2, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Blanca-López, N.; del Carmen Plaza-Serón, M.; Cornejo-García, J.A.; Perkins, J.R.; Canto, G.; Blanca, M. Drug-induced anaphylaxis. Curr. Treat. Options Allergy 2015, 2, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Macy, E.M. Current Epidemiology and Management of Radiocontrast-Associated Acute- and Delayed-Onset Hypersensitivity: A Review of the Literature. Perm. J. 2018, 22, 17–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaddy Norton, A.; Broyles, A.D. Drug allergy in children and adults: Is it the double X chromosome? Ann. Allergy Asthma Immunol. 2019, 122, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeil, M.M. Vaccine-Associated Anaphylaxis. Curr. Treat. Options Allergy 2019, 6, 297–308. [Google Scholar] [CrossRef]
- McNeil, M.M.; Weintraub, E.S.; Duffy, J.; Sukumaran, L.; Jacobsen, S.J.; Klein, N.P.; Hambidge, S.J.; Lee, G.M.; Jackson, L.A.; Irving, S.A.; et al. Risk of anaphylaxis after vaccination in children and adults. J. Allergy Clin. Immunol. 2016, 137, 868–878. [Google Scholar] [CrossRef] [Green Version]
- Griffioen, M.; Halsey, N. Gender differences in immediate hypersensitivity reactions to vaccines: A review of the literature. Public Health Nurs. 2014, 31, 206–214. [Google Scholar] [CrossRef]
- Harris, T.; Nair, J.; Fediurek, J.; Deeks, S.L. Assessment of sex-specific differences in adverse events following immunization reporting in Ontario, 2012–15. Vaccine 2017, 35, 2600–2604. [Google Scholar] [CrossRef]
- Sirufo, M.M.; Suppa, M.; Ginaldi, L.; De Martinis, M. Does Allergy Break Bones? Osteoporosis and Its Connection to Allergy. Int. J. Mol. Sci. 2020, 21, 712. [Google Scholar] [CrossRef] [Green Version]
- Muraro, A.; Lemanske, R.F., Jr.; Castells, M.; Torres, M.J.; Khan, D.; Simon, H.U.; Bindslev-Jensen, C.; Burks, W.; Poulsen, L.K.; Sampson, H.A.; et al. Precision medicine in allergic disease-food allergy, drug allergy, and anaphylaxis-PRACTALL document of the European Academy of Allergy and Clinical Immunology and the American Academy of Allergy, Asthma and Immunology. Allergy 2017, 72, 1006–1021. [Google Scholar] [CrossRef] [Green Version]
- Labella, M.; Garcia-Neuer, M.; Castells, M. Application of precision medicine to the treatment of anaphylaxis. Curr. Opin. Allergy Clin. Immunol. 2018, 18, 190–197. [Google Scholar] [CrossRef]
- Fava, G.A.; Cosci, F.; Sonino, N. Current Psychosomatic Practice. Psychother. Psychosom. 2017, 86, 13–30. [Google Scholar] [CrossRef]
- Adler, N.E.; Stead, W.W. Patients in context--EHR capture of social and behavioral determinants of health. N. Engl. J. Med. 2015, 372, 698–701. [Google Scholar] [CrossRef] [Green Version]
- Billi, A.C.; Kahlenberg, J.M.; Gudjonsson, J.E. Sex bias in autoimmunity. Curr. Opin. Rheumatol. 2019, 31, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Golubnitschaja, O.; Baban, B.; Boniolo, G.; Wang, W.; Bubnov, R.; Kapalla, M.; Krapfenbauer, K.; Mozaffari, M.S.; Costigliola, V. Medicine in the early twenty-first century: Paradigm and anticipation—EPMA position paper 2016. EPMA J. 2016, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swedin, L.; Saarne, T.; Rehnberg, M.; Glader, P.; Niedzielska, M.; Johansson, G.; Hazon, P.; Catley, M.C. Patient stratification and the unmet need in asthma. Pharmacol. Ther. 2017, 169, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Golubnitschaja, O.; Zhan, X. Chronic inflammation: Key player and biomarker-set to predict and prevent cancer development and progression based on individualized patient profiles. EPMA J. 2019, 10, 365–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golubnitschaja, O.; Flammer, J. Individualised patient profile: Clinical utility of Flammer syndrome phenotype and general lessons for predictive, preventive and personalised medicine. EPMA J. 2018, 9, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baban, B.; Golubnitschaja, O. The potential relationship between Flammer and Sjögren syndromes: The chime of dysfunction. EPMA J. 2017, 8, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, C.V.; Acosta, E.P.; Strykowski, J.M. Gender differences in human pharmacokinetics and pharmacodynamics. J. Adolesc. Health 1994, 15, 619–629. [Google Scholar] [CrossRef]
- Roberts, G.; Almqvist, C.; Boyle, R.; Crane, J.; Hogan, S.P.; Marsland, B.; Saglani, S.; Woodfolk, J.A. Developments in the field of clinical allergy in 2018 through the eyes of Clinical and Experimental Allergy, Part II. Clin. Exp. Allergy 2019, 49, 1550–1557. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Martinis, M.; Sirufo, M.M.; Suppa, M.; Di Silvestre, D.; Ginaldi, L. Sex and Gender Aspects for Patient Stratification in Allergy Prevention and Treatment. Int. J. Mol. Sci. 2020, 21, 1535. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041535
De Martinis M, Sirufo MM, Suppa M, Di Silvestre D, Ginaldi L. Sex and Gender Aspects for Patient Stratification in Allergy Prevention and Treatment. International Journal of Molecular Sciences. 2020; 21(4):1535. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041535
Chicago/Turabian StyleDe Martinis, Massimo, Maria Maddalena Sirufo, Mariano Suppa, Daniela Di Silvestre, and Lia Ginaldi. 2020. "Sex and Gender Aspects for Patient Stratification in Allergy Prevention and Treatment" International Journal of Molecular Sciences 21, no. 4: 1535. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041535