Functional Consequences of Low Activity of Transport System A for Neutral Amino Acids in Human Bone Marrow Mesenchymal Stem Cells

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
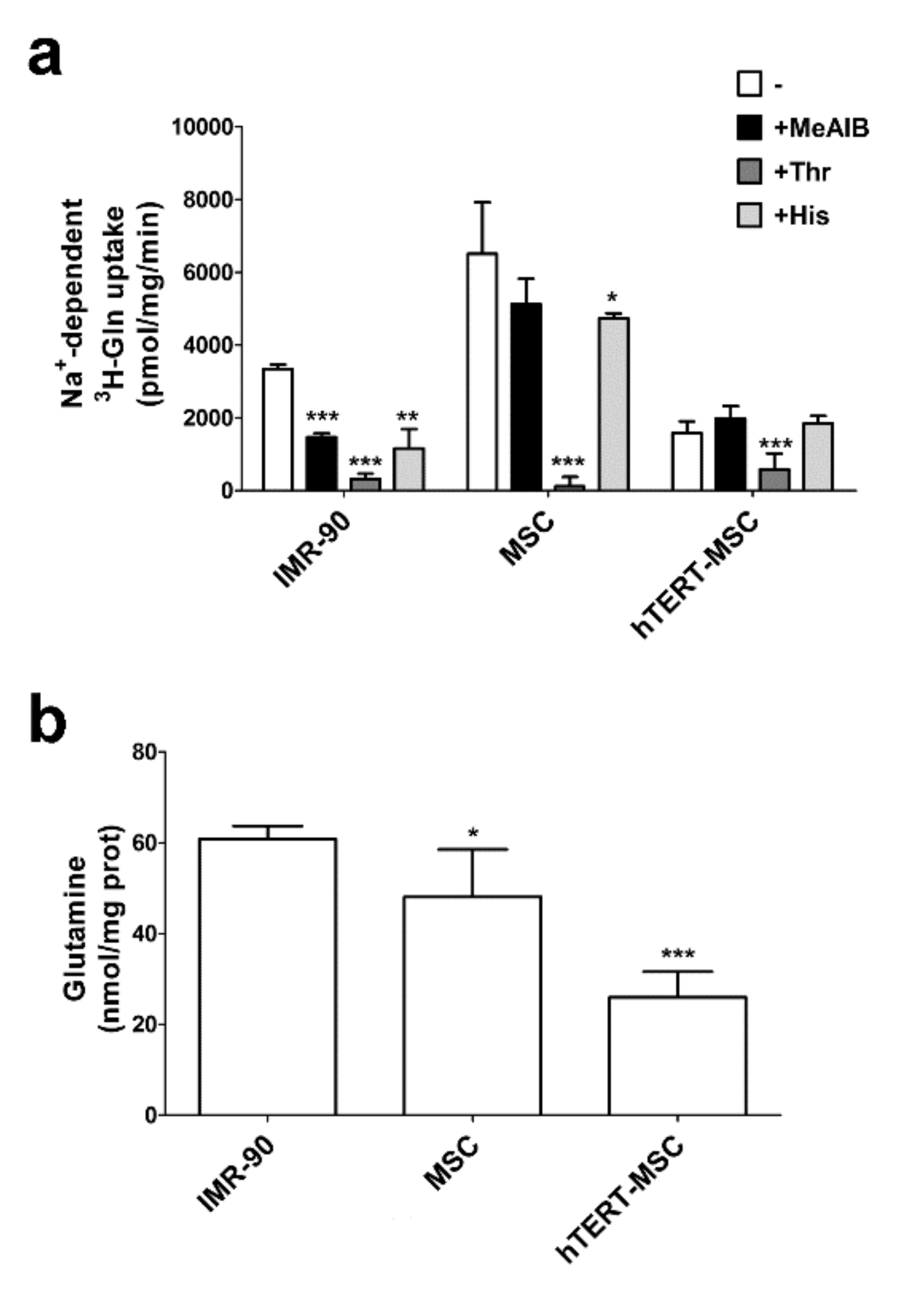
2.1. MeAIB Does Not Inhibit Glutamine Transport in Human Mesenchymal Stem Cells
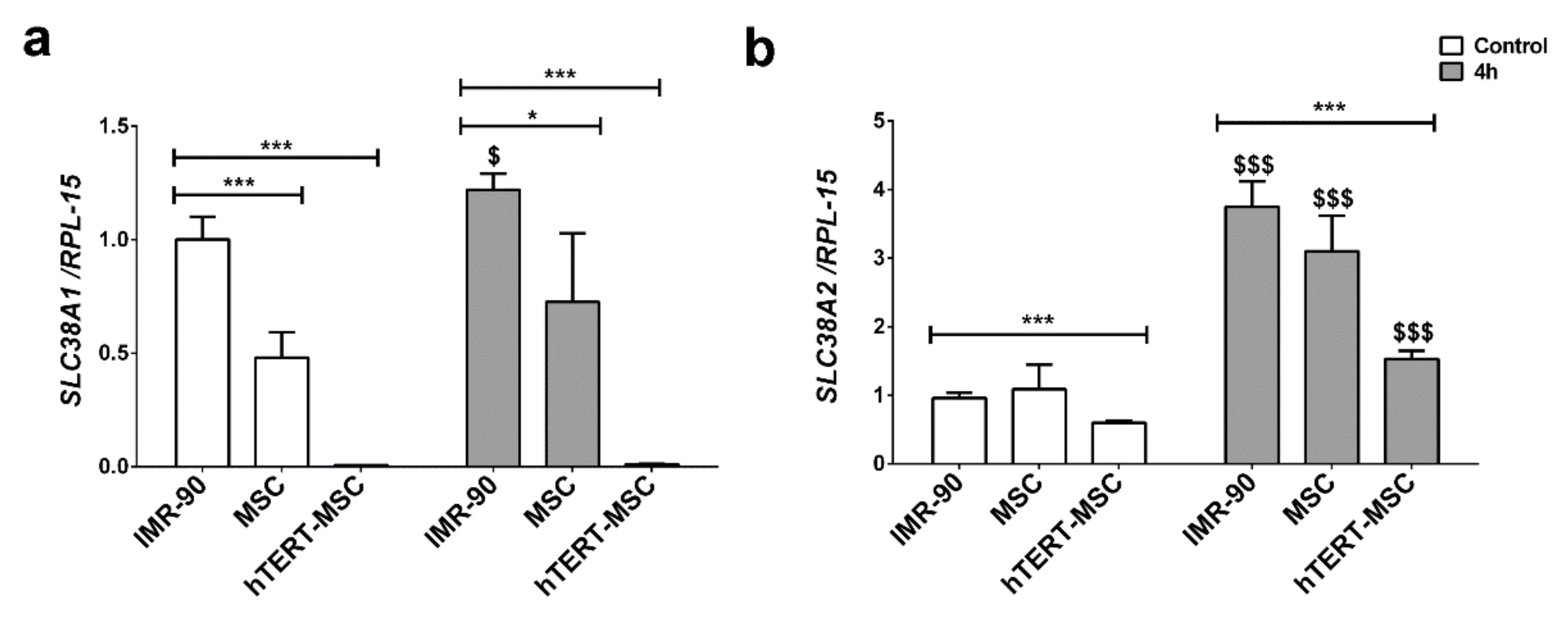
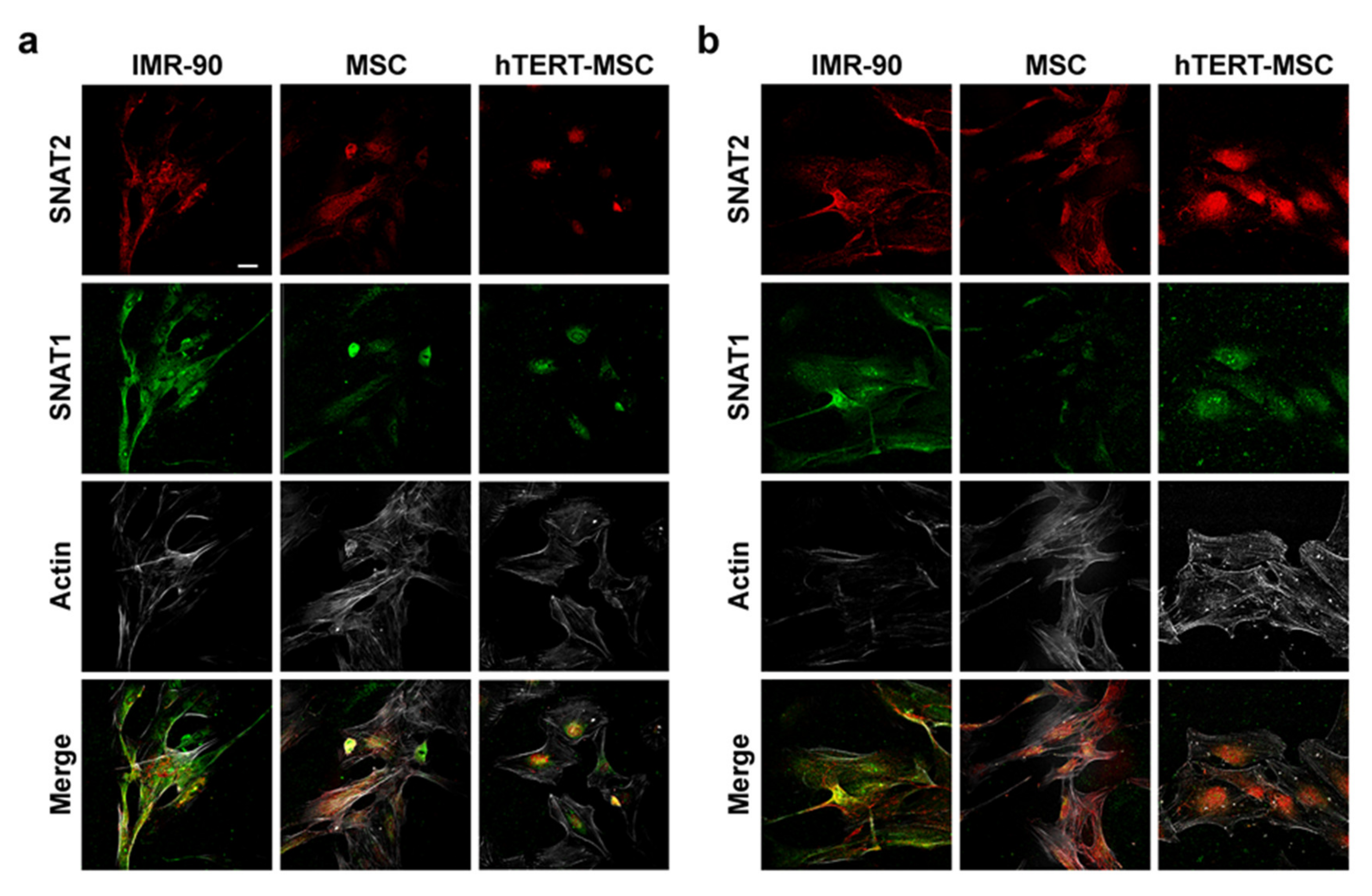
2.2. Mesenchymal Stem Cells Have a Low Expression of the System A Transporters
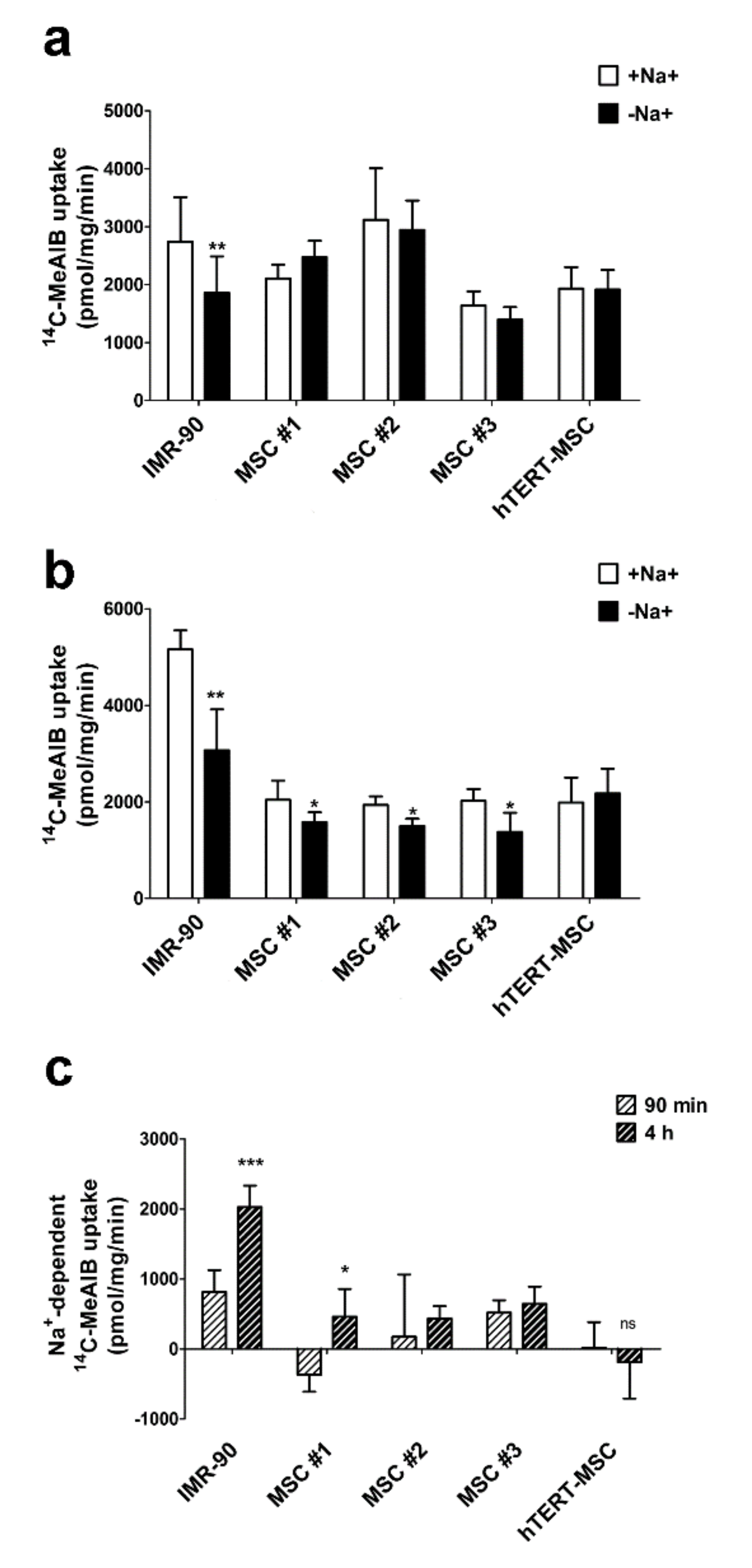
2.3. Mesenchymal Stem Cells Do Not Exhibit an Increase in System A Activity upon Amino Acid Starvation
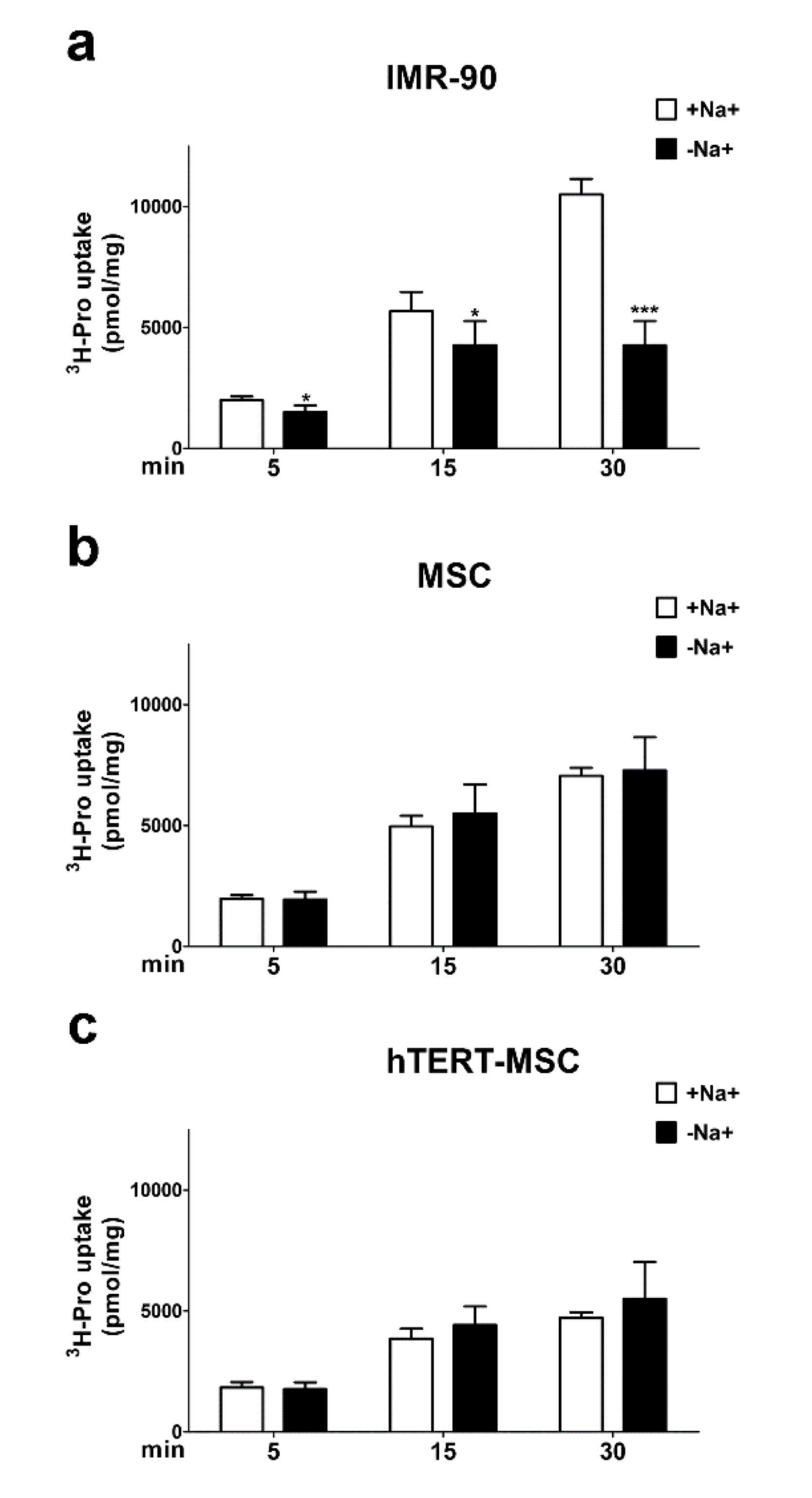
2.4. Mesenchymal Stem Cells Do Not Show a Sodium-Dependent Proline Accumulation
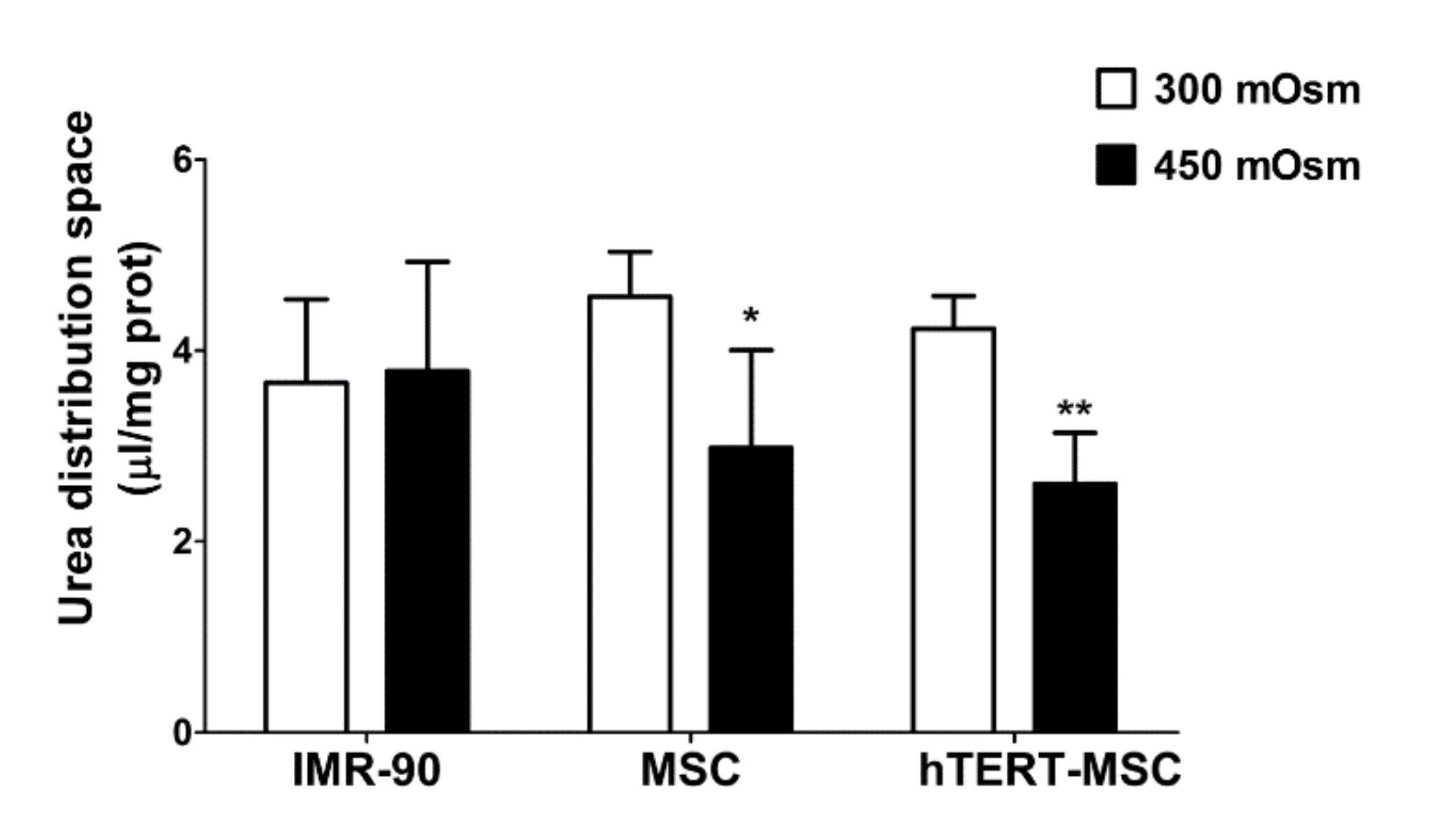
2.5. Mesenchymal Stem Cells Do Not Recover Cell Volume after Hypertonic Stress
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RT-PCR Analysis
4.3. Amino Acid Uptake
4.4. Immunofluorescence
4.5. Intracellular Gln Content
4.6. Cell Volume
4.7. Statistical Analysis
4.8. Reagents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, J.T.; Mostacada, K.; de Lima, S.; Martinez, A.M. Bone marrow mesenchymal stem cell transplantation for improving nerve regeneration. Int. Rev. Neurobiol. 2013, 108, 59–77. [Google Scholar] [PubMed]
- Karamini, A.; Bakopoulou, A.; Andreadis, D.; Gkiouras, K.; Kritis, A. Therapeutic potential of mesenchymal stromal stem cells in rheumatoid arthritis: A systematic review of in vivo studies. Stem Cell Rev. Rep. 2020. [Epub ahead of print]. