Characterization and Cytotoxicity Assessment of the Lipophilic Fractions of Different Morphological Parts of Acacia dealbata

 , , , , and
, , , , and 
Abstract
:
1. Introduction
2. Results and Discussion
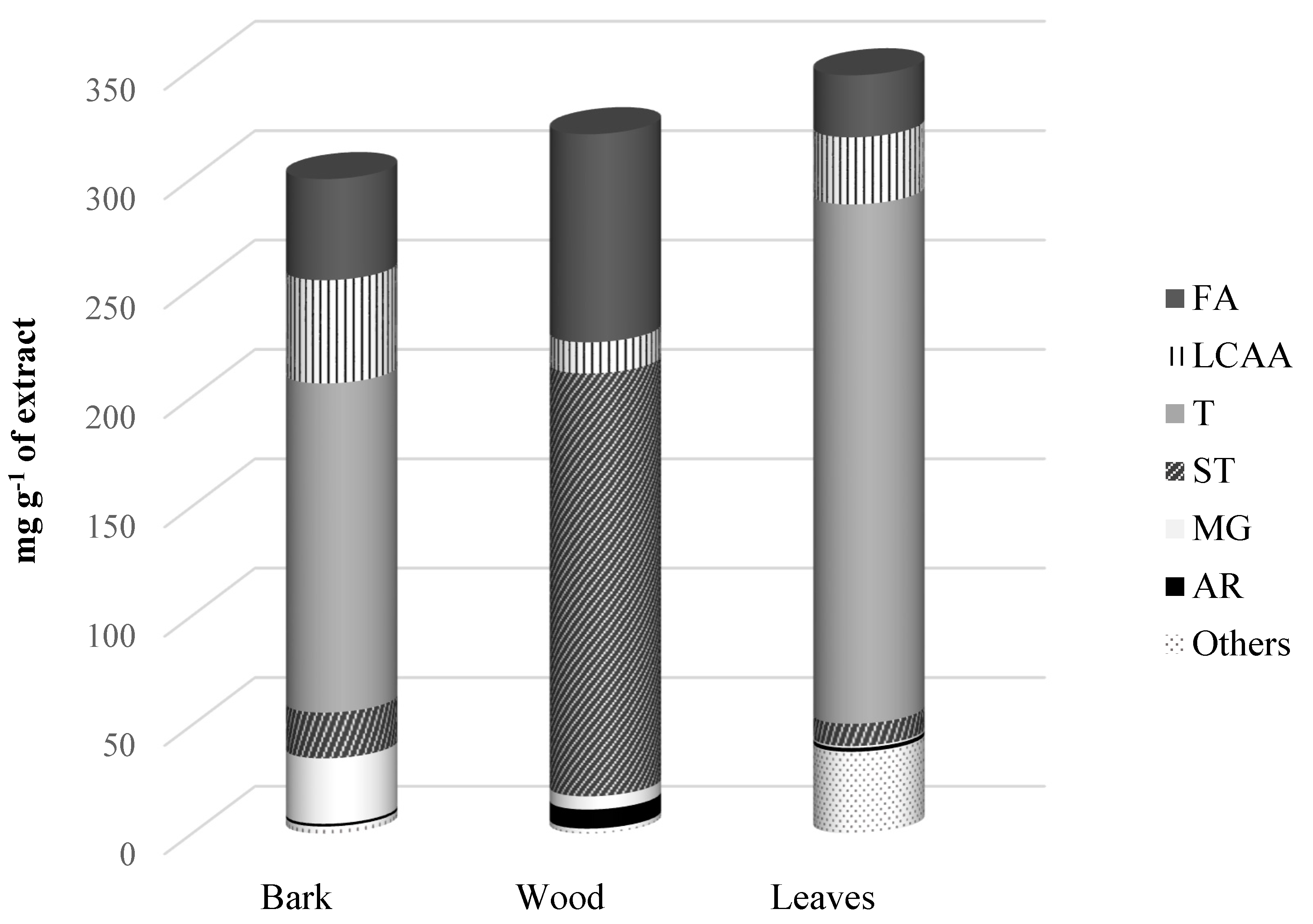
2.1. Lipophilic Extractives Yield of A. dealbata Bark, Wood and Leaves
2.2. Chemical Characterization of the Lipophilic Extract
2.2.1. Fatty Acids
2.2.2. Long-Chain Aliphatic Alcohols
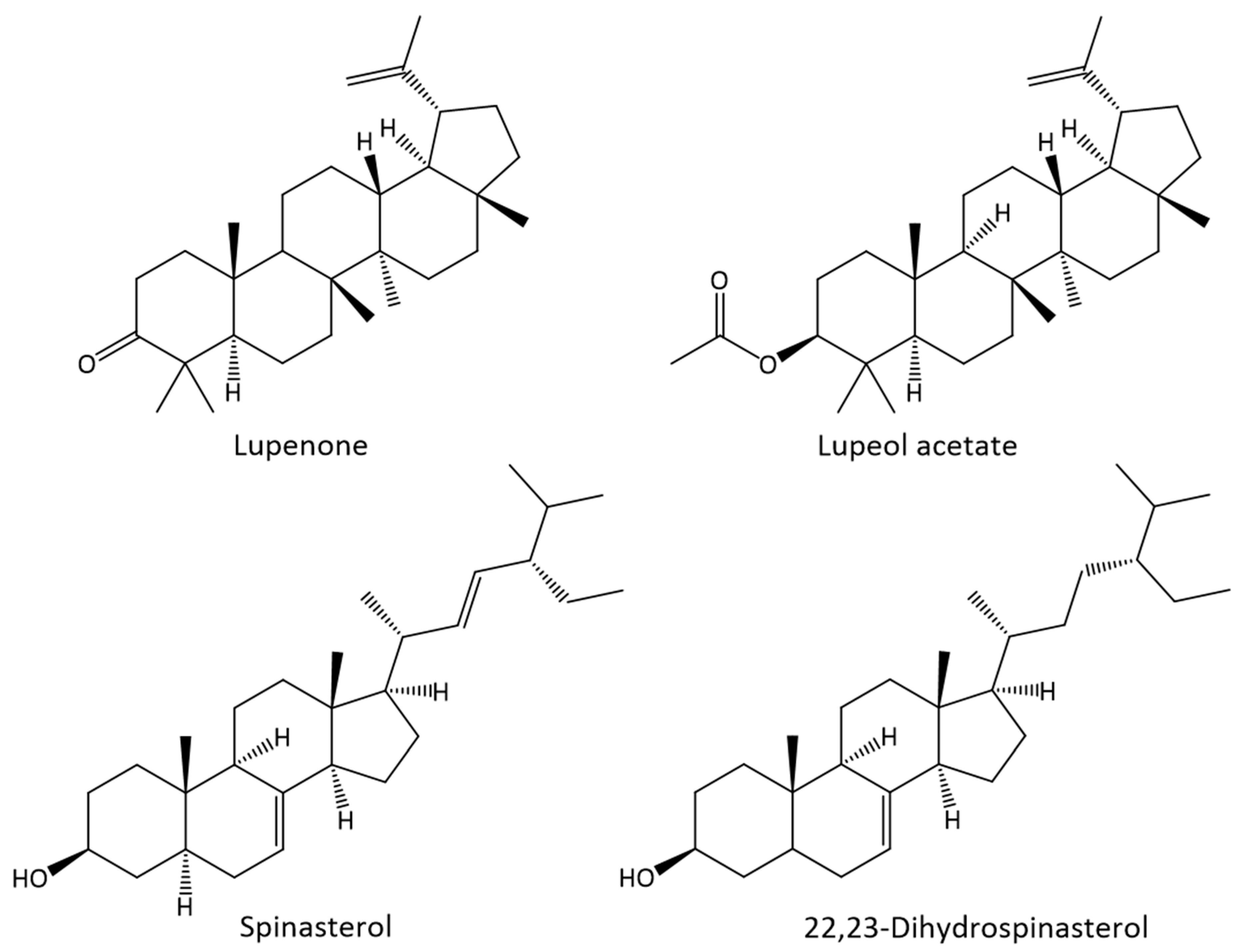
2.2.3. Terpenic Compounds
2.2.4. Sterols
2.2.5. Monoglycerides
2.2.6. Aromatic Compounds
2.2.7. Other Components
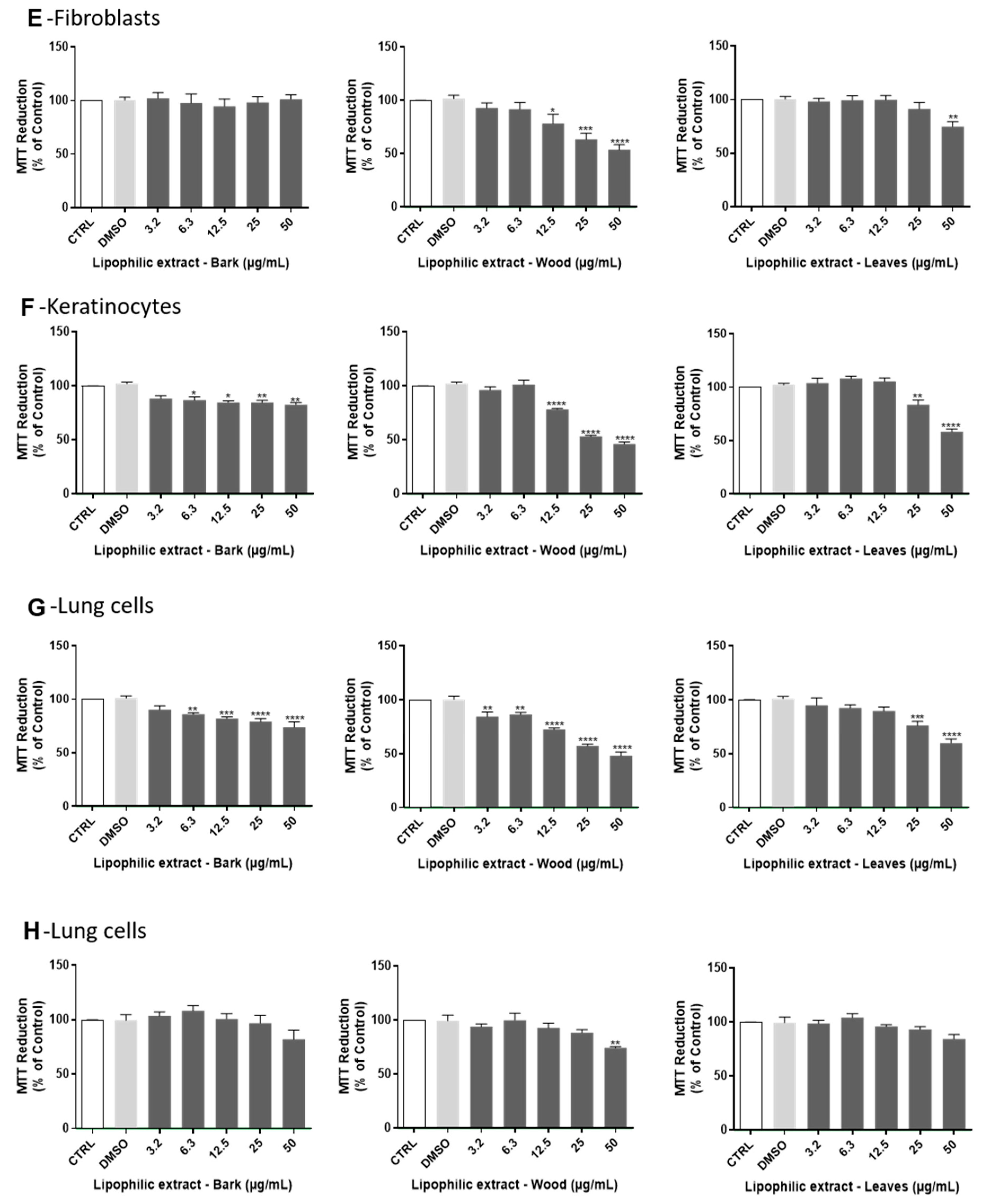
2.3. Cytotoxity Evaluation of A. dealbata Lipophilic Extracts
3. Materials and Methods
3.1. Reagents
3.2. Sample Collection
3.3. Characterization of Lipophilic Extracts
3.3.1. Lipophilic Compounds Extraction
