Promoting Roles of Embryonic Signals in Embryo Implantation and Placentation in Cooperation with Endocrine and Immune Systems

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Main Factors Regulating Endometrial Receptivity for Embryo Implantation
1.1. Endocrine System and Embryo Signals
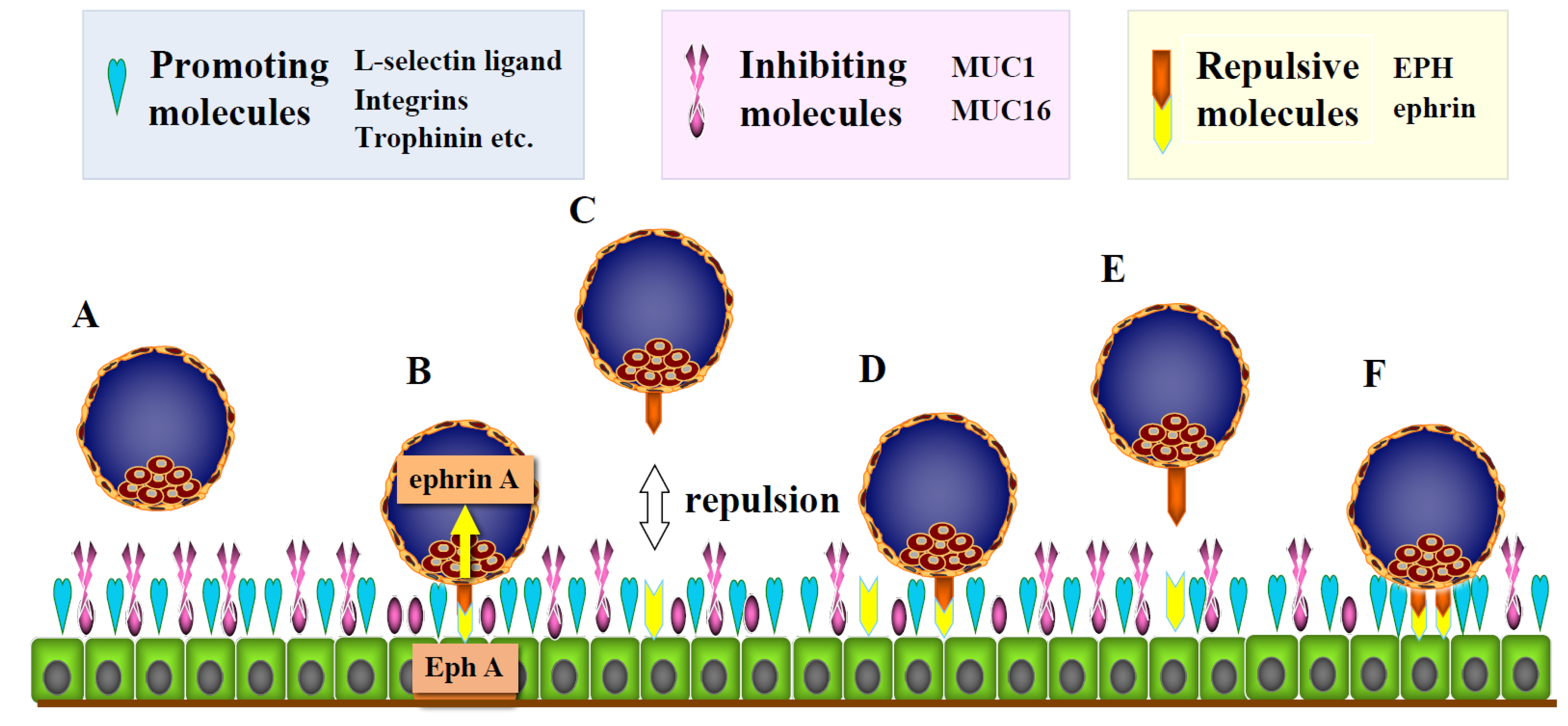
1.2. Adhesion-Promoting and -Inhibiting Molecules
1.3. Repulsive Molecules
2. Invasion Process of Human Embryos after Attachment to Endometrial Epithelial Cells
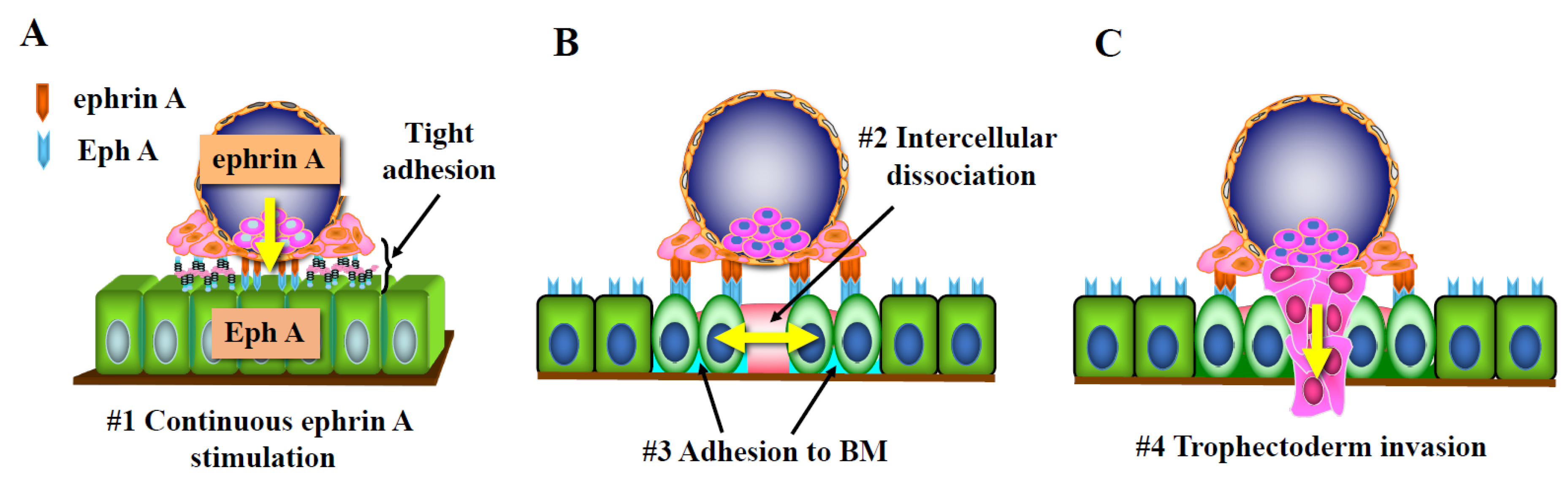
2.1. Activation of the Trophectoderm after Attachment and Acquisition of Invasive Properties
2.2. Opening of Tight Junctions in the Endometrial Epithelial Layer
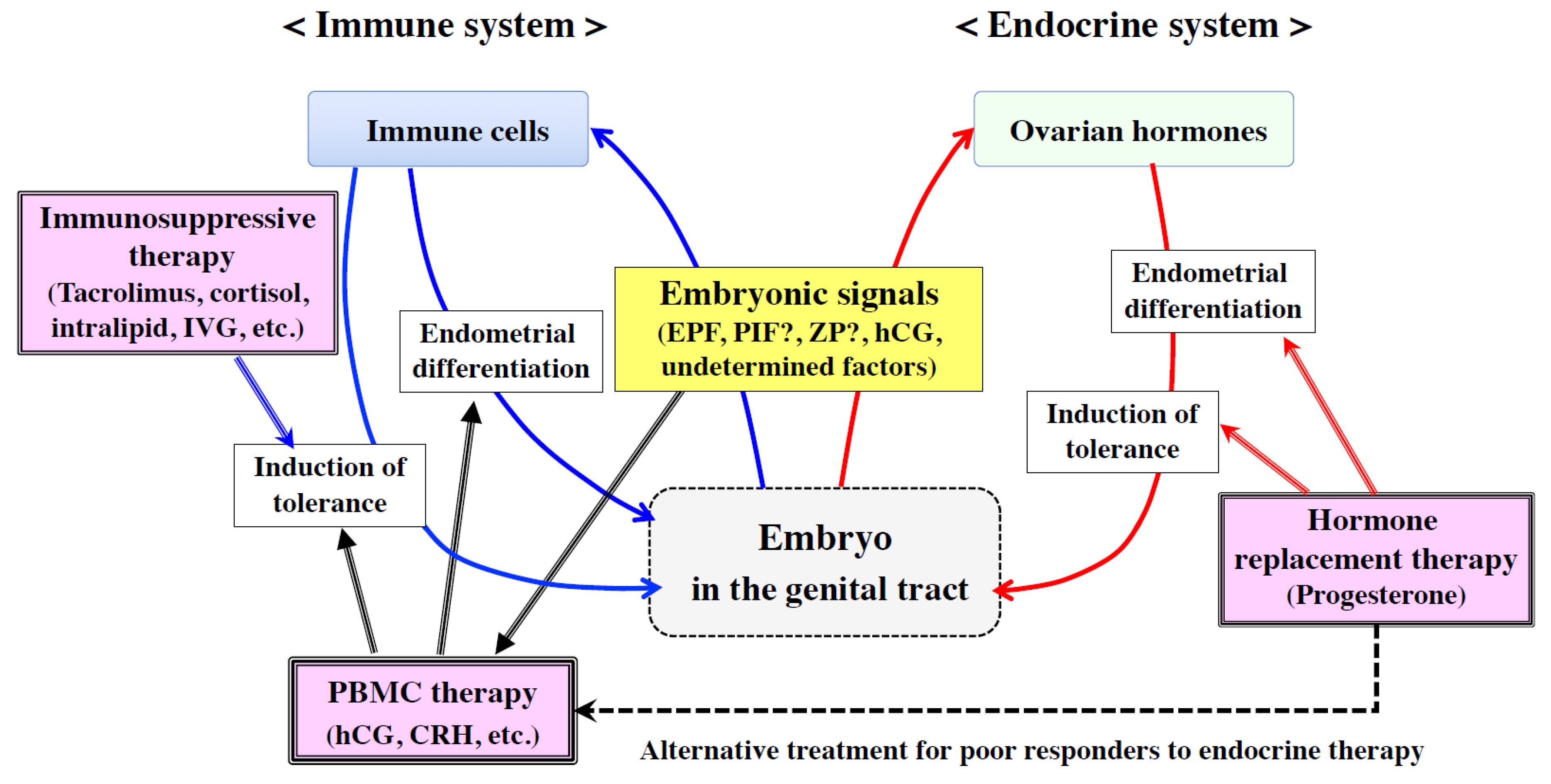
3. Positive Role of the Immune System in Embryo Implantation and Placentation
3.1. Regulation of Endometrial Receptivity and Embryo Invasion by Embryonic Signals
3.2. Regulation of Extravillous Trophoblast (EVT) Invasion by Embryonic Signals
4. Topics of Clinical Application of Immune Therapy
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALCAM | activated leukocyte cell adhesion molecule |
| BM | basement membrane |
| ECM | extracellular matrix |
| EMT | epithelial–mesenchymal transition |
| EVT | extravillous trophoblast |
| hCG | human chorionic gonadotropin |
| IGFBP-1 | insulin-like growth factor binding protein-1 |
| IL | interleukin |
| IVF-ET | in vitro fertilization and embryo transfer |
| LIF | leukemia inhibitory factor |
| M-CSF | macrophage colony stimulating factor |
| MMP | matrix metalloproteinase |
| mTOR | mammalian target of rapamycin |
| PBMC | peripheral blood mononuclear cell |
| PIF | pre-implantation factor |
| PTENP1 | phosphatase and tensin homolog pseudogene 1 |
| TIMP | tissue inhibitor of metalloproteinase |
| VCAM1 | vascular cell adhesion molecule 1 |
| VEGF | vascular endothelial growth factor |
| ZP | zona pellucida |
References
- Wetendorf, M.; DeMayo, F.J. The progesterone receptor regulates implantation, decidualization, and glandular development via a complex paracrine signaling network. Mol. Cell Endocrinol. 2012, 357, 108–118. [Google Scholar]
- Large, M.J.; DeMayo, F.J. The regulation of embryo implantation and endometrial decidualization by progesterone receptor signaling. Mol. Cell Endocrinol. 2012, 358, 155–165. [Google Scholar]
- Yoshinaga, K. Uterine receptivity for blastocyst implantation. Ann. N. Y. Acad. Sci. 1988, 541, 424–431. [Google Scholar]
- Sarantis, L.; Roche, D.; Psychoyos, A. Displacement of receptivity for nidation in the rat by the progesterone antagonist RU 486: A scanning electron microscopy study. Hum. Reprod. 1988, 3, 251–255. [Google Scholar]
- Psychoyos, A. The Implantation Window: BASIC and Clinical Aspects; Ares-Serono Symposia Publications: Roma, Italy, 1993; Volume 4, pp. 57–63. [Google Scholar]
- Fukui, Y.; Hirota, Y.; Matsuo, M.; Gebril, M.; Akaeda, S.; Hiraoka, T.; Osuga, Y. Uterine receptivity, embryo attachment, and embryo invasion: Multistep processes in embryo implantation. Reprod. Med. Biol. 2019, 18, 234–240. [Google Scholar]
- Edwards, R.G. Human uterine endocrinology and the implantation window. Ann. N. Y. Acad. Sci. 1988, 541, 445–454. [Google Scholar]
- Lessey, B.A. The role of the endometrium during embryo implantation. Hum. Reprod. 2000, 15 (Suppl 6), 39–50. [Google Scholar]
- Kosaka, K.; Fujiwara, H.; Tatsumi, K.; Yoshioka, S.; Higuchi, T.; Sato, Y.; Nakayama, T.; Fujii, S. Human peripheral blood mononuclear cells enhance cell-cell interaction between human endometrial epithelial cells and BeWo-cell spheroids. Hum. Reprod. 2003, 18, 19–25. [Google Scholar]
- Tur-Kaspa, I.; Confino, E.; Dudkiewicz, A.B.; Myers, S.A.; Friberg, J.; Gleicher, N. Ovarian stimulation protocol for in vitro fertilization with gonadotropin-releasing hormone agonist widens the implantation window. Fertil. Steril. 1990, 53, 859–864. [Google Scholar]
- Murphy, C.R. Understanding the apical surface markers of uterine receptivity: Pinopods-or uterodomes? Hum. Reprod. 2000, 15, 2451–2454. [Google Scholar]
- Quinn, K.E.; Matson, B.C.; Wetendorf, M.; Caron, K.M. Pinopodes: Recent advancements, current perspectives, and future directions. Mol. Cell Endocrinol. 2019, 501, 110644. [Google Scholar] [PubMed]
- Tabibzadeh, S.; Babaknia, A. The signals and molecular pathways involved in implantation, a symbiotic interaction between blastocyst and endometrium involving adhesion and tissue invasion. Hum. Reprod. 1995, 10, 1579–1602. [Google Scholar] [PubMed]
- Simon, C.; Dominguez, F.; Remohi, J.; Pellicer, A. Embryo effects in human implantation: Embryonic regulation of endometrial molecules in human implantation. Ann. N. Y. Acad. Sci. 2001, 943, 1–16. [Google Scholar]
- Daikoku, T.; Cha, J.; Sun, X.; Tranguch, S.; Xie, H.; Fujita, T.; Hirota, Y.; Lydon, J.; DeMayo, F.; Maxson, R.; et al. Conditional deletion of Msx homeobox genes in the uterus inhibits blastocyst implantation by altering uterine receptivity. Dev. Cell 2011, 21, 1014–1025. [Google Scholar]
- Namiki, T.; Ito, J.; Kashiwazaki, N. Molecular mechanisms of embryonic implantation in mammals: Lessons from the gene manipulation of mice. Reprod. Med. Biol. 2018, 17, 331–342. [Google Scholar]