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Cho, S.G. Overcoming immunoregulatory plasticity of mesenchymal stem cells for accelerated clinical applications. Int. J. Hematol. 2016, 103, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naji, A.; Eitoku, M.; Favier, B.; Deschaseaux, F.; Rouas-Freiss, N.; Suganuma, N. Biological functions of mesenchymal stem cells and clinical implications. Cell Mol. Life Sci. 2019, 76, 3323–3348. [Google Scholar] [CrossRef]
- Yuan, X.; Logan, T.M.; Ma, T. Metabolism in human mesenchymal stromal cells: A missing link between hmsc biomanufacturing and therapy? Front. Immunol. 2019, 10, 977. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, S.; Mihara, K.; Downing, J.R.; Pui, C.H.; Campana, D. Mesenchymal cells regulate the response of acute lymphoblastic leukemia cells to asparaginase. J. Clin. Investig. 2007, 117, 1049–1057. [Google Scholar] [CrossRef]
- Ehsanipour, E.A.; Sheng, X.; Behan, J.W.; Wang, X.; Butturini, A.; Avramis, V.I.; Mittelman, S.D. Adipocytes cause leukemia cell resistance to l-asparaginase via release of glutamine. Cancer Res. 2013, 73, 2998–3006. [Google Scholar] [CrossRef] [Green Version]
- Chiu, M.; Taurino, G.; Bianchi, M.G.; Kilberg, M.S.; Bussolati, O. Asparagine synthetase in cancer: Beyond acute lymphoblastic leukemia. Front. Oncol. 2019, 9, 1480. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Cheng, T. Q’s next: The diverse functions of glutamine in metabolism, cell biology and cancer. Oncogene 2010, 29, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Bhutia, Y.D.; Ganapathy, V. Glutamine transporters in mammalian cells and their functions in physiology and cancer. Biochim. Biophys. Acta 2016, 