Molecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male Infertility

 , , and
, , and
Abstract
:1. Introduction
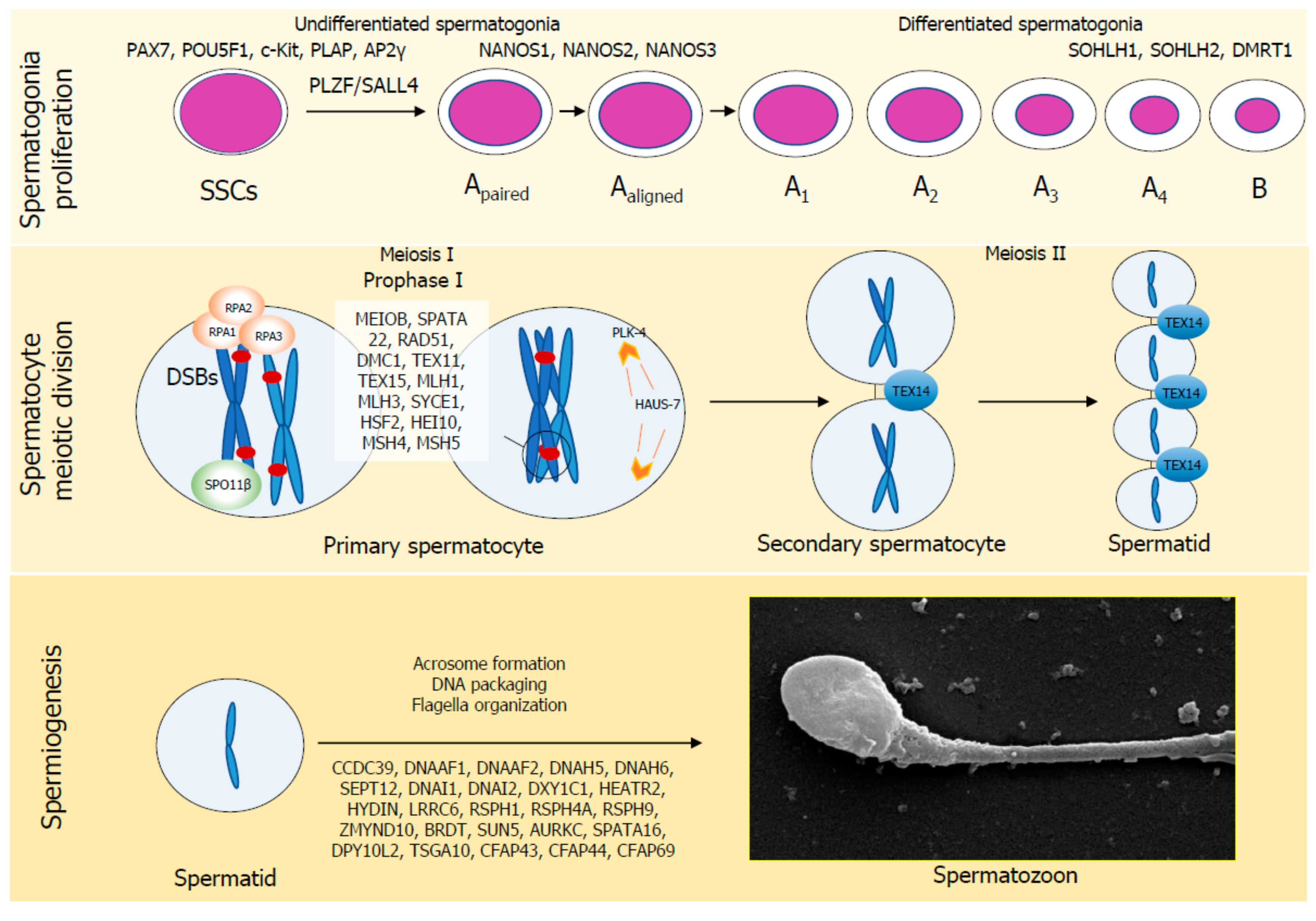
2. Physiology of Spermatogenesis
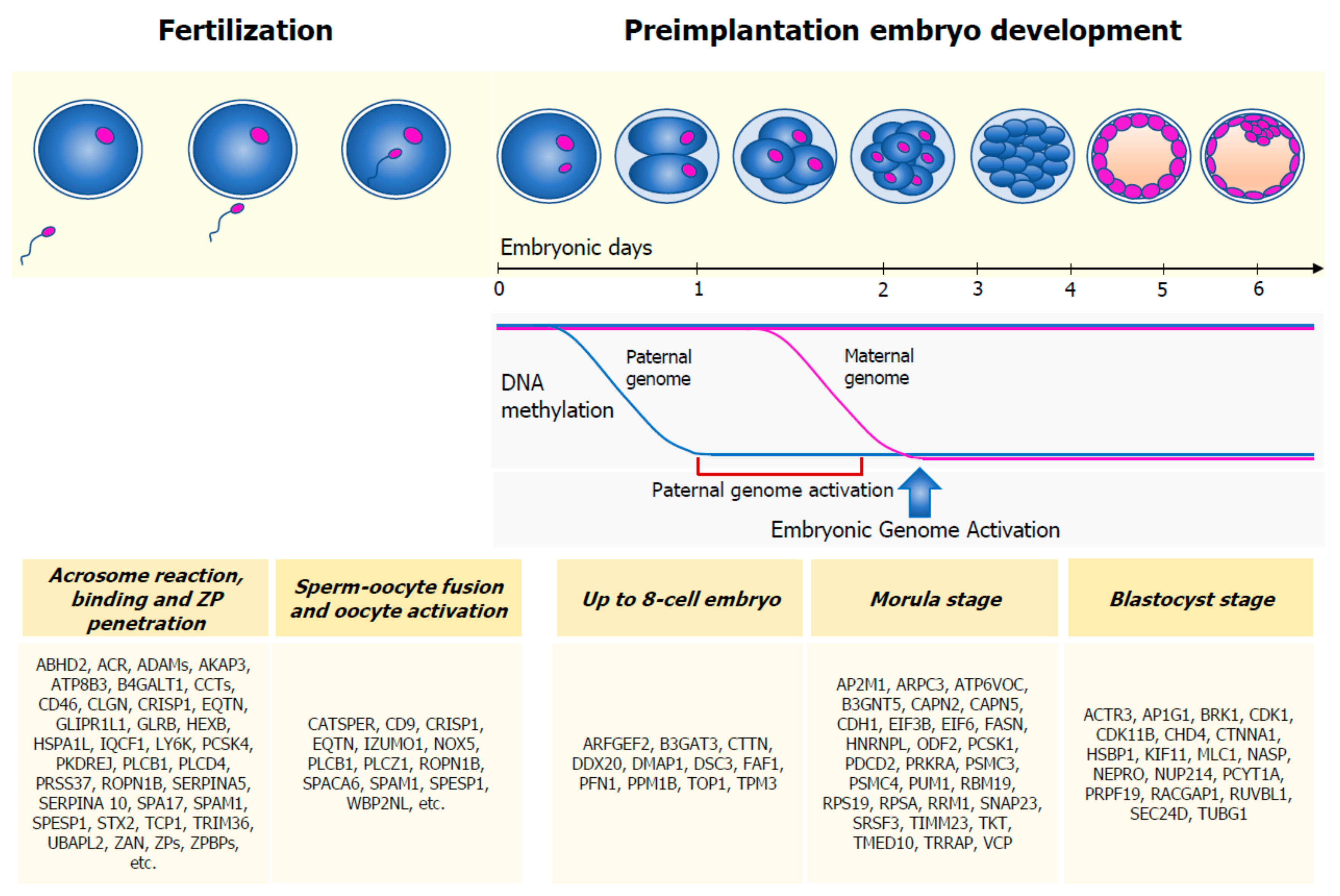
3. Physiology of Fertilization and Embryo Development
4. Sperm’s Contribution to Human Fertility
4.1. Sperm Genome
4.2. Sperm Transcriptome
4.3. Sperm Proteome
4.4. Sperm Metabolome
5. Discussion
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Report of the Meeting on the Prevention of Infertility at the Primary Health Care Levels; WHO: Geneva, Switzerland, 1983. [Google Scholar]
- ESHRE Guideline Group on RPL; Atik, R.B.; Christiansen, O.B.; Elson, J.; Kolte, A.M.; Lewis, S.; Middeldorp, S.; Nelen, W.; Peramo, B.; Quenby, S.; et al. ESHRE guideline: Recurrent pregnancy loss. Hum. Reprod. Open 2018, 2018, hoy004. [Google Scholar]
- Valenti, D.; La Vignera, S.; Condorelli, R.A.; Rago, R.; Barone, N.; Vicari, E.; Calogero, A.E. Follicle-stimulating hormone treatment in normogonadotropic infertile men. Nat. Rev. Urol. 2013, 10, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Punab, M.; Poolamets, O.; Paju, P.; Vihljajev, V.; Pomm, K.; Ladva, R.; Korrovits, P.; Laan, M. Causes of male infertility: A 9-year prospective monocentre study on 1737 patients with reduced total sperm counts. Hum. Reprod. 2017, 32, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Tüttelmann, F.; Ruckert, C.; Röpke, A. Disorders of spermatogenesis: Perspectives for novel genetic diagnostics after 20 years of unchanged routine. Med. Genet. 2018, 30, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Yan, M.; Ge, R.; Cheng, C.Y. Crosstalk between Sertoli and Germ Cells in Male Fertility. Trends Mol. Med. 2020, 26, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.R.; Liu, Y.X. Regulation of spermatogonial stem cell self-renewal and spermatocyte meiosis by Sertolicell signaling. Reproduction 2015, 149, R159–R167. [Google Scholar] [CrossRef] [Green Version]
- Cannarella, R.; Condorelli, R.A.; Duca, Y.; La Vignera, S.; Calogero, A.E. New insights into the genetics of spermatogenic failure: A review of the literature. Hum. Genet. 2019, 138, 125–140. [Google Scholar] [CrossRef]