- Fazleabas, A.T.; Kim, J.J.; Strakova, Z. Implantation: Embryonic signals and the modulation of the uterine environment—A review. Placenta 2004, 25 (Suppl. A), S26–S31. [Google Scholar]
- Rao, C.V.; Lei, Z.M. The past, present and future of nongonadal LH/hCG actions in reproductive biology and medicine. Mol. Cell Endocrinol. 2007, 269, 2–8. [Google Scholar]
- Fazleabas, A.T.; Donnelly, K.M.; Srinivasan, S.; Fortman, J.D.; Miller, J.B. Modulation of the baboon (Papio anubis) uterine endometrium by chorionic gonadotrophin during the period of uterine receptivity. Proc. Natl. Acad. Sci. USA 1999, 96, 2543–2548. [Google Scholar]
- Licht, P.; Russu, V.; Wildt, L. On the role of human chorionic gonadotropin (hCG) in the embryo-endometrial microenvironment: Implications for differentiation and implantation. Semin. Reprod. Med. 2001, 19, 37–47. [Google Scholar]
- Licht, P.; Fluhr, H.; Neuwinger, J.; Wallwiener, D.; Wildt, L. Is human chorionic gonadotropin directly involved in the regulation of human implantation? Mol. Cell Endocrinol. 2007, 269, 85–92. [Google Scholar]
- Evans, J. Hyperglycosylated hCG: A Unique Human Implantation and Invasion Factor. Am. J. Reprod. Immunol. 2016, 75, 333–340. [Google Scholar] [PubMed] [Green Version]
- Makrigiannakis, A.; Vrekoussis, T.; Zoumakis, E.; Kalantaridou, S.N.; Jeschke, U. The Role of HCG in Implantation: A Mini-Review of Molecular and Clinical Evidence. Int. J. Mol. Sci. 2017, 18, 1305. [Google Scholar]
- Cuman, C.; Van Sinderen, M.; Gantier, M.P.; Rainczuk, K.; Sorby, K.; Rombauts, L.; Osianlis, T.; Dimitriadis, E. Human Blastocyst Secreted microRNA Regulate Endometrial Epithelial Cell Adhesion. EBioMedicine 2015, 2, 1528–1535. [Google Scholar] [PubMed] [Green Version]
- Es-Haghi, M.; Godakumara, K.; Haling, A.; Lattekivi, F.; Lavrits, A.; Viil, J.; Andronowska, A.; Nafee, T.; James, V.; Jaakma, U.; et al. Specific trophoblast transcripts transferred by extracellular vesicles affect gene expression in endometrial epithelial cells and may have a role in embryo-maternal crosstalk. Cell Commun. Signal. 2019, 17, 146. [Google Scholar] [PubMed] [Green Version]
- Takamura, M.; Zhou, W.; Rombauts, L.; Dimitriadis, E. The long noncoding RNA PTENP1 regulates human endometrial epithelial adhesive capacity in vitro: Implications in infertility. Biol. Reprod. 2020, 102, 53–62. [Google Scholar]
- Sugihara, K.; Sugiyama, D.; Byrne, J.; Wolf, D.P.; Lowitz, K.P.; Kobayashi, Y.; Kabir-Salmani, M.; Nadano, D.; Aoki, D.; Nozawa, S.; et al. Trophoblast cell activation by trophinin ligation is implicated in human embryo implantation. Proc. Natl. Acad. Sci. USA 2007, 104, 3799–3804. [Google Scholar]
- Fukuda, M.N.; Sugihara, K. Signal transduction in human embryo implantation. Cell Cycle 2007, 6, 1153–1156. [Google Scholar]
- Fukuda, M.N.; Sugihara, K. An integrated view of L-selectin and trophinin function in human embryo implantation. J. Obstet. Gynaecol. Res. 2008, 34, 129–136. [Google Scholar]
- Genbacev, O.D.; Prakobphol, A.; Foulk, R.A.; Krtolica, A.R.; Ilic, D.; Singer, M.S.; Yang, Z.Q.; Kiessling, L.L.; Rosen, S.D.; Fisher, S.J. Trophoblast L-selectin-mediated adhesion at the maternal-fetal interface. Science 2003, 299, 405–408. [Google Scholar]
- Lai, T.H.; Zhao, Y.; Shih Ie, M.; Ho, C.L.; Bankowski, B.; Vlahos, N. Expression of L-selectin ligands in human endometrium during the implantation window after controlled ovarian stimulation for oocyte donation. Fertil. Steril. 2006, 85, 761–763. [Google Scholar]
- Foulk, R.A.; Zdravkovic, T.; Genbacev, O.; Prakobphol, A. Expression of L-selectin ligand MECA-79 as a predictive marker of human uterine receptivity. J. Assist. Reprod. Genet. 2007, 24, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Ma, X.; Deng, L.; Yao, B.; Xiong, Y.; Wu, Y.; Wang, L.; Ma, Q.; Ma, F. Role of selectins and their ligands in human implantation stage. Glycobiology 2017, 27, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessey, B.A.; Castelbaum, A.J.; Sawin, S.W.; Sun, J. Integrins as markers of uterine receptivity in women with primary unexplained infertility. Fertil. Steril. 1995, 63, 535–542. [Google Scholar] [CrossRef]
- Lessey, B.A.; Arnold, J.T. Paracrine signaling in the endometrium: Integrins and the establishment of uterine receptivity. J. Reprod. Immunol. 1998, 39, 105–116. [Google Scholar] [CrossRef]
- Chung, T.W.; Park, M.J.; Kim, H.S.; Choi, H.J.; Ha, K.T. Integrin alphaVbeta3 and alphaVbeta5 are required for leukemia inhibitory factor-mediated the adhesion of trophoblast cells to the endometrial cells. Biochem. Biophys. Res. Commun. 2016, 469, 936–940. [Google Scholar] [PubMed]
- Elnaggar, A.; Farag, A.H.; Gaber, M.E.; Hafeez, M.A.; Ali, M.S.; Atef, A.M. AlphaVBeta3 Integrin expression within uterine endometrium in unexplained infertility: A prospective cohort study. BMC Women’s Health 2017, 17, 90. [Google Scholar] [CrossRef] [Green Version]
- Aplin, J.D.; Seif, M.W.; Graham, R.A.; Hey, N.A.; Behzad, F.; Campbell, S. The endometrial cell surface and implantation. Expression of the polymorphic mucin MUC-1 and adhesion molecules during the endometrial cycle. Ann. N. Y. Acad. Sci. 1994, 734, 103–121. [Google Scholar] [CrossRef]