1863, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Broer, S. The slc38 family of sodium-amino acid co-transporters. Pflüg. Arch. 2014, 466, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Shotwell, M.A.; Jayme, D.W.; Kilberg, M.S.; Oxender, D.L. Neutral amino acid transport systems in chinese hamster ovary cells. J. Biol. Chem. 1981, 256, 5422–5427. [Google Scholar] [PubMed]
- Ericsson, A.; Hamark, B.; Jansson, N.; Johansson, B.R.; Powell, T.L.; Jansson, T. Hormonal regulation of glucose and system a amino acid transport in first trimester placental villous fragments. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R656–R662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, H.N.; Oxender, D.L.; Liang, M.; Vatz, K.A. The use of n-methylation to direct route of mediated transport of amino acids. J. Biol. Chem. 1965, 240, 3609–3616. [Google Scholar]
- Mackenzie, B.; Erickson, J.D. Sodium-coupled neutral amino acid (system n/a) transporters of the slc38 gene family. Pflüg. Arch. 2004, 447, 784–795. [Google Scholar] [CrossRef]
- Takahashi, Y.; Nishimura, T.; Maruyama, T.; Tomi, M.; Nakashima, E. Contributions of system a subtypes to alpha-methylaminoisobutyric acid uptake by placental microvillous membranes of human and rat. Amino Acids 2017, 49, 795–803. [Google Scholar] [CrossRef]
- Foster, A.C.; Farnsworth, J.; Lind, G.E.; Li, Y.X.; Yang, J.Y.; Dang, V.; Penjwini, M.; Viswanath, V.; Staubli, U.; Kavanaugh, M.P. D-serine is a substrate for neutral amino acid transporters asct1/slc1a4 and asct2/slc1a5, and is transported by both subtypes in rat hippocampal astrocyte cultures. PLoS ONE 2016, 11, e0156551. [Google Scholar] [CrossRef]