- Sironen, A.; Uimari, P.; Venhoranta, H.; Andersson, M.; Vilkki, J. An exonic insertion within Tex14 gene causes spermatogenic arrest in pigs. BMC Genom. 2011, 12, 591. [Google Scholar] [CrossRef] [Green Version]
- Duca, Y.; Calogero, A.E.; Cannarella, R.; Condorelli, R.A.; La Vignera, S. Current and emerging medical therapeutic agents for idiopathic male infertility. Exp. Opin. Pharmacother. 2019, 20, 55–67. [Google Scholar] [CrossRef]
- Calogero, A.E.; Condorelli, R.A.; Russo, G.I.; La Vignera, S. Conservative Nonhormonal Options for the Treatment of Male Infertility: Antibiotics, Anti-Inflammatory Drugs, and Antioxidants. Biomed. Res. Int. 2017, 2017, 4650182. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Yang, Y.; Feng, G.H.; Sun, B.F.; Chen, J.Q.; Li, Y.F.; Chen, Y.S.; Zhang, X.X.; Wang, C.X.; Jiang, L.Y.; et al. Mettl3-mediated m6A regulates spermatogonial differentiation and meiosis initiation. Cell Res. 2017, 27, 1100–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, J.; Chen, M.; Ji, S.; Wang, X.; Wang, Y.; Huang, X.; Yang, L.; Wang, Y.; Cui, X.; Lv, L.; et al. Equatorin is not essential for acrosome biogenesis but is required for the acrosome reaction. Biochem. Biophys. Res. Commun. 2014, 444, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Kanemori, Y.; Koga, Y.; Sudo, M.; Kang, W.; Kashiwabara, S.; Ikawa, M.; Hasuwa, H.; Nagashima, K.; Ishikawa, Y.; Ogonuki, N.; et al. Biogenesis of sperm acrosome is regulated by pre-mRNA alternative splicing of Acrbp in the mouse. Proc. Natl. Acad. Sci. USA 2016, 113, E3696–E3705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stival, C.; Molina, L.P.; Del, C.; Paudel, B.; Buffone, M.G.; Visconti, P.E.; Krapf, D. Sperm Capacitation and Acrosome Reaction in Mammalian Sperm. In Sperm Acrosome Biogenesis and Function during Fertilization; Springer: Cham, Switzerland, 2016; Volume 220, pp. 93–106. [Google Scholar]
- Shimada, M.; Yanai, Y.; Okazaki, T.; Noma, N.; Kawashima, I.; Mori, T.; Richards, J.S. Hyaluronan fragments generated by sperm-secreted hyaluronidase stimulate cytokine/chemokine production via the TLR2 and TLR4 pathway in cumulus cells of ovulated COCs, which may enhance fertilization. Development 2008, 135, 2001–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanii, I.; Aradate, T.; Matsuda, K.; Komiya, A.; Fuse, H. PACAP-mediated