Altered Cofactor Preference of Thermostable StDAPDH by a Single Mutation at K159


Abstract
:1. Introduction
2. Results and Discussion
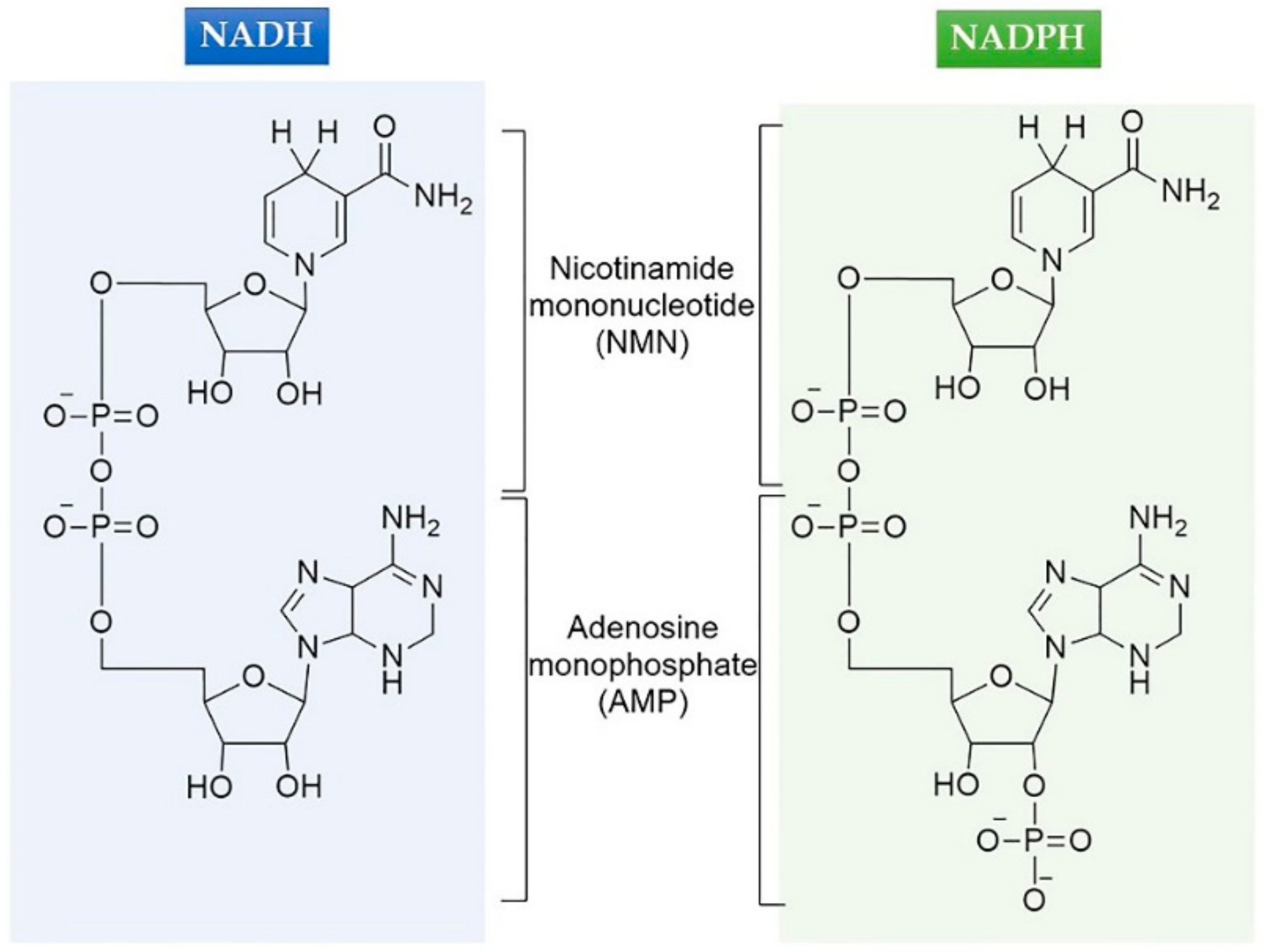
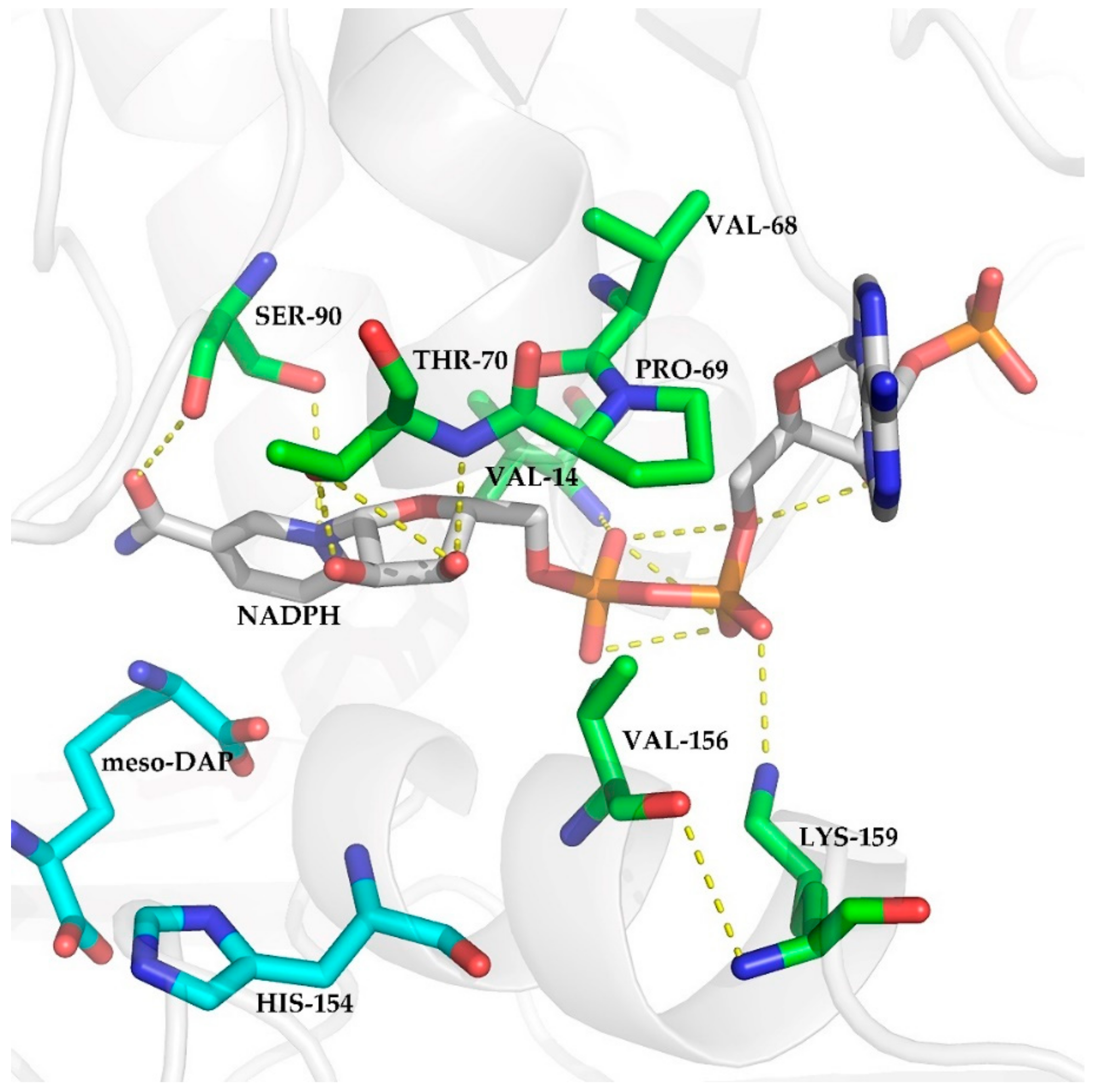
2.1. Selection of Mutations
2.2. Determination of Kinetic Parameters with NADH as Cofactor.
2.3. MD Simulation Analysis
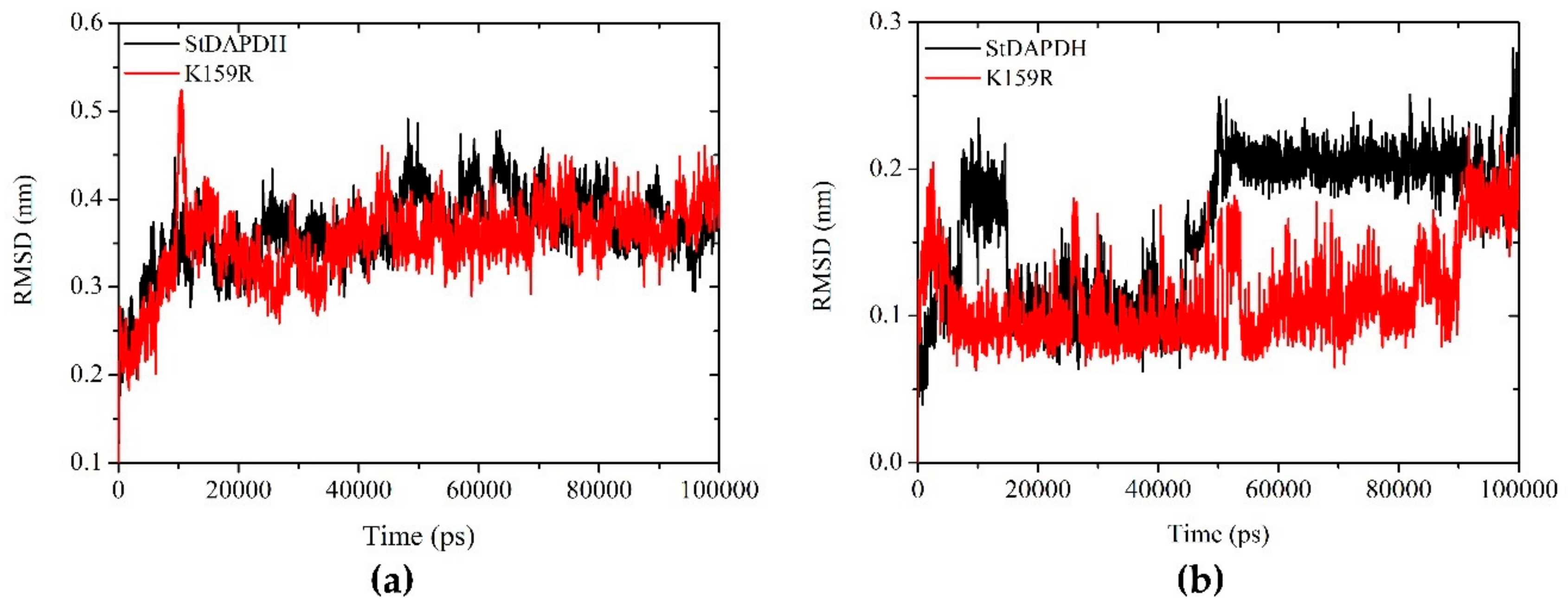
2.3.1. Root Mean Square Deviation Analysis
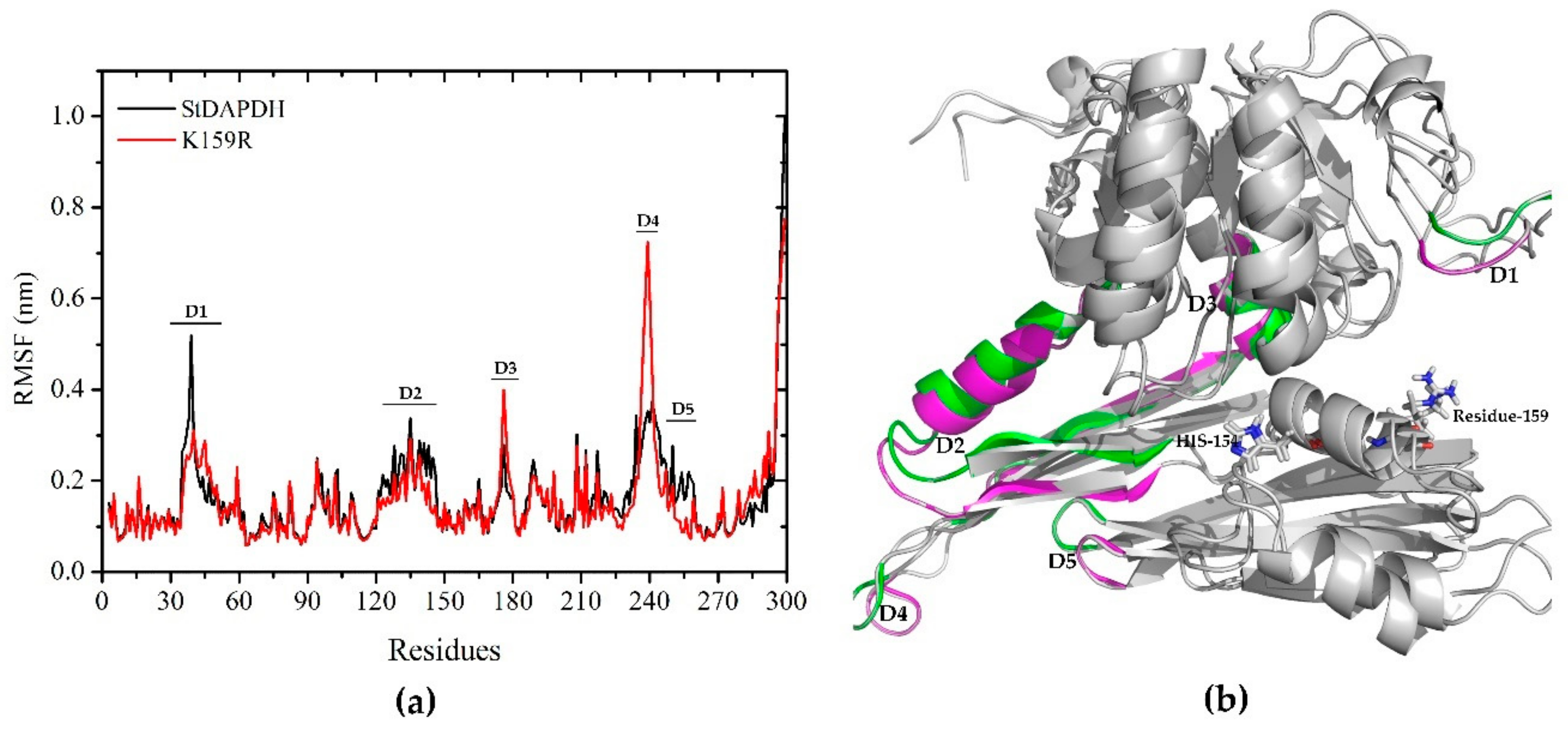
2.3.2. Root Mean Square Fluctuation Analysis
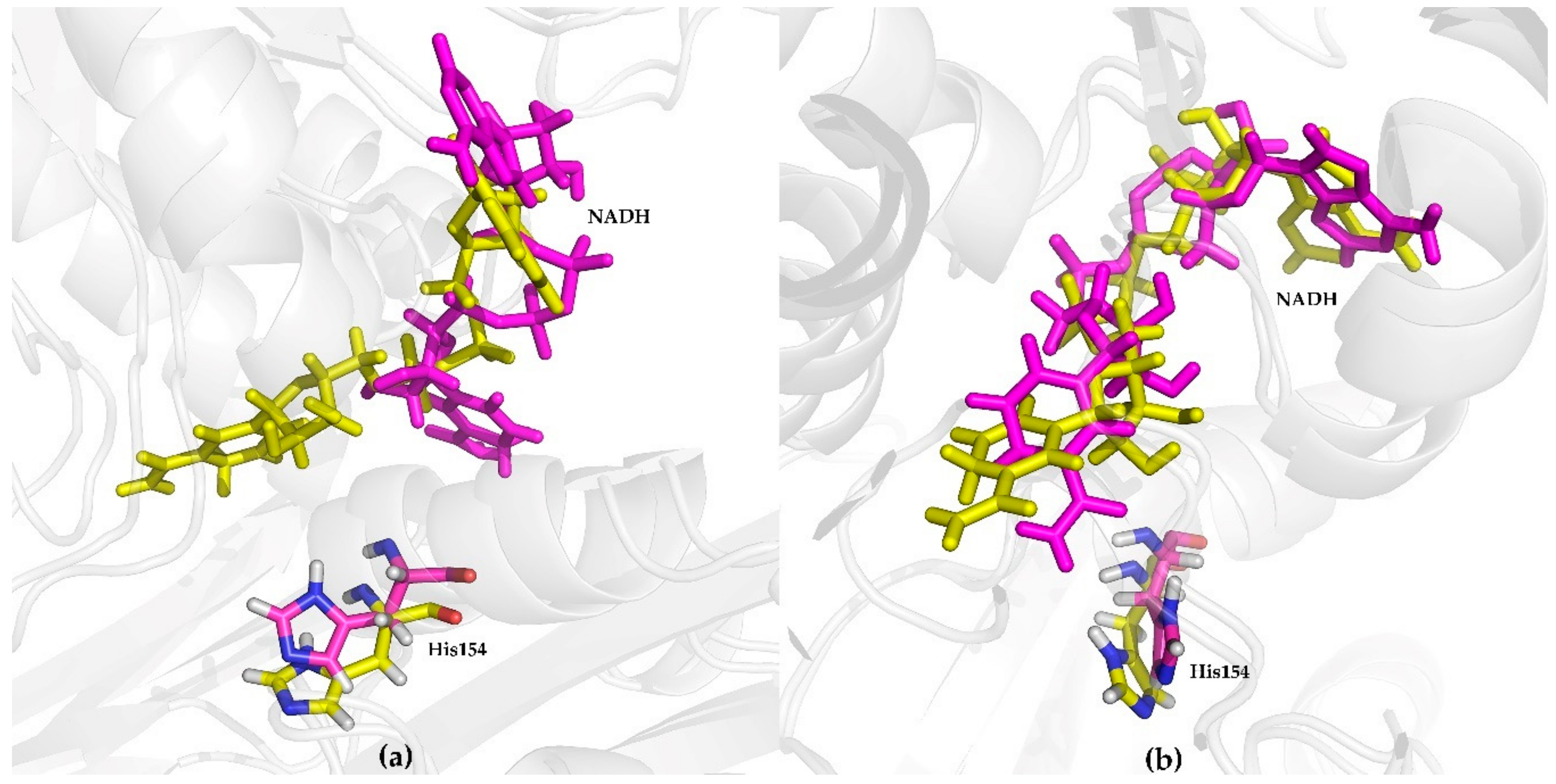
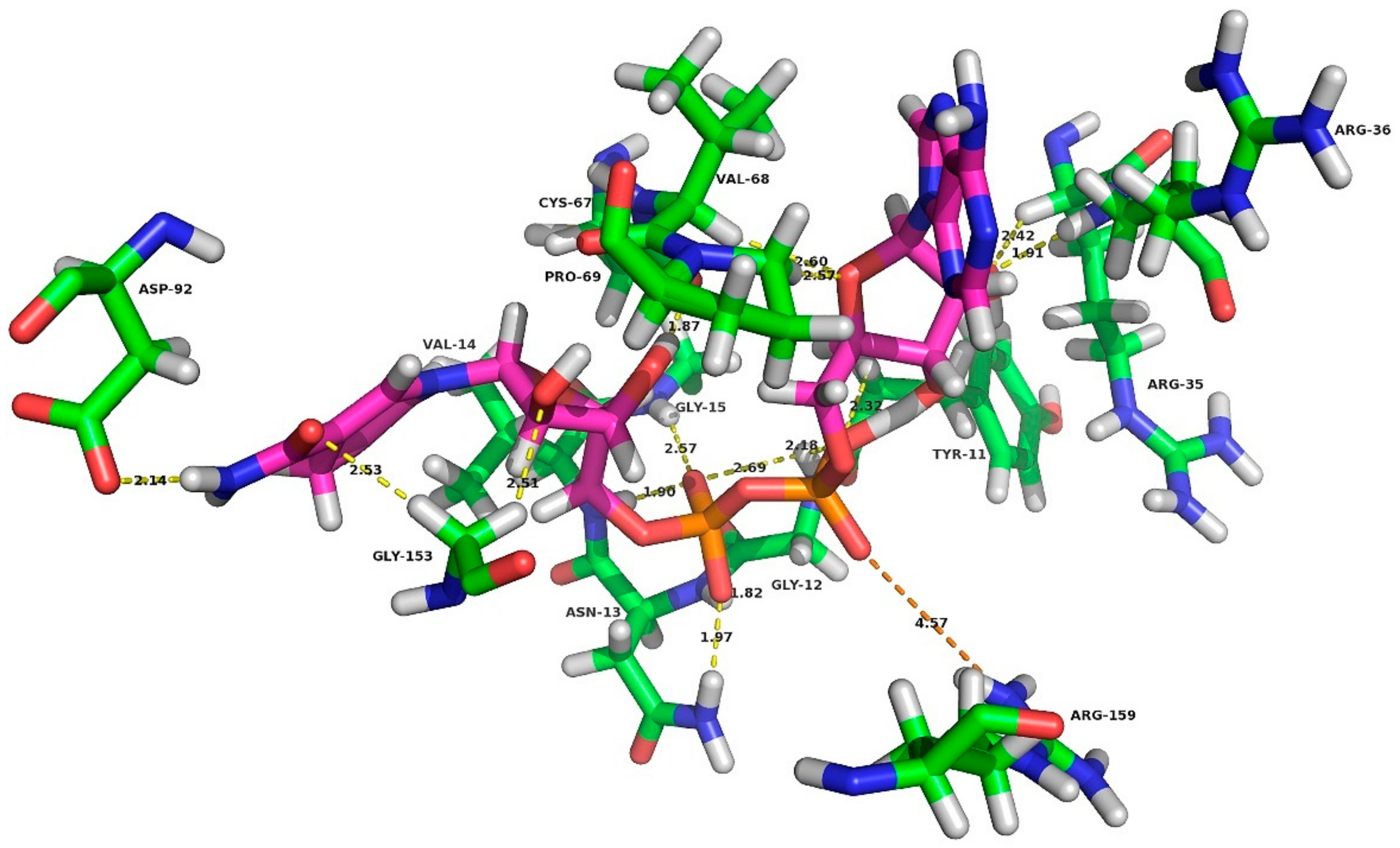
2.3.3. Conformational Analysis
3. Materials and Methods
3.1. Materials and Strains
3.2. Overexpression and Purification of Proteins
3.3. Determination of Kinetic Constants
3.4. MD Simulations
3.5. Analysis of Molecular Dynamics Trajectories
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NADP(H) | Nicotinamide adenine dinucleotide phosphate |
| NAD(H) | Nicotinamide adenine dinucleotide |
| AMP | Adenosine monophosphate |
| NMN | Nicotinamide mononucleotide |
| meso-DAPDH | meso-Diaminopimelate dehydrogenase |
| StDAPDH | meso-DAPDH from Symbiobacterium thermophilum IAM14863 |
| UtDAPDH | meso-DAPDH from Ureibacillus thermophaericus strain A1 |
| meso-DAP | meso-2,6-Diaminopimelate |
| MD | Molecular dynamics |
| NVT | Constant number of particles, volume, and temperature |
| NPT | Constant number of particles, pressure, and temperature |
References