- Bussolati, O.; Rotoli, B.M.; Laris, P.C.; Dall’Asta, V.; Gazzola, G.C. The preferential interaction of l-threonine with transport system asc in cultured human fibroblasts. Biochim. Biophys. Acta 1991, 1070, 305–312. [Google Scholar] [CrossRef]
- Chaudhry, F.A.; Reimer, R.J.; Krizaj, D.; Barber, D.; Storm-Mathisen, J.; Copenhagen, D.R.; Edwards, R.H. Molecular analysis of system n suggests novel physiological roles in nitrogen metabolism and synaptic transmission. Cell 1999, 99, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Rae, C.; Hare, N.; Bubb, W.A.; McEwan, S.R.; Broer, A.; McQuillan, J.A.; Balcar, V.J.; Conigrave, A.D.; Broer, S. Inhibition of glutamine transport depletes glutamate and gaba neurotransmitter pools: Further evidence for metabolic compartmentation. J. Neurochem. 2003, 85, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, M.; Bussolati, O. Department of Medicine and Surgery, University of Parma: Parma, Italy. 2020; Manuscript in preparation. [Google Scholar]
- Franchi-Gazzola, R.; Gaccioli, F.; Bevilacqua, E.; Visigalli, R.; Dall’Asta, V.; Sala, R.; Varoqui, H.; Erickson, J.D.; Gazzola, G.C.; Bussolati, O. The synthesis of snat2 transporters is required for the hypertonic stimulation of system a transport activity. Biochim. Biophys. Acta 2004, 1667, 157–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takanaga, H.; Mackenzie, B.; Suzuki, Y.; Hediger, M.A. Identification of mammalian proline transporter sit1 (slc6a20) with characteristics of classical system imino. J. Biol. Chem. 2005, 280, 