sperm-cumulus cell interaction promotes fertilization. Reproduction 2011, 141, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Bleil, J.D.; Wassarman, P.M. Structure and function of the zona pellucida: Identification and characterization of the proteins of the mouse oocyte’s zona pellucida. Dev. Biol. 1980, 76, 185–202. [Google Scholar] [CrossRef]
- Sun, T.C.; Wang, J.H.; Wang, X.X.; Liu, X.M.; Zhang, C.L.; Hao, C.F.; Ma, W.Z.; Deng, S.L.; Liu, Y.X. Effects of sperm proteins on fertilization in the female reproductive tract. Front. Biosci. (Landmark Ed.) 2019, 24, 735–749. [Google Scholar]
- Wen, L.; Liu, Q.; Xu, J.; Liu, X.; Shi, C.; Yang, Z.; Zhang, Y.; Xu, H.; Liu, J.; Yang, H.; et al. Recent advances in mammalian reproductive biology. Sci. China Life Sci. 2019, 1–41. [Google Scholar] [CrossRef]
- Castillo, J.; Jodar, M.; Oliva, R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum. Reprod. Update 2018, 24, 535–555. [Google Scholar] [CrossRef] [Green Version]
- Farhadova, S.; Gomez-Velazquez, M.; Feil, R. Stability and Lability of Parental Methylation Imprints in Development and Disease. Genes 2019, 10, 999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannarella, R.; Condorelli, R.A.; La Vignera, S.; Bellucci, C.; Luca, G.; Calafiore, R.; Calogero, A.E. IGF2 and IGF1R mRNAs Are Detectable in Human Spermatozoa. World J. Men Health 2019, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacone, F.; Cannarella, R.; Mongioì, L.M.; Alamo, A.; Condorelli, R.A.; Calogero, A.E.; La Vignera, S. Epigenetics of Male Fertility: Effects on Assisted Reproductive Techniques. World J. Men Health 2019, 37, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, F.C.; Charalambous, M. Genomic imprinting, growth and maternal-fetal interactions. J. Exp. Biol. 2018, 221, jeb164517. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Taskin, O.; Albert, A.; Bedaiwy, M.A. Association between sperm DNA fragmentation and idiopathic recurrent pregnancy loss: A systematic review and meta-analysis. Reprod. Biomed. Online 2019, 38, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Deng, C.; Li, T.; Xie, Y.; Guo, Y.; Yang, Q.Y.; Liang, X.; Deng, C.H.; Liu, G.H. Sperm DNA fragmentation index influences assisted reproductive technology outcome: A systematic review and meta-analysis combined with a retrospective cohort study. Andrologia 2019, 51, e13263. [Google Scholar] [CrossRef]