3.3.2. GC–MS Analysis
3.4. Cytotoxicity Evaluation of Lipophilic Extracts
3.4.1. Cell Culture
3.4.2. Cell Viability Assay
3.4.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lorenzo, P.; Palomera-Pérez, A.; Reigosa, M.J.; González, L. Allelopathic interference of invasive Acacia dealbata Link on the physiological parameters of native understory species. Plant Ecol. 2011, 212, 403–412. [Google Scholar] [CrossRef]
- Sheppard, A.W.; Shaw, R.H.; Sforza, R. Top 20 environmental weeds for classical biological control in Europe: a review of opportunities, regulations and other barriers to adoption. Weed Res. 2006, 46, 93–117. [Google Scholar] [CrossRef]
- Lorenzo, P.; González, L.; Reigosa, M.J. The genus Acacia as invader: the characteristic case of Acacia dealbata Link in Europe. Ann. For. Sci. 2010, 67, 101. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, N.; Becerra, J.; Villaseñor-Parada, C.; Lorenzo, P.; González, L.; Hernández, V. Effects and identification of chemical compounds released from the invasive Acacia dealbata Link. Chem. Ecol. 2015, 31, 479–493. [Google Scholar] [CrossRef]
- WorldWideWattle Species Gallery. Acacia dealbata Link. Available online: http://worldwidewattle.com/speciesgallery/species-intro.php?id=17858 (accessed on 5 January 2020).
- Santos, A.J.A.; Anjos, O.M.S.; Simões, R.M.S. Papermaking potential of Acacia dealbata and Acacia melanoxylon. Appita. J. 2006, 59, 58–64. [Google Scholar]
- CABI Acacia dealbata. In: Invasive Species Compendium. Available online: https://www.cabi.org/isc/datasheet/2207#37C102CF-ECB0-4B32-B37E-3B3AC4ACA344 (accessed on 10 January 2020).