- Albers, A.; Thie, M.; Hohn, H.P.; Denker, H.W. Differential expression and localization of integrins and CD44 in the membrane domains of human uterine epithelial cells during the menstrual cycle. Acta Anat. (Basel) 1995, 153, 12–19. [Google Scholar] [CrossRef]
- Berneau, S.C.; Ruane, P.T.; Brison, D.R.; Kimber, S.J.; Westwood, M.; Aplin, J.D. Investigating the role of CD44 and hyaluronate in embryo-epithelial interaction using an in vitro model. Mol. Hum. Reprod. 2019, 25, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Gimeno, M.J.; Mercader, A.; O’Connor, J.E.; Remohi, J.; Polan, M.L.; Pellicer, A. Embryonic regulation of integrins beta 3, alpha 4, and alpha 1 in human endometrial epithelial cells in vitro. J. Clin. Endocrinol. Metab. 1997, 82, 2607–2616. [Google Scholar]
- Takahashi, H.; Takizawa, T.; Matsubara, S.; Ohkuchi, A.; Kuwata, T.; Usui, R.; Matsumoto, H.; Sato, Y.; Fujiwara, H.; Okamoto, A.; et al. Extravillous trophoblast cell invasion is promoted by the CD44-hyaluronic acid interaction. Placenta 2014, 35, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Horie, A.; Tani, H.; Ueda, M.; Okunomiya, A.; Suginami, K.; Kondoh, E.; Baba, T.; Konishi, I.; Shinomura, T.; et al. Versican V1 in human endometrial epithelial cells promotes BeWo spheroid adhesion in vitro. Reproduction 2019, 157, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, R.; Bai, H.; Kuse, M.; Ideta, A.; Aoyagi, Y.; Fujiwara, H.; Okuda, K.; Imakawa, K.; Sakurai, T. Involvement of VCAM1 in the bovine conceptus adhesion to the uterine endometrium. Reproduction 2014, 148, 119–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.R.; Inoue, T.; Ueda, M.; Hirano, T.; Higuchi, T.; Maeda, M.; Konishi, I.; Fujiwara, H.; Fujii, S. CD9 is expressed on human endometrial epithelial cells in association with integrins alpha(6), alpha(3) and beta(1). Mol. Hum. Reprod. 2000, 6, 252–257. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.M.; Cao, Y.J.; Yang, Y.J.; Li, J.; Hu, Z.; Duan, E.K. Tetraspanin CD9 regulates invasion during mouse embryo implantation. J. Mol. Endocrinol. 2006, 36, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, F.; Simon, C.; Quinonero, A.; Ramirez, M.A.; Gonzalez-Munoz, E.; Burghardt, H.; Cervero, A.; Martinez, S.; Pellicer, A.; Palacin, M.; et al. Human endometrial CD98 is essential for blastocyst adhesion. PLoS ONE 2010, 5, e13380. [Google Scholar] [CrossRef]
- Iwai, M.; Hamatani, T.; Nakamura, A.; Kawano, N.; Kanai, S.; Kang, W.; Yoshii, N.; Odawara, Y.; Yamada, M.; Miyamoto, Y.; et al. Membrane protein CD9 is repositioned and released to enhance uterine function. Lab. Investig. 2019, 99, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Higuchi, T.; Katsuragawa, H.; Inoue, T.; Kataoka, N.; Park, K.R.; Ueda, M.; Maeda, M.; Fujiwara, H.; Fujii, S. CD9 is involved in invasion of human trophoblast-like choriocarcinoma cell line, BeWo cells. Mol. Hum. Reprod. 1999, 5, 168–174. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Higuchi, T.; Ueda, M.; Inoue, T.; Kataoka, N.; Maeda, M.; Fujiwara, H.; Fujii, S. CD9 is expressed in extravillous trophoblasts in association with integrin alpha3 and integrin alpha5. Mol. Hum. Reprod. 1999, 5, 162–167. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Sato, Y.; Horie, A.; Suginami, K.; Tani, H.; Hattori, A.; Araki, Y.; Kagami, K.; Konishi, I.; Fujiwara, H. CD9 suppresses human extravillous trophoblast invasion. Placenta 2016, 47, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Hey, N.A.; Graham, R.A.; Seif, M.W.; Aplin, J.D. The polymorphic epithelial mucin MUC1 in human endometrium is regulated with maximal expression in the implantation phase. J. Clin. Endocrinol. Metab. 1994, 78, 337–342. [Google Scholar] [PubMed]
- Aplin, J.D.; Hey, N.A.; Graham, R.A. Human endometrial MUC1 carries keratan sulfate: Characteristic glycoforms in the luminal epithelium at receptivity. Glycobiology 1998, 8, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meseguer, M.; Aplin, J.D.; Caballero-Campo, P.; O’Connor, J.E.; Martin, J.C.; Remohi, J.; Pellicer, A.; Simon, C. Human endometrial mucin MUC1 is up-regulated by progesterone and down-regulated in vitro by the human blastocyst. Biol. Reprod. 2001, 64, 590–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gipson, I.K.; Blalock, T.; Tisdale, A.; Spurr-Michaud, S.; Allcorn, S.; Stavreus-Evers, A.; Gemzell, K. MUC16 is lost from the uterodome (pinopode) surface of the receptive human endometrium: In vitro evidence that MUC16 is a barrier to trophoblast adherence. Biol. Reprod. 2008, 78, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paria, B.C.; Reese, J.; Das, S.K.; Dey, S.K. Deciphering the cross-talk of implantation: Advances and challenges. Science 2002, 296, 2185–2188. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, Y.; Elad, D.; Jaffa, A.J.; Cao, Y.; Ye, X.; Duan, E. Navigating the site for embryo implantation: Biomechanical and molecular regulation of intrauterine embryo distribution. Mol. Aspects Med. 2013, 34, 1024–1042. [Google Scholar] [CrossRef]