8974–8984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Asta, V.; Rossi, P.A.; Bussolati, O.; Gazzola, G.C. Response of human fibroblasts to hypertonic stress. Cell shrinkage is counteracted by an enhanced active transport of neutral amino acids. J. Biol. Chem. 1994, 269, 10485–10491. [Google Scholar]
- Breborowicz, A.; Polubinska, A.; Oreopoulos, D.G. Changes in volume of peritoneal mesothelial cells exposed to osmotic stress. Perit. Dial. Int. 1999, 19, 119–123. [Google Scholar] [CrossRef]
- Dall’Asta, V.; Bussolati, O.; Sala, R.; Parolari, A.; Alamanni, F.; Biglioli, P.; Gazzola, G.C. Amino acids are compatible osmolytes for volume recovery after hypertonic shrinkage in vascular endothelial cells. Am. J. Physiol. 1999, 276, C865–C872. [Google Scholar] [CrossRef]
- Maallem, S.; Mutin, M.; Gonzalez-Gonzalez, I.M.; Zafra, F.; Tappaz, M.L. Selective tonicity-induced expression of the neutral amino-acid transporter snat2 in oligodendrocytes in rat brain following systemic hypertonicity. Neuroscience 2008, 153, 95–107. [Google Scholar] [CrossRef]
- Franchi-Gazzola, R.; Visigalli, R.; Dall’Asta, V.; Sala, R.; Woo, S.K.; Kwon, H.M.; Gazzola, G.C.; Bussolati, O. Amino acid depletion activates tonebp and sodium-coupled inositol transport. Am. J. Physiol. Cell Physiol. 2001, 280, C1465–C1474. [Google Scholar] [CrossRef]
- Dall’asta, V.; Franchi-Gazzola, R.; Bussolati, O.; Sala, R.; Rotoli, B.M.; Rossi, P.A.; Uggeri, J.; Belletti, S.; Visigalli, R.; Gazzola, G.C. Emerging roles for sodium dependent amino acid transport in mesenchymal cells. Amino Acids 1996, 11, 117–133. [Google Scholar]