- McQueen, D.B.; Zhang, J.; Robins, J.C. Sperm DNA fragmentation and recurrent pregnancy loss: A systematic review and meta-analysis. Fertil. Steril. 2019, 112, 54–60. [Google Scholar] [CrossRef]
- Nanassy, L.; Liu, L.; Griffin, J.; Carrell, D.T. The clinical utility of the protamine 1/protamine 2 ratio in sperm. Protein Pept. Lett. 2011, 8, 772–777. [Google Scholar] [CrossRef]
- Muratori, M.; De Geyter, C. Chromatin condensation, fragmentation of DNA and differences in the epigenetic signature of infertile men. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 117–126. [Google Scholar] [CrossRef]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Baskaran, S.; Dutta, S.; Sengupta, P.; Khorshid, H.R.K.; Esteves, S.; Gilany, K.; Hedayati, M.; et al. Reactive oxygen species-induced alterations in H19-Igf2 methylation patterns, seminal plasma metabolites, and semen quality. J. Assist. Reprod. Genet. 2019, 36, 241–253. [Google Scholar] [CrossRef]
- Ni, W.; Pan, C.; Pan, Q.; Fei, Q.; Huang, X.; Zhang, C. Methylation levels of IGF2 and KCNQ1 in spermatozoa from infertile men are associated with sperm DNA damage. Andrologia 2019, 51, e13239. [Google Scholar] [CrossRef] [PubMed]
- Montjean, D.; Zini, A.; Ravel, C.; Belloc, S.; Dalleac, A.; Copin, H.; Boyer, P.; McElreavey, K.; Benkhalifa, M. Sperm global DNA methylation level: Association with semen parameters and genome integrity. Andrology 2015, 3, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Santi, D.; De Vincentis, S.; Magnani, E.; Spaggiari, G. Impairment of sperm DNA methylation in male infertility: A meta-analytic study. Andrology 2017, 5, 695–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, R.P. Post-meiotic gene expression. Trends Genet. 1990, 8, 2664–2669. [Google Scholar] [CrossRef] [Green Version]
- Vibranovski, M.D.; Chalopin, D.S.; Lopes, H.F.; Long, M.; Karr, T.L. Direct evidence for postmeiotic transcription during Drosophila melanogaster spermatogenesis. Genetics 2010, 186, 431–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Chao, S.B.; Xiao, L.; Wang, Z.B.; Meng, T.G.; Li, Y.Y.; Han, Z.M.; Ouyang, Y.C.; Hou, Y.; Sun, Q.Y.; et al. Sperm-carried RNAs play critical roles in mouse embryonic development. Oncotarget 2017, 8, 67394–67405. [Google Scholar] [CrossRef] [Green Version]