- Imakawa, K.; Bai, R.; Kusama, K. Integration of molecules to construct the processes of conceptus implantation to the maternal endometrium. J. Anim. Sci. 2018, 96, 3009–3021. [Google Scholar] [CrossRef]
- Sanchez, J.M.; Mathew, D.J.; Passaro, C.; Fair, T.; Lonergan, P. Embryonic maternal interaction in cattle and its relationship with fertility. Reprod. Domest. Anim. 2018, 53 (Suppl. 2), 20–27. [Google Scholar] [CrossRef]
- Simintiras, C.A.; Sanchez, J.M.; McDonald, M.; Lonergan, P. The biochemistry surrounding bovine conceptus elongationdagger. Biol. Reprod. 2019, 101, 328–337. [Google Scholar] [CrossRef]
- Yoshinaga, K.; Adams, C.E. Delayed implantation in the spayed, progesterone treated adult mouse. J. Reprod. Fertil. 1966, 12, 593–595. [Google Scholar] [CrossRef] [Green Version]
- Hamatani, T.; Daikoku, T.; Wang, H.; Matsumoto, H.; Carter, M.G.; Ko, M.S.; Dey, S.K. Global gene expression analysis identifies molecular pathways distinguishing blastocyst dormancy and activation. Proc. Natl. Acad. Sci. USA 2004, 101, 10326–10331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, H.; Yoshioka, S.; Tatsumi, K.; Kosaka, K.; Satoh, Y.; Nishioka, Y.; Egawa, M.; Higuchi, T.; Fujii, S. Human endometrial epithelial cells express ephrin A1: Possible interaction between human blastocysts and endometrium via Eph-ephrin system. J. Clin. Endocrinol. Metab. 2002, 87, 5801–5807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquale, E.B. Eph receptor signalling casts a wide net on cell behaviour. Nat. Rev. Mol. Cell Biol. 2005, 6, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Kania, A.; Klein, R. Mechanisms of ephrin-Eph signalling in development, physiology and disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Darling, T.K.; Lamb, T.J. Emerging Roles for Eph Receptors and Ephrin Ligands in Immunity. Front. Immunol. 2019, 10, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.H.; Klein, R. Eph receptors and ephrin ligands. essential mediators of vascular development. Trends. Cardiovasc. Med. 2000, 10, 183–188. [Google Scholar] [CrossRef]
- Wolf, K.; Hu, H.; Isaji, T.; Dardik, A. Molecular identity of arteries, veins, and lymphatics. J. Vasc. Surg. 2019, 69, 253–262. [Google Scholar] [CrossRef]
- Klein, R. Bidirectional modulation of synaptic functions by Eph/ephrin signaling. Nat. Neurosci. 2009, 12, 15–20. [Google Scholar] [CrossRef]
- Fujii, H.; Tatsumi, K.; Kosaka, K.; Yoshioka, S.; Fujiwara, H.; Fujii, S. Eph-ephrin A system regulates murine blastocyst attachment and spreading. Dev. Dyn. 2006, 235, 3250–3258. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Fu, J.; Ren, Q.; Chen, X.; Wang, A. Expression of Eph A molecules during swine embryo implantation. Mol. Biol. Rep. 2012, 39, 2179–2185. [Google Scholar] [CrossRef]
- Fu, Y.; Li, L.; Fang, X.; Li, B.; Zhao, W.; Zhou, L.; Ren, S. Investigation of Eph-ephrin A1 in the regulation of embryo implantation in sows. Reprod. Domest. Anim. 2018, 53, 1563–1574. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Knox, R.V.; Li, L.; Ren, S. Differential gene expression of Eph-ephrin A1 and LEPR-LEP with high or low number of embryos in pigs during implantation. Reprod. Domest. Anim. 2018, 53, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, H.; Tatsumi, K.; Kosaka, K.; Sato, Y.; Higuchi, T.; Yoshioka, S.; Maeda, M.; Ueda, M.; Fujii, S. Human blastocysts and endometrial epithelial cells express activated leukocyte cell adhesion molecule (ALCAM/CD166). J. Clin. Endocrinol. Metab. 2003, 88, 3437–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, M.A.; Aruffo, A.A.; Bajorath, J. Cell surface receptors and their ligands: In vitro analysis of CD6-CD166 interactions. Proteins 2000, 40, 420–428. [Google Scholar] [CrossRef]
- Swart, G.W. Activated leukocyte cell adhesion molecule (CD166/ALCAM): Developmental and mechanistic aspects of cell clustering and cell migration. Eur. J. Cell Biol. 2002, 81, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Gilsanz, A.; Sanchez-Martin, L.; Gutierrez-Lopez, M.D.; Ovalle, S.; Machado-Pineda, Y.; Reyes, R.; Swart, G.W.; Figdor, C.G.; Lafuente, E.M.; Cabanas, C. ALCAM/CD166 adhesive function is regulated by the tetraspanin CD9. Cell Mol. Life Sci. 2013, 70, 475–493. [Google Scholar] [CrossRef]
- Haouzi, D.; Dechaud, H.; Assou, S.; Monzo, C.; de Vos, J.; Hamamah, S. Transcriptome analysis reveals dialogues between human trophectoderm and endometrial cells during the implantation period. Hum. Reprod. 2011, 26, 1440–1449. [Google Scholar] [CrossRef] [Green Version]
- Zen, K.; Parkos, C.A. Leukocyte-epithelial interactions. Curr. Opin. Cell Biol. 2003, 15, 557–564. [Google Scholar] [CrossRef]
- Agace, W.W.; Higgins, J.M.; Sadasivan, B.; Brenner, M.B.; Parker, C.M. T-lymphocyte-epithelial-cell interactions: Integrin alpha(E)(CD103)beta(7), LEEP-CAM and chemokines. Curr. Opin. Cell Biol. 2000, 12, 563–568. [Google Scholar] [CrossRef]
- Luissint, A.C.; Parkos, C.A.; Nusrat, A. Inflammation and the Intestinal Barrier: Leukocyte-Epithelial Cell Interactions, Cell Junction Remodeling, and Mucosal Repair. Gastroenterology 2016, 151, 616–632. [Google Scholar] [CrossRef] [Green Version]