- Pereira, F.B.M.; Domingues, F.M.J.; Silva, A.M.S. Triterpenes from Acacia dealbata. Nat. Prod. Lett. 1996, 8, 97–103. [Google Scholar] [CrossRef]
- Silva, E.; Fernandes, S.; Bacelar, E.; Sampaio, A. Antimicrobial activity of aqueous, ethanolic and methanolic leaf extracts from Acacia spp. and Eucalyptus nicholii. African J. Tradit. Complement. Altern. Med. 2016, 13, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Gouws, A.J.; Shackleton, C.M. Abundance and correlates of the Acacia dealbata invasion in the northern Eastern Cape, South Africa. For. Ecol. Manage. 2019, 432, 455–466. [Google Scholar] [CrossRef]
- Ngorima, A.; Shackleton, C.M. Livelihood benefits and costs from an invasive alien tree (Acacia dealbata) to rural communities in the Eastern Cape, South Africa. J. Environ. Manage. 2019, 229, 158–165. [Google Scholar] [CrossRef]
- Rodolphe, P.; Katharina, B.; Meierhenrich, U.J.; Elise, C.; Georges, F.; Nicolas, B. Chemical composition of french mimosa absolute oil. J. Agric. Food Chem. 2010, 58, 1844–1849. [Google Scholar] [CrossRef]
- Yáñez, R.; Romaní, A.; Garrote, G.; Alonso, J.L.; Parajó, J.C. Processing of Acacia dealbata in aqueous media: First step of a wood biorefinery. Ind. Eng. Chem. Res. 2009, 48, 6618–6626. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Meireles, C.I.R.; Pinto Gomes, C.J.; Almeida Ribeiro, N.M.C. Historical development of the portuguese forest: The introduction of invasive species. Forests 2019, 10, 974. [Google Scholar] [CrossRef] [Green Version]
- Freire, C.S.R.; Coelho, D.S.C.; Santos, N.M.; Silvestre, A.J.D.; Pascoal Neto, C. Identification of Δ7 phytosterols and phytosteryl glucosides in the wood and bark of several Acacia species. Lipids 2005, 40, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Pinto, P.C.R.; Oliveira, C.; Costa, C.A.; Gaspar, A.; Faria, T.; Ataíde, J.; Rodrigues, A.E. Kraft delignification of energy crops in view of pulp production and lignin valorization. Ind. Crops Prod. 2015, 71, 153–162. [Google Scholar] [CrossRef]
- Freire, C.S.R.; Silvestre, A.J.D.; Neto, C.P. Demonstration of long-chain n-alkyl caffeates and Δ7-steryl glucosides in the bark of Acacia species by gas chromatography–mass spectrometry. Phytochem. Anal. 2007, 18, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Huang, X.; Wu, H.; Wang, X. Beneficial health effects of lupenone triterpene: A review. Biomed. Pharmacother. 2018, 103, 198–203. [Google Scholar] [CrossRef]
- Borges, F.R.M.; Silva, M.D.; Córdova, M.M.; Schambach, T.R.; Pizzolatti, M.G.; Santos, A.R.S. Anti-inflammatory action of hydroalcoholic extract, dichloromethane fraction and steroid α-spinasterol from Polygala sabulosa in LPS-induced peritonitis in mice. J. Ethnopharmacol. 2014, 151, 144–150. [Google Scholar] [CrossRef]
- Meneses-Sagrero, S.E.; Navarro-Navarro, M.; Ruiz-Bustos, E.; Del-Toro-Sánchez, C.L.; Jiménez-Estrada, M.; Robles-Zepeda, R.E. Antiproliferative activity of spinasterol isolated of Stegnosperma halimifolium (Benth, 1844). Saudi Pharm. J. 2017, 25, 1137–1143. [Google Scholar] [CrossRef]