- Conine, C.C.; Sun, F.; Song, L.; Rivera-Pérez, J.A.; Rando, O.J. MicroRNAs Absent in Caput Sperm Are Required for Normal Embryonic Development. Dev. Cell 2019, 50, 7–8. [Google Scholar] [CrossRef]
- García-Herrero, S.; Meseguer, M.; Martínez-Conejero, J.A.; Remohí, J.; Pellicer, A.; Garrido, N. The transcriptome of spermatozoa used in homologous intrauterine insemination varies considerably between samples that achieve pregnancy and those that do not. Fertil. Steril. 2010, 94, 1360–1373. [Google Scholar] [CrossRef]
- Arney, K.L. H19 and Igf2—enhancing the confusion? Trends Genet. 2003, 19, 17–23. [Google Scholar] [CrossRef]
- Sakian, S.; Louie, K.; Wong, E.C.; Havelock, J.; Kashyap, S.; Rowe, T.; Taylor, B.; Ma, S. Altered gene expression of H19 and IGF2 in placentas from ART pregnancies. Placenta 2015, 36, 1100–1105. [Google Scholar] [CrossRef]
- Garrod, D.; Chidgey, M. Desmosome structure, composition and function. Biochim. Biophys. Acta Biomembr. 2008, 1778, 572–587. [Google Scholar] [CrossRef] [PubMed]
- Engel, K.M.; Baumann, S.; Rolle-Kampczyk, U.; Schiller, J.; von Bergen, M.; Grunewald, S. Metabolomic profiling reveals correlations between spermiogram parameters and the metabolites present in human spermatozoa and seminal plasma. PLoS ONE 2019, 14, e0211679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shettles, L.B. The Relation of Dietary Deficiencies to Male Fertility. Fertil. Steril. 1960, 11, 88–99. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Rhoads, J.M.; Satterfield, C.; Smith, S.; Spencer, T.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, J. Studies on arginine in human semen. II. The effects of medication with L-arginine-HCL on male infertility. Bull. Osaka Med. Sch. 1967, 13, 84–89. [Google Scholar] [PubMed]
- Aliabadi, E.; Mehranjani, M.S.; Borzoei, Z.; Talaei-Khozani, T.; Mirkhani, H.; Tabesh, H. Effects of L-carnitine and L-acetyl-carnitine on testicular sperm motility and chromatin quality. Iran. J. Reprod. Med. 2012, 10, 77–82. [Google Scholar]