- Matthews, J.D.; Weight, C.M.; Parkos, C.A. Leukocyte-epithelial interactions and mucosal homeostasis. Toxicol. Pathol. 2014, 42, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rescigno, M.; Urbano, M.; Valzasina, B.; Francolini, M.; Rotta, G.; Bonasio, R.; Granucci, F.; Kraehenbuhl, J.P.; Ricciardi-Castagnoli, P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat. Immunol. 2001, 2, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Akyaw, A.; Krishnamoorthy, K.; Goldsmith, L.T.; Morelli, S.S. The role of mesenchymal-epithelial transition in endometrial function. Hum. Reprod. Update 2019, 25, 114–133. [Google Scholar] [CrossRef] [PubMed]
- Imakawa, K.; Bai, R.; Fujiwara, H.; Ideta, A.; Aoyagi, Y.; Kusama, K. Continuous model of conceptus implantation to the maternal endometrium. J. Endocrinol. 2017, 233, R53–R65. [Google Scholar] [CrossRef] [Green Version]
- Imakawa, K.; Bai, R.; Fujiwara, H.; Kusama, K. Conceptus implantation and placentation: Molecules related to epithelial-mesenchymal transition, lymphocyte homing, endogenous retroviruses, and exosomes. Reprod. Med. Biol. 2016, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Uchida, H.; Maruyama, T.; Nishikawa-Uchida, S.; Oda, H.; Miyazaki, K.; Yamasaki, A.; Yoshimura, Y. Studies using an in vitro model show evidence of involvement of epithelial-mesenchymal transition of human endometrial epithelial cells in human embryo implantation. J. Biol. Chem. 2012, 287, 4441–4450. [Google Scholar] [CrossRef] [Green Version]
- Yoshinaga, K. Review of factors essential for blastocyst implantation for their modulating effects on the maternal immune system. Semin. Cell Dev. Biol. 2008, 19, 161–169. [Google Scholar] [CrossRef]
- Fujii, H.; Fujiwara, H.; Horie, A.; Sato, Y.; Konishi, I. Ephrin A1 induces intercellular dissociation in Ishikawa cells: Possible implication of the Eph-ephrin A system in human embryo implantation. Hum. Reprod 2011, 26, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Kamata, R.; Sakai, R. EphA2 phosphorylates the cytoplasmic tail of Claudin-4 and mediates paracellular permeability. J. Biol. Chem. 2005, 280, 42375–42382. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Liu, Z.; Peng, W.; Gao, Z.; Ouyang, H.; Yan, T.; Ding, S.; Cai, Z.; Zhao, B.; Mao, L.; et al. Activation of EphA4 induced by EphrinA1 exacerbates disruption of the blood-brain barrier following cerebral ischemia-reperfusion via the Rho/ROCK signaling pathway. Exp. Ther. Med. 2018, 16, 2651–2658. [Google Scholar] [CrossRef]
- Carter, N.; Nakamoto, T.; Hirai, H.; Hunter, T. EphrinA1-induced cytoskeletal re-organization requires FAK and p130(cas). Nat. Cell Biol. 2002, 4, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Fujiwara, H.; Horie, A.; Suginami, K.; Sato, Y.; Konishi, I. EphrinA1 stimulates cell attachment and inhibits cell aggregation through the EphA receptor pathway in human endometrial carcinoma-derived Ishikawa cells. Hum. Reprod. 2011, 26, 1163–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.; Bae, H.; Bazer, F.W.; Song, G. Functional Roles of Eph A-Ephrin A1 System in Endometrial Luminal Epithelial Cells During Early Pregnancy. J. Cell Physiol. 2017, 232, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, H.; Nishioka, Y.; Matsumoto, H.; Suginami, K.; Horie, A.; Tani, H.; Matsumura, N.; Baba, T.; Sato, Y.; Araki, Y.; et al. Eph-ephrin A system regulates human choriocarcinoma-derived JEG-3 cell invasion. Int. J. Gynecol. Cancer 2013, 23, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Min, J. Effect of ephrin-A1/EphA2 on invasion of trophoblastic cells. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2011, 31, 824–827. [Google Scholar] [CrossRef]
- Simon, C.; Frances, A.; Piquette, G.; Hendrickson, M.; Milki, A.; Polan, M.L. Interleukin-1 system in the materno-trophoblast unit in human implantation: Immunohistochemical evidence for autocrine/paracrine function. J. Clin. Endocrinol. Metab. 1994, 78, 847–854. [Google Scholar]
- Salamonsen, L.A.; Hannan, N.J.; Dimitriadis, E. Cytokines and chemokines during human embryo implantation: Roles in implantation and early placentation. Semin. Reprod. Med. 2007, 25, 437–444. [Google Scholar] [CrossRef]
- Saito, S. Cytokine network at the feto-maternal interface. J. Reprod. Immunol. 2000, 47, 87–103. [Google Scholar] [CrossRef]
- Robertson, S.A.; Chin, P.Y.; Glynn, D.J.; Thompson, J.G. Peri-conceptual cytokines—Setting the trajectory for embryo implantation, pregnancy and beyond. Am. J. Reprod. Immunol. 2011, 66 (Suppl. 1), 2–10. [Google Scholar] [CrossRef]
- Schjenken, J.E.; Robertson, S.A. Seminal fluid and immune adaptation for pregnancy—Comparative biology in mammalian species. Reprod. Domest. Anim. 2014, 49 (Suppl. 3), 27–36. [Google Scholar] [CrossRef]