- Villaseñor, I.M.; Domingo, A.P. Anticarcinogenicity potential of spinasterol isolated from squash flowers. Teratog. Carcinog. Mutagen. 2000, 20, 99–105. [Google Scholar] [CrossRef]
- El Kharrassi, Y.; Samadi, M.; Lopez, T.; Nury, T.; El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; Vamecq, J.; Moustaid, K.; Latruffe, N.; et al. Biological activities of schottenol and spinasterol, two natural phytosterols present in argan oil and in cactus pear seed oil, on murine miroglial BV2 cells. Biochem. Biophys. Res. Commun. 2014, 446, 798–804. [Google Scholar] [CrossRef]
- Vijayasekhar, V.E.; Prasad, M.S.; Joshi, D.S.D.S.; Narendra, K.; Satya, A.K.; Rao, K.R.S.S. Evaluation of phytochemicals and in vitro pharmacological activity of Acacia sinuata plant leaves. Int. J. Pharm. Sci. Res. 2016, 7, 2423–2434. [Google Scholar] [CrossRef]
- Cordeiro, N.; Freitas, N.; Faria, M.; Gouveia, M. Ipomoea batatas (L.) Lam.: A rich source of lipophilic phytochemicals. J. Agric. Food Chem. 2013, 61, 12380–12384. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Ching, C.; Wu, T.-S.; Wu, C.-R.; Hsieh, W.-T.; Tsai, H.-Y. Balanophora spicata and lupeol acetate possess antinociceptive and anti-inflammatory activities in vivo and in vitro. Evid. Based Complement. Altern. Med. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lucetti, D.L.; Lucetti, E.C.P.; Bandeira, M.A.M.; Veras, H.N.H.; Silva, A.H.; Leal, L.K.A.M.; Lopes, A.A.; Alves, V.C.C.; Silva, G.S.; Brito, G.A.; et al. Anti-inflammatory effects and possible mechanism of action of lupeol acetate isolated from Himatanthus drasticus (Mart). Plumel. J. Inflamm. 2010, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, G.-C.; Park, M.-S.; Yoon, D.-Y.; Shin, C.-H.; Sin, H.-S.; Um, S.-J. Antitumor activity of spinasterol isolated from Pueraria roots. Exp. Mol. Med. 2005, 37, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Klein, L.C.; Gandolfi, R.B.; Santin, J.R.; Lemos, M.; Filho, V.C.; De Andrade, S.F. Antiulcerogenic activity of extract, fractions, and some compounds obtained from Polygala cyparissias St. Hillaire & Moquin (Polygalaceae). Naunyn. Schmiedebergs. Arch. Pharmacol. 2010, 381, 121–126. [Google Scholar] [CrossRef]
- Ardenghi, J.V.; Pretto, J.B.; Souza, M.M.; Junior, A.C.; Soldi, C.; Pizzolatti, M.G.; Meotti, F.C.; Moura, J.; d‘Ávila; Santos, A.R.S. Antinociceptive properties of coumarins, steroid and dihydrostyryl-2-pyrones from Polygala sabulosa (Polygalaceae) in mice. J. Pharm. Pharmacol. 2006, 58, 107–112. [Google Scholar] [CrossRef]
- Jeong, G.-S.; Li, B.; Lee, D.-S.; Kim, K.H.; Lee, I.K.; Lee, K.R.; Kim, Y.-C. Cytoprotective and anti-inflammatory effects of spinasterol via the induction of heme oxygenase-1 in murine hippocampal and microglial cell lines. Int. Immunopharmacol. 2010, 10, 1587–1594. [Google Scholar] [CrossRef]
- Luís, A.; Gil, N.; Amaral, M.E.; Duarte, A.P. Antioxidant activities of extracts from Acacia melanoxylon, Acacia dealbata and Olea europaea and alkaloids estimation. Int. J. Pharm. Pharm. Sci. 2012, 4, 225–231. [Google Scholar]
