Gene Polymorphisms in Boar Spermatozoa and Their Associations with Post-Thaw Semen Quality



Abstract
:1. Introduction
2. Results
2.1. Assessment of Semen Quality
2.2. RNA-Seq Mapping and Variant Calling
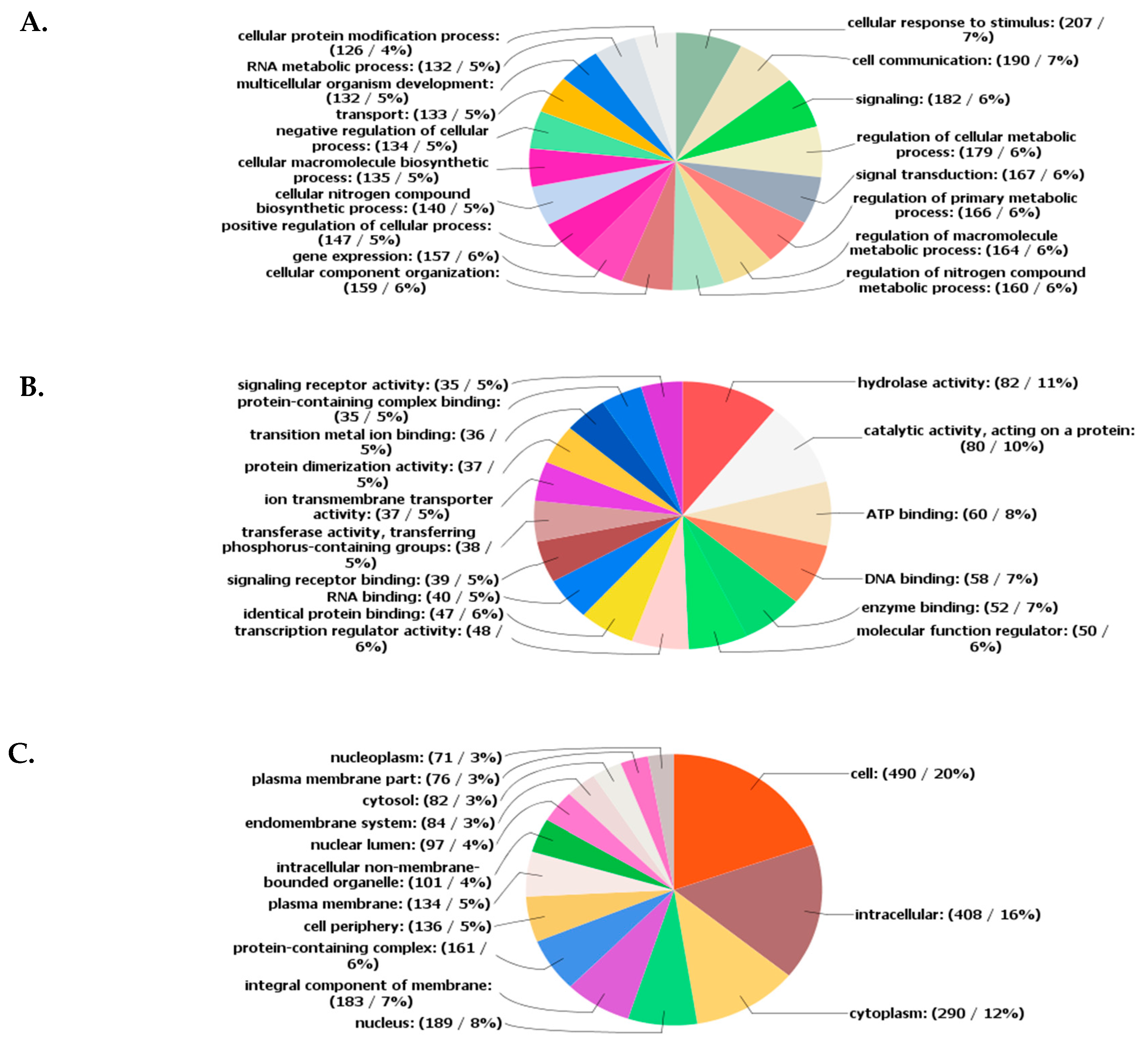
2.3. SNP Abundance in Functional Categories of Candidate Genes (CGs)
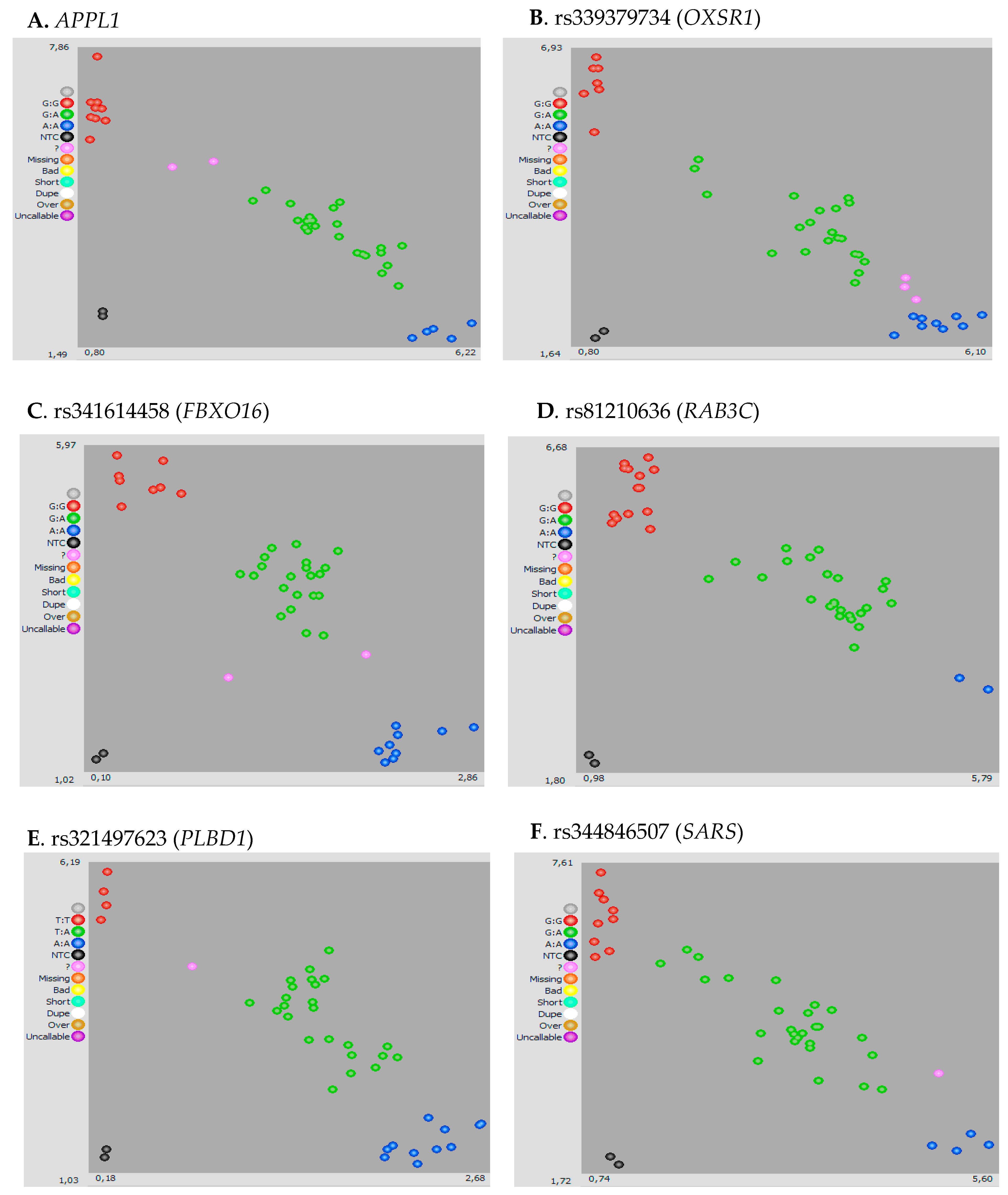
2.4. KASP Genotyping and Validation
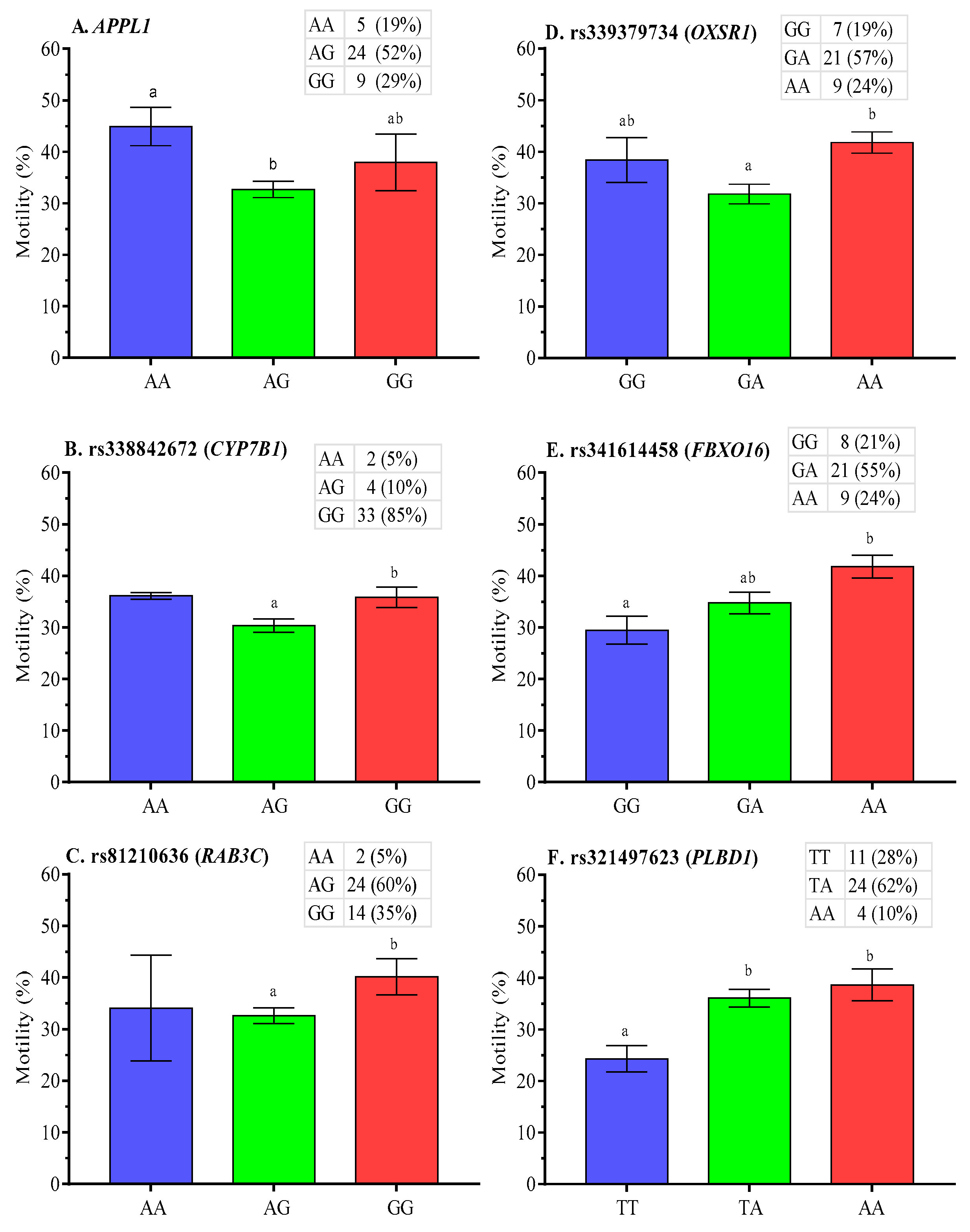
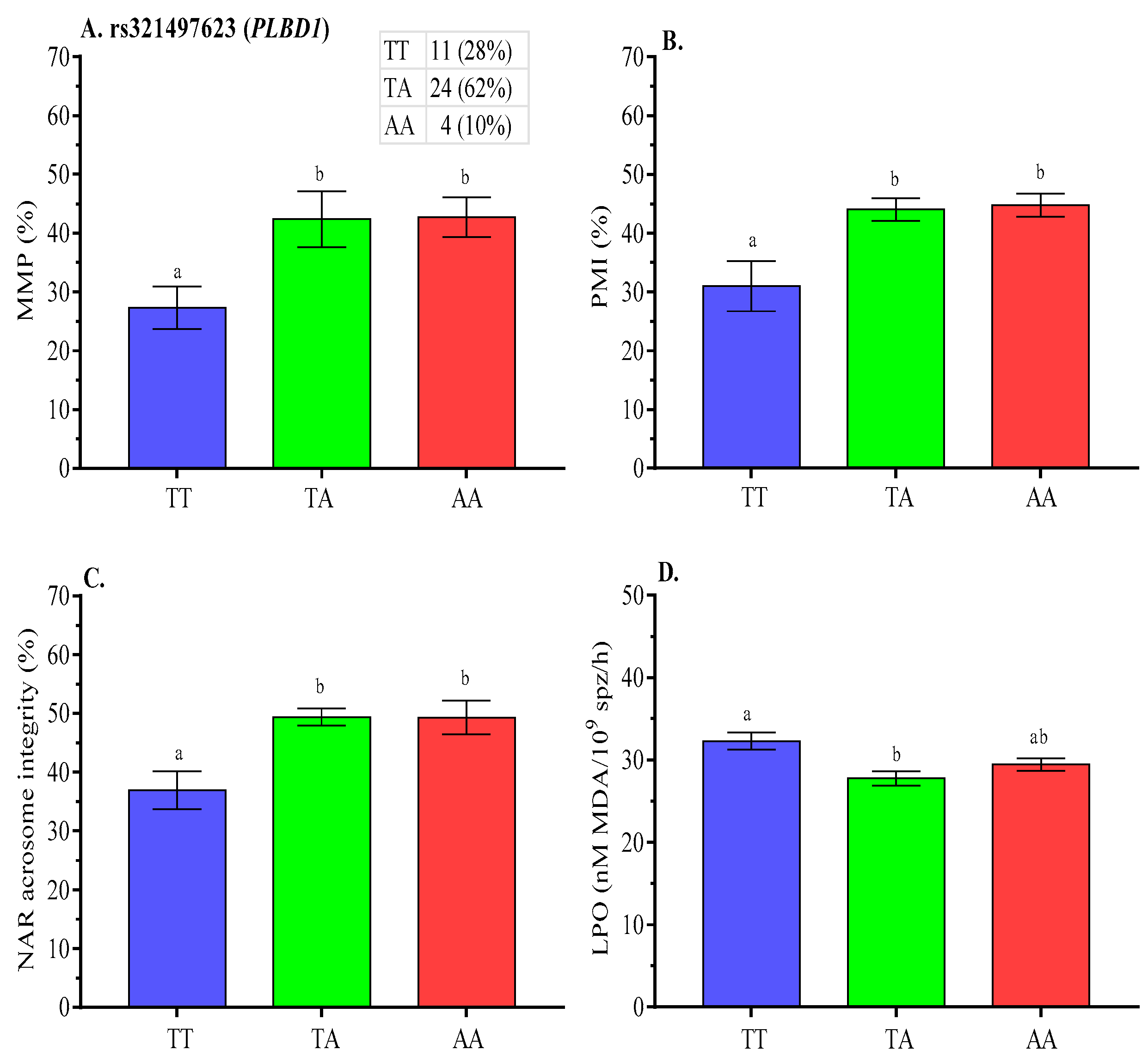
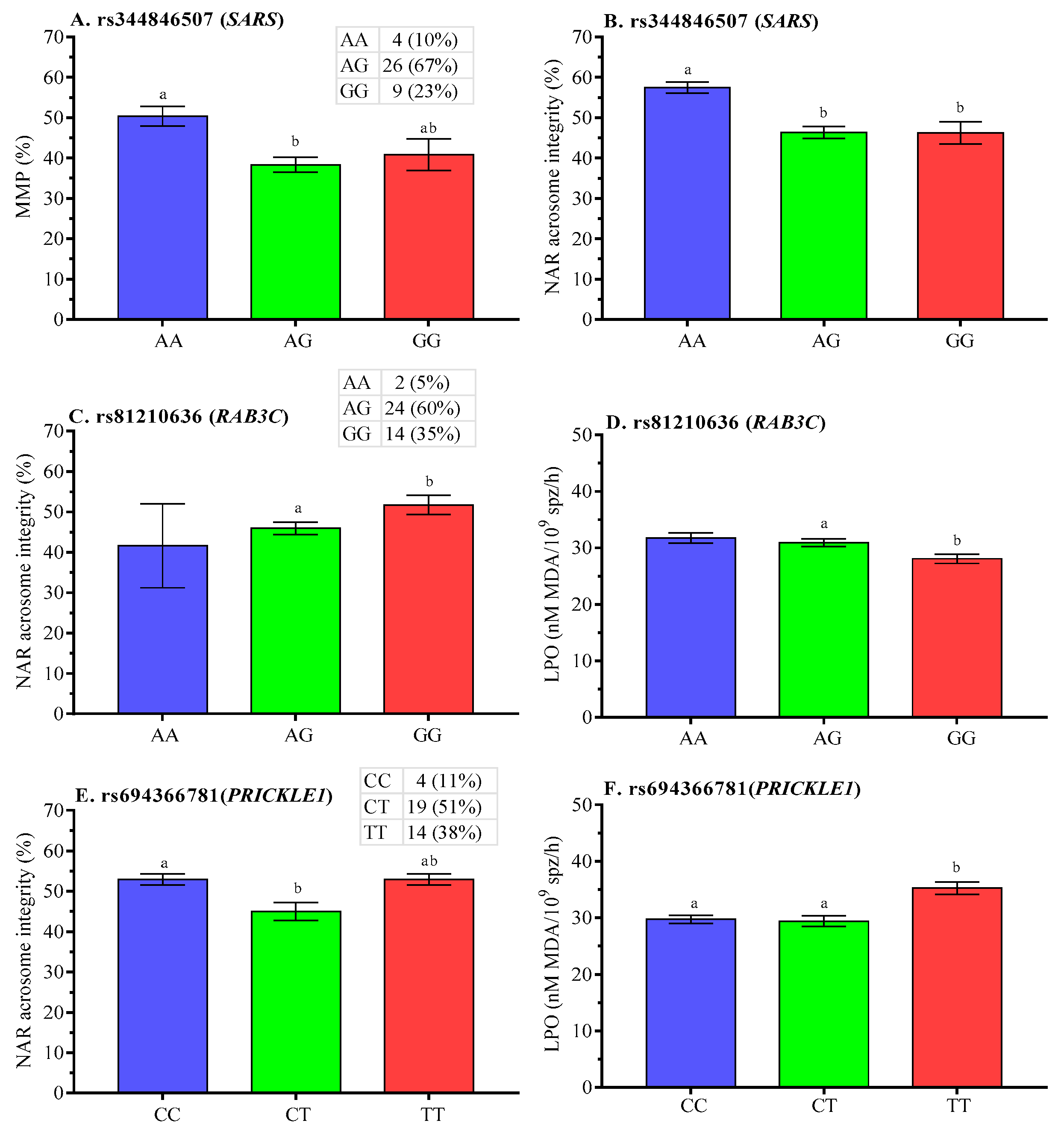
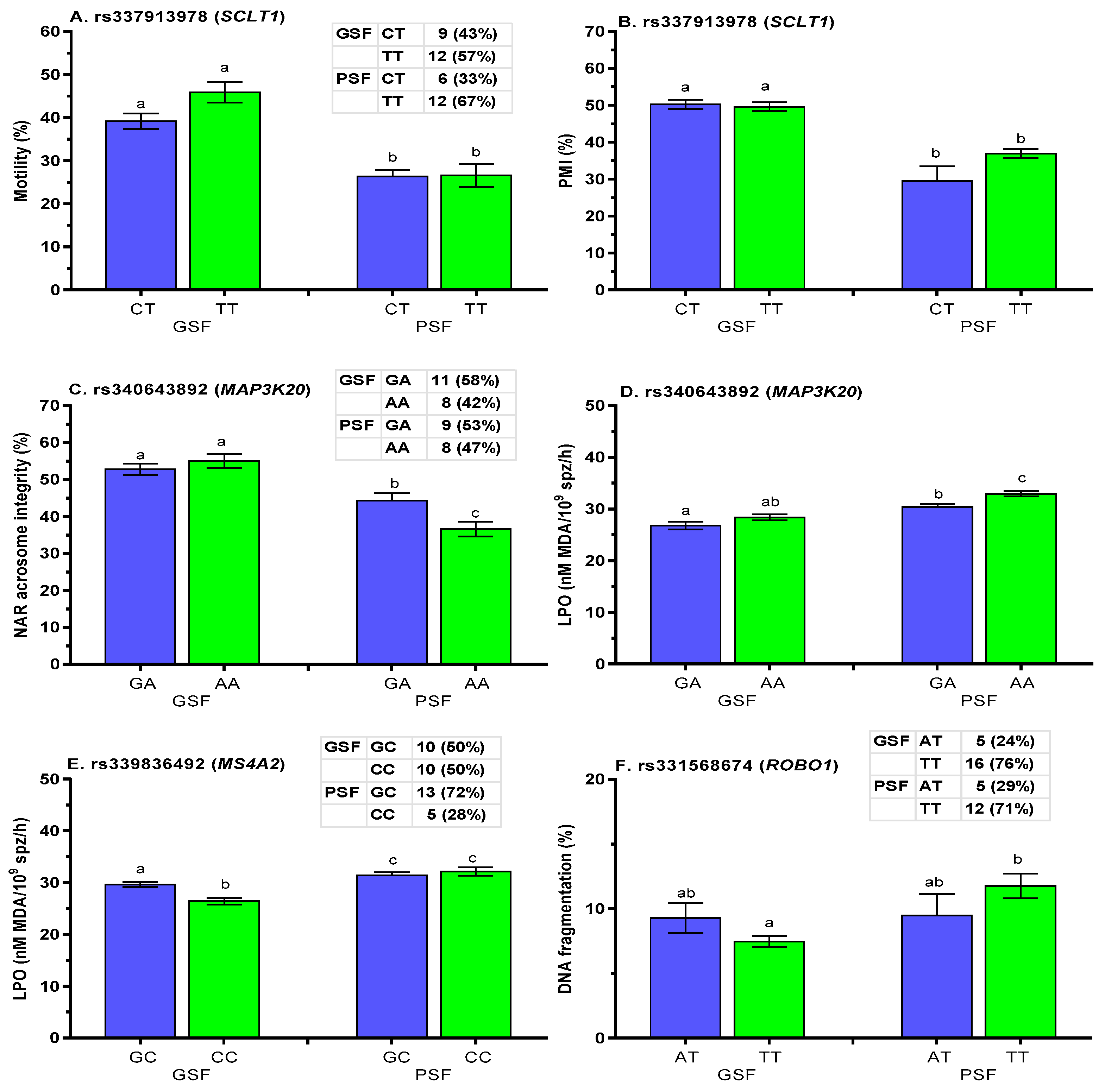
2.5. Association Analysis of SNPs with Post-Thaw Semen Quality
3. Discussion
3.1. SNP Functional Classes and Validation
3.2. SNP Associations with Post-Thaw Semen Quality
4. Materials and Methods
4.1. Animals and Semen Collections
4.2. Cryopreservation Procedure
4.3. Semen Quality Assessment
4.3.1. Sperm Motility
4.3.2. Mitochondrial Membrane Potential (MMP)
4.3.3. Plasma Membrane Integrity (PMI)
4.3.4. Normal Apical Ridge (NAR) Acrosome Integrity
4.3.5. DNA Fragmentation
4.3.6. Lipid Peroxidation (LPO)
4.4. Genomic DNA Isolation
4.5. Variant Calling and Filtering Analysis
4.6. KEGG Pathways and GO Enrichment Analysis
4.7. SNP KASP Genotyping Assay
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of data and materials
References
- Didion, B.A.; Braun, G.D.; Duggan, M.V. Field fertility of frozen boar semen: A retrospective report comprising over 2600 AI services spanning a four year period. Anim. Reprod. Sci. 2013, 137, 189–196. [Google Scholar] [CrossRef]
- Yeste, M.; Joan, E.; Rodríguez-Gil, J.E.; Bonet, S. Artificial insemination frozen-thawed boar sperm. Mol. Reprod. Dev. 2017, 84, 802–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roca, J.; Broekhuijse, M.L.; Parrilla, I.; Rodríguez-Martínez, H.; Martinez, E.A.; Bolarin, A. Boar differences in artificial insemination outcomes: Can they be minimized? Reprod. Domest. Anim. 2015, 50 (Suppl. 2), 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, W.V.; Medrano, A.; Thurston, L.M.; Watson, P.F. The significance of cooling rates and animal variability for boar sperm cryopreservation: Insights from the cryomicroscope. Theriogenology 2005, 63, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.; Strzeżek, J.; Kordan, W. Post-thaw sperm characteristics following long-term storage of boar semen in liquid nitrogen. Anim. Reprod. Sci. 2014, 147, 119–127. [Google Scholar] [CrossRef]
- Thurston, L.M.; Siggins, K.; Mileham, A.J.; Watson, P.F.; Holt, W.V. Identification of amplified restriction fragment length polymorphism markers linked to genes controlling boar sperm viability following cryopreservation. Biol. Reprod. 2002, 66, 545–554. [Google Scholar] [CrossRef]
- Van Son, M.; Tremoen, N.H.; Gaustad, A.H.; Myromslien, F.D.; Våge, D.I.; Stenseth, E.-B.; Zeremichael, T.T.; Grindflek, E. RNA sequencing reveals candidate genes and polymorphisms related to sperm DNA integrity in testis tissue from boars. BMC Vet. Res. 2007, 13, 362. [Google Scholar] [CrossRef] [Green Version]
- Fraser, L.; Brym, P.; Pareek, C.S.; Mogielnicka-Brzozowska, M.; Paukszto, Ł.; Jastrzębski, J.P.; Wasilewska-Sakowska, K.; Mańkowska, A.; Sobiech, P.; Żukowski, K. Transcriptome analysis of boar spermatozoa with different freezability using RNA-Seq. Theriogenology 2020, 142, 400–413. [Google Scholar] [CrossRef]
- Dai, D.-H.; Qazi, I.H.; Ran, M.-X.; Liang, K.; Zhang, Y.; Zhang, M.; Zhou, G.-B.; Angel, C.; Zeng, C.-J. Exploration of miRNA and mRNA profiles in fresh and frozen-thawed boar sperm by transcriptome and small RNA sequencing. Int. J. Mol. Sci. 2019, 20, 802. [Google Scholar] [CrossRef] [Green Version]