- Robertson, S.A. Seminal fluid signaling in the female reproductive tract: Lessons from rodents and pigs. J. Anim. Sci. 2007, 85 (Suppl. 13), E36–E44. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A.; Sharkey, D.J. Seminal fluid and fertility in women. Fertil. Steril. 2016, 106, 511–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takabatake, K.; Fujiwara, H.; Goto, Y.; Nakayama, T.; Higuchi, T.; Maeda, M.; Mori, T. Intravenous administration of splenocytes in early pregnancy changes the implantation window in mice. Hum. Reprod. 1997, 12, 583–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takabatake, K.; Fujiwara, H.; Goto, Y.; Nakayama, T.; Higuchi, T.; Fujita, J.; Maeda, M.; Mori, T. Splenocytes in early pregnancy promote embryo implantation by regulating endometrial differentiation in mice. Hum. Reprod. 1997, 12, 2102–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, M.A. Embryo Transfer Surgery. Methods Mol. Biol. 2020, 2066, 101–106. [Google Scholar] [PubMed]
- Fujiwara, H.; Araki, Y.; Toshimori, K. Is the zona pellucida an intrinsic source of signals activating maternal recognition of the developing mammalian embryo? J. Reprod. Immunol. 2009, 81, 1–8. [Google Scholar] [CrossRef]
- Fujiwara, H.; Araki, Y.; Imakawa, K.; Saito, S.; Daikoku, T.; Shigeta, M.; Kanzaki, H.; Mori, T. Dual Positive Regulation of Embryo Implantation by Endocrine and Immune Systems—Step-by-Step Maternal Recognition of the Developing Embryo. Am. J. Reprod. Immunol. 2016, 75, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H. Do circulating blood cells contribute to maternal tissue remodeling and embryo-maternal cross-talk around the implantation period? Mol. Hum. Reprod. 2009, 15, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H. Immune cells contribute to systemic cross-talk between the embryo and mother during early pregnancy in cooperation with the endocrine system. Reprod. Med. Biol. 2006, 5, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Morton, H.; Hegh, V.; Clunie, G.J. Immunosuppression detected in pregnant mice by rosette inhibition test. Nature 1974, 249, 459–460. [Google Scholar] [CrossRef]
- Clarke, F.M. Identification of molecules and mechanisms involved in the ‘early pregnancy factor’ system. Reprod. Fertil. Dev. 1992, 4, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Morton, H. Early pregnancy factor: An extracellular chaperonin 10 homologue. Immunol. Cell Biol. 1998, 76, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhu, X.; Chen, R.; Liu, J.; Liu, P.; Hu, A.; Wu, L.; Hua, H.; Yuan, H. Early Pregnancy Factor Enhances the Generation and Function of CD4(+)CD25(+) Regulatory T Cells. Tohoku. J. Exp. Med. 2016, 240, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatkin, C.W.; Roussev, R.G.; Stout, M.; Absalon-Medina, V.; Ramu, S.; Goodman, C.; Coulam, C.B.; Gilbert, R.O.; Godke, R.A.; Barnea, E.R. PreImplantation Factor (PIF) correlates with early mammalian embryo development-bovine and murine models. Reprod. Biol. Endocrinol. 2011, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Barnea, E.R. Applying embryo-derived immune tolerance to the treatment of immune disorders. Ann. N. Y. Acad. Sci. 2007, 1110, 602–618. [Google Scholar] [CrossRef]
- Barnea, E.R.; Almogi-Hazan, O.; Or, R.; Mueller, M.; Ria, F.; Weiss, L.; Paidas, M.J. Immune regulatory and neuroprotective properties of preimplantation factor: From newborn to adult. Pharmacol. Ther. 2015, 156, 10–25. [Google Scholar] [CrossRef]
- Hayrabedyan, S.; Shainer, R.; Yekhtin, Z.; Weiss, L.; Almogi-Hazan, O.; Or, R.; Farnsworth, C.L.; Newsome, S.; Todorova, K.; Paidas, M.J.; et al. Synthetic PreImplantation Factor (sPIF) induces posttranslational protein modification and reverses paralysis in EAE mice. Sci. Rep. 2019, 9, 12876. [Google Scholar] [CrossRef]
- Clark, G.F. A role for carbohydrate recognition in mammalian sperm-egg binding. Biochem. Biophys. Res. Commun. 2014, 450, 1195–1203. [Google Scholar] [CrossRef]
- Schumacher, A.; Brachwitz, N.; Sohr, S.; Engeland, K.; Langwisch, S.; Dolaptchieva, M.; Alexander, T.; Taran, A.; Malfertheiner, S.F.; Costa, S.D.; et al. Human chorionic gonadotropin attracts regulatory T cells into the fetal-maternal interface during early human pregnancy. J. Immunol. 2009, 182, 5488–5497. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, A.; Heinze, K.; Witte, J.; Poloski, E.; Linzke, N.; Woidacki, K.; Zenclussen, A.C. Human chorionic gonadotropin as a central regulator of pregnancy immune tolerance. J. Immunol. 2013, 190, 2650–2658. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, A. Human Chorionic Gonadotropin as a Pivotal Endocrine Immune Regulator Initiating and Preserving Fetal Tolerance. Int. J. Mol. Sci. 2017, 18, 2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adcock, E.W., 3rd; Teasdale, T.; August, C.S.; Cox, S.; Meschia, G.; Ballaglia, T.C.; Naughton, M.A. Human chorionic gonadotropin: Its possible role in maternal lymphocyte suppression. Science 1973, 181, 845–847. [Google Scholar] [CrossRef] [PubMed]
- Muchmore, A.V.; Blaese, R.M. Immunoregulatory properties of fractions from human pregnancy urine: Evidence that human chorionic gonadotropin is not responsible. J. Immunol. 1977, 118, 881–886. [Google Scholar]