- Johnson, W. Final report of the safety assessment of Acacia Catechu gum, Acacia Concinna fruit extract, Acacia dealbata leaf extract, Acacia dealbata leaf wax, Acacia decurrens extract, Acacia farnesiana extract, Acacia farnesiana flower wax, Acacia farnesiana gum, Acacia senegal extract, Acacia senegal gum, and Acacia Senegal gum extract. Int. J. Toxicol. 2005, 24, 75–118. [Google Scholar] [CrossRef]
- Mohammad Alharbi, W.D.; Azmat, A. Anticonvulsant and neuroprotective effects of the Acacia tortilis growing in KSA. Pak. J. Pharm. Sci. 2015, 28, 531–534. [Google Scholar]
- Stohs, S.J.; Bagchi, D. Antioxidant, anti-inflammatory, and phemoprotective properties of Acacia catechu heartwood extracts. Phyther. Res. 2015, 29, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Freire, C.S.R.; Silvestre, A.J.D.; Neto, C.P. Identification of new hydroxy fatty acids and ferulic acid esters in the wood of Eucalyptus globulus. Holzforschung 2002, 56, 143–149. [Google Scholar] [CrossRef]
- Patinha, D.J.S.; Domingues, R.M.A.; Villaverde, J.J.; Silva, A.M.S.; Silva, C.M.; Freire, C.S.R.; Neto, C.P.; Silvestre, A.J.D. Lipophilic extractives from the bark of Eucalyptus grandis x globulus, a rich source of methyl morolate: Selective extraction with supercritical CO2. Ind. Crops Prod. 2013, 43, 340–348. [Google Scholar] [CrossRef]
- Ramos, P.A.B.; Moreirinha, C.; Santos, S.A.O.; Almeida, A.; Freire, C.S.R.; Silva, A.M.S.; Silvestre, A.J.D. Valorisation of bark lipophilic fractions from three Portuguese Salix species: A systematic study of the chemical composition and inhibitory activity on Escherichia coli. Ind. Crop. Prod. 2019, 132, 245–252. [Google Scholar] [CrossRef]
- Ramos, P.A.B.; Guerra, A.R.; Guerreiro, O.; Freire, C.S.R.; Silva, A.M.S.; Duarte, M.F.; Silvestre, A.J.D. Lipophilic extracts of Cynara cardunculus L. var. altilis (DC): A source of valuable bioactive terpenic compounds. J. Agric. Food Chem. 2013, 61, 8420–8429. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt(min) | Compound | mg g−1 of Extract | mg kg−1 of dw | ||||
|---|---|---|---|---|---|---|---|
| Bark | Wood | Leaves | Bark | Wood | Leaves | ||
| Fatty acids | 46.2 | 95.1 | 28.3 | 1060.0 | 290.0 | 1747.4 | |
| Saturated fatty acids | 41.2 | 55.4 | 23.0 | 946.1 | 168.7 | 1415.9 | |
| 24.12 | Dodecanoic acid | 0.1 | 0.3 | 0.2 | 2.9 | 0.9 | 12.9 |
| 29.26 | Tetradecanoic acid | 0.1 | 0.3 | 0.4 | 2.7 | 0.9 | 24.0 |
| 31.65 | Pentadecanoic acid | 0.1 | 0.5 | n.d. | 1.7 | 1.4 | n.d. |
| 33.95 | Hexadecanoic acid | 5.4 | 22.9 | 6.3 | 124.3 | 69.7 | 389.1 |
| 36.15 | Heptadecanoic acid | 0.1 | 1.1 | 0.1 | 3.0 | 3.3 | 7.2 |
| 38.26 | Octadecanoic acid | 0.8 | 8.4 | 1.0 | 17.4 | 25.6 | 59.4 |
| 40.28 | Nonadecanoic acid | 0.1 | 0.6 | n.d. | 1.4 | 1.7 | n.d. |
| 42.25 | Eicosanoic acid | 0.4 | 2.3 | 0.7 | 8.4 | 6.9 | 41.5 |
| 44.14 | Heneicosanoic acid | 0.2 | 1.6 | 0.3 | 3.5 | 5.0 | 21.4 |
| 45.96 | Docosanoic acid | 1.6 | 4.6 | 0.9 | 36.5 | 14.1 | 55.1 |
| 47.73 | Tricosanoic acid | 0.5 | 2.8 | 0.6 | 12.6 | 8.5 | 37.2 |
| 49.43 | Tetracosanoic acid | 6.1 | 6.2 | 1.3 | 141.0 | 18.8 | 80.0 |
| 51.10 | Pentacosanoic acid | 0.6 | 1.7 | n.d. | 13.9 | 5.1 | n.d. |
| 52.90 | Hexacosanoic acid | 3.7 | 2.2 | 1.4 | 85.9 | 6.6 | 86.3 |
| 54.80 | Heptacosanoic acid | 1.1 | n.d. | n.d. | 24.5 | n.d. | n.d. |