- Gao, X.; Chen, X.; Liu, W.; Feng, J.; Wu, Q.; Hua, L.; Zhu, D. A novel meso-diaminopimelate dehydrogenase from Symbiobacterium thermophilum: Overexpression, characterization, and potential for D-amino acid synthesis. Appl. Environ. Microbiol. 2012, 78, 8595–8600. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Ma, Q.; Zhu, H. Distribution, industrial applications, and enzymatic synthesis of D-amino acids. Appl. Microbiol. Biotechnol. 2015, 99, 3341–3349. [Google Scholar] [CrossRef]
- Marcone, G.L.; Rosini, E.; Crespi, E.; Pollegioni, L. D-amino acids in foods. Appl. Microbiol. Biotechnol. 2020, 104, 555–574. [Google Scholar] [CrossRef] [PubMed]
- Akita, H.; Suzuki, H.; Doi, K.; Ohshima, T. Efficient synthesis of D-branched-chain amino acids and their labeled compounds with stable isotopes using D-amino acid dehydrogenase. Appl. Microbiol. Biotechnol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhu, Z.; Huang, R.; Zhang, Y.-H.P. Coenzyme engineering of a hyperthermophilic 6-phosphogluconate dehydrogenase from NADP to NAD with its application to biobatteries. Sci. Rep. 2016, 6, 36311. [Google Scholar] [CrossRef] [Green Version]
- Antranikian, G.; Vorgias, C.E.; Bertoldo, C. Extreme environments as a resource for microorganisms and novel biocatalysts. Adv. Biochem. Eng. Biotechnol. 2005, 96, 219–262. [Google Scholar] [PubMed]
- Akita, H.; Fujino, Y.; Doi, K.; Ohshima, T. Highly stable meso-diaminopimelate dehydrogenase from an Ureibacillus thermosphaericus strain A1 isolated from a Japanese compost: Purification, characterization and sequencing. AMB Express 2011, 1, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Zhang, Z.; Zhang, Y.n.; Li, Y.; Zhu, H.; Wang, S.; Li, C. A newly determined member of the meso-diaminopimelate dehydrogenase family with a broad substrate spectrum. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Akita, H.; Doi, K.; Kawarabayasi, Y.; Ohshima, T. Creation of a thermostable NADP+-dependent D-amino acid dehydrogenase from Ureibacillus thermosphaericus strain A1 meso-diaminopimelate dehydrogenase by site-directed mutagenesis. Biotechnol. Lett. 2012, 34, 1693–1699. [Google Scholar] [CrossRef]
- Akita, H.; Hayashi, J.; Sakuraba, H.; Ohshima, T. Artificial thermostable D-amino acid dehydrogenase: Creation and application. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Suzuki, S.; Horinouchi, S.; Beppu, T. Growth of a Tryptophanase-producing thermophile, Symbiobacterium thermophilum gen. nov., sp. nov., is dependent on co-culture with a Bacillus sp. J. Gen. Microbiol. 1988, 134, 2353–2362. [Google Scholar] [CrossRef]
- Ueda, K.; Ohno, M.; Yamamoto, K.; Nara, H.; Mori, Y.; Shimada, M.; Hayashi, M.; Oida, H.; Terashima, Y.; Nagata, M.; et al. Distribution and diversity of symbiotic thermophiles, Symbiobacterium thermophilum and related bacteria, in natural environments. Appl. Environ. Microbiol. 2001, 67, 3779–3784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, M.; Okano, I.; Watsuji, T.-O.; Kakinuma, T.; Ueda, K.; Beppu, T. Establishing the independent culture of a strictly symbiotic bacterium Symbiobacterium thermophilum from its supporting Bacillus strain. Biosci. Biotechnol. Biochem. 