- Kosaka, K.; Fujiwara, H.; Tatsumi, K.; Yoshioka, S.; Sato, Y.; Egawa, H.; Higuchi, T.; Nakayama, T.; Ueda, M.; Maeda, M.; et al. Human chorionic gonadotropin (HCG) activates monocytes to produce interleukin-8 via a different pathway from luteinizing hormone/HCG receptor system. J. Clin. Endocrinol. Metab. 2002, 87, 5199–5208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, N.; Kelly, R.; Saunders, P.T.; Critchley, H.O. Proliferation of uterine natural killer cells is induced by human chorionic gonadotropin and mediated via the mannose receptor. Endocrinology 2009, 150, 2882–2888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, L.A. Hyperglycosylated hCG. Placenta 2007, 28, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, A.; Zenclussen, A.C. Human Chorionic Gonadotropin-Mediated Immune Responses That Facilitate Embryo Implantation and Placentation. Front. Immunol. 2019, 10, 2896. [Google Scholar] [CrossRef]
- Rolle, L.; Memarzadeh Tehran, M.; Morell-Garcia, A.; Raeva, Y.; Schumacher, A.; Hartig, R.; Costa, S.D.; Jensen, F.; Zenclussen, A.C. Cutting edge: IL-10-producing regulatory B cells in early human pregnancy. Am. J. Reprod. Immunol. 2013, 70, 448–453. [Google Scholar] [CrossRef]
- Guzman-Genuino, R.M.; Eldi, P.; Garcia-Valtanen, P.; Hayball, J.D.; Diener, K.R. Uterine B Cells Exhibit Regulatory Properties During the Peri-Implantation Stage of Murine Pregnancy. Front. Immunol. 2019, 10, 2899. [Google Scholar] [CrossRef] [Green Version]
- Koushaeian, L.; Ghorbani, F.; Ahmadi, M.; Eghbal-Fard, S.; Zamani, M.; Danaii, S.; Yousefi, B.; Jadidi-Niaragh, F.; Hamdi, K.; Yousefi, M. The role of IL-10-producing B cells in repeated implantation failure patients with cellular immune abnormalities. Immunol. Lett. 2019, 214, 16–22. [Google Scholar] [CrossRef]
- Egawa, H.; Fujiwara, H.; Hirano, T.; Nakayama, T.; Higuchi, T.; Tatsumi, K.; Mori, T.; Fujii, S. Peripheral blood mononuclear cells in early pregnancy promote invasion of human choriocarcinoma cell line, BeWo cells. Hum. Reprod. 2002, 17, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, T.; Fujiwara, H.; Maeda, M.; Inoue, T.; Yoshioka, S.; Mori, T.; Fujii, S. Human peripheral blood mononuclear cells (PBMC) in early pregnancy promote embryo invasion in vitro: HCG enhances the effects of PBMC. Hum. Reprod. 2002, 17, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, N.; Yan, W.; Yin, T.; Wang, Y.; Guo, Y.; Zhou, D.; Xu, M.; Ding, J.; Yang, J. HCG-Activated Human Peripheral Blood Mononuclear Cells (PBMC) Promote Trophoblast Cell Invasion. PLoS ONE 2015, 10, e0125589. [Google Scholar] [CrossRef] [PubMed]
- O’Tierney-Ginn, P.F.; Lash, G.E. Beyond pregnancy: Modulation of trophoblast invasion and its consequences for fetal growth and long-term children’s health. J. Reprod. Immunol. 2014, 104–105, 37–42. [Google Scholar]
- Sato, Y. Endovascular trophoblast and spiral artery remodeling. Mol. Cell Endocrinol. 2019, 110699. [Google Scholar] [CrossRef] [PubMed]
- Yagel, S.; Geva, T.E.; Solomon, H.; Shimonovitz, S.; Reich, R.; Finci-Yeheskel, Z.; Mayer, M.; Milwidsky, A. High levels of human chorionic gonadotropin retard first trimester trophoblast invasion in vitro by decreasing urokinase plasminogen activator and collagenase activities. J. Clin. Endocrinol. Metab. 1993, 77, 1506–1511. [Google Scholar]
- Zygmunt, M.; Hahn, D.; Munstedt, K.; Bischof, P.; Lang, U. Invasion of cytotrophoblastic JEG-3 cells is stimulated by hCG in vitro. Placenta 1998, 19, 587–593. [Google Scholar] [CrossRef]
- Zygmunt, M.; McKinnon, T.; Herr, F.; Lala, P.K.; Han, V.K. HCG increases trophoblast migration in vitro via the insulin-like growth factor-II/mannose-6 phosphate receptor. Mol. Hum. Reprod. 2005, 11, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Handschuh, K.; Guibourdenche, J.; Tsatsaris, V.; Guesnon, M.; Laurendeau, I.; Evain-Brion, D.; Fournier, T. Human chorionic gonadotropin produced by the invasive trophoblast but not the villous trophoblast promotes cell invasion and is down-regulated by peroxisome proliferator-activated receptor-gamma. Endocrinology 2007, 148, 5011–5019. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.L.; Chiu, P.C.; Hautala, L.; Salo, T.; Yeung, W.S.; Stenman, U.H.; Koistinen, H. Human chorionic gonadotropin and its free beta-subunit stimulate trophoblast invasion independent of LH/hCG receptor. Mol. Cell Endocrinol. 2013, 375, 43–52. [Google Scholar] [CrossRef]
- Fujiwara, H.; Higuchi, T.; Yamada, S.; Hirano, T.; Sato, Y.; Nishioka, Y.; Yoshioka, S.; Tatsumi, K.; Ueda, M.; Maeda, M.; et al. Human extravillous trophoblasts express laeverin, a novel protein that belongs to membrane-bound gluzincin metallopeptidases. Biochem. Biophys. Res. Commun. 2004, 313, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Hattori, A.; Goto, Y.; Ueda, M.; Maeda, M.; Fujiwara, H.; Tsujimoto, M. Laeverin/aminopeptidase Q, a novel bestatin-sensitive leucine aminopeptidase belonging to the M1 family of aminopeptidases. J. Biol. Chem. 2007, 282, 20088–20096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, M.; Arisaka, N.; Goto, Y.; Ohsawa, Y.; Inoue, H.; Fujiwara, H.; Hattori, A.; Tsujimoto, M. Histidine 379 of human laeverin/aminopeptidase Q, a nonconserved residue within the exopeptidase motif, defines its distinctive enzymatic properties. J. Biol. Chem. 2009, 284, 34692–34702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, A.; Fujiwara, H.; Sato, Y.; Suginami, K.; Matsumoto, H.; Maruyama, M.; Konishi, I.; Hattori, A. Laeverin/aminopeptidase Q induces trophoblast invasion during human early placentation. Hum. Reprod. 2012, 27, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, S.A.; Care, A.S.; Moldenhauer, L.M. Regulatory T cells in embryo implantation and the immune response to pregnancy. J. Clin. Investig. 2018, 128, 4224–4235. [Google Scholar] [CrossRef] [Green Version]