- Lu, J.-C.; Jing, J.; Yao, Q.; Fan, K.; Wang, G.-H.; Feng, R.-X.; Liang, Y.J.; Chen, L.; Ge, Y.F.; Yao, B. Relationship between Lipids Levels of Serum and Seminal Plasma and Semen Parameters in 631 Chinese Subfertile Men. PLoS ONE 2016, 11, e0146304. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sperm Parameter | Genes |
|---|---|
| Sperm number | CATSPER1, CCDC39, DAZ1, DAZ2, DAZ3, DAZ4, DBY, DMC1, DMRT1, DNAH6, FANCM, HAUS7, HSF2, KLHL10, MEIOB, NR5A1, PLK-4, SEPT12, SOHLH1, SPINK2, SYCE1, SYCP3, TAF4B, TDRD6, TEX11, TEX14, TEX15, USP26, ZMYND15 |
| Sperm motility | AK7, CCDC39, CEP135, CFAP43, CFAP44, CFAP69, DNAAF1, DNAAF2, DNAAF3, DNAH1, DNAH5, DNAI1, DNAI2, DNAJB13, DYX1C1, FSIP2, HEATR2, HYDIN, LRRC6, PIH1D3, RSPH1, RSPH4A, RSPH9, SLC26A8, WDR66, ZMYND10 |
| Sperm morphology | AURKC, BRDT, DPY19L2, SPATA16, SUN5, TSGA10 |
| Process | Imprinted Genes |
|---|---|
| Imprinted genes involved in placenta and embryo development and growth in mice | |
| Placenta establishing | Peg10 |
| Nutrient transport capacity and surface area for exchange | Igf2, Grb10 |
| Fetal growth | Igf2, Igf2r, Cdkn1c, Grb10 |
| Imprinted genes involved in sperm quality | |
| CREM | Increased methylation is associated with decreased semen quality |
| DAZL | Increased methylation in OAT patients compared to controls |
| FAM50B | Decreased methylation levels are associated with asthenozoospermia |
| GNAS | Decreased methylation levels are associated with asthenozoospermia |
| GLT2 | Abnormal methylation levels are associated with oligozoospermia |
| H19 | Decreased methylation is associated with male infertility |
| KCNQ1OT1 | Increased methylation in patients with abnormal sperm parameters |
| MEST | Increased methylation is associated with male infertility |
| RHOX | Increased methylation is associated with male infertility |
| SNRPN | Increased methylation is associated with male infertility |
| ZAC | Increased methylation is associated with oligozoospermia |
| CREM | Increased methylation is associated with decreased semen quality |
| DAZL | Increased methylation in OAT patients compared to controls |
| FAM50B | Decreased methylation levels are associated with asthenozoospermia |
| Process | Proteins |
|---|---|
Fertilization