1999, 63, 1083–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misono, H.; Ogasawara, M.; Nagasaki, S. Purification and properties of meso-diaminopimelate dehydrogenase from Brevibacterium sp. Agric. Biol. Chem. 1986, 50, 1329–1330. [Google Scholar] [CrossRef]
- White, P. The essential role of diaminopimelate dehydrogenase in the biosynthesis of lysine by Bacillus sphaericus. J. Gen. Microbiol. 1983, 129, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Misono, H.; Togawa, H.; Yamamoto, T.; Soda, K. Occurrence of meso-α,ε-diaminopimelate dehydrogenase in Bacillus sphaericus. Biochem. Biophys. Res. Commun. 1976, 72, 89–93. [Google Scholar] [CrossRef]
- Woodyer, R.; van der Donk, W.A.; Zhao, H. Relaxing the nicotinamide cofactor specificity of phosphite dehydrogenase by rational design. Biochemistry 2003, 42, 11604–11614. [Google Scholar] [CrossRef]
- Rollin, J.A.; Tam, T.K.; Zhang, Y.H.P. New biotechnology paradigm: Cell-free biosystems for biomanufacturing. Green Chem. 2013, 15, 1708–1719. [Google Scholar] [CrossRef]
- Banta, S.; Swanson, B.A.; Wu, S.; Jarnagin, A.; Anderson, S. Alteration of the specificity of the cofactor-binding pocket of Corynebacterium 2,5-diketo-D-gluconic acid reductase A. Protein Eng. 2002, 15, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, P. Cofactor regeneration for sustainable enzymatic biosynthesis. Biotechnol. Adv. 2007, 25, 369–384. [Google Scholar] [CrossRef]