- Ran, M.X.; Zhou, Y.M.; Liang, K.; Wang, W.C.; Zhang, Y.; Zhang, M.; Yang, J.D.; Zhou, G.B.; Wu, K.; Wang, C.D.; et al. Comparative analysis of microRNA and mRNA profiles of sperm with different freeze tolerance capacities in boar (Sus scrofa) and giant panda (Ailuropoda melanoleuca). Biomolecules 2019, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Ropka-Molik, K.; Bereta, A.; Żukowski, K.; Tyra, M.; Piórkowska, K.; Żak, G.; Oczkowicz, M. Screening for candidate genes related with histological microstructure, meat quality and carcass characteristics in pig based on RNA-seq data. Asian Aust. J. Anim. Sci. 2018, 31, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Wimmers, K.; Lin, C.L.; Tholen, E.; Jennen, G.J.; Schellander, K.; Ponsuksili, S. Polymorphisms in candidate genes as markers for sperm quality and boar fertility. Anim. Genet. 2005, 36, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Diniz, D.B.; Lopes, M.S.; Broekhuijse, M.L.W.J.; Lopes, P.S.; Harlizius, B.; Guimaraes, S.E.F.; Duijvesteijn, N.; Knol, E.F.; Silva, F.F. A genome-wide association study reveals a novel candidate gene for sperm motility in pigs. Anim. Reprod. Sci. 2014, 151, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Mucha, A.; Piórkowska, K.; Ropka-Molik, K.; Szyndler-Nędza, M. New polymorphic changes in the WNT7A gene and their effect on reproductive traits in pigs. Ann. Anim. Sci. 2018, 18, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Marques, D.B.D.; Bastiaansen, J.W.M.; Broekhuijse, M.L.W.J.; Lopes, M.S.; Knol, E.F.; Harlizius, B.; Guimarães, S.E.F.; Silva, F.F.; Lopes, P.S. Weighted single-step GWAS and gene network analysis reveal new candidate genes for semen traits in pigs. Genet. Sel. Evol. 2018, 50, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Ren, J.; Ren, D.; Guo, Y.; Wu, Y.; Yang, G.; Mao, H.; Brenig, B.; Huang, L. A whole genome scanning for quantitative trait loci on traits related to sperm quality and ejaculation in pigs. Anim. Reprod. Sci. 2008, 114, 210. [Google Scholar] [CrossRef]
- Matoulkova, E.; Michalova, E.; Borivoj Vojtesek, B.; Hrstka, R. The role of the 3′ untranslated region in post-transcriptional regulation of protein expression in mammalian cells. RNA Biol. 2012, 9, 563–576. [Google Scholar] [CrossRef] [Green Version]
- Mayr, C. Evolution and biological roles of alternative 3′UTRs. Trends Cell Biol. 2016, 26, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sun, Y.; Yang, C.; Zhang, Y.; Jiang, Q.; Huang, J.; Ju, Z.; Wang, X.; Zhong, J.; Wang, C. Functional SNPs of INCENP affect semen quality by alternative splicing mode and binding affinity with the target bta-mir-378 in Chinese Holstein bulls. PLoS ONE 2016, 11, e0162730. [Google Scholar] [CrossRef]
- Liu, L.; Yu, S.; Chen, R.; Lv, X.; Pan, C. A novel synonymous SNP (A47A) of the TMEM95 gene is significant;y associated with the reproductive tracts related to testis in male piglets. Arch. Anim. Breed. 2017, 60, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Wu, W.; Xu, B.; Niu, X.; Cui, H.; Zhang, Y.; Wang, Z.; Wang, X. Variants in the SRD5A2 gene are associated with quality of semen. Mol. Med. Rep. 2012, 6, 639–644. [Google Scholar] [CrossRef] [PubMed]
- KBiosciences. PrimerPicker Lite for KASPar v.0.26; KBiosciences Ltd.: Hoddesdon, UK, 2009. [Google Scholar]
- Pareek, C.S.; Smoczyński, R.; Kadarmideen, H.N.; Dziuba, P.; Blaszczyk, P.; Sikora, M.; Walendzik, P.; Grzybowski, T.; Pierzchała, M.; Horbańczuk, J.; et al. Single nucleotide polymorphism discovery in bovine pituitary gland using RNA-Seq technology. PLoS ONE 2016, 11, e0161370. [Google Scholar] [CrossRef] [PubMed]
- Fardo, D.W.; Becker, K.D.; Bertram, L.; Tanzi, R.E.; Lange, C. Recovering unused information in gemone-wide association studies: The benefit of analyzing SNPs out of Hardy-Weinberg equilibrium. Europ. J. Hum. Genet. 2009, 17, 1676–1682. [Google Scholar] [CrossRef] [PubMed]
- Minelli, C.; Hompson, J.R.; Abrams, K.R.; Thakkinstian, A.; Attia, J. How should we use information about HWE in the meta-analyses of genetic association studeis? Int. J. Epidemiol. 2007, 37, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.L.; Ponsuksili, S.; Tholen, E.; Jennen, D.G.J.; Schellander, K.; Wimmers, K. Candidate gene markers for sperm quality and fertility of boars. Anim. Reprod. Sci. 2006, 349–363. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, H. Laboratory semen assessment and predictions of fertility: Still Utopia. Reprod. Domest. Anim. 2003, 38, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Gadea, J. Sperm factors related to in vitro and in vivo porcine fertility. Theriogenology 2005, 63, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Daigneault, B.W.; McNamara, K.A.; Purdy, P.H.; Krisher, R.L.; Knox, R.V.; Rodriguez-Zas, S.; Miller, D.J. Enhanced fertility prediction of cryopreserved boar spermatozoa using novel sperm function assessment. Andrology 2015, 3, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Gadea, J.; Sellés, E.; Marco, M. The predictive value of porcine seminal parameters on fertility outcome under commercial conditions. Reprod. Domest. Anim. 2004, 239, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Zhao, Z.; Zhao, R.; Xiao, S.; Jiang, H.; Yue, X.; Li, X.; Gao, Y.; Liu, J.; Zhang, J. Effects of novel single nucleotide polymorphisms of the FSH beta-subunit gene on semen quality and fertility in bulls. Anim. Reprod. Sci. 2009, 114, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ju, Z.; Wang, L.; Zhang, Y.; Huang, J.; Li, Q.; Zhong, J.; An, L.; Wang, C. Six novel single-nucleotide polymorphisms in SPAG11 gene and their association with sperm quality traits in Chinese Holstein bull. Anim. Reprod. Sci. 2011, 129, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.; Dziekońska, A.; Strzeżek, R.; Strzeżek, J. Dialysis of boar semen prior to freezing thawing: Its effects on post-thaw sperm characteristics. Theriogenology 2007, 67, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Kasimanickam, V.R.; Kasimanickam, R.K.; Kastelic, J.P.; Stevenson, J.S. Associations of adiponectin and fertility estimates in Holstein bulls. Theriogenology 2013, 79, 766–777. [Google Scholar] [CrossRef]
- Thomas, S.; Kratzsch, D.; Schaab, M.; Scholz, M.; Grunewald, S.; Thiery, J. Seminal plasma adipokine levels are correlated with functional characteristics of spermatozoa. Fertil. Steril. 2013, 99, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Kadivar, A.; Heidari Khoei, H.; Hassanpour, H.; Golestanfar, A.; Ghanaei, H. Correlation of adiponectin mRNA abundance and its receptors with quantitative parameters of sperm motility in rams. Int. J. Fertil. Steril. 2016, 10, 127–135. [Google Scholar]
- Yung-Liang, L.; Sung-Sen, Y.; Shyi-Jou, C.; Yu-Chun, L.; Chin-Chen, C.; Hsin-Hui, H.; Chang, F.W.; Yu, M.H.; Lin, S.H.; Wu, G.J.; et al. OSR1 and SPAK cooperatively modulate Sertoli cell support of mouse spermatogenesis. Sci. Rep. 2016, 6, 37205. [Google Scholar]
- Fraser, L.; Strzeżek, J. Effect of different procedures of ejaculate collection, extenders and packages on DNA integrity of boar spermatozoa following freezing-thawing. Anim. Reprod. Sci. 2007, 99, 317–329. [Google Scholar] [CrossRef]
- Li, M.W.M.; Mruk, D.D.; Cheng, C.Y. Mitogen-activated protein kinases in male reproductive function. Trends Mol. Med. 2009, 15, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, T.; Toshimori, K.; Lida, H. Subcellular localization of MS4A13 isoform 2 in mouse spermatozoa. Reproduction 2017, 154, 843–857. [Google Scholar] [CrossRef]
- Schuster, A.; Tang, C.; Xie, Y.; Ortogero, N.; Yua, S.; Yan, W. SpermBase—A database for sperm-borne RNA contents. Biol. Reprod. 2016, 95, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neild, D.M.; Brouwers, J.F.H.M.; Colenbrander, B.; Agüero, A.; Gadella, B.M. Lipid peroxide formation in relation to membrane stability of fresh and frozen thawed stallion spermatozoa. Mol. Reprod. Dev. 2005, 72, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Garde, J.; Roldan, E.R.S. rab3-Peptide stimulates exocytosis of the ram sperm acrosome via interaction with cyclic AMP and phospholipase A2 metabolites. FEBS Lett. 1996, 391, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Sutovsky, P.; Moreno, R.; Ramalho-Santos, J.; Dominko, T.; Thompson, W.E.; Schatten, G.A. Putative, ubiquitin-dependent mechanism for the recognition and elimination of defective spermatozoa in the mammalian epididymis. J. Cell Sci. 2001, 114, 1665–1675. [Google Scholar] [PubMed]
- Suryawanshi, A.R.; Khan, S.A.; Gajbhiye, R.K.; Gurav, M.Y.; Khole, V.V. Differential proteomics leads to identification of domain-specific epididymal sperm proteins. J. Androl. 2011, 32, 240–259. [Google Scholar] [CrossRef]