| 56.83 | Octacosanoic acid | 5.3 | n.d. | 5.4 | 120.5 | n.d. | 334.1 |
| 58.87 | Nonacosanoic acid | 3.1 | n.d. | n.d. | 72.0 | n.d. | n.d. |
| 61.10 | Triacontanoic acid | 8.2 | n.d. | 4.3 | 188.9 | n.d. | 267.5 |
| 65.98 | Dotriacontanoic acid | 3.7 | n.d. | n.d. | 84.9 | n.d. | n.d. |
| Unsaturated fatty acids | 4.2 | 38.7 | 5.4 | 95.7 | 118.0 | 331.5 | |
| 33.34 | Hexadec-9-enoic acid | 0.1 | 0.2 | 0.1 | 1.3 | 0.7 | 6.1 |
| 37.38 | Octadeca-9,12-dienoic acid | 1.7 | 31.9 | 1.7 | 39.6 | 97.2 | 104.4 |
| 37.42 | Octadeca-9,12,15-trienoic acid | 0.4 | 1.2 | 1.9 | 9.4 | 3.7 | 116.2 |
| 37.58 | cis-Octadec-9-enoic acid | 1.6 | 4.1 | 1.5 | 36.3 | 12.6 | 89.6 |
| 37.73 | trans-Octadec-9-enoic acid | 0.4 | 1.3 | 0.2 | 9.2 | 3.8 | 15.1 |
| w-Hydroxyacids | 0.8 | 1.1 | n.d | 18.2 | 3.3 | n.d. | |
| 52.01 | 22-Hydroxydocosanoic | 0.8 | 1.1 | n.d. | 18.2 | 3.3 | n.d. |
| Long-chain aliphatic alcohols | 47.2 | 14.2 | 30.7 | 1082.5 | 43.7 | 1891.5 | |
| 22.02 | Dodecan-1-ol | 0.01 | 0.5 | 0.02 | 0.3 | 1.4 | 1.4 |
| 24.73 | Tridecan-1-ol | 0.1 | 1.4 | 0.05 | 1.2 | 4.2 | 3.1 |
| 27.35 | Tetradecan-1-ol | 0.04 | 2.2 | n.d. | 0.9 | 6.8 | n.d. |
| 29.85 | Pentadecan-1-ol | 0.7 | 2.0 | n.d. | 15.9 | 6.2 | n.d. |
| 32.19 | Hexadecan-1-ol | 0.04 | 0.6 | 0.1 | 0.8 | 1.8 | 8.1 |
| 36.63 | Octadecan-1-ol | 0.1 | 0.4 | 0.1 | 2.0 | 1.4 | 3.1 |
| 40.71 | Eicosan-1-ol | 0.1 | n.d. | n.d. | 2.0 | n.d. | n.d. |
| 44.50 | Docosan-1-ol | 0.2 | 0.3 | n.d. | 4.3 | 1.2 | n.d. |
| 46.30 | Tricosan-1-ol | 0.2 | n.d. | n.d. | 3.7 | n.d. | n.d. |
| 48.05 | Tetracosan-1-ol | 3.8 | n.d. | n.d. | 86.5 | n.d. | n.d. |
| 49.73 | Pentacosan-1-ol | 0.9 | n.d. | n.d. | 19.8 | n.d. | n.d. |
| 51.43 | Hexacosan-1-ol | 11.6 | n.d. | 3.0 | 265.5 | n.d. | 184.1 |
| 53.21 | Heptacosan-1-ol | 1.5 | n.d. | n.d. | 34.3 | n.d. | n.d. |
| 55.15 | Octacosan-1-ol | 9.6 | 1.8 | 4.2 | 221.0 | 5.6 | 261.0 |
| 57.17 | Nonacosan-1-ol | 2.3 | n.d. | n.d. | 53.2 | n.d. | n.d. |
| 59.25 | Triacontan-1-ol | 10.8 | 5.0 | 16.8 | 247.9 | 15.2 | 1035.2 |
| 63.71 | Dotricontan-1-ol | 5.4 | n.d. | 6.4 | 123.3 | n.d. | 395.6 |
| Terpenic compounds | 150.5 | n.d. | 237.4 | 3450.8 | n.d. | 14,635.3 | |
| 29.00 | Neophytadiene | n.d. | n.d. | 2.6 | n.d. | n.d. | 158.8 |
| 37.02 | Phytol | n.d. | n.d. | 5.5 | n.d. | n.d. | 340.6 |
| 48.89 | Squalene | n.d. | n.d. | 28.4 | n.d. | n.d. | 1747.6 |
| 56.66 | β-Amyrone | n.d. | n.d. | 20.4 | n.d. | n.d. | 1256.5 |
| 57.57 | Lupenone | 56.4 | n.d. | 112.7 | 1293.7 | n.d. | 6946.8 |
| 58.36 | β-Amyrin | n.d. | n.d. | 26.1 | n.d. | n.d. | 1606.8 |
| 58.97 | α-Amyrin | n.d. | n.d. | 41.8 | n.d. | n.d. | 2578.2 |
| 60.52 | Lupenyl acetate | 94.0 | n.d. | n.d. | 2157.1 | n.d. | n.d. |
| Sterols | 21.1 | 193.7 | 10.3 | 484.2 | 590.4 | 635.7 | |
| 58.39 | Spinasterol | 8.6 | 94.6 | n.d. | 198.3 | 288.5 | n.d. |
| 58.69 | Sitostanol | n.d. | 7.0 | n.d. | n.d. | 21.2 | n.d. |
| 59.63 | 22,23-Dihydrospinasterol | 12.5 | 92.1 | 10.3 | 285.8 | 280.7 | 635.7 |
| Monoglycerides | 30.2 | 6.0 | 0.8 | 691.6 | 18.4 | 49.4 | |
| 45.14 | 1-Monohexadecenoin | n.d. | n.d. | 0.3 | n.d. | n.d. | 16.7 |
| 45.36 | 1-Monohexadecanoin | 0.1 | 2.8 | 0.5 | 3.4 | 8.4 | 32.7 |