- Nicklin, P.; Bergman, P.; Zhang, B.; Triantafellow, E.; Wang, H.; Nyfeler, B.; Yang, H.; Hild, M.; Kung, C.; Wilson, C.; et al. Bidirectional transport of amino acids regulates mtor and autophagy. Cell 2009, 136, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Timmerman, L.A.; Holton, T.; Yuneva, M.; Louie, R.J.; Padro, M.; Daemen, A.; Hu, M.; Chan, D.A.; Ethier, S.P.; van ‘t Veer, L.J.; et al. Glutamine sensitivity analysis identifies the xct antiporter as a common triple-negative breast tumor therapeutic target. Cancer Cell 2013, 24, 450–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchi-Gazzola, R.; Dall’Asta, V.; Sala, R.; Visigalli, R.; Bevilacqua, E.; Gaccioli, F.; Gazzola, G.C.; Bussolati, O. The role of the neutral amino acid transporter snat2 in cell volume regulation. Acta Physiol. 2006, 187, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.M.; Cwiklinski, E.; Shah, D.S.; Stretton, C.; Hyde, R.; Taylor, P.M.; Hundal, H.S. Effects of sodium and amino acid substrate availability upon the expression and stability of the snat2 (slc38a2) amino acid transporter. Front. Pharmacol. 2018, 9, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaccioli, F.; Huang, C.C.; Wang, C.; Bevilacqua, E.; Franchi-Gazzola, R.; Gazzola, G.C.; Bussolati, O.; Snider, M.D.; Hatzoglou, M. Amino acid starvation induces the snat2 neutral amino acid transporter by a mechanism that involves eukaryotic initiation factor 2alpha phosphorylation and cap-independent translation. J. Biol. Chem. 2006, 281, 17929–17940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, S.; Park, S.H.; Patel, A.; Carn, J.; Lee, K.; Kaplan, D.L. Hypoxia and amino acid