- Torabi, F.; Bogle, O.A.; Estanyol, J.M.; Oliva, R.; Miller, D. Zona pellucida-binding protein 2 (ZPBP2) and several proteins containing BX7B motifs in human sperm may have hyaluronic acid binding or recognition properties. Mol. Human Reprod. 2017, 23, 803–816. [Google Scholar] [CrossRef]
- Morris, R.L.; Hoffman, M.P.; Obar, R.A.; McCafferty, S.S.; Gibbons, I.R.; Leone, A.D.; Cool, J.; Allgood, E.L.; Musante, A.M.; Judkins, K.M.; et al. Analysis of cytoskeletal and motility proteins in the sea urchin genome assembly. Dev. Biol. 2006, 300, 219–237. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Cheng, C.Y. Planar cell polarity (PCP) proteins and spermatogenesis. Semin. Cell Dev. Biol. 2016, 59, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Wasilewska, K.; Fraser, L. Boar variability in sperm cryo-tolerance after cooling of semen in different long term extenders at various temperatures. Anim. Reprod. Sci. 2017, 185, 161–173. [Google Scholar] [CrossRef]
- McReynolds, S.; Dzieciatkowska, M.; Stevens, J.; Hansen, K.C.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Toward the identification of a subset of unexplained infertility: A sperm proteomic approach. Fertil. Steril. 2014, 102, 692–699. [Google Scholar] [CrossRef]
- Yuan, S.; Liu, Y.; Peng, H.; Tang, C.; Hennig, G.W.; Wang, Z.; Wang, L.; Yu, T.; Klukovich, R.; Zhang, Y.; et al. Motile cilia of the male reproductive system require miR-34/miR-449 for development and function to generate luminal turbulence. Proc. Natl. Acad. Sci. USA 2019, 116, 3584–3593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Gao, N.; Li, X.; El-Ashram, S.; Wang, Z.; Zhu, L.; Jiang, W.; Peng, X.; Zhang, C.; Chen, Y.; et al. Identifying candidate genes associated with sperm morphology abnormalities using weighted single-step GWAS in a Duroc boar population. Theriogenology 2020, 141, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Beck, D.; Sadler-Riggleman, I.; Skinner, M.K. Generational comparisons (F1 versus F3) of vinclozolin induced epigenetic transgenerational inheritance of sperm differential DNA methylation regions (epimutations) using MeDIP-Seq. Environ. Epigenetics 2017, 3, dvx016. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.; Parda, A.; Filipowicz, K.; Strzeżek, J. Comparison of post-thaw DNA integrity of boar spermatozoa assessed with the neutral Comet assay and sperm-Sus Halomax test kit. Reprod. Domest. Anim. 2010, 45, e155–e160. [Google Scholar] [CrossRef] [PubMed]
- Dziekońska, A.; Fraser, L.; Strzeżek, J. Effect of different storage temperatures on the metabolic activity of spermatozoa following liquid storage of boar semen. J. Anim. Feed Sci. 2009, 18, 638–649. [Google Scholar] [CrossRef]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Pan, Y.; Shen, S.; Lin, L.; Xing, Y. rMATS-DVR: rMATS discovery of differential variants in RNA. Bioinformatics 2017, 33, 2216–2217. [Google Scholar] [CrossRef]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNP in the genome Drosophilla melanogaster w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl variant effect predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F. geneopop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Vitku, J.; Kolatorova, L.; Ricco, C.; Ferroud, C.; Hennebert, O.; Skodova, T.; Heracek, J.; Starka, L. The quantitation of 7 beta-hydroxy-epiandrosterone in the plasma and seminal plasma of men with different degrees of fertility. Physiol. Res. 2018, 67 (Suppl. 3), S511–S519. [Google Scholar] [CrossRef] [PubMed]
- Hilz, S.; Fogarty, E.A.; Modzelewska, A.J.; Cohen, P.E.; Grimson, A. Transcriptome profiling of the developing male germ line identifies the miR-29 family as a global regulator during meiosis. RNA Biol. 2017, 14, 219–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.W.; Herr, D.R.; Noguchi, K.; Yung, Y.C.; Lee, C.W.; Mutoh, T.; Lin, M.E.; Teo, S.T.; Park, K.E.; Mosley, A.N.; et al. LPA receptors: Subtypes and biological actions. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 157–186. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, C. Redox regulation of mammalian sperm capacitation. Asian J. Androl. 2015, 17, 583–590. [Google Scholar] [CrossRef]