- Ledee, N.; Petitbarat, M.; Chevrier, L.; Vitoux, D.; Vezmar, K.; Rahmati, M.; Dubanchet, S.; Gahery, H.; Bensussan, A.; Chaouat, G. The Uterine Immune Profile May Help Women With Repeated Unexplained Embryo Implantation Failure After In Vitro Fertilization. Am. J. Reprod. Immunol. 2016, 75, 388–401. [Google Scholar] [CrossRef] [Green Version]
- Abdolmohammadi-Vahid, S.; Danaii, S.; Hamdi, K.; Jadidi-Niaragh, F.; Ahmadi, M.; Yousefi, M. Novel immunotherapeutic approaches for treatment of infertility. Biomed. Pharmacother. 2016, 84, 1449–1459. [Google Scholar] [CrossRef]
- Nakagawa, K.; Kwak-Kim, J.; Ota, K.; Kuroda, K.; Hisano, M.; Sugiyama, R.; Yamaguchi, K. Immunosuppression with tacrolimus improved reproductive outcome of women with repeated implantation failure and elevated peripheral blood TH1/TH2 cell ratios. Am. J. Reprod. Immunol. 2015, 73, 353–361. [Google Scholar] [CrossRef]
- Nakagawa, K.; Kwak-Kim, J.; Kuroda, K.; Sugiyama, R.; Yamaguchi, K. Immunosuppressive treatment using tacrolimus promotes pregnancy outcome in infertile women with repeated implantation failures. Am. J. Reprod. Immunol. 2017, 78, e12682. [Google Scholar] [CrossRef]
- Yamaguchi, K. Tacrolimus treatment for infertility related to maternal-fetal immune interactions. Am. J. Reprod. Immunol. 2019, 81, e13097. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; Abdolmohamadi-Vahid, S.; Ghaebi, M.; Dolati, S.; Abbaspour-Aghdam, S.; Danaii, S.; Berjis, K.; Madadi-Javid, R.; Nouri, Z.; Siahmansouri, H.; et al. Sirolimus as a new drug to treat RIF patients with elevated Th17/Treg ratio: A double-blind, phase II randomized clinical trial. Int. Immunopharmacol. 2019, 74, 105730. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, S.; Fujiwara, H.; Nakayama, T.; Kosaka, K.; Mori, T.; Fujii, S. Intrauterine administration of autologous peripheral blood mononuclear cells promotes implantation rates in patients with repeated failure of IVF-embryo transfer. Hum. Reprod. 2006, 21, 3290–3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makrigiannakis, A.; BenKhalifa, M.; Vrekoussis, T.; Mahjub, S.; Kalantaridou, S.N.; Gurgan, T. Repeated implantation failure: A new potential treatment option. Eur. J. Clin. Investig. 2015, 45, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Achilli, C.; Duran-Retamal, M.; Saab, W.; Serhal, P.; Seshadri, S. The role of immunotherapy in in vitro fertilization and recurrent pregnancy loss: A systematic review and meta-analysis. Fertil. Steril. 2018, 110, 1089–1100. [Google Scholar] [CrossRef] [Green Version]
- Maleki-Hajiagha, A.; Razavi, M.; Rezaeinejad, M.; Rouholamin, S.; Almasi-Hashiani, A.; Pirjani, R.; Sepidarkish, M. Intrauterine administration of autologous peripheral blood mononuclear cells in patients with recurrent implantation failure: A systematic review and meta-analysis. J. Reprod. Immunol. 2019, 131, 50–56. [Google Scholar] [CrossRef]
- Yakin, K.; Oktem, O.; Urman, B. Intrauterine administration of peripheral mononuclear cells in recurrent implantation failure: A systematic review and meta-analysis. Sci. Rep. 2019, 9, 3897. [Google Scholar] [CrossRef]
- Pourmoghadam, Z.; Abdolmohammadi-Vahid, S.; Pashazadeh, F.; Aghebati-Maleki, L.; Ansari, F.; Yousefi, M. Efficacy of intrauterine administration of autologous peripheral blood mononuclear cells on the pregnancy outcomes in patients with recurrent implantation failure: A systematic review and meta-analysis. J. Reprod. Immunol. 2019, 137, 103077. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujiwara, H.; Ono, M.; Sato, Y.; Imakawa, K.; Iizuka, T.; Kagami, K.; Fujiwara, T.; Horie, A.; Tani, H.; Hattori, A.; et al. Promoting Roles of Embryonic Signals in Embryo Implantation and Placentation in Cooperation with Endocrine and Immune Systems. Int. J. Mol. Sci. 2020, 21, 1885. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051885
Fujiwara H, Ono M, Sato Y, Imakawa K, Iizuka T, Kagami K, Fujiwara T, Horie A, Tani H, Hattori A, et al. Promoting Roles of Embryonic Signals in Embryo Implantation and Placentation in Cooperation with Endocrine and Immune Systems. International Journal of Molecular Sciences. 2020; 21(5):1885. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051885
Chicago/Turabian StyleFujiwara, Hiroshi, Masanori Ono, Yukiyasu Sato, Kazuhiko Imakawa, Takashi Iizuka, Kyosuke Kagami, Tomoko Fujiwara, Akihito Horie, Hirohiko Tani, Akira Hattori, and et al. 2020. "Promoting Roles of Embryonic Signals in Embryo Implantation and Placentation in Cooperation with Endocrine and Immune Systems" International Journal of Molecular Sciences 21, no. 5: 1885. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051885