| 47.55 | 2-Monolinolein | n.d. | 0.5 | n.d. | n.d. | 1.6 | n.d. |
| 48.13 | 1-Monolinolein | n.d. | 2.8 | n.d. | n.d. | 8.4 | n.d. |
| 55.92 | 1-Monodocosanoin | 6.5 | n.d. | n.d. | 149.5 | n.d. | n.d. |
| 60.06 | 1-Monotetracosanoin | 23.5 | n.d. | n.d. | 538.7 | n.d. | n.d. |
| Aromatic compounds | 1.3 | 8.8 | 1.8 | 29.0 | 26.9 | 112.0 | |
| Aromatic aldehydes | 0.2 | 5.8 | 0.1 | 4.4 | 17.6 | 9.1 | |
| 14.86 | 4-Hydroxybenzaldehyde | 0.1 | 0.3 | 0.1 | 1.3 | 0.8 | 7.3 |
| 19.75 | Vanillin | 0.1 | 2.2 | n.d. | 2.5 | 6.8 | n.d. |
| 24.42 | Syringaldehyde | 0.02 | 1.4 | 0.03 | 0.6 | 4.2 | 1.8 |
| 24.53 | 2,5-Hydroxybenzaldehyde | n.d. | 0.8 | n.d. | n.d. | 2.5 | n.d. |
| 28.02 | Coniferaldehyde | n.d. | 0.5 | n.d. | n.d. | 1.5 | n.d. |
| 31.89 | Sinapaldehyde | n.d. | 0.6 | n.d. | n.d. | 1.8 | n.d. |
| Aromatic acids | 0.7 | 2.1 | 0.8 | 15.9 | 6.4 | 47.7 | |
| 10.98 | Benzoic acid | n.d. | n.d. | 0.1 | n.d. | n.d. | 7.0 |
| 23.24 | p-Hydroxybenzoic acid | 0.1 | 0.04 | 0.1 | 1.3 | 0.1 | 8.4 |
| 26.85 | Vanillic acid | 0.2 | 1.2 | 0.2 | 5.6 | 3.8 | 12.4 |
| 26.97 | Homovanillic acid | 0.1 | 0.1 | n.d. | 1.2 | 0.3 | n.d. |
| 30.18 | Syringic acid | 0.3 | 0.7 | n.d. | 7.8 | 2.2 | n.d. |
| 31.09 | p-Coumaric acid | n.d. | n.d. | 0.3 | n.d. | n.d. | 19.8 |
| Other aromatic compounds | 0.4 | 1.0 | 0.9 | 8.8 | 2.9 | 55.3 | |
| 16.07 | Resorcinol | 0.1 | n.d. | 0.1 | 2.8 | n.d. | 6.8 |
| 21.75 | Tyrosol | 0.02 | 0.1 | 0.8 | 0.5 | 0.3 | 48.5 |
| 23.62 | Vanillyl alcohol | 0.2 | 0.4 | n.d. | 5.5 | 1.2 | n.d. |
| 27.76 | p-Coumaric alcohol | n.d. | 0.5 | n.d. | n.d. | 1.4 | n.d. |
| Others | 3.2 | 2.3 | 37.7 | 74.4 | 6.9 | 2323.2 | |
| 13.44 | Glycerol | 0.9 | 1.5 | 5.5 | 20.2 | 4.6 | 339.7 |
| 15.57 | trans-Erythronoic acid-γ-lactone | 0.2 | 0.5 | 0.4 | 4.2 | 1.6 | 21.6 |
| 17.01 | cis-Erythronoic acid-γ-lactone | 0.2 | 0.2 | 0.4 | 3.8 | 0.7 | 25.7 |
| 54.54 | α-Tocopherol | 2.0 | n.d. | 31.4 | 46.1 | n.d. | 1936.1 |
| Total | 299.6 | 320.2 | 347.1 | 6872.4 | 976.3 | 21,394.5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, C.S.D.; Moreira, P.; Resende, J.; Cruz, M.T.; Pereira, C.M.F.; Silva, A.M.S.; Santos, S.A.O.; Silvestre, A.J.D. Characterization and Cytotoxicity Assessment of the Lipophilic Fractions of Different Morphological Parts of Acacia dealbata. Int. J. Mol. Sci. 2020, 21, 1814. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051814
Oliveira CSD, Moreira P, Resende J, Cruz MT, Pereira CMF, Silva AMS, Santos SAO, Silvestre AJD. Characterization and Cytotoxicity Assessment of the Lipophilic Fractions of Different Morphological Parts of Acacia dealbata. International Journal of Molecular Sciences. 2020; 21(5):1814. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051814
Chicago/Turabian StyleOliveira, Cátia S. D., Patrícia Moreira, Judite Resende, Maria T. Cruz, Cláudia M. F. Pereira, Artur M. S. Silva, Sónia A. O. Santos, and Armando J. D. Silvestre. 2020. "Characterization and Cytotoxicity Assessment of the Lipophilic Fractions of Different Morphological Parts of Acacia dealbata" International Journal of Molecular Sciences 21, no. 5: 1814. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051814