- Jamsai, D.; Bianco, D.M.; Smith, S.J.; Merriner, D.J.; Ly-Huynh, J.D.; Herlihy, A.; Niranjan, B.; Gibbs, G.M.; O’Bryan, M.K. Characterization of gametogenetin 1 (GGN1) and its potential role in male fertility through the interaction with the ion channel regulator, cysteine-rich secretory protein 2 (CRISP2) in the sperm tail. Reproduction 2008, 135, 751–759. [Google Scholar] [CrossRef] [Green Version]
- Naz, R.K.; Sellamuthu, R. Receptors in spermatozoa: Are they real? J. Androl. 2006, 27, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Song, P. Molecular cloning of a novel gene ZAhi-1 and its expression analysis during zebrafish gametogenesis. Mol. Biol. Rep. 2006, 33, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Parte, P. Membrane bound glucose regulated protein 78 interacts with alpha-2-macroglobulin to promote actin reorganization in sperm during epididymal maturation. Mol. Hum. Reprod. 2019, 25, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.V.; Korrodi-Gregorio, L.; Luers, G.; Cardosa, M.J.; Patricio, A.; Maia, N.; da E Silva Cruz, E.F.; Fardilha, M. Characterisation of several ankyrin repeat protein variant 2, a phosphoprotein phosphatase 1-interacting protein, in testis and spermatozoa. Reprod. Fertil. Dev. 2016, 7, 1009–1019. [Google Scholar] [CrossRef]
- Owa, M.; Uchihashi, T.; Yanagisaw, H.A.; Yamano, T.; Iguchi, H.; Fukuzawa, H.; Wakabayashi, K.I.; Ando, T.; Kikkawa, M. Inner lumen proteins stabilize doublet microtubules in cilia and flagella. Nat. Commun. 2019, 1143, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Shen, J.; Kugan, N.; Furth, E.E.; Lombard, D.B.; Cheung, C.; Pak, S.; Luo, G.; Pignolo, R.J.; DePinho, R.A.; et al. Telomere shortening exposes functions for the mouse Werner and Bloom Syndrome genes. Mol. Cell. Biol. 2004, 24, 8437–8446. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Cheng, L.; Gu, Y.; Xin, Y.; Wu, B.; Zhou, S.; Guo, S.; Liu, Y.; Diao, H.; Shi, H.; et al. A human lectin microarray for sperm surface glycosylation analysis. Mol. Cell. Proteom. 2016, 15, 2839–2851. [Google Scholar] [CrossRef] [Green Version]
- Kropp, J.; Carrillo, J.A.; Namous, H.; Daniels, A.; Salih, S.M.; Song, J.; Khatib, H. Male fertility status is associated with DNA methylation signatures in sperm and transcriptomic profiles of bovine preimplantation embryos. BMC Genom. 2017, 18, 280. [Google Scholar] [CrossRef] [Green Version]
- Augustine, L.M.; Markelewicz, R.J.; Boekelheide, K.; Cherrington, N.J. Xenobiotic and endobiotic transporter mRNA expression in the blood–testis barrier. Drug Metab. Dispos. 2005, 33, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E. The function of spermine. IUBMB Life 2014, 66, 8–18. [Google Scholar] [CrossRef]
- Mikolcevic, P.; Sigl, R.; Rauch, V.; Hess, M.W.; Pfaller, K.; Barisic, M.; Pelliniemi, L.J.; Boesl, M.; Geley, S. Cyclin-dependent kinase 16/PCTAIRE kinase 1 is activated by cyclin Y and is essential for spermatogenesis. Mol. Cell. Biol. 2012, 32, 868–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.E.; Saraceno, G.E.; Capani, F.; Castilla, R. Long-chain acyl-CoA synthetase 4 is regulated by phosphorylation. Biochem. Biophys. Res. Commun. 2013, 430, 272–277. [Google Scholar]
- Moscatelli, N.; Lunetti, P.; Braccia, C.; Armirotti, A.; Pisanello, F.; De Vittorio, M.; Zara, V.; Ferramosca, A. Comparative proteomic analysis of proteins involved in bioenergetics pathways associated with human sperm motility. Int. J. Mol. Sci. 2019, 20, 3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinaci, M.; Volpe, S.; Bernardini, C.; De Ambrogi, M.; Tamanini, C.; Seren, E.; Galeati, G. Immunolocalization of heat shock protein 70 (Hsp 70) in boar spermatozoa and its role during fertilization. Mol. Reprod. Dev. 2005, 72, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Braas, K.M.; Harakall, S.A.; Ouafik, L.; Eipper, B.A.; May, V. Expression of peptidylglycine alpha-amidating monooxygenase: An in situ hybridization and immunocytochemical study. Endocrinology 1992, 130, 2778–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Q.; Feng, T.; Coleman, M.L. Modifying the maker: Oxygenases target ribosome biology. Translation 2015, 3, e1009331. [Google Scholar] [CrossRef] [Green Version]