- Chenault, H.K.; Whitesides, G.M. Regeneration of nicotinamide cofactors for use in organic synthesis. Appl. Biochem. Biotechnol. 1987, 14, 147–197. [Google Scholar] [CrossRef] [PubMed]
- Liese, A.; Seelbach, K.; Wandrey, C. Industrial Biotransformations; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Vedha-Peters, K.; Gunawardana, M.; Rozzell, J.D.; Novick, S.J. Creation of a broad-range and highly stereoselective D-amino acid dehydrogenase for the one-step synthesis of D-amino acids. J. Am. Chem. Soc. 2006, 128, 10923–10929. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.T.; Wu, L.H.; Knight, J.A. Stability of NADPH: Effect of various factors on the kinetics of degradation. Clin. Chem. 1986, 32, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.-H.; Whitesides, G.M. Enzyme-catalyzed organic synthesis: NAD(P)H cofactor regeneration by using glucose-6-phosphate and the glucose-5-phosphate dehydrogenase from Leuconostoc mesenteroides. J. Am. Chem. Soc. 1981, 103, 4890–4899. [Google Scholar] [CrossRef]
- Cahn, J.K.B.; Werlang, C.A.; Baumschlager, A.; Brinkmann-Chen, S.; Mayo, S.L.; Arnold, F.H. A general tool for engineering the NAD/NADP cofactor preference of oxidoreductases. ACS Synth. Biol. 2017, 6, 326–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, E.; Wheeldon, I.R.; Banta, S. Broadening the cofactor specificity of a thermostable alcohol dehydrogenase using rational protein design introduces novel kinetic transient behavior. Biotechnol. Bioeng. 2010, 107, 763–774. [Google Scholar] [CrossRef]