| AAAS, ABHD2, ACR, ADAM2, ADAM20, ADAM21, ADAM30, AKAP3, AKAP4, APOB, ASH1L, ATP1A4, ATP8B3, B4GALT1, BAX, BCL2L1, BSPH1, CATSPER1, CATSPER2, CATSPER4, CATSPERB, CATSPERD, CATSPERG, CCDC136, CCT2, CCT3, CCT4, CCT5, CCT7, CCT8, CD46, CD9, CDK1, CLGN, CLIC4, CRISP1, DEFB126, DNALI1, DUOX2, ELSPBP1, EQTN, GLIPR1L1, GLRB, GNPDA1, H3F3A, HEXB, HSPA1L, HVCN1, INSL6, IQCF1, IZUMO1, KCNU1, KLHL10, LY6K, MAEL, MFGE8, NLRP5, NOX5, PARK7, PCSK4, PKDREJ, PLCB1, PLCD4, PLCZ1, PRSS37, RNASE10, ROPN1B, SERPINA10, SERPINA5, SLC22A16, SMAD4, SMCP, SPA17, SPACA3, SPACA6, SPACA7, SPAG1, SPAG8, SPAM1, SPESP1, SPINK2, SPTBN4, STX2, SYCP2, TARBP2, TCP1, TCP11, TDRKH, TEKT3, TEX11, TEX15, TRIM36, TRPC7, TUBGCP3, UBAP2L, UBE3A, UBXN8, WBP2NL, ZAN, ZP1, ZP2, ZPBP, ZPBP2 |
Preimplantation embryo development
| AP1G1, AP2M1, ARFGEF2, ARHGDIB, ARPC3, ATP6V0C, B3GAT3, B3GNT5, BRCA2, BRK1, BSG, C1QBP, C2orf61, CALCA, CAPN2, CAPN5, CDH1, CDK11B, CENPF, CHD4, CTNNA1, CTTN, CUL3, DAD1, DDR1, DDX20, DMAP1, DSC3, EIF3B, EIF6, FAF1 FASN, FKBP4, HNRNPL, HSBP1, IGFBP7, KIF11, LATS1, MCL1, MMP2, MMP9, NASP, NDEL1, NEPRO, NUP214, ODF2, PCSK1, PCSK5, PCYT1A, PDCD2, PFN1, PPM1B, PRKRA, PRLR, PRPF19, PSMC3, PSMC4, PTGS2, PUM1, RACGAP1, RBM19, RPL7L1, RPS19, RPSA, RRM1, RUVBL1, SBDS, SCGB1A1, SEC24D, SMURF2, SNAP23, SOD1, SPP1, SRSF3, TBP, TGFBR1, TGFBR2, TIMM23, TIMP1, TKT, TMED10, TOP1, TPM3, TRIM28, TRRAP, TUBG1, VCP, ZPR1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cannarella, R.; Condorelli, R.A.; Mongioì, L.M.; La Vignera, S.; Calogero, A.E. Molecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male Infertility. Int. J. Mol. Sci. 2020, 21, 1728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051728
Cannarella R, Condorelli RA, Mongioì LM, La Vignera S, Calogero AE. Molecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male Infertility. International Journal of Molecular Sciences. 2020; 21(5):1728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051728
Chicago/Turabian StyleCannarella, Rossella, Rosita A. Condorelli, Laura M. Mongioì, Sandro La Vignera, and Aldo E. Calogero. 2020. "Molecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male Infertility" International Journal of Molecular Sciences 21, no. 5: 1728. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051728