- Bragado, M.J.; Gil, M.C.; Martin-Hidalgo, D.; Hurtado de Llera, A.; Bravo, N.; Moreno, A.D.; Garcia-Marin, L.L. Src family tyrosine kinase regulates acrosome reaction but not motility in porcine spermatozoa. Reproduction 2012, 144, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Modarressi, M.H.; Behnam, B.; Cheng, M.; Taylor, K.E.; Wolfe, J.; van der Hoorn, F.A. Tsga10 Encodes a 65-kilodalton protein that is processed to the 27-kilodalton fibrous sheath protein. Biol. Reprod. 2004, 70, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Vicens, A.; Luke, L.; Roldan, E.R. Proteins involved in motility and sperm-egg interaction evolve more rapidly in mouse spermatozoa. PLoS ONE 2014, 9, e91302. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, R.E.; Duncan, W.C. The SLIT–ROBO pathway: A regulator of cell function with implications for the reproductive system. Reproduction 2010, 139, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Demain, L.A.; Conway, G.S.; Newman, W.G. Genetics of mitochondrial dysfunction and infertility. Clin. Genet. 2017, 91, 199–207. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters | Fresh, Pre-Freeze Semen | Post-Thaw Semen | Total | |
|---|---|---|---|---|
| GSF | PSF | GSF + PSF | ||
| Motility (%) | 75.78 ± 0.28 | 42.96 ± 1.63 a | 26.85 ± 1.28 b | 35.31 ± 1.65 |
| MMP (%) | 87.83 ± 2.64 | 48.82 ± 1.04 a | 31.55 ± 1.55 b | 40.62 ± 1.64 |
| PMI (%) | 87.43 ± 1.22 | 49.91 ± 0.84 a | 34.55 ± 1.62 b | 42.64 ± 1.51 |
| NAR acrosome integrity (%) | 91.62 ± 1.07 | 53.93 ± 0.60 a | 40.85 ± 0.86 b | 47.72 ± 1.38 |
| DNA fragmentation (%) | 2.97 ± 0.13 | 7.85 ± 0.45 a | 10.56 ± 0.83 b | 9.16 ± 0.50 |
| LPO (nMol MDA/109spz/h) | 21.34 ± 0.68 | 27.37 ± 0.47 a | 31.55 ± 0.48 b | 29.36 ± 0.45 |
| Sample ID | Raw SNV | ≥10% Reads ≥90% | % Raw Reads | ≥10 Reads = 100% | Raw dbSNPs | ≥10% Reads ≥90% | % Raw Reads | ≥10 Reads = 100% |
|---|---|---|---|---|---|---|---|---|
| G01 | 941,355 | 80,714 | 8.6% | 66,625 | 632,893 | 68,577 | 10.8% | 57,182 |
| G09 | 873,327 | 78,084 | 8.9% | 73,323 | 583,610 | 66,131 | 11.3% | 62,796 |
| G17 | 941,859 | 144,415 | 15.3% | 135,223 | 618,586 | 122,917 | 19.9% | 116,302 |
| P30 | 866,451 | 100,806 | 11.6% | 94,399 | 575,549 | 84,148 | 14.6% | 79,695 |
| P38 | 890,984 | 110,574 | 12.4% | 103,347 | 591,527 | 94,160 | 15.9% | 88,990 |
| P39 | 898,293 | 110,753 | 12.3% | 103,664 | 594,402 | 94,107 | 15.8% | 89,169 |
| SNP Effect | SNP Count | Percent (%) |
|---|---|---|
| 3 prime UTR variant | 919 | 67.031 |
| 5 prime UTR variant | 120 | 8.752 |
| Synonymous variant | 100 | 7.294 |
| Missense variant | 74 | 5.398 |
| Non coding transcript exon variant | 57 | 4.158 |
| Splice region variant | 4 | 0.292 |
| Intron variant, non-coding transcript variant | 3 | 0.219 |
| Stop gained | 2 | 0.145 |
| Others | 4 | 0.292 |
| Unknown | 88 | 6.419 |
| Table 4A | |||||||||||
| SNP ID | Locus | Freezability Groups | Allele Counts | Total | Allele Frequencies | p-value | HWE | ||||
| A | G | A | G | p-value | χ2 | Probability | |||||
| APPL1 | GSF | 19 | 23 | 42 | 0.452 | 0.548 | 1.00000 | 1.0000 | 5.8659 | 0.2094 | |
| unknown | PSF | 15 | 19 | 34 | 0.441 | 0.559 | 0.0532 | ||||
| Total | 34 | 42 | 76 | 0.447 | 0.553 | ||||||
| CYP7B1 | GSF | 5 | 35 | 40 | 0.125 | 0.875 | 0.71180 | 0.0105 | 9.1044 | 0.0585 | |
| rs338842672 | PSF | 3 | 35 | 38 | 0.079 | 0.921 | 1.0000 | ||||
| Total | 8 | 70 | 78 | 0.103 | 0.897 | ||||||
| EML5 | GSF | 12 | 30 | 42 | 0.286 | 0.714 | 0.06221 | 0.1283 | 6.1640 | 0.1872 | |
| rs345056502 | PSF | 17 | 17 | 34 | 0.500 | 0.500 | 0.3575 | ||||
| Total | 29 | 47 | 76 | 0.382 | 0.618 | ||||||
| LPAR1 | GSF | 26 | 14 | 40 | 0.650 | 0.350 | 0.62845 | 0.6347 | 2.0228 | 0.7316 | |
| unknown | PSF | 24 | 10 | 34 | 0.706 | 0.294 | 0.5731 | ||||
| Total | 50 | 24 | 74 | 0.676 | 0.324 | ||||||
| RAB3C | GSF | 11 | 31 | 42 | 0.262 | 0.738 | 0.10038 | 1.0000 | 7.5732 | 0.1085 | |
| rs81210636 | PSF | 17 | 21 | 38 | 0.447 | 0.553 | 0.0227 | ||||
| Total | 38 | 52 | 80 | 0.350 | 0.650 | ||||||
| SARS | GSF | 19 | 21 | 40 | 0.475 | 0.525 | 0.50110 | 1.0000 | 8.8484 | 0.0650 | |
| rs344846507 | PSF | 15 | 23 | 38 | 0.395 | 0.605 | 0.0120 | ||||