supplementation synergistically promote the osteogenesis of human mesenchymal stem cells on silk protein scaffolds. Tissue Eng. Part A 2010, 16, 3623–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comes, S.; Gagliardi, M.; Laprano, N.; Fico, A.; Cimmino, A.; Palamidessi, A.; De Cesare, D.; De Falco, S.; Angelini, C.; Scita, G.; et al. L-proline induces a mesenchymal-like invasive program in embryonic stem cells by remodeling h3k9 and h3k36 methylation. Stem Cell Rep. 2013, 1, 307–321. [Google Scholar] [CrossRef] [Green Version]
- D’Aniello, C.; Fico, A.; Casalino, L.; Guardiola, O.; Di Napoli, G.; Cermola, F.; De Cesare, D.; Tate, R.; Cobellis, G.; Patriarca, E.J.; et al. A novel autoregulatory loop between the gcn2-atf4 pathway and (l)-proline [corrected] metabolism controls stem cell identity. Cell Death Differ. 2015, 22, 1094–1105. [Google Scholar] [CrossRef]
- Yi, X.; Liu, J.; Wu, P.; Gong, Y.; Xu, X.; Li, W. The whole transcriptional profiling of cellular metabolism during adipogenesis from hmscs. J. Cell. Physiol. 2020, 235, 349–363. [Google Scholar] [CrossRef]
- Meier, C.; Camargo, S.M.; Hunziker, S.; Moehrlen, U.; Gros, S.J.; Bode, P.; Leu, S.; Meuli, M.; Holland-Cunz, S.; Verrey, F.; et al. Intestinal imino transporter sit1 is not expressed in human newborns. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G887–G895. [Google Scholar] [CrossRef]
- Andre, V.; Longoni, D.; Bresolin, S.; Cappuzzello, C.; Dander, E.; Galbiati, M.; Bugarin, C.; Di Meglio, A.; Nicolis, E.; Maserati, E.; et al. Mesenchymal stem cells from shwachman-diamond syndrome patients display normal functions and do not contribute to hematological defects. Blood Cancer J. 2012, 2, e94. [Google Scholar] [CrossRef]