- Zeng, Q.-K.; Du, H.-L.; Wang, J.-F.; Wei, D.-Q.; Wang, X.-N.; Li, Y.-X.; Lin, Y. Reversal of coenzyme specificity and improvement of catalytic efficiency of Pichia stipitis xylose reductase by rational site-directed mutagenesis. Biotechnol. Lett. 2009, 31, 1025–1029. [Google Scholar] [CrossRef]
- Scrutton, N.S.; Berry, A.; Perham, R.N. Redesign of the coenzyme specificity of a dehydrogenase by protein engineering. Nature 1990, 343, 38–43. [Google Scholar] [CrossRef]
- Gao, X.; Ma, Q.; Chen, M.; Dong, M.; Pu, Z.; Zhang, X.; Song, Y. Insight into the highly conserved and differentiated cofactor-binding sites of meso-diaminopimelate dehydrogenase StDAPDH. J. Chem. Inf. Model. 2019, 59, 2331–2338. [Google Scholar] [CrossRef]
- Mesecar, A.D.; Stoddard, B.L.; Koshland, D.E. Orbital steering in the catalytic power of enzymes: Small structural changes with large catalytic consequences. Science 1997, 277, 202–206. [Google Scholar] [CrossRef]
- Cahn, J.K.B.; Baumschlager, A.; Brinkmann-Chen, S.; Arnold, F.H. Mutations in adenine-binding pockets enhance catalytic properties of NAD(P)H-dependent enzymes. Protein Eng. Des. Sel. PEDS 2016, 29, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddock, D.J.; Patrick, W.M.; Gerth, M.L. Substitutions at the cofactor phosphate-binding site of a clostridial alcohol dehydrogenase lead to unexpected changes in substrate specificity. Protein Eng. Des. Sel. PEDS 2015, 28, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Segura, L.; Riveros-Rosas, H.; Julián-Sánchez, A.; Muñoz-Clares, R.A. Residues that influence coenzyme preference in the aldehyde dehydrogenases. Chem. Biol. Interact. 