| Total | 34 | 44 | 78 | 0.436 | 0.564 | ||||||
| TXNIP | GSF | 12 | 30 | 42 | 0.286 | 0.714 | 0.10631 | 0.1301 | 14.0970 | 0.0070 | |
| rs340075321 | PSF | 18 | 20 | 38 | 0.474 | 0.526 | 0.0067 | ||||
| Total | 30 | 50 | 80 | 0.375 | 0.625 | ||||||
| Table 4B | |||||||||||
| SNP ID | Locus | Freezability Groups | Allele Counts | Total | Allele Frequencies | p-value | HWE | ||||
| G | A | G | A | p-value | χ2 | Probability | |||||
| OXSR1 | GSF | 18 | 24 | 42 | 0.429 | 0.571 | 0.48138 | 0.3775 | 8.4171 | 0.0774 | |
| rs339379734 | PSF | 17 | 15 | 32 | 0.531 | 0.469 | 0.0394 | ||||
| Total | 35 | 39 | 74 | 0.473 | 0.527 | ||||||
| A2M | GSF | 8 | 34 | 42 | 0.190 | 0.810 | 0.36658 | 0.0001 | 26.9432 | 0.0000 | |
| rs339026428 | PSF | 2 | 36 | 38 | 0.053 | 0.947 | 0.0271 | ||||
| Total | 10 | 70 | 80 | 0.125 | 0.875 | ||||||
| ANKRD42 | GSF | 16 | 26 | 42 | 0.381 | 0.619 | 0.49879 | 0.6413 | 18.6650 | 0.0009 | |
| rs81210697 | PSF | 20 | 18 | 38 | 0.526 | 0.474 | 0.0015 | ||||
| Total | 36 | 44 | 80 | 0.450 | 0.550 | ||||||
| CCDC149 | GSF | 17 | 25 | 42 | 0.405 | 0.595 | 0.48804 | 0.3681 | 7.5901 | 0.1078 | |
| rs332902509 | PSF | 17 | 17 | 34 | 0.500 | 0.500 | 0.0611 | ||||
| Total | 34 | 42 | 76 | 0.447 | 0.553 | ||||||
| CFAP52 | GSF | 28 | 12 | 40 | 0.700 | 0.300 | 0.79820 | 0.6178 | 3.5245 | 0.4742 | |
| unknown | PSF | 25 | 9 | 34 | 0.735 | 0.265 | 0.2779 | ||||
| Total | 53 | 21 | 74 | 0.716 | 0.284 | ||||||
| COMMD2 | GSF | 19 | 21 | 40 | 0.475 | 0.525 | 0.64518 | 0.0050 | 19.7578 | 0.0006 | |
| rs318435440 | PSF | 14 | 20 | 34 | 0.412 | 0.588 | 0.0102 | ||||
| Total | 33 | 41 | 74 | 0.446 | 0.554 | ||||||
| FBXO16 | GSF | 16 | 24 | 40 | 0.400 | 0.600 | 0.16668 | 1.0000 | 0.9449 | 0.9180 | |
| rs341614458 | PSF | 21 | 15 | 36 | 0.583 | 0.417 | 0.6235 | ||||
| Total | 37 | 39 | 76 | 0.487 | 0.513 | ||||||
| MAP3K20 | GSF | 11 | 27 | 38 | 0.289 | 0.711 | 1.00000 | 0.2531 | 5.3360 | 0.2545 | |
| rs340643892 | PSF | 9 | 25 | 34 | 0.265 | 0.735 | 0.2741 | ||||
| Total | 20 | 52 | 72 | 0.278 | 0.722 | ||||||
| WRN | GSF | 9 | 29 | 38 | 0.237 | 0.763 | 0.76470 | 1.0000 | 0.0000 | 1.0000 | |
| rs319208708 | PSF | 5 | 21 | 26 | 0.192 | 0.808 | 1.0000 | ||||
| Total | 14 | 50 | 64 | 0.212 | 0.788 | ||||||
| Table 4C | |||||||||||
| SNP ID | Locus | Freezability Groups | Allele Counts | Total | Allele Frequencies | p-value | HWE | ||||
| A | C | A | C | p-value | χ2 | Probability | |||||
| CLEC7A | GSF | 15 | 27 | 42 | 0.357 | 0.643 | 1.00000 | 0.6647 | 3.2122 | 0.5230 | |
| rs325939188 | PSF | 13 | 21 | 34 | 0.382 | 0.618 | 0.3019 | ||||
| Total | 28 | 48 | 76 | 0.368 | 0.632 | ||||||
| EML6 | GSF | 10 | 30 | 40 | 0.250 | 0.750 | 0.60588 | 1.0000 | 4.4370 | 0.3501 | |
| rs322659685 | PSF | 11 | 23 | 34 | 0.324 | 0.676 | 0.3501 | ||||
| Total | 21 | 53 | 73 | 0.284 | 0.716 | ||||||
| C | A | C | A | ||||||||
| ABCB11 | GSF | 16 | 24 | 40 | 0.400 | 0.600 | 0.62897 | 0.0005 | 15.2834 | 0.0041 | |
| rs324930519 | PSF | 11 | 23 | 34 | 0.324 | 0.676 | 1.0000 | ||||
| Total | 27 | 47 | 74 | 0.635 | 0.365 | ||||||
| SMS | GSF | 12 | 26 | 38 | 0.316 | 0.684 | 0.33352 | 0.0015 | 13.8162 | 0.0079 | |
| rs343122214 | PSF | 16 | 20 | 36 | 0.444 | 0.556 | 0.6577 | ||||
| Total | 28 | 46 | 74 | 0.378 | 0.622 | ||||||
| Table 4D | |||||||||||
| SNP ID | Locus | Freezability Group | Allele Counts | Allele Frequencies | Total | p-value | HWE | ||||
| C | G | C | G | p-value | χ2 | Probability | |||||
| TMEM177 | GSF | 21 | 17 | 38 | 0.553 | 0.447 | 0.81666 | 0.0001 | 23.0661 | 0.0001 | |
| rs80954196 | PSF | 18 | 18 | 36 | 0.500 | 0.500 | 0.1750 | ||||
| Total | 39 | 35 | 74 | 0.527 | 0.473 | ||||||
| G | C | G | C | ||||||||
| MS4A2 | GSF | 10 | 30 | 40 | 0.250 | 0.750 | 0.32355 | 0.2784 | 9.0353 | 0.0602 | |
| rs339836492 | PSF | 13 | 23 | 36 | 0.361 | 0.639 | 0.0392 | ||||
| Total | 23 | 53 | 76 | 0.303 | 0.697 | ||||||
| Table 4E | |||||||||||
| SNP ID | Locus | Freezability Groups | Allele Counts | Total | Allele Frequencies | p-value | HWE | ||||
| C | T | C | T | p-value | χ2 | Probability | |||||
| PRICKLE1 | GSF | 15 | 27 | 42 | 0.357 | 0.643 | 1.00000 | 1.0000 | 2.2918 | 0.6823 | |
| rs694366781 | PSF | 12 | 20 | 32 | 0.375 | 0.625 | 0.3179 | ||||