- Bustin, S.A. Absolute quantification of mrna using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, M.; Taurino, G.; Bianchi, M.G.; Ottaviani, L.; Andreoli, R.; Ciociola, T.; Lagrasta, C.A.M.; Tardito, S.; Bussolati, O. Oligodendroglioma cells lack glutamine synthetase and are auxotrophic for glutamine, but do not depend on glutamine anaplerosis for growth. Int. J. Mol. Sci. 2018, 19, 1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, M.; Taurino, G.; Bianchi, M.G.; Dander, E.; Fallati, A.; Giuliani, N.; D’Amico, G.; Bussolati, O. Functional Consequences of Low Activity of Transport System A for Neutral Amino Acids in Human Bone Marrow Mesenchymal Stem Cells. Int. J. Mol. Sci. 2020, 21, 1899. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051899
Chiu M, Taurino G, Bianchi MG, Dander E, Fallati A, Giuliani N, D’Amico G, Bussolati O. Functional Consequences of Low Activity of Transport System A for Neutral Amino Acids in Human Bone Marrow Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2020; 21(5):1899. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051899
Chicago/Turabian StyleChiu, Martina, Giuseppe Taurino, Massimiliano G. Bianchi, Erica Dander, Alessandra Fallati, Nicola Giuliani, Giovanna D’Amico, and Ovidio Bussolati. 2020. "Functional Consequences of Low Activity of Transport System A for Neutral Amino Acids in Human Bone Marrow Mesenchymal Stem Cells" International Journal of Molecular Sciences 21, no. 5: 1899. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051899