2015, 234, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Oliveira, M.A.; Tai, H.-H. Critical residues for the coenzyme specificity of NAD+-dependent 15-hydroxyprostaglandin dehydrogenase. Arch Biochem. Biophys. 2003, 419, 139–146. [Google Scholar] [CrossRef]
- Liu, W.; Li, Z.; Huang, C.-H.; Guo, R.-T.; Zhao, L.; Zhang, D.; Chen, X.; Wu, Q.; Zhu, D. Structural and mutational studies on the unusual substrate specificity of meso-diaminopimelate dehydrogenase from Symbiobacterium thermophilum. ChemBioChem 2014, 15, 217–222. [Google Scholar] [CrossRef]
- Akita, H.; Seto, T.; Ohshima, T.; Sakuraba, H. Structural insight into the thermostable NADP+-dependent meso-diaminopimelate dehydrogenase from Ureibacillus thermosphaericus. Acta Crystallogr. Sect. D 2015, 71, 1136–1146. [Google Scholar] [CrossRef]
- Wang, F.; Scapin, G.; Blanchard, J.S.; Angeletti, R.H. Substrate binding and conformational changes of Clostridium glutamicum diaminopimelate dehydrogenase revealed by hydrogen/deuterium exchange and electrospray mass spectrometry. Protein Sci. 1998, 7, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ma, Q.; Dong, M.; Zhang, X.; Chen, Y.; Gao, X.; Song, Y. Essential role of amino acid position 71 in substrate preference by meso-diaminopimelate dehydrogenase from Symbiobacterium thermophilum IAM14863. Enzym. Microb. Technol. 2018, 111, 57–62. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Mark, P.; Nilsson, L. Structure and dynamics of the TIP3P, SPC, and SPC/E water models at 298 K. J. Phys. Chem. A 2001, 105, 9954–9960. [Google Scholar] [CrossRef]