| Total | 27 | 47 | 74 | 0.365 | 0.635 | ||||||
| SCLT1 | GSF | 9 | 33 | 42 | 0.214 | 0.786 | 0.77416 | 0.5322 | 1.2614 | 0.8679 | |
| rs337913978 | PSF | 6 | 30 | 36 | 0.167 | 0.833 | 1.0000 | ||||
| Total | 15 | 63 | 78 | 0.192 | 0.808 | ||||||
| T | C | T | C | ||||||||
| ACSL4 | GSF | 5 | 37 | 42 | 0.119 | 0.881 | 0.36037 | 1.0000 | 0.0000 | 1.0000 | |
| rs334625232 | PSF | 8 | 28 | 36 | 0.222 | 0.778 | 1.0000 | ||||
| Total | 13 | 65 | 78 | 0.167 | 0.833 | ||||||
| ATP5F1A | GSF | 22 | 20 | 42 | 0.524 | 0.476 | 0.06697 | 0.0000 | >43.1295 | <0.0000 | |
| rs328079913 | PSF | 28 | 10 | 38 | 0.737 | 0.263 | 0.0036 | ||||
| Total | 50 | 30 | 80 | 0.625 | 0.375 | ||||||
| HSPA13 | GSF | 11 | 19 | 30 | 0.367 | 0.633 | 0.57756 | 0.2960 | 3.7925 | 0.4348 | |
| rs335938037 | PSF | 8 | 22 | 30 | 0.267 | 0.733 | 0.5072 | ||||
| Total | 19 | 41 | 60 | 0.317 | 0.683 | ||||||
| PAM | GSF | 11 | 29 | 40 | 0.275 | 0.725 | 0.77866 | 0.2555 | 2.7292 | 0.6041 | |
| rs81217594 | PSF | 6 | 20 | 26 | 0.231 | 0.769 | 1.0000 | ||||
| Total | 17 | 49 | 66 | 0.258 | 0.742 | ||||||
| RIOX2 | GSF | 9 | 29 | 38 | 0.237 | 0.763 | 0.57428 | 1.0000 | 0.0000 | 1.0000 | |
| unknown | PSF | 8 | 18 | 26 | 0.308 | 0.692 | 1.0000 | ||||
| Total | 17 | 47 | 64 | 0.266 | 0.734 | ||||||
| SKAP2 | GSF | 16 | 26 | 42 | 0.381 | 0.619 | 0.21618 | 0.6422 | 2.1935 | 0.7002 | |
| rs336351767 | PSF | 8 | 26 | 34 | 0.235 | 0.765 | 0.5200 | ||||
| Total | 24 | 52 | 76 | 0.316 | 0.684 | ||||||
| Table 4F | |||||||||||
| SNP ID | Locus | Freezability group | Allele counts | Total | Allele frequencies | p-value | HWE | ||||
| A | T | A | T | p-value | χ2 | Probability | |||||
| ROBO1 | GSF | 5 | 37 | 42 | 0.119 | 0.881 | 0.74597 | 1.0000 | 0.0000 | 1.0000 | |
| rs331568674 | PSF | 5 | 29 | 34 | 0.147 | 0.853 | 1.0000 | ||||
| Total | 10 | 66 | 76 | 0.132 | 0.868 | ||||||
| T | A | T | A | ||||||||
| PLBD1 | GSF | 14 | 28 | 42 | 0.333 | 0.667 | 0.16654 | 0.0487 | 6.0432 | 0.1959 | |
| rs321497623 | PSF | 18 | 18 | 36 | 0.500 | 0.500 | 1.0000 | ||||
| Total | 32 | 46 | 78 | 0.410 | 0.590 | ||||||
| Table 4G | |||||||||||
| SNP ID | Locus | Freezability group | Allele counts | Total | Allele frequencies | p-value | HWE | ||||
| G | T | G | T | p-value | χ2 | Probability | |||||
| HARS2 | GSF | 8 | 32 | 40 | 0.200 | 0.800 | 0.78401 | 0.5482 | 2.5152 | 0.6419 | |
| rs336003721 | PSF | 8 | 26 | 34 | 0.235 | 0.765 | 0.5187 | ||||
| Total | 16 | 58 | 74 | 0.216 | 0.784 | ||||||
| A. rs337913978 (SCLT1) | Motility | Plasma Membrane Integrity (PMI) | ||||
| F | G | F × G | F | G | F × G | |
| p-value | p-value | p-value | p-value | p-value | p-value | |
| <0.001 | >0.305 | <0.043 | <0.001 | >0.063 | <0.028 | |
| B. rs340643892 (MAP3K20) | NAR Acrosome Integrity | Lipid Peroxidation (LPO) | ||||
| F | G | F × G | F | G | F × G | |
| p-value | p-value | p-value | p-value | p-value | p-value | |
| <0.001 | >0.146 | <0.010 | <0.001 | <0.022 | >0.981 | |
| C. rs339836492 (MS4A2) | Lipid Peroxidation (LPO) | |||||
| F | G | F × G | ||||
| p-value | p-value | p-value | ||||
| <0.001 | <0.039 | >0.302 | ||||
| D. rs331568674 (ROBO1) | DNA Fragmentation | |||||
| F | G | F × G | ||||
| p-value | p-value | p-value | ||||
| <0.030 | >0.800 | <0.047 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mańkowska, A.; Brym, P.; Paukszto, Ł.; Jastrzębski, J.P.; Fraser, L. Gene Polymorphisms in Boar Spermatozoa and Their Associations with Post-Thaw Semen Quality. Int. J. Mol. Sci. 2020, 21, 1902. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051902
Mańkowska A, Brym P, Paukszto Ł, Jastrzębski JP, Fraser L. Gene Polymorphisms in Boar Spermatozoa and Their Associations with Post-Thaw Semen Quality. International Journal of Molecular Sciences. 2020; 21(5):1902. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051902
Chicago/Turabian StyleMańkowska, Anna, Paweł Brym, Łukasz Paukszto, Jan P. Jastrzębski, and Leyland Fraser. 2020. "Gene Polymorphisms in Boar Spermatozoa and Their Associations with Post-Thaw Semen Quality" International Journal of Molecular Sciences 21, no. 5: 1902. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051902