- Miyamoto, S.; Kollman, P.A. Settle: An analytical version of the SHAKE and RATTLE algorithm for rigid water models. J. Comput. Chem. 1992, 13, 952–962. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | KM (mM) | kcat (s–1) | kcat/KM (mM-1·s–1) | Cofactor |
|---|---|---|---|---|
| StDAPDH | ND 1 | ND | ND | NADH |
| 8.86 ± 0.45 | 7.61 ± 0.18 | 0.86 | NADPH 2 | |
| V14L | ND | ND | ND | NADH |
| 10.08 ± 0.96 | 1.63 ± 0.08 | 0.16 | NADPH 2 | |
| K159R | 8.04 ± 0.85 | 1.34 ± 0.20 | 0.17 | NADH |
| 7.58 ± 0.45 | 8.07 ± 0.21 | 1.06 | NADPH 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Ma, Q.; Song, H.; Sun, X.; Li, Z.; Liu, M. Altered Cofactor Preference of Thermostable StDAPDH by a Single Mutation at K159. Int. J. Mol. Sci. 2020, 21, 1788. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051788
Gao X, Ma Q, Song H, Sun X, Li Z, Liu M. Altered Cofactor Preference of Thermostable StDAPDH by a Single Mutation at K159. International Journal of Molecular Sciences. 2020; 21(5):1788. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051788
Chicago/Turabian StyleGao, Xiuzhen, Qinyuan Ma, Huihui Song, Xinming Sun, Zhiyun Li, and Mingfei Liu. 2020. "Altered Cofactor Preference of Thermostable StDAPDH by a Single Mutation at K159" International Journal of Molecular Sciences 21, no. 